Von
JACK FRAZIER, DIETER GRAMENTZ und UWE FRITZ
Monotypische Meeresschildkroten-Gattung und einziger rezenter Vertreter der Familie Dermochelyidae. Fur eine Gattungsdiagnose sei auf die einzige Art, Dermochelys coriacea, verwiesen.
Dermochelys coriacea (Vandellius, 1761) - Lederschildkrote
Diagnose. Grofite rezente Meeresschildkrotenart. Die SCL von 25 betragt meist 124 bis 176 cm, die maximale SCL liegt bei 252 cm (GlRONDOT und FRETEY 1996). ERNST et al. (2000) geben sogar eine maximale CL von 291 cm an, wobei unklar ist, ob sich dies auf cfcf oder 22 bezieht. Adulte Lederschildkroten wie- gen meist 300 bis 600 kg; der Rekord liegt bei 916 kg (ECKERT und LUGINBUHL 1988, MORGAN 1989).
Der Kopf ist grofi; die Rhamphotecae am Ober- und Unterkiefer sind diinn und schwach. Der Oberkiefer ist mittig tief eingekerbt, die Mediankerbe wird links und rechts durch eine deutliche Zacke begrenzt, auf die hinten wieder eine Kerbe im Oberkiefer folgt, so dafi an der Schnauze der Rand des Oberkiefers
„2K-formig ist; die Schnauze ist stumpf. Bin sekundarer Gaumen ist nicht ausge- bildet; die beiden Choanen sind daher im Unterschied zu Cheloniiden in der vorderen Oberkieferhalfte voll sichtbar. Der Schlund ist ausgesprochen volumi- nos und wie das Gaumendach und der Mundboden mit deudichen hakenartigen Papillen bedeckt. Der von oben leierfbrmige Carapax tragt sieben Langskiele, wovon einer auf der Korperlangsachse und jeweils drei rechts und links davon verlaufen. Der jeweils aufierste Kiel fallt mit der Carapax-Aufienkante zusam- men. Das Plastron weist drei schwache Langskiel-Paare auf. Bei adulten Leder-
1 Das zugrunde liegende englische Manuskript wurde von JACK FRAZIER 1985 geschrieben und an WOLFGANG BOHME geschickt. Spater wurde der Text von DIETER GRAMENTZ ins Deutsche iibersetzt und bearbeitet. Etwa 20 Jahre nach Abschlul? des originalen Manuskripts wurde der Inhalt von UWE FRITZ grundlegend neu bearbeitet und in die vorliegende Form gebracht.
250 8. Dermochelyidae - Lederschildkroten
schildkroten sind auf dem Panzer kerne Homschilder vorhanden. Die Panzer- oberflache wird von glatter Haut mit wachsartiger Oberflache bedeckt, unter der eine Schicht kleiner polygonaler Mosaikknochen liegt. Die Vorderflossen sind sehr lang; die Hinterflossen auffallig breit; Krallen fehlen (bis auf einige Ausnah- men bei Embryonen und Schliipflingen). Die Grundfarbe ausgewachsener Tiere ist dorsal dunkelbraun bis schwarz, worauf weifie Tupfen liegen; die Kopfoberseite tragt aufierdem haufig einen rosafarbenen Fleck; die Unterseite der Schildkroten ist meist hell gefarbt. Schlupflinge sind uberwiegend dunkelgrau bis schwarz gefarbt, die Rander des Carapax, der Flossen und die Carapax-Langskiele sind weifilich.
Beschreibung. Allgemeine Beschreibungen, Abbildungen und Fotografien von Dermochelys coriacea finden sich u. a. bei DERANIYAGALA (1939), CARR (1952),
LOVERIDGE und WILLIAMS (1957), VILLIERS (1958), PRITCHARD (1971, 1979, 1980), BRONGERSMA (1972), ERNST und BARBOUR (1972, 1989), REBEL (1974),
SMITH und SMITH (1980), PRITCHARD und TREBBAU (1984), MARQUEZ (1990) und ERNST et al. (1994, 2000).
Mafie: Nistende 99 haben meist eine SCL von 124 bis 176 cm, wobei die Durchschnittswerte verschiedener Populationen zwischen 134 und 162 cm lie- gen. Die SCL von 1.328 99 aus Franzosisch Guayana lag bei durchschnittlich 155 cm (127-252 cm, SO = 9), die SCB lag bei durchschnittlich 87 cm (67-199
100 110 120 130 140 150 160 170 180
Carapaxlange (cm)
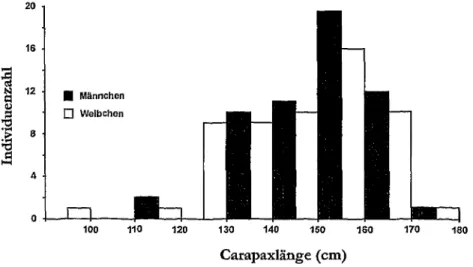
Abb. 45: Carapaxlangen von Dermochelys coriacea von der franzosischen Atlantikkiiste (n = 114); nach DUGUY (1997).
cm, SD = 6; GlRONDOT und FRETEY 1996). Die BGL variiert im allgemeinen zwischen 125 und 185 cm; die Durchschnittswerte bei verschiedenen Brut- kolonien liegen hier zwischen 143 und 164 cm. Das Gewicht adulter 22 liegt in der Regel zwischen 224 und 646 kg (DERANIYAGALA 1939, PRITCHARD 1969, HUGHES 1974a, 1996, BRONGERSMA1972, FRETEY 1978, BUSKIRKund CHOWDER 1994, LESLIE et al. 1996, STEYERMARK et al. 1996). Adulte cfcf ahneln hinsicht- lich der Panzermafie den 92 (Abb. 45). Bin in Wales gestrandetes cf soil eine BCL von 256,5 cm gehabt haben (ECKERT und LUGINBUHL 1988), womit die- ses Tier das grofite bekannte cf ware. MORGAN (1989) und DAVENPORT et al.
(1990a) geben fur dieses Tier allerdings eine korrigierte BCL von nur 159 cm an.
Die durchschnittliche BCL von 20 in Schottland, Wales und England gestrande- ten Lederschildkroten betrug 152 ± 25 cm (Spanne: 120-210 cm; GODLEY et al.
1998). Die Carapaxlangen von 114 Dermochelys coriacea von der franzosischen Atlantikkuste und von 84 Exemplaren aus dem Mittelmeer sind Abbildung 45 und 46 zu entnehmen. Aus dem Mittelmeergebiet sind nur extrem wenige Fun- de kleinerer Tiere bekannt (siehe „Fortpflanzung im Mittelmeergebiet", S. 277).
Die Spannweite der Vorderflossen soil bei Lederschildkroten maximal etwa 270 cm betragen konnen (PRITCHARD 1979). Die Breite von Spuren am Niststrand liegt meist zwischen 180 und 210 cm (PRITCHARD 1979).
Obwohl fur nistende Lederschildkroten-22 zahlreiche MeSwerte veroffendicht wurden, gibt es nur wenige Untersuchungen, die sich mit Proportionsverhaltnissen beschaftigen. Bei 22 sind Carapaxlange und -breite positiv korreliert, wie HUGHES (1974a) anhand von Bandmessungen und FRETEY (1978) anhand von Stockmafi-
20
l
N 16B. 12 S
i
a 8-4-
I I HI i
• CL n = 39 P BCL n = 44II
111-120 121-130 131-140 141-150 151-160 161-170 171-180 181-190
Carapaxlange (cm)
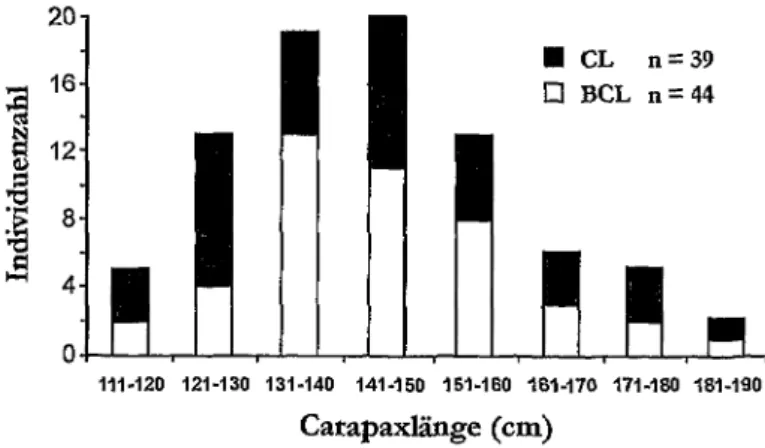
Abb. 46: Carapaxlangen von Dermochelys coriacea aus dem Mittelmeer. BCL: Bandmafi- Carapaxlange, CL: Carapaxlange ohne Angabe, ob es sich um Band- oder StockmafJ
handelt; nach CASALE et al. (2003).
252 8. Dermochelyidae - Lederschildkroten
Angaben zeigten. BENABIB (1983) konnte diesen Zusammenhang mit beiden Mefiverfahren an denselben Exemplaren bestatigen. Kopfbreite und Carapaxlange sind ebenfalls positiv korreliert (HUGHES 1974a). Bin grofies, haufig iibersehe- nes Problem ist, dafi verschiedene Forscher unterschiedliche Mefimethoden an- wenderij so dafi die Daten mitunter nur bedingt vergleichbar sind. Nach HUGHES (1971) unterscheiden sich Bandmafi- und StockmaiKangaben der Carapaxlangen um bis zu 6 °/o, bei der Carapaxbreite sogar um bis zu 32 %!
Au£ere Merkmale:SowohlderwissenschaftlicheNameDermocbelyscoriacea
= Jedrige Hautschildkrote" als auch der deutsche und englische Trivialname
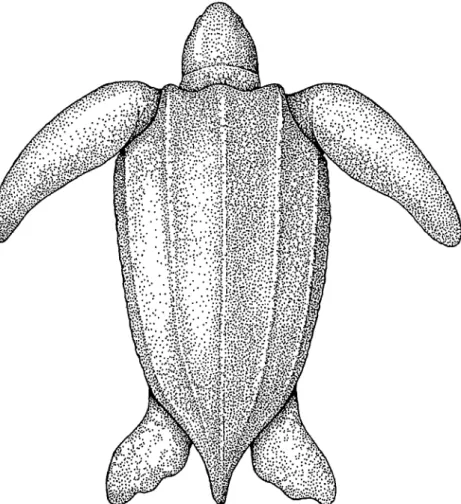
Abb. 47a: Adulte Dermocbelys coriacea in Dorsalansicht; Original J. MOBAVEC.
„Lederschildkr6te" bzw. „Leatherback Turtle" beziehen sich auf die wohl auftal- ligste externmorphologische Eigenschaft der Art, namlich, dafl der Panzer von glatter Haut start mit Hornschildern bedeckt wird (Abb. 47). Allerdings stimmt es nicht, dafi bei D. coriacea gar keine verhornten Hautstrukturen vorkommen, wie es manchmal zu lesen steht (PR1TCHARD 1979, PR1TCHARD und TREBBAU 1984). Auch die Angabe, dafS der Art Krallen und am Kiefer Hornschneiden vollig fