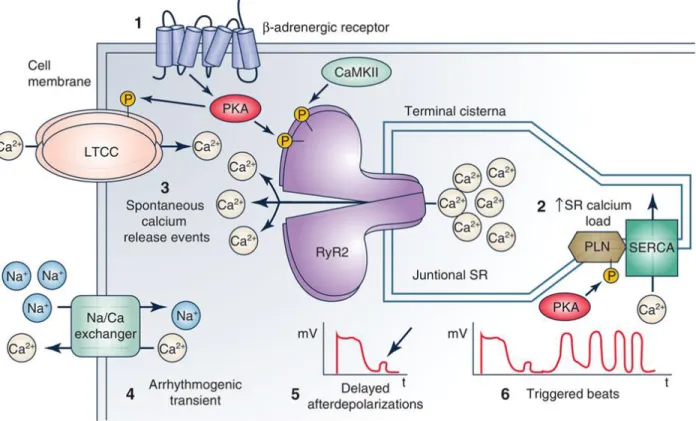
Calsequestrin (Casq) is the most abundant calcium-binding protein in the sarcoplasmic reticulum (SR) of skeletal and cardiac muscle (Rossi et al., 2021). Instead, FKBPs still bind to mutant RyR2 but cannot inhibit it (Zhang et al., 2016). The resulting cellular and SR calcium overload causes an increase in spontaneous calcium release, especially during catecholaminergic stimulation (Chopra et al., 2009).
The mutations caused a frameshift in the TRDN gene, leading to a syndrome called “triadin knockout syndrome” (Altmann et al., 2015). Increased HRC can activate RyR2 calcium release channels and contribute to CPVT (Liu et al., 2015). In vivo, artificially increasing resting heart rate with atropine or by overdrive stimulation prevented CPVT (Faggioni et al., 2013).
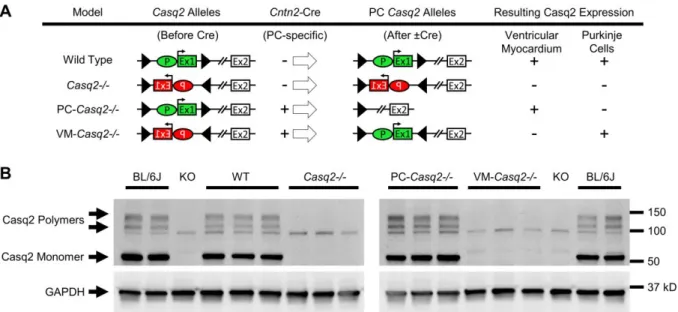
Surprisingly, deletion of Casq2 in the Purkinje network did not produce a CPVT phenotype (Figure 4) (Flores et al., 2018). Propafenone, another class 1C antiarrhythmic drug, has also been used to treat CPVT (Marx et al., 2019).

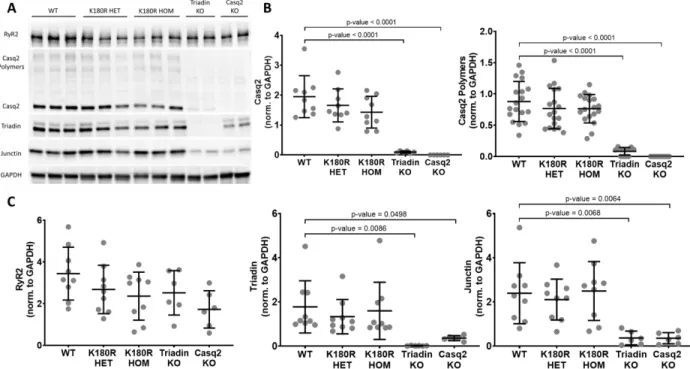
Functionally, Casq2 regulates the amount of calcium released from the SR through RyR2 during E-C coupling by buffering intra-SR calcium ( Zhang et al., 1997 ; Bers, 2002 ). K180R impairs the extent of calcium binding to the Casq2 polymer within the SR without altering the total SR calcium buffer or SR calcium content. The resulting NCX current integral represented total SR calcium, and the corresponding fluorescence signal amplitude represented free calcium during SR calcium.
The spontaneous opening rate of RyR2 and thus the calcium spark frequency increases with increasing SR calcium content ( Satoh et al., 1997 ). The height of the caffeine-induced calcium transient amplitude is a well-established measure of total SR calcium content (Bers, 2001). Given its role as the main calcium buffer in the SR, a decrease in Casq2 protein causes a reduction in total SR calcium content (Knollmann et al., 2006).
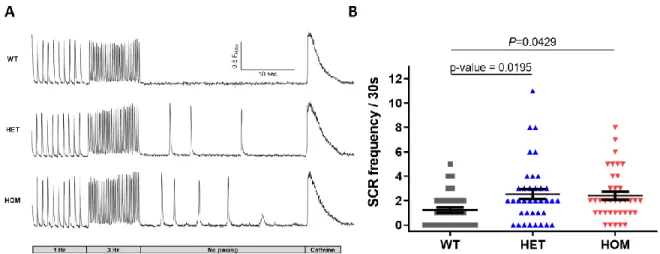
Due to the abnormal calcium processing that occurs in the K180R cardiomyocytes, we would hypothesize that the total SR calcium content is altered. The integral of NCX currents generated by SR calcium release is an established quantitative measure of total SR calcium content ( Bers, 2001 ). Our experiments showed that the K180R variant produces CPVT in vivo and increases spontaneous calcium release in vitro without affecting Casq2 levels or total SR calcium content.
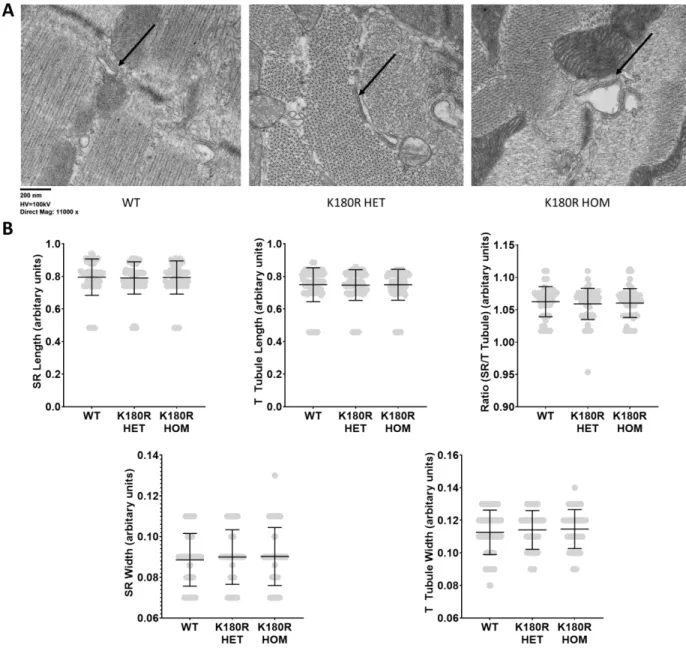
To test whether K180R alters the refractoriness of SR calcium release, isolated cardiomyocytes were loaded with fluo-4 calcium. Given that K180R reduced the refractoriness of SR calcium release, we reasoned that K180R impairs the ability of the SR to buffer calcium appropriately after calcium release. On the other hand, the amplitude of the SR calcium depletion signal (ie, the maximal change in intra-SR free calcium concentration during the SR calcium release event) was not different between groups.
We next measured intra-SR calcium buffering in intact cardiomyocytes at a fixed rate of 1 Hz (Figure 18). By affecting the dynamic buffering, K180R reduces the SR calcium release resistance of the cell. The impaired dynamic buffering increases the rate of calcium rise in the SR and thereby reduces SR calcium release.

Experimental and modeling studies have suggested that Purkinje cells in the cardiac conduction system are the cellular source responsible for arrhythmia generation in CPVT (Cerrone et al., 2007; Herron et al., 2010; Kang et al., 2010). Experiments in isolated Purkinje cells and intact hearts from the Ryr2/Ryr2R4496C mouse model have suggested that Purkinje cells in the cardiac conduction system are the cellular source responsible for inducing CPVT (Cerrone et al., 2007; Kang et al., 201; & Rudy, 2011). Optical mapping, commonly used to study the anatomical origin of ventricular arrhythmias, cannot resolve Purkinje cells from the surrounding ventricular cardiomyocytes in the sub-endocardium.
To generate transgenic mice expressing cre under the control of the Purkinje-specific Cntn2 gene, we used BAC recombination, as the approach used in the GENSAT program to generate Cntn2-EGFP BAC reporter mice ( Gong et al., 2003 ). These results suggest that loss of Casq2 in the conduction system alone is not sufficient to cause ventricular arrhythmias. The incidence of positive staining for Casq2 decreased with increasing distance from the nearest Purkinje cell, indicating that additional Casq2 expression was only in subendocardial myocytes located in front of Purkinje cells.
Casq2 expression in ventricular myocardium paired with Purkinje cells protects against arrhythmias, whereas Casq2 expression in Purkinje cells alone does not (Table 3). Cx43 is a gap junction protein expressed only in the ventricular myocardium but not in the atria and conduction system. Taken together, our experimental data indicate that ectopic beats in the CPVT mouse model are generated near or at the Purkinje–myocardial junction.
Given the unique geometric properties of the Purkinje myocardial junction, which facilitate retrograde conduction (Wiedmann et al., 1996; Huelsing et al., 1998), we hypothesized that ectopic beats are triggered by subthreshold membrane depolarizations (ie, the DADs) in the juxta -Purkinje ventricular myocytes. Retrograde excitation occurred as subthreshold DADs in the ventricular myocardium elicited action potentials in the PF (Figure 30A). Subthreshold delayed afterdepolarization (DAD)-like activity in ventricular cells of the Purkinje myocardial junction causes retrograde excitation of Purkinje fibers. A) Representation of the tissue block (left) used in the computational model.
Finally, in silico modeling further supported the experimental findings, showing that only DADs near the Purkinje-myocardial junction trigger action potentials in the PF. Analogous to our results, Casq2 deletion alone in the cardiac conduction system was not sufficient to generate a CPVT phenotype. The low rate of ventricular arrhythmias observed by Flores et al after Casq2 rescue in the sinoatrial node could be attributed to the sinus tachycardia in this model.
Although these studies were pivotal in showing that hiPSC-CMs are useful for studying CPVT, all CPVT hiPSC lines are from families with a specific CASQ2 mutation (Itzhaki et al., 2012). SgRNAs were inserted into the pSpCas9 vector and the insertion of sgRNAs was confirmed by sequence analysis (Wang et al., 2018). Cardiac differentiation was induced from hiPSC monolayers using small molecules CHIR99021 (Selleck Chemicals) and IWR-1 (Sigma) (Wang et al., 2021).
2 days after seeding, hiPSC-CMs were transduced with channelrhodopsin-2 (ChR2) adeno-associated virus (AAV)1 expression with H134R mutation fused to mCherry at a multiplicity of infection (MOI) of 1.2×104 viral genome/cell (Addgene viral prep #100054-AAV1) (Chavali et al., 2019). HiPSC-CMs were loaded with Fura-2 AM (ThermoFisher Scientific) as previously described in (Parikh et al., 2017). It is well known that Casq2 interacts with multiple proteins within the SR such as RyR2, triadin, and junctin ( Zhang et al., 1997 ).
Casq2 ablation has been reported to be associated with an increase in spontaneous calcium release events, resulting in delayed depolarization (DAD), triggered beats, and CPVT (Knollmann et al., 2006). CPVT is a disease caused by abnormal calcium handling leading to increased spontaneous calcium release that is pathological (Knollmann et al., 2006; Cerrone et al., 2009). To examine the effect of Casq2 deletion on RyR2 function and intracellular calcium release, we performed intracellular calcium spark measurements in permeabilized hiPSC-CMs using previously established protocols (Parikh et al., 2017).
These data are congruent with previous findings in mice ( Knollmann et al., 2006 ) and suggest that loss of Casq2 increases RyR2 calcium release. Variants in CASQ2 have been shown to cause a lethal arrhythmia syndrome, catecholaminergic polymorphic ventricular tachycardia (CPVT) (Lahat et al., 2001a; Lahat et al., 2001b). This suggests that the functional capacity of the SR to store calcium remains unchanged; a result similar to the observations in mouse cardiomyocytes lacking Casq2 (Knollmann et al., 2006).
Importantly, our findings in the heterozygous Casq2 model confirmed our previous reports that subtle reductions in Casq2 protein increase calcium leakage at the cellular level (Chopra et al., 2007).

While the mechanism of CPVT is generally agreed upon, the anatomical origin of the disease remains unclear. A better understanding of the mechanism of CPVT and the anatomical origin of the disease is important for developing treatments for CPVT patients. A limitation of most iPSC work is that all current lines have been generated directly from families with specific CPVT mutations (Itzhaki et al., 2012).
K180R was recently studied in vitro using the latest crystal structure of the Casq2 polymer (Titus et al., 2020). As previously discussed, one of the more commonly debated points regrading CPVT is the cellular origin of the ventricular arrhythmias that occur in the disease. However, there are also data generated to support that the origin of CPVT is outside the Purkinje cells.
One of the best tools to address these issues is human induced pluripotent stem cells (hiPSC). In the mouse model, the absence of Casq2 was shown to reduce junctin and triadin levels, most likely a consequence of the proteins interacting with each other (Knollmann et al., 2006). The aim of the experiment was address localization and protein interaction between the calcium processing proteins in the SR.
Previous work has shown that in mouse models, changes in calcium handling proteins within the SR typically decrease the levels of other proteins located within the calcium release unit (Knollmann et al., 2006; Chopra et al., 2009). . A quantitative structural and morphometric analysis of the Purkinje network and Purkinje-myocardial junctions in porcine hearts. Autosomal recessive catecholamine or exercise-induced polymorphic ventricular tachycardia: clinical features and assignment of the disease gene to chromosome 1p13-21.
Corrigendum: Antiarrhythmic Effects of Dantrolene in Patients with Catecholaminergic Polymorphic Ventricular Tachycardia and Replication of the Responses Using iPSC Models. Characterization of the first induced pluripotent stem cell line generated from a patient with autosomal dominant. Therapeutic approach for patients with catecholaminergic polymorphic ventricular tachycardia: state of the art and future developments.