I especially want to thank Professor Arnold for helping me understand some of the intricacies of natural evolution, directed evolution and high-throughput screening. James Link helped me perform enzymatic assays with radioactive chemicals and taught me some fundamentals of molecular biology to support studies of aliphatic ketone-containing amino acids. Her advocacy and encouragement have been invaluable in helping me persevere and persevere through all the challenges of completing a doctorate at Caltech.
The first part of this thesis describes established methods to incorporate non-canonical amino acids into proteins and attempts to exploit the properties of non-canonical amino acids in areas such as protein structure determination, protein and organism evolution, immune system modulation and proteomics . Replacement of leucine with homoisoleucine in coiled-coil peptides stabilizes these proteins, as evidenced by the increase in the thermal denaturation temperature of the coiled-coil. The properties of the single-chain variable fragment form of a model anti-digoxin antibody have been studied after replacing the methionine residues of the protein with methionine analogs containing alkyne, azide, and aliphatic side chains.
Experiments with antibody fragments displayed on the surface of Escherichia coli cells showed that replacement of the methionine residues of the fragment with an analog containing an alkyne side chain reduced the fluorescence levels of cells treated with fluorescently labeled antigen to background levels, indicating loss of binding function. The replacement of methionine with azidohomoalanine in several variants lowers the dissociation constant of the fragment by a factor of two.
LIST OF TABLES
Expanding the Scope of Protein Science and Engineering with Noncanonical Amino Acids
Methods for incorporating ncAAs into peptides and proteins are either chemical, biosynthetic, or a combination of the two. The inability of the aaRSs of the in vitro translation system to recognize the tRNA (chemically acylated with the ncAA of interest) renders the tRNA orthogonal to the translation system, a recurring concept in site-specific incorporation of ncAAs into proteins. Finally, the ncAA (figure 1.3c) to be added to the genetic code must not be a substrate for any of the wild-type aaRSs in the cell.
This technique can be used on proteins by residue-specifically replacing one of the canonical amino acids in the molecule with an amino acid containing a heavy atom. Residues 7–10 have been incorporated residue-specifically into proteins with high substitution of the corresponding canonical amino acids. Judicious incorporation of ncAA residue- and site-specifically into protein samples can eliminate some of the complexities associated with NMR studies of proteins.
Fluorescent amino acid 53 has been used in FRET studies of the GPCR tachykinin neurokinin 2 (NK2) to study the structure of the protein (210, 211). One report on the successful incorporation of a glycosylated amino acid into proteins using an in vitro translation system has appeared in the literature (279), suggesting that future attempts to incorporate glycosylated amino acids directly into proteins may yet prove fruitful. Interestingly, several of the mutations observed in the final isolated mutant were also observed after screening.
Many clones isolated after three rounds of panning were found to contain amino acid 85. The versatility of bioorthogonal chemistry allows the same set of ncAAs to be used to visualize and identify newly synthesized proteins. Tang Y & Tirrell DA (2002) Reduction of Escherichia coli leucyl-tRNA synthetase editing activity allows incorporation of novel amino acids into proteins in vivo.
Hohsaka T, Ashizuka Y, Sasaki H, Murakami H, & Sisido M (1999) Incorporation of two different non-natural amino acids independently into a single protein through extension of the genetic code. 2010). Investigation of the structure and function of the tachykinin neurokinin-2 receptor through biosynthetic incorporation of fluorescent amino acids at specific sites. A new strategy for the synthesis of glycoproteins. 2009) Site-specific incorporation of mucin-type N-acetylgalactosamine-alpha-O-threonine into protein in Escherichia coli.
2008) Immunochemical interruption of self-tolerance. 2009) Mechanistic studies of immunochemical interruption of self-tolerance by unnatural amino acids. Knight RD, Freeland SJ and Landweber LF (2001) Rewiring the keyboard evolvability of the genetic code. A set of codons specifying one of the twenty canonical amino acids is "reprogrammed" to code for ncAA.
The amino acid to be "added" to the genetic code should not be a substrate for any of the endogenous aaRSs.

Homoisoleucine: A Translationally Active Leucine Surrogate of Expanded Hydrophobic Surface Area
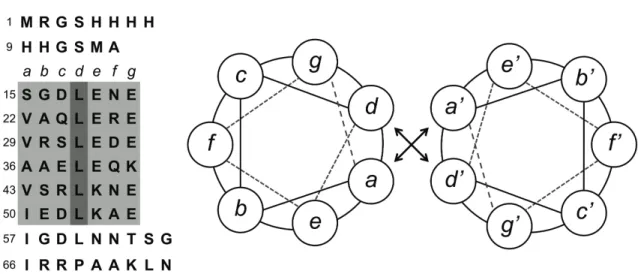
Replacement of Leu by Hil stabilizes coiled-coil peptides, as shown by increasing the thermal denaturation temperature. A1 contains six heptad repeats, designated (abcdefg), which mediate self-association of the peptide in aqueous solutions. Previous studies have shown that replacement of leucine by (2S,4R)-trifluoroleucine (Tfl, 3) or hexafluoroleucine (Hfl, 4) at the d positions of the heptad repeats results in significant stabilization of the coiled coil structure of A, presumably by enhanced hydrophobic interactions between fluorinated peptide strands.
Marsh and colleagues have argued that the “efficient packing” of the larger fluorinated amino acids into helix bundle cores may be more important than fluorination itself (18). Expansion of hydrophobic side chain volume at the d-position of the heptad repeat constitutes an effective strategy for. Although Hil and Tfl behave similarly in raising the melting temperature of A1, other experiments suggest important differences in the behavior of water adjacent to hydrocarbon and fluorocarbon side chains.
Aliquots of the resuspended cultures were supplemented with Hi (0.5 mM) or Leu (0.3 mM), shaken at 37 °C for 15 min, and induced by the addition of IPTG (1 mM final concentration). The purified samples were then analyzed by matrix-assisted laser desorption ionization (MALDI) mass spectrometry on an Applied Biosystems Voyager DE Pro instrument at the mass spectrometry facility in the Caltech Division of Chemistry and Chemical Engineering. For quantitative analysis, the rest of the trypsinized samples (not desalted) were sent to the Protein and Peptide Mass Analytical Laboratory (PPMAL) of the Beckman Institute at Caltech for liquid chromatography tandem mass spectrometry (LC/MS/MS) analysis.
Assuming that substitution of Hil in place of Leu is a random event, the ratio of the peak areas yields the quantitative substitution level in a given sample. The ratios of the areas of two substituted peaks are more relevant in this particular case because the total area A cannot be determined from the available experimental data. The identities of the peptides in the SLEDEAAELEQK series were confirmed by tandem mass spectrometry.
Structures of amino acids were minimized using Hartree-Fock calculations with a 6-31G* basis in vacuum, followed by determination of the molecular surfaces. The portions of the spectra shown include the m/z region in which the peptide LKNEIEDLKAEIGDLNNTSGIR occurs with or without Hil substitution. Kinetic parameters are reported as means determined from three independent experiments with errors reported as means of the 95% confidence intervals.
Hydration Dynamics at Fluorinated Protein Surfaces
Fluorinated amino acids have attracted special attention (4-16) because of the unusual solubility properties of fluorinated hydrocarbons. The nature of the protein hydration layer can be influenced not only by the topographic and electrostatic properties of the protein surface (24) but also by the physical and chemical properties of individual surface-exposed residues (27, 30). Self-association of the peptide juxtaposes the a and d positions and results in the formation of a hydrophobic core.
Analysis of each protein showed that the overall structural properties of the molecules were generally insensitive to genetic mutations and the incorporation of non-canonical amino acids. The steady-state fluorescence emission spectrum of Trp depends on the degree of exposure of the Trp side chain to water (36). These observations indicate that the Trp residues are exposed to the aqueous environment (consistent with the original design) and are not involved in the oligomerization of the proteins.
The minimal change in the steady-state fluorescence spectrum caused by replacing Leu with Tf1 or H1 suggests similar features of the hydration region probed by Trp (eg, the effective number of water molecules in the hydration shell). Despite these concerns, the dynamics results for A37L-L remain consistent with the conclusions of the paper (see below). The hydration dynamics of the proteins were well represented by triple-exponential decays with characteristic time scales of and 10-61 picoseconds.
The 70% increase in the relaxation energy (333 cm−1) of the slowest component (E3) compared to the nonfluorinated S31L-L (194 cm−1) is indicative of the dramatic slowing down of the hydration dynamics near. On the other hand, for A37L, increasing the size of the hydrophobic surface does not seem to affect the hydration dynamics as much. Therefore, the delay of the dynamics upon fluorination is much more pronounced for A37L-T than for the corresponding fluorination of S31L.
In the case of the S31L-T protein, one sample was made according to the procedure described below for the production of Hil-containing proteins. The uncertainty in the anisotropy amplitude, rTrp and r∞, is not affected by the limited time resolution of our data. The regions in the three XICs allow determination of the extent of incorporation of non-canonical amino acids in place of leucine.