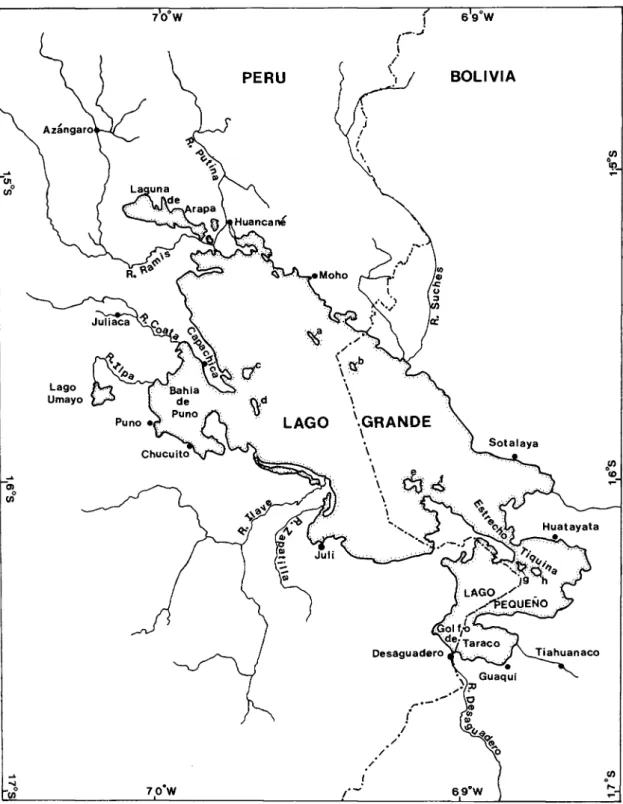
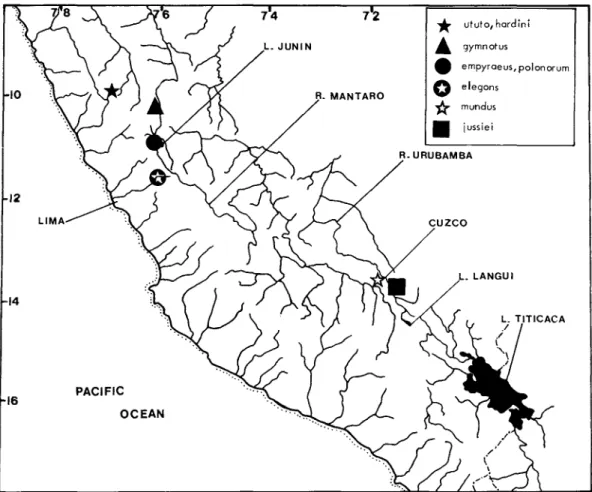
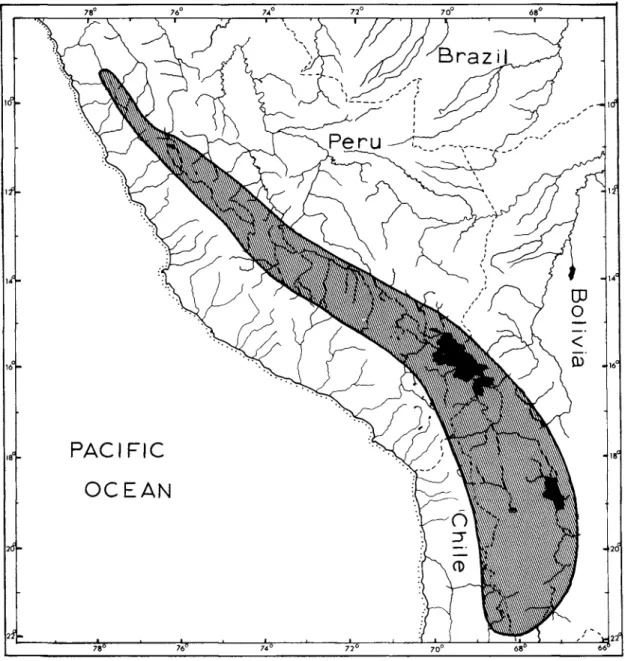
In the last revision of the genus Orestias, Chernavin (1944a) recognized 20 species and five subspecies. Limits of the known range of the genus Orestias, cross-hatched and outlined by a solid line. They were distributed, many unsorted and uncatalogued, to the collections of the California Academy of Sciences and the Field Museum of Natural History.
The Percy Sladen Expedition of 1937 greatly added to our knowledge of the Lago Titicaca ichthyofauna (see Transactions of the Linnean Society for a comprehensive report on the Percy Sladen Expedition). In a recent review of the killerfishes, order Cyprinodontiformes (Parenti, 1981), I suggested that Orestias has no close South American relatives, but is rather closely related to Anatolian cyprinodonts, and most closely to Kosswigichthys (including Anatolichthys ). I defer a discussion of the species assemblage concept and its relation to the phylogeny and biogeography of Orestias species to another paper (Parenti, in press).
Thus, the aims of this paper are to review the evidence for the relationship of Orestia with Anatolian cyprinodonts and the rest of the subfamily Cyprinodontinae;. John Simmons of the University of Kansas, Museum of Natural History, prepared Figures 52 and 53.
NOTE ON MATERIALS AND METHODS
Measurements were primarily used to aid species identification in conjunction with descriptions of what I defined as consistent differences, such as, for example, broad head (as in Orestias luteus, Fig. 7) versus a narrow head. All descriptions of cartilaginous structures are based on the examination of such preparations with comparisons made to the collections of exclusively alizarin red S-stained preparations of cyprinodontiforms in the BMNH and USNM.
RELATIONSHIPS OF KILLIFISHES OF THE TRIBE ORESTIINI
Aphanius species, which were judged to be more closely related to Kosswigichthys and Orestias than to other Aphanius species, could not be defined as monophyletic. Kosswigichthys asquamatus is unsealed; other species in the genus show a derived state of pore pattern and squamation as in "A." mint. General features of the specimen include a series of small neuromasts arranged in a "lyre-shape".
In both Aphanius fasciatus and the species Cyprinodontini, the dermosphenotic is a long bone with a deep trough for the sensory canal. Regarding the development of the gonads in Aphanius fasciatus, he stated (pp. The ovary consists of two components, the right and left gonads. Because goodeids are distantly related to orestiines, the early association of the gonads in Orestias, " Aphanius.”
The consistently high number of vertebrae in Orestias (30 or more) may be an additional defining character of the genus;. Pelvic fins and fin rings are absent in some members of the Cyprinodontini and Aphanius species, as well as in all Orestias (character 21).

PHYLOGENETIC ANALYSIS
Allen (Eigenmann and Allen, 1942) presented a dendrogram of Orestias species, together with his brief revision of the genus. A number of species of the mulleri complex, two species of the cuvieri complex (ispi and forgeti), and O. Aspects of the pattern can best be seen in the photograph of the holotype of O.
Distribution of the number of dorsal fin rays in Orestias Number of dorsal fin rays. Distribution of the number of anal fin rays in Orestias Number of anal fin rays. Frequency distribution of head length in Orestias (expressed in thousandths of standard length).
Frequency distribution of the width of the head in Orestias (expressed in thousandths of standard length). Frequency distribution of the depth of the body at the back of the head (expressed in thousandths of standard length). Depth of the body (table 10) is correlated in a number of species with the depth of the tail tree (table 11).
On average, all members of the agassii complex have a relatively deep caudal peduncle.

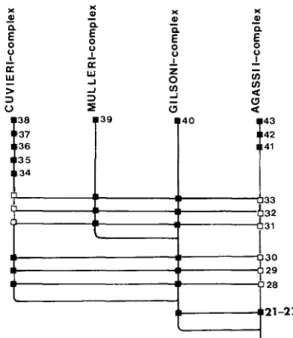
EXPLANATION OF SYNAPOMORPHY DIAGRAMS
Being deep-bodied is also a synapomorphy of a group within the agassii complex; that is, species of the luteus group and O. A deep caudal peduncle also occurs in crawfordi and incae, two relatively deep-bodied species of the mulleri complex. This derived character can also be described as an increase in the length of the lower jaw (fig. 32) which is the longest in the genus in cuvieri.
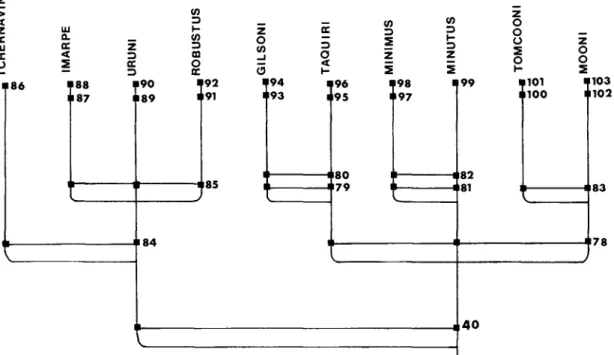
Therefore, this possibly further derived state is not used as a further defining character of the gilsoni complex. Also, in these two large species of the cuvieri complex, the dorsal scales are never as large relative to those lateral to the median dorsal ridge as in the agassii complex. In ispi and forgeti and species of the mulleri and gilsoni complexes, a median spine can be distinguished, but these scales are not thickened, nor smooth or granulated.
The four species of the cuvieri complex (cuvieri, pentlandii, ispi, and forgeti) are defined as a monophyletic group by the five synapomorhies (characters 34 through 38) discussed above for Figure 26. Relationships among species of the gilsoni- complex (fig. 29): the 10 species of the gilsoni complex are defined as monophyletic by the unique structure of the caudal fin, described as character 40 (fig. 26). There is a slight rise in the dorsal profile from the tip of the snout to the base of the dorsal fin, and a slight downward slope in the ventral profile from the tip of the mandible to the base of the anal fin (Fig. 72). ).
Species of the agassii complex are divided into three groups based on the squamation patterns described below. The number of scale rows increases posteriorly, becoming a series of about four to five rows at the base of the caudal fin. Pigmentation pattern characterized by a dark band at the base of the dorsal, anal and especially the pectoral fin rays (see for example Figure 46).
Orestias tschudii and richersoni are defined as sister species by a synapomorphy. 113) The body is laterally compressed, especially behind the origin of the dorsal fin. In the largest adults, body depth reaches nearly 35 percent of SL (Table 10), and is typically more than 29 percent. Orestias olivaceus, albus, farfani, rotundipinnis and luteus (the luteus group; Fig. 31) form a less inclusive monophyletic group defined by one synapomorphy. 121) Scales from the snout to the base of the dorsal fin, the preopercular and opercular regions and dorsal to the pectoral fin are thick and covered with dense granulations (Fig. 7).
SYSTEMATIC ACCOUNTS
DIAGNOSIS: A broad-headed (width varying between 29 and 38 percent of SL, Table 9), deep-bodied (depth at posterior extent of head varying between 29 and 38 percent of SL, Table 10) species of the luteus group of the agassii complex, which is most closely related to O. Orestias olivaceus, like all other species in the luteus group, has thick granular scales on the dorsal surface of the head and body. DIAGNOSIS: One of the four species (agassii, parinacotensis, ascotanensis, in addition to laucaensis) of Orestias with members in Chile.
The model can be seen in the photograph of the holotype (fig. 55), an adult female. Scales are missing from the abdomen, and the basal parts of the pectoral, dorsal and anal fins. DIAGNOSIS: A slender species of the agassii complex, in which males possess extremely ctenoid scales (up to 10 ctenii per scale).
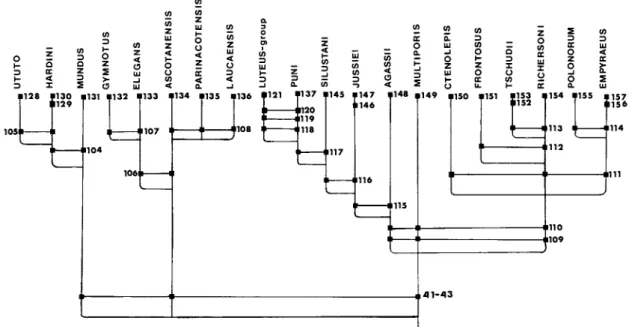
Orestias ascotanensis shares with other species of the agassii complex the sexual dichromatism described for O. A member of the agassii complex that does not differ from most other species in the complex by any morphometric or meristic characteristics (data for this species are summarized in Tables 3 to 12). The species of the genus Orestias are divided into four monophyletic groups, called species complexes.
Luteus species usually have an extremely broad body and a broad head.