Introduction
Development of a tunable integrase mediated differentiation archi-
Experimental considerations for differentiation architectures
By strategically placing integrase binding sites in the genome, a single integrase-mediated recombination event can simultaneously activate and deactivate the expression of desired genes [ 20 – 22 ]. To allow differentiation to control different expression programs, genes regulated by the recombination event must encode proteins that control the expression of multiple genes, as in the case of transcription factors, sigma factors or orthogonal RNA polymerases.
Deterministic modeling of integrase-mediated differentiation
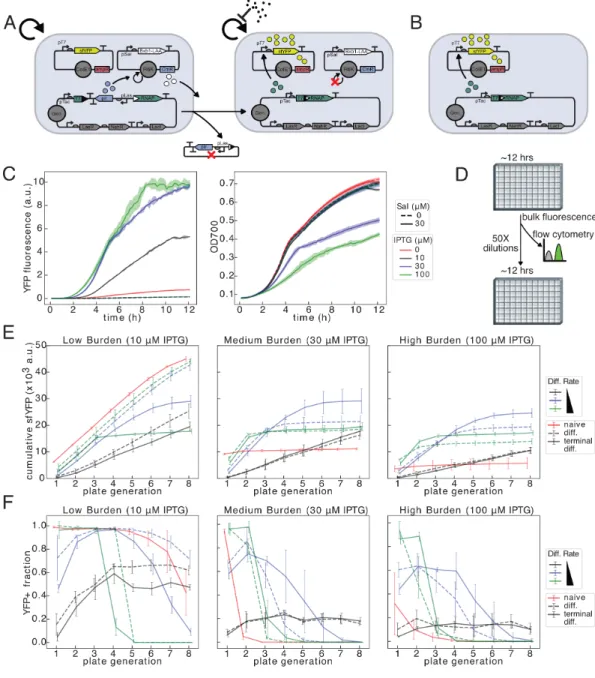
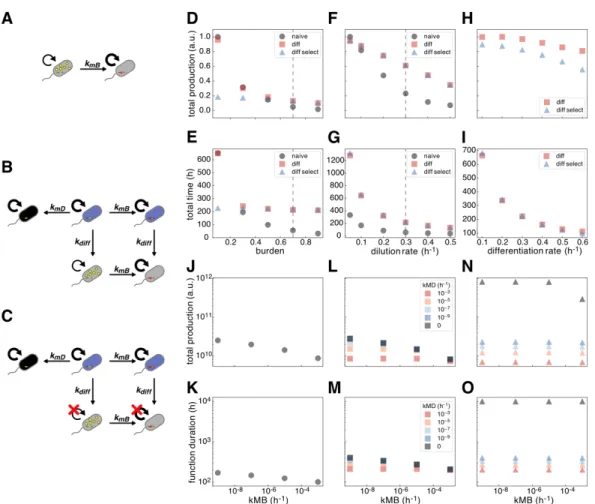
In the case of differentiation alone, we propose a class of mutations in progenitor cells that would destroy the cell's ability to undergo differentiation (Figure 2.1B-C). Because the behavior of the two architectures involving differentiation is also affected by the differentiation rate, we optimized this parameter in each case to maximize the total production (Figure 2.2A) or the duration of the population function (Figure 2.2B).
