I would like to thank the current and former members of the Tirrell lab for fostering a fantastic lab culture that I have thoroughly enjoyed over the years. I would like to thank the members of the MCME subgroup for their valuable suggestions on my work, Dr.
THE BACILLUS SUBTILIS SPORE AND ITS APPLICATION IN BIOTECHNOLOGY
Introduction
Genetic Regulation of Spore Formation
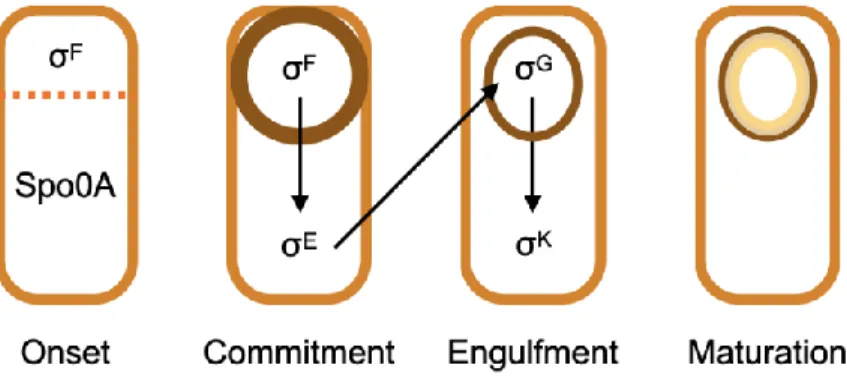
Following asymmetric septum formation, distinct gene expression programs occur in the pro-spore and mother cell compartments to drive further morphological development (Figure 1.2). Another set of compartment-specific σ-factors – σG in the pro-spore compartment and σK in the mother cell compartment – become active as early-phase σ-factors are removed by proteases.
Structure and Properties of the Spore
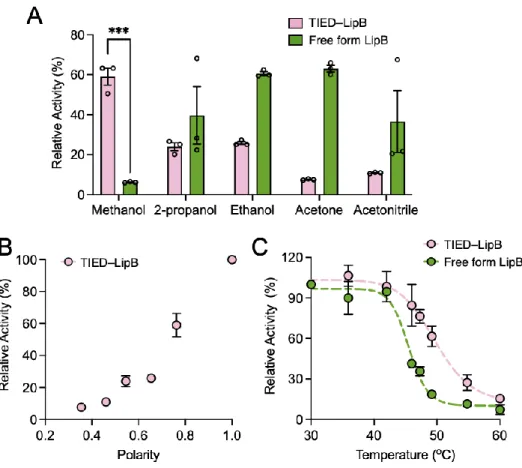
The partially dehydrated nature of the core, coupled with the small acid-soluble proteins that bind to the genome, makes the spores highly resistant to heat and desiccation. CotA, a copper-dependent laccase, produces a dark pigment, which effectively protects the spores from UV radiation.53 In addition, the pigment resembles melanin, which is known to confer resistance to peroxide in many microbes.54 A recent study found that major cystine-rich crust proteins, CotY and CotZ, in purified form, can each self-assemble into highly ordered secondary structures, implying their importance for the structural integrity of the spore.55 In addition, the formation of a crust polysaccharide layer is believed to enhance the protective the spores against biological degradation and clearance.52 1.4 Germination.
Engineered Spores as Platforms for Heterologous Protein Display
Relying on this approach, many studies have shown success in the display of bioactive molecules on B. subtilis spores. Chen and colleagues presented the first example of using B.subtilis to generate water-sensitive materials.104 The spore cortex surrounding the dehydrated core swells upon absorbing water.
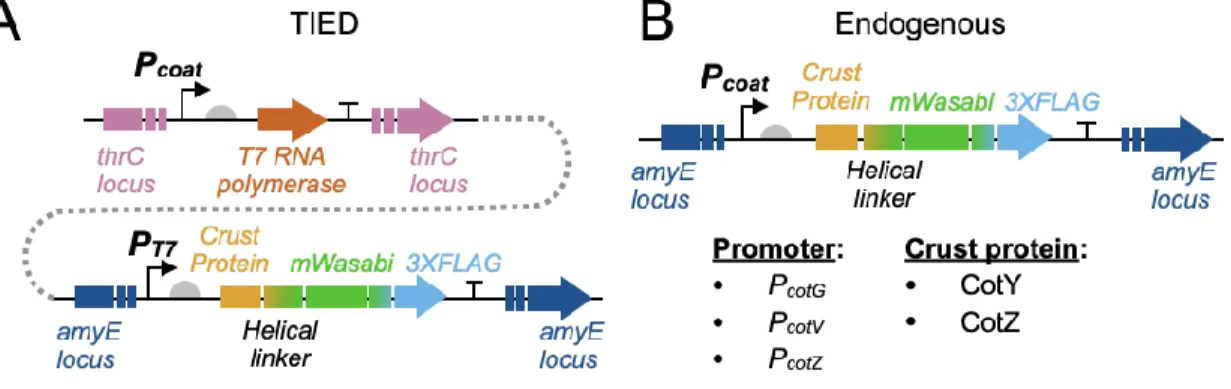
Expression and display of UreA from Helicobacter acinonychis on the surface of Bacillus subtilis spores. Surface representation of lipolytic enzyme, lipase A and lipase B from Bacillus subtilis on the Bacillus subtilis spore.
T7 RNA POLYMERASE ENABLED HIGH DENSITY PROTEIN DISPLAY ON BACILLUS SUBTILIS SPORES
- Introduction
- Results and Discussion
- Conclusions
- Materials and Methods
- Supplementary Tables and Figures Table S2.1 Protein and peptide sequences Table S2.1 Protein and peptide sequences
- Acknowledgements
- References
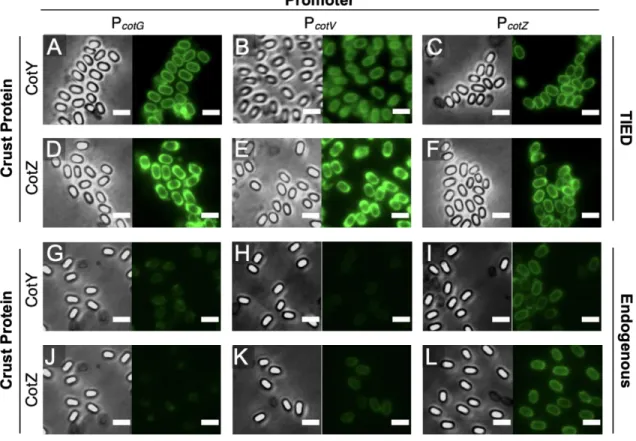
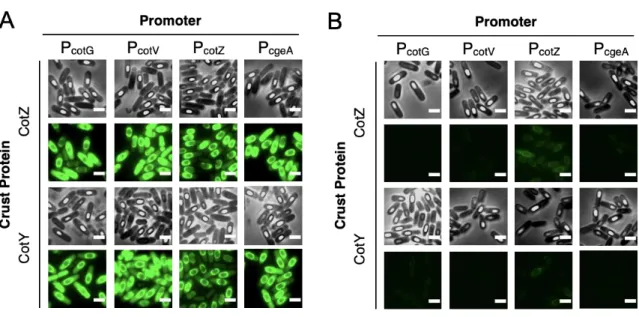
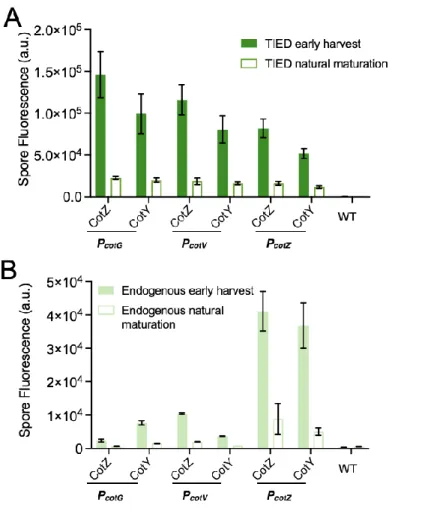
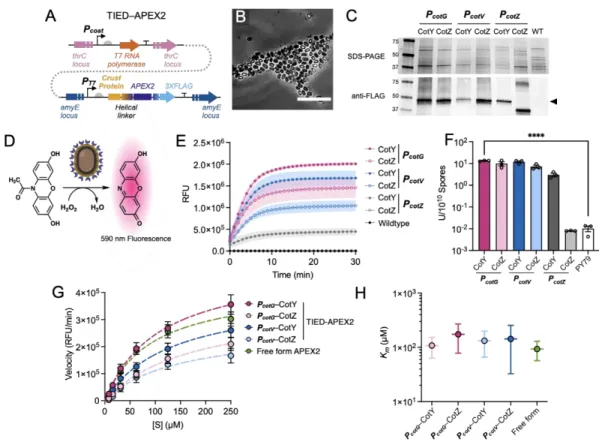
We further investigated the distribution of fluorescent fusion proteins by microscopic analysis and measurement of bulk fluorescence of purified spore solutions. Although the TIED system shows a higher overall level of expression of the fluorescent fusion proteins (Figure 2.2A), as the prespores approach maturation, fluorescence emission from the spore. Bulk spore fluorescence of TIED variants prepared by the early harvest method as a function of total fluorescent protein expression.
A total of six TIED combinations and their endogenous counterparts were constructed, with the coat protein promoters directly driving the expression of the fusion protein. Competence was induced in selected clones for subsequent transformation of the construct for T7 RNAP expression. Data were fitted with linear regression (dotted line). B), Bulk spore fluorescence of the endogenous counterparts prepared via the early harvest method as a function of total fluorescent protein expression.
STRESS-TOLERANT, RECYCLABLE, AND RENEWABLE
BIOCATALYST PLATFORM ENABLED BY ENGINEERED BACILLUS SUBTILIS SPORES
Introduction
Due to its specificity, efficiency and environmental friendliness, enzyme catalysis offers unique advantages for a wide range of chemical transformations.1-4 With the advancement of computational tools and directed development in recent years, enzyme engineering has expanded the portfolio of available biocatalysts beyond the repertoire of nature.5-8 Despite their apparent advantages in chemical and pharmaceutical production processes, the use of enzymes in a scalable and economical way remains a challenge for a wide range of applications. In addition to expensive purification, enzymes are less attractive in terms of stability and reusability compared to synthetic catalysts. So far, a series of enzymes including α-amylase, lipase and β-galactosidase have been placed on the surface of B.
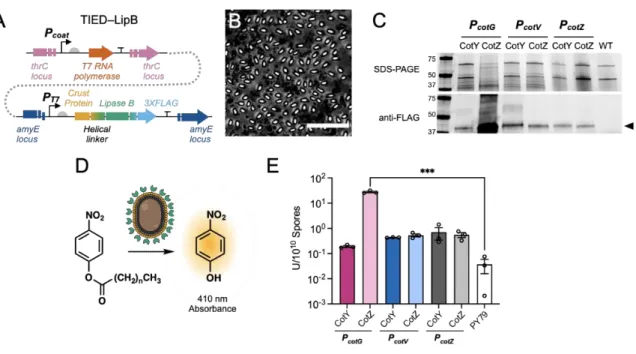
Furthermore, quantitative and substrate-specific characterizations of catalytic behavior of spore-immobilized enzymes compared to their respective free forms remain elusive. This development was made possible by T7 RNA polymerase-activated high-density protein display on bacterial spores (TIED) technology. In addition to being recyclable for multiple uses, enzyme-displaying spores can also be completely renewed when their activity is exhausted, allowing unlimited reuse of immobilized biocatalysts.
Results and Discussion
- Catalytic Performance of TIED-LipB
- Catalytic Performance of TIED-LipA
- Catalytic Performance of TIED-APEX2
- Recycling and Renewal of TIED Enzymes
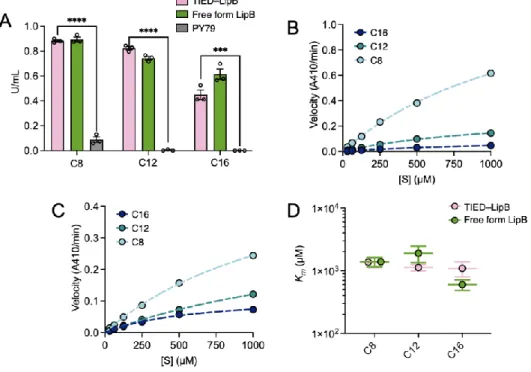
The apparent Michaelis-Menten (Km) constants of TIED-LipB and free-form LipB were similar to each other for all substrates (Figures 3.2D, S3.5C). Michaelis Menten constant, Km, of TIED-LipB (combination of PCotG and fusion partner CotZ) and free-form LipB for hydrolysis of each substrate, determined by Michaelis Menten fitting. Enzymatic activity of TIED-LipA variants and PY79 spores for hydrolysis of p-nitrophenol palmitate (C16, 1 mM).
Michaelis Menten constant, Km, of TIED-LipA (combination of PCotV and CotZ fusion partner) and free forms of LipB for hydrolysis of each substrate, determined by Michaelis Menten adjustment. P values were determined by two-tailed t-tests relative to baseline TIED-LipA activity (cycle 1). P values were determined by two-tailed t-tests relative to baseline TIED-LipA activity (cycle 1).
Conclusions
The maximum reaction rate (Vmax) and the Michaelis Menten constant (Km) for the reactions catalyzed by TIED-LipB and free form LipB. The constants were determined by fitting the reaction rates at different substrate concentrations with the Michaelis Menten model. The maximum reaction rate (Vmax) and the Michaelis Menten constant (Km) for the reactions catalyzed by TIED-LipA and free-form LipA. P values were determined by two-tailed t-test against the intact activity of TIED enzymes (cycle 1).
Recycling and renewal of TIED-LipB. A), recycling of TIED-LipB (combination of PcotG promoter and CotZ fusion partner, trace OD600 of 1.0) for conversion of C8 (1.0 mM) to Tris-HCl. P-values were determined by two-tailed t-tests against the pristine activity of TIED enzymes (first cycle). P-values were determined by two-tailed t-tests against the pristine activity of TIED enzymes (1st cycle).
Acknowledgements
New insights into the efficiency of alpha-amylase enzyme presentation on the Bacillus subtilis spore surface by adsorption and covalent immobilization. Transgalactosylation in a water-solvent biphasic reaction system with β-galactosidase displayed on the surfaces of Bacillus subtilis spores. Probing interactions between the Bacillus subtilis spore coat proteins CotY and CotZ using single molecule force spectroscopy.
The Bacillus BioBrick Box 2.0: expansion of the genetic toolbox for standardized work with Bacillus subtilis. The Bacillus BioBrick Box: generation and evaluation of essential genetic building blocks for standardized work with Bacillus subtilis. Flexibility of the spore layer formation program in Bacillus subtilis: bypassing the CotE requirement through CotH overproduction.
LEAK-FREE CELLULAR COMPOSITES FOR PRODUCTION OF RECOMBINANT PROTEINS
Introduction
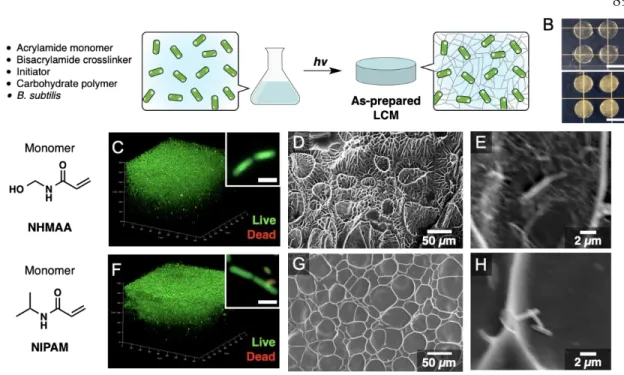
Recent advances in synthetic biology have expanded our ability to engineer cells for specific functional outcomes that may exceed those of natural systems.1,2 Inspired by the possibility of engineering "living" materials, several strategies for incorporating cells into smart three-dimensional materials have been One of the remaining challenges in this area is the development of methods to achieve safe cell confinement so that cells do not escape into the environment, while allowing the functional biomacromolecules produced to readily diffuse out of the materials and be collected for use. A recent strategy using a hydrogel with a core–shell architecture has been shown to achieve effective bioretention; however, the cross-linked network was designed to be permeable to nutrients but not to biomacromolecules, including proteins.11 The search for molecular design principles for scaffolds that provide safe cell confinement while allowing the permeability of recombinant proteins will open new opportunities for using the engineering life of materials. When reimmersed in an aqueous medium, trapped cells and spores in pre-dried LCMs exhibit metabolic activity, including the synthesis and secretion of recombinant proteins.
In particular, one of the scaffolds developed in this study, based on photopolymerization of N-(hydroxymethyl)acrylamide (NHMAA), achieves effective cellular confinement, showing no evidence of cellular leakage over a 72-hour period.
Results and Discussion
Image analysis of z-stacks revealed that 64% of the emission signals originated from SYTO-9 in samples reswelled in LB medium compared to 21% after reswelling in PBS (Figures 4.2B, 4.2C). Consistent with this result, high-resolution confocal imaging showed that the majority of cells in the PNHMAACell samples reswelled in PBS. In contrast, a characteristic pattern of alternating red and green staining was observed in samples of PNHMAACell cells reswollen in LB on high-resolution imaging (Figure 4.2B, inset).
Interestingly, Spo0A knockout cells in PNHMAASpo0A or PNIPAMSpo0A re-swollen in LB did not show the red-green alternating pattern (Figures S4.2B, S4.2D). Representative confocal image stacks of PNHMAAmRFP before (left) and after (right) desiccation and re-swelling in LB. Image analysis of the dehydrated and re-swollen PNHMAACell samples further suggests that cross-linked cells do not divide in LB (Figure S4.5).
Conclusions
Materials and Methods
Samples were immersed in 100 μL CTC solution (5 mM), incubated at 37°C for 30 min, and subsequently washed with PBS before imaging. Cell and spore composites were immersed in a 2.5% glutaraldehyde solution in 50 mM sodium cacodylate buffer for 2 h at 37 °C. Before the measurement, freeze-dried samples were transferred to a conductive carbon tape, broken with a blade and coated with a Pd layer of 10 nm in thickness before imaging.
After dehydration on PTFE plates with desiccants (Drierite®) for 36 hours, the composites were immersed in LB or PBS (1.5 ml/composite) at 37 hours. Three dehydrated cell or spore composites were immersed in 15 ml LB and placed in a shaking incubator. at 37°C, 250 rpm. Twelve dried cell composites carrying AmyQ-mRFP or mRFP-expressing cells were immersed in 5 ml LB.
Supplementary Tables and Figures
Characterization of PNIPAMSpores Representative confocal laser scanning microscopy images of PNIPAMSpore samples prepared from the dual fluorescence (DF) strain. A), Pristine PNIPAMSpores, (B), PNIPAMSpores reswollen in PBS after dehydration and (C), PNIPAMSpores reswollen in LB after dehydration. Re-swelling of PNIPAMSpores in LB for 12 h resulted in biofilm formation on the surface: Percentages of cells and spores were not determined for samples containing large cell clusters. Representative confocal laser scanning microscopy images of PNHMAACell samples prepared from the double fluorescence (DF) strain.
Representative confocal laser scanning microscopy image stacks (left: low resolution, right: high resolution) after 12 h re-swelling of dehydrated LCMs prepared with B. A), PNHMAASpore re-swollen in PBS, (B), PNHMAASpore re-swollen in LB, (C), PNIPAMSpore re-swelled in PBS, and (D), PNIPAMSspore in LB.

Acknowledgements
Effect of chain transfer agent on the crosslinking of poly (n-butyl methacrylate-co-N-methylol acrylamide) latex particles and films. Performance of cross-linked poly(vinyl acetate-co-N-methylolacrylamide) latex adhesive, II. part: Fracture mechanics and microscopic studies of durability. Live cell imaging of germination and outgrowth of individual spores of Bacillus subtilis; effect of heat stress quantitatively analyzed with SporeTracker.