The OFFICIAL DATE OF PUBLICATION is handstamped in a limited number of initial copies and is recorded in the institution's annual report, Annals of the Smithsonian Institution. A world classification of the Harmacloninae, a new subfamily of Tineidae (Lepidoptera: Tineidea) / Donald R. Smithsonian contributions to zoology ; no. 597). Diakonoff (1968) retained Harmaclon in the Tineidae, as did Nielsen and Common (in CSIRO, 1991) and Robinson and Nielsen (1993) in their treatment of the Australian fauna.
USNM collections of the former United States National Museum, now deposited in the National Museum of Natural History, Smithsonian Institution, Washington, D.C. In the most recently published overview of the subject to date, Robinson (in Robinson and Nielsen, 1993) acknowledged 11 subfamilies within the questionably monophyletic family Tineidae.
Unique Forewing/Hindwing Secondary Coupling Sys- tem: Bradley (1953a) first described this very effective wing
Adults are nocturnal and phototropic, and the majority of species apparently fly year-round in tropical areas. Nine were assumed to be monophyletic, and the remaining two, Meessiinae and Myrmecozelinae, were not defined by any known synapomorphies. Myrmecozelinae in particular were thought to be polyphyletic; however, they may contain a larger monophyletic unit characterized by Myrmecozela (type species: ochraceella TengstrSm) on the basis of certain shared male genitalia.
Harmaclona was recently grouped with Myrmecozelinae (Robinson and Nielsen, 1993), more or less by default, because several characters excluded it from other, more well-defined subfamilies. Partly because I agree with Robinson about the polyphyly of Myrmecozelinae, I believe it is prudent and more informative to recognize Harmaclona together with its revived sister genus Micrerethista as a separate subfamily, defined by the following synapomorphies .
Loss of Pretarsal Arolium and Pseudempodial Seta: The associated loss of both the arolium and pseudempo-
Robinson (in Robinson and Nielsen, 1993) observed females ovipositing between cork and glass in a collection vial, with the eggs covered by the female's nuchal hairs.
Abdominal Tympanic Organs: Auditory organs have evolved in only about 15 of the 119 families of Lepidoptera
The tympanic organs of the Harmacloninae are situated at the base of the abdomen on the anterior half of the second sternum, in a position similar to those of the Pyraloidea. The tympana are elliptical in outline and extend approximately half the length of the second sternite in both sexes. A shallow, external cavity is formed immediately anteroventral to the tympanum by the concavity of the adjacent metacoxae.
Dissection of the abdomen revealed a single air chamber, lined with membrane, beneath the tympanic membrane. A broad, thin band of tissue, perhaps containing the scoloparium, was attached to the inner wall of the tympanic membrane in the region of the spinula (Figure 107).
Enlargement of Sternal Apophyses: The slender apo- physes projecting anteriorly from abdominal sternum II
Harmaclona cossidella: 9, male; 10, female; 11, enlargement of figure 9, showing spherical sensory cavities in flagellomeres 1–5 (see arrow); 12, enlargement of figure 10, showing sensory cavities in flagellomeres 1–9 (see arrow). As a result, attempts to remove the abdomen for genital dissection often result in a separation between the meso- and metathorax. A cursory examination of dried and cleaned specimens revealed no evidence of hard-walled internal cavities or remnants of scolopia attached to the tympanic spinula.
Aedoeagus with Basal Midventral Keel: The ridge-like keel developed at the base of the aedoeagus (Figures 166,184)
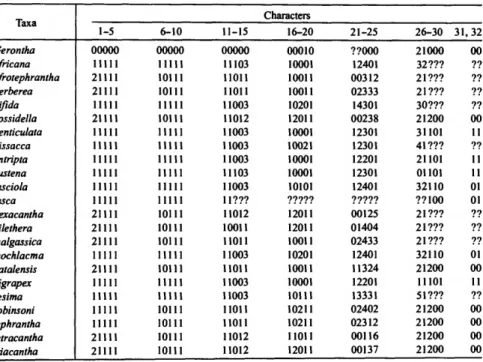
Analysis of the data for 22 taxa (excluding Micrerethista fused) by the implicit summary option (ie*) of Hennig86 initially resulted in 496 trees (length = 106; Harmaclona hilethera is believed to be more derived (Figure 91) than suggested by his more apomorphic anellus—the most specialized form within the genus. The apex of the valva is entire in Gerontha (Moriuti, 1989) and is rarely divided or deeply notched in Myrmecozelinae (Zagulajev, 1975).
The presence of the basal midventral keel on the aedoeagus constitutes an autapomorphy for the Harmacloninae. This characteristic cluster of spines constitutes a synapomorphy of the Micrere-Thista entripta species group (Figs.
KhD-
MHHl-i-i-
HHHH]-
HKHH>
HHHhH-
HJ-HHM-
Mesofurcasternum (Figure 96) with paired lateral and mesal arms of similar size; anterior process curved ventrad, ~1.2 length of lateral arms. Foreleg with epiphysis pectinate, well developed (indicates length of tibia; foretibia short, about equal to first tarsomere; Male genitalia [Figure 163] with gnathos present; apotheca absent; ventral saccular flap reduced, less than 0.75 length of spine patch.
Valve with dorsal lobe moderately thin, width -0.3 its length; apex slightly inverted; ventral lobe -0.5 length of dorsal lobe. Valve with dorsal lobe moderately slender at base, width less than 0.25 length; apex slightly inverted; ventral lobe ~ 0.55 length of dorsal lobe. Aedoeagus with 1-3 pairs of minute spines along lateral margin of phallotrem; basal keel well developed, ~ 0.28 length of aedoeagus; base of aedoeagus cylindrical; ejaculatory duct not coiled.
Valva with dorsal lobe relatively slender at the base, width less than 0.4 of the length; ventral lobe -0.64 the length of the dorsal lobe, with a narrowly rounded apex. Ductus bursae short, about the same length as the anterior apophyses; antrum subquadrate, slightly tapering anteriorly; linear band of spicules absent. Valva variable (figures dorsal lobe moderately broad to slender, greatest width 0.3 to 0.37 the length of the lobe, apex subacute to moderate.
Valva with dorsal lobe very slender, ~0.5 times the length of valva; greatest width of the lobe at the base -2.2 of the length; apex of dorsal lobe rounded, slightly curved upwards; ventral lobe ~0.8 the length of the dorsal lobe. Valva with dorsal lobe moderately slender, ~0.4 the length of valva; greatest width of lobe at base ~0.4 of length; apex of lobe tapered and slightly curved upwards; ventral lobe ~0.75 the length of the dorsal lobe. Ductus bursae is very elongated, ~2-2.5x the length of the anterior apophyses, with a dense band of spicules extending along one side of the duct to the corpus bursae.
0.4 the length of forewing; dorsum of basal l k white, lightly irrorated with fuscous; apical 3 /4
Valve with dorsal lobe elongate and thin, ~ 0.6 valve length; greatest width of lobe at base ~0.25 its length; ventral lobe ~ 0.61 length of dorsal lobe. Aedoeagus with 6-8 elongate subapical spines projecting laterally and curved ventrally (Fig. Basal midventral keel well developed, -0.29 length of aedoeagus; Ventral (saccular) lobe of valve well developed, ranging from 0.85 to 1 . cucullar) lobe.
Female genitalia with caudal edge of convex ostium [Figure 338]; antrum elongated, ~0.3 the length of the entire ductus bursae H. Caudal edge of female ostium concave [Figure 336]; antrum shorter, less than 0.2 times the length of the ductus bursae H. MALE GENITALIA (Figures 271-276). – Anterior margin slightly concave on either side of the relatively short sac, the latter ~0.25 times the length of the valva.
Aedoeagus with ventral keel moderately well developed, ~XU length of entire aedoeagus (Figure 256); apex expanded as in H. MALE GENITALIA (Figures 283-288).—The anterior margin of the vinculum is deeply concave on the other side of the elongate saccus, the latter nearly 0.9 the length of the valve (Figure 283). MALE GENITALIA (Figs. 289-294).—Anterior margin of vinculum slightly concave on other side of elongate saccus, the latter about 0.7 length of valva (Fig. 289).
Anellus with a pair of short, conical, caudal lobes less than 0.15 times as long as the entire anellus. Aedoeagus with elongated basal keel, almost 0.4 times the length of the entire aedoeagus (Figure 292); distal end with a pair of ventrally curved and laterally diverging apical spines and a subapical pair of ventrolateral spines approximately the same length as the apical pair (Figs. 293,294). Aedoeagus with elongated basal keel, ~0.42 the length of the entire aedoeagus (Figure 298); apex with a divergent pair of acute or minutely bidentate apical arms and a single, shorter mid-dorsal spine (Figs. 299,300); even smaller dorsal spines are sometimes present near or on the base of the terminal arms (Figs. 301, 302).
Literature Cited
Martin, editor, Contributions to West Coast Paleontology and Geology in Honor of Standish Mallory. Manuscripts intended for serial publication receive substantive review at their original Smithsonian museums or offices and are submitted to the Smithsonian Institution Press on Form SI-36, which must show approval by the appropriate authority designated by the sponsoring organizational unit. Requests for special treatment—use of color, folds, box-bound covers, etc.—require, on the same form, the added approval of the sponsoring authority.
Review of manuscripts and art by the press for series format and style requirements, completeness and clarity of copy, and arrangement of all material, as described below, will govern, at the discretion of the press, acceptance or rejection of manuscripts and art . The first page of the text should have the title and author at the top of the page; The second page should have only the author's name and professional mailing address, to be used as an unnumbered footnote on the first page of the printed text. If cross-referencing between key and text is required, do not include page references within the key, but number the detached taxa, using the same numbers as their corresponding heads in the text.
Synonymy in zoology should use the short form (taxon, author, year page), with full reference at the end of the paper under Literature Cited." For botany, the long form (taxon, author, abbreviated journal or book title, volume, page, year, without reference in cited literature") is optional. Text referencing system (author, year used in the text, with full citation in "Literature Cited" at the end of the text) should be used instead of bibliographical footnotes in all contribution series and is strongly recommended in the Study Series : (Jones or ". Extensive notes should be collected and placed at the end of the text in a note section.
For titles of books and articles, use capital letters in sentence form, according to the rules of the language used (exception: capitalize all important words in English). Use the bracket/comma system for volume(number): Pagination For alignment and arrangement of elements, follow the format of recent publications in the series for which the manuscript is intended. Legends for illustrations should be submitted at the end of the manuscript, with as many legends typed, double-spaced, on a page as is convenient.
They should be called figures and should be numbered consecutively as they will appear in the monograph. Use of the metric system of measurement is preferable; where use of the English system is unavoidable, provide metric equivalents in brackets.