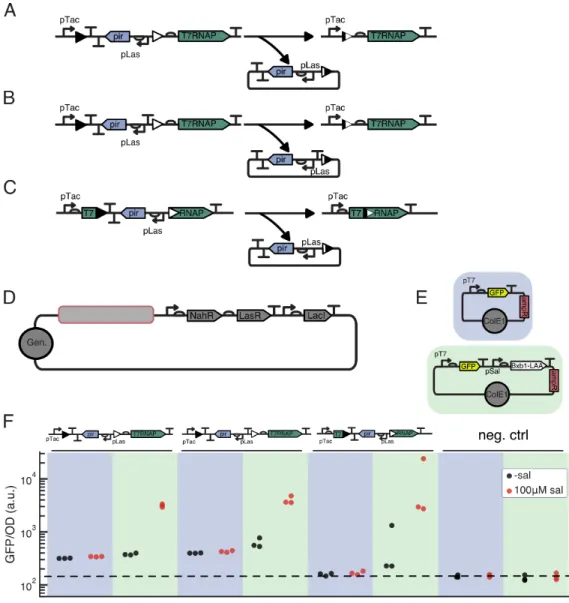
Integrase mediated differentiation allows tuning of population distri-
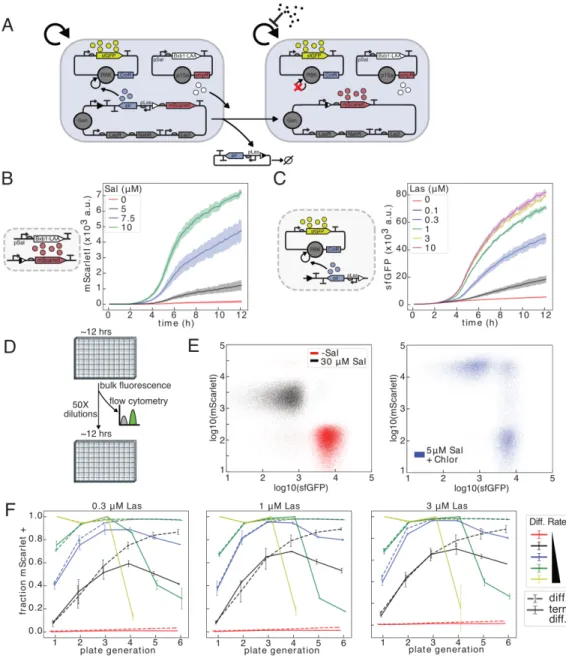
The abundance of differentiated cells is inferred from mScarletI fluorescence, and the relative abundance of the R6K plasmid is inferred from the expression of the encoded constitutively produced green fluorescent protein sfGFP. This is revealed at all Las-AHL concentrations in flow cytometry of the fifth plate generation for 10 µM salicylate, and in the fourth plate generation for 15 µM salicylate (Figure 2.3F).
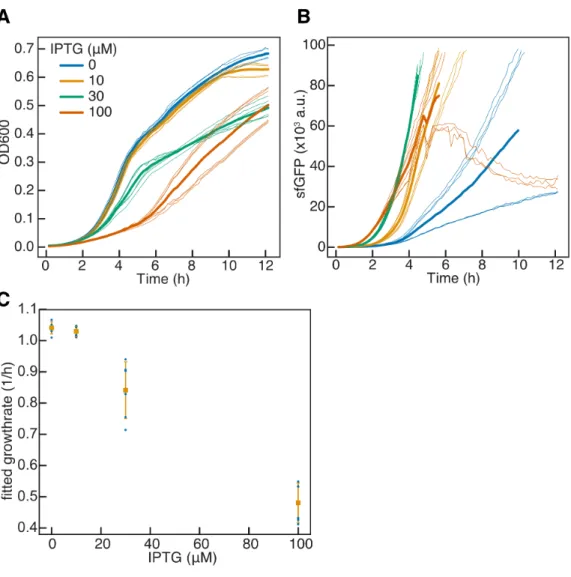
Differentiation-activated T7 RNAP expression improves burdensome
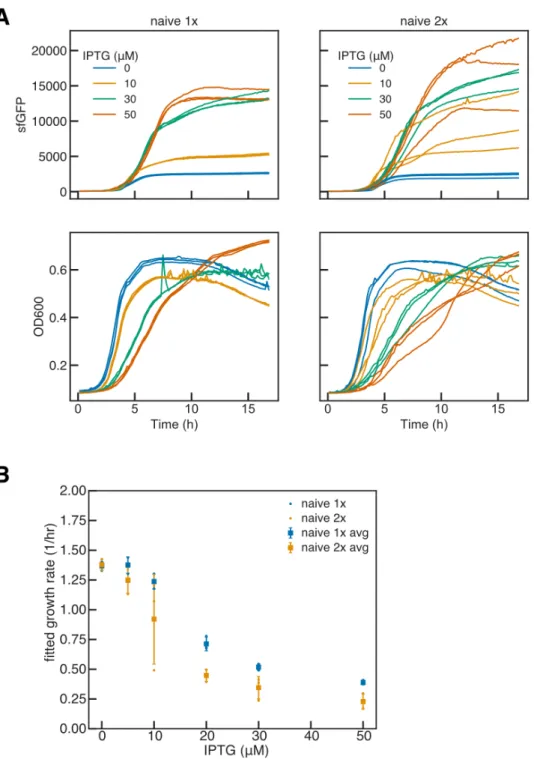
With higher load generation, the benefit of differentiation for both cycle duration and total generation becomes apparent. This benefit to total production is increased at the highest induction level of IPTG, by ~3.4x and.
Discussion
Although this may not be surprising, we recognize that differentiation mutation may be facilitated by errors during integrase-mediated recombination, the rate of which has not been quantified to our knowledge. This can be achieved by the integration of two modified copies of the differentiation cassette or by two redundant differentiation mechanisms using orthogonal integrases.
Model implementation
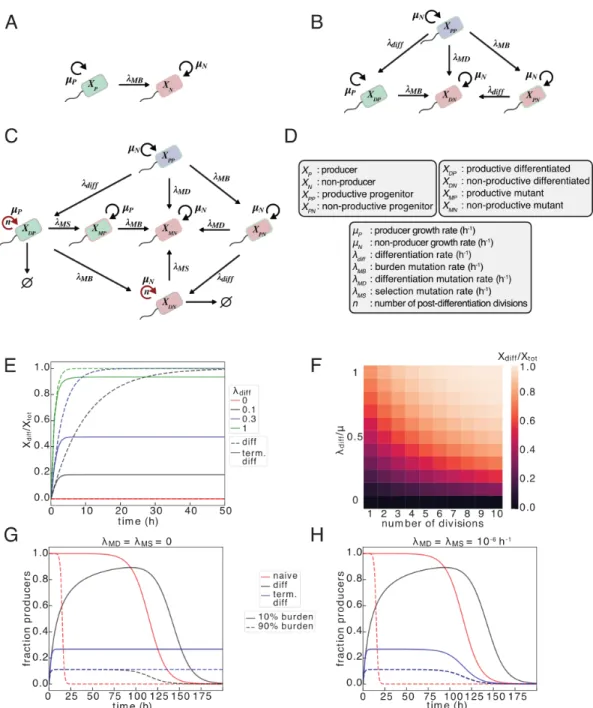
Selection Mutation (𝜆𝑀 . 𝑆): Mutation occurring in differentiated cells (either producers or non-producers) that allows cells to evade selection and proliferate indefinitely. In the modeling performed for this paper, we assume that all differentiated cells capable of further cell divisions (𝑖 < 𝑛) grow and produce at the same rate.
Materials and methods
For long-term experiments, cells were diluted 1:50 after ∼12 h growth in the same media conditions in a replicate dish. 50,000 unrelated events were recorded for each sample and the results were analyzed with the custom Python code available in the supplementary information listed in the Github repository.
Supplementary Information
This was done similarly for naïve (0 and 30 𝜇M IPTG) and differentiation without selection (7.5 𝜇M salicylate/0 𝜇M IPTG and 7.5 𝜇M salicylate/30 𝜇M IPTG), but PCRs were unsuccessful.
Acknowledgements
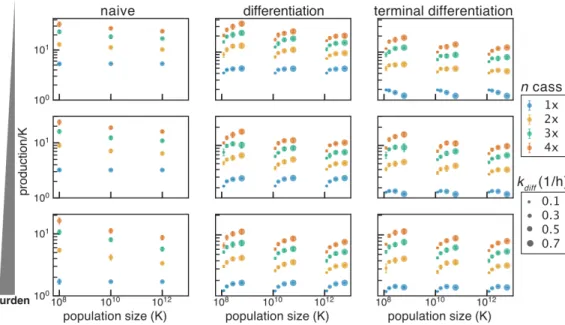
In the naive case, additive selection results from the expression of the function depending on the number of copies. However, in the case of terminal differentiation, additive selection exists only for integrase and differentiation mutations. We further look at the effect of copy number in the case of recessive selection as the population size changes.
Improving the evolutionary stability of differentiation circuit
Introduction
This served as a useful initial demonstration of the viability of this strategy, but the circuit design and implementation was lacking in several ways. Chief among these shortcomings was the susceptibility of the differentiation circuit to being broken by a variety of single mutations. Furthermore, by placing the expression of the integrase responsible for catalyzing the differentiation event on a plasmid, it is inherently subject to both more opportunities for mutation and to random plasmid partitioning effects that can accelerate the expansion and fixation of beneficial mutations [11].
Reviewing intuition from deterministic modeling
However, exponential increases in the load mutation rate only result in a linear decrease in duration and total production. -C) Gray cells containing product (yellow dots) are producers and grow more slowly than all other cells that are non-producers. (A) Naive inducible or constitutive expression in which producing cells undergo a last mutation at a rate 𝑘𝑀 𝐵(h −1)which abolishes expression and restores wild-type growth in non-producing cells. Progenitor cells and differentiated producers incur charge mutations at a rate of 𝑘𝑀 𝐵 (h−1), and progenitor cells incur differentiation mutations at a rate of 𝑘𝑀 𝐷 (h−1).
Differentiation circuit development
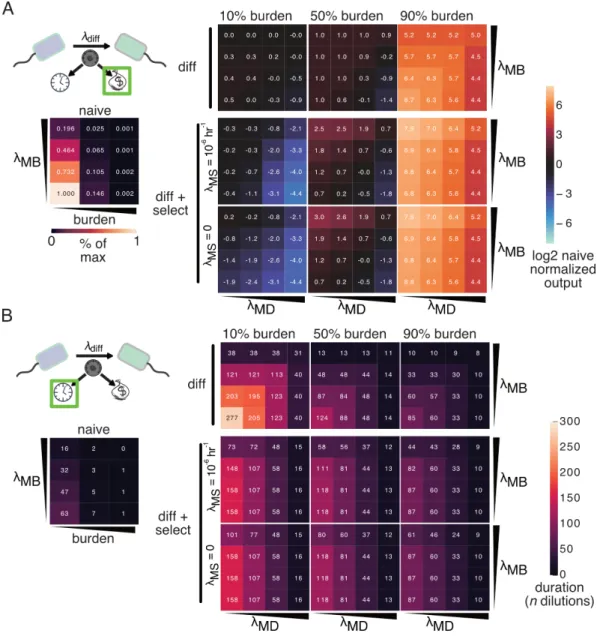
Decreasing the differentiation mutation rate increases both metrics regardless of the load mutation rate, and critically, if this rate is zero, we achieve undefined function (Fig 3.1N-O). From our initial deterministic modeling, we observed that reducing the differentiation mutation rate or probability improves the duration of terminal differentiation circuit function, regardless of the mutation rate of the load. If a second identical cassette were to be integrated, recombination of both cassettes would be required to stop replication of the R6K plasmid and allow growth restriction mediated by antibiotic selection.
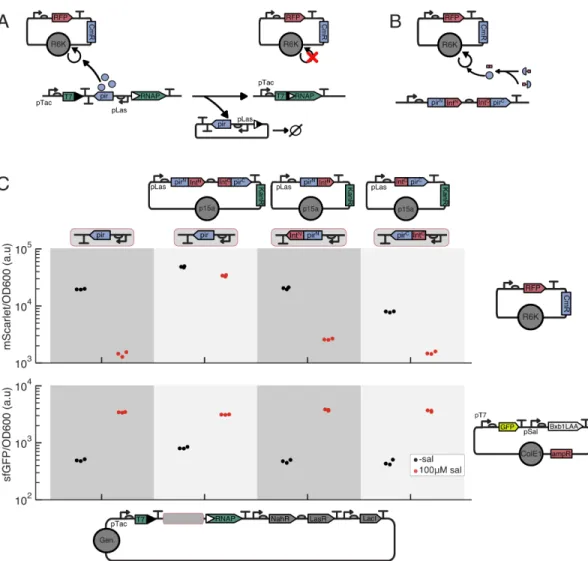
Experimental evaluation of differentiation and terminal differentia-
At low load (higher 𝜇𝑃), terminal differentiation works worse, but better as the load increases (Figure 3.17). Indeed, we see that the inclusion of stochastic plasmid loss in conjunction with a model of antibiotic degradation and growth inhibition can negatively affect the performance of naïve and differentiation architectures in a strain-dependent manner, and that this effect is much larger in the case of two cassettes (Figure 3.17). We see this both in the total production achieved (Figure 3.17) and in tracing the population of cells that have lost the plasmid (Figures 3.21-3.26).

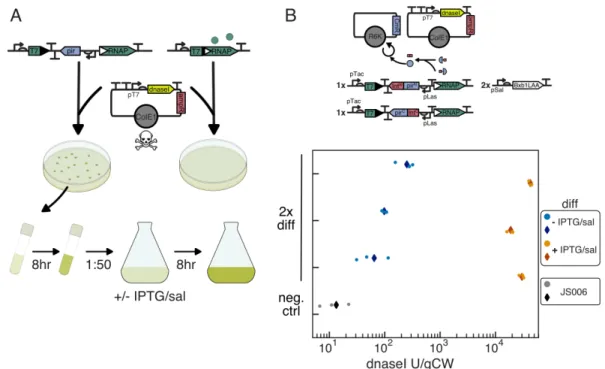
Differentiation enables expression of toxic functions
Cells with one (naive 1x) or two (naive 2x) copies of genomically integrated T7 RNAP were transformed with ColE1 KanR pT7 GFP, plated in LB + kan, and grown in LB + kan before dilution under experimental conditions. 1000 cfu respectively, while the naive 1x and naive 2x strains yielded 1 and 0 colonies respectively, compared to > 104 cfu for both when transformed with the Cole1 AmpR pT7 GFP control (Figure 3.6A, Table 3.1). Growth of induced and uninduced cultures showed that the dnaseI plasmid minimally affected growth when T7 RNAP is not expressed and that 20 m m salicylate induction likely resulted in more complete differentiation in a large shake culture compared to small volumes in 96-well microplates in the former. experiments.
Discussion
Differentiation was particularly beneficial compared to naïve expression with a higher burden as expected from modeling, and the redundancy and robustness for differentiation mutations provided with the split-𝜋 protein terminal differentiation was effective. We further demonstrated computationally that the robustness of terminal differentiation circuits to burden mutations affecting T7 RNAP expression and extend plasmid loss to the general case of plasmid mutations disrupting the function of interest. But because a system like this with terminal differentiation where effects of plasmid instability can be substantially or completely mitigated, we can potentially get the stability benefits of genomic integration with the ease of plasmid transformation.
Model implementation
Since the mutation rate is also proportional to the number of loci that could be mutated, the general case of the 𝑛cassette is this. The naive case is treated identically with the number of 𝑃 cassettes considered instead of 𝐷 𝑃with differentiation. The cell count is then subtracted from the source genotype and added to the target genotype.
Materials and methods
Differentiation strains were cotransformed with R6KCmR-mScarletI and ColE1 AmpR pT7 GFP or ColE1 KanR pT7 GFP, recovered in SOC with 30 nM Las-AHL and plated on LB agar containing 34 µg/ml chloramphenicol, 30 nM Las-AHL and 100 𝜇g/ml carbenicillin or 50 µg/ml kanamycin. Naive strains were grown in LB with the appropriate antibiotic, and differentiation strains were grown in LB with chlorine and carb or jug with 10 nM Las-AHL. For dnaseI expression experiments, 2x split-pir differentiation cells were cotransformed with an empty R6KCmR plasmid and the isolated ColE1 AmpR dnaseI expression plasmid, recovered in SOC with 30 nM Las-AHL and plated on LB agar with carb/chlorine/30 nM Las-AHL.
Acknowledgements
Buffers used for assay were as described in Kunitz [43], although to allow simultaneous measurement of many samples and to avoid problems we observed with background absorption in russel lysate when performing the Kunitz assay, we developed a fluorescence-based assay similar to Vogel and Frantz [44]. Standards were prepared by adding various amounts of dnaseI (Invitrogen AM2222) to JS006 lysate diluted 1:10 in 0.85 percent NaCl. Samples for testing were diluted 1:10 or 1:50 in 0.85 percent NaCl, and 10❜L of sample or standard pipetted with a multi-channel pipette into triplicate wells immediately prior to testing.
Supplemental Figures
In addition, the effect of the degree of differentiation in the case of terminal differentiation is also interesting. Here we see that in the absence of plasmid mutation or shared antibiotic resistance, recessive selection generally outperforms additive selection (no difference between additive and recessive with copy number 1). If we model the case where we have just achieved integrase expression independent of copy number, we see no benefit in the context of terminal differentiation.
Development of differentiation circuit architectures for scaling in
Introduction
We demonstrated both computationally and experimentally that this redundancy and robustness to mutation improves the evolutionary stability of the terminal differentiation architecture and further demonstrated that this system can enable the expression of a toxic protein. In doing so, we are motivated by the important feature of terminal differentiation architecture, which protects cargo mutations—those that disrupt engineered functions of interest—from evolutionary forces. By addressing the evolutionary stability of terminal differentiation architecture, we can address this for any biologically possible function.
Considerations for scaling terminal differentiation
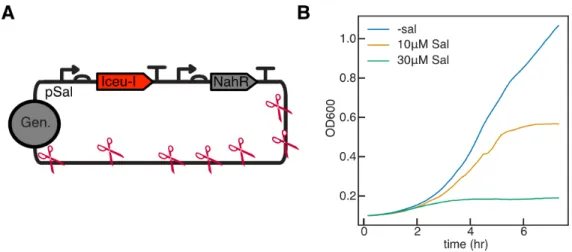
We consider additional means to extend this redundancy in the terminal differentiation architecture beyond what we achieved with the split-𝜋 protein, and demonstrate a proof of concept experimental circuit design that should allow scaling to arbitrary𝑛. In the ideal case, an identical construct could be integrated into the genome repeatedly, with each integration providing an additional layer of redundancy. In a demonstration of the utility of chromosome-free bacterial cells, the homing endonuclease I-ceuI was used to induce double-strand breaks in the genome with subsequent genome loss mediated by endogenous nucleases.
ETERNAL: A terminal differentiation architecture with scalable re-
To scale up the redundancy of this circuit, it is fully integrated into the genome. We further propose ways to incorporate delay in I-CeuI expression via an intermediate step of T7 RNAP-driven 𝜙C31 integrase expression, with 𝜙C31 subsequently activating I-CeuI expression (Figure 4.2B). However, using a medium strength RBS (B0032) did not allow sufficiently high expression of Bxb1 integrase, and using a high strength RBS (B0034), although allowing sufficient expression, resulted in leaky expression of the integrase in the absence of induction.
Model exploration of redundant architectures
In the case of a single cassette with a lower load (30%), increasing the differentiation rate above 0.1 h−1 reduces the total production. However, in the case of two or more cassettes, increasing the differentiation rate between 0.1 and 0.7 hours universally increases the total output. In the differentiation system we have implemented here, the differentiation rate is significantly lower and dependent on the steady-state concentration of integrase protein.
Materials and methods
Acknowledgements
Determination of the optimal aligned spacing between the glans-dalgarno sequence and the translation initiation codon of escherichia coli m RNAs. Multicellularity makes somatic differentiation evolutionarily stable. Proceedings of the National Academy of Sciences of the United States of America. Overproduction of the toxic protein, bovine pancreatic DNasel, in Escherichia coli using a tightly controlled T7 promoter-based vector.Gene136,337–340.
Conclusion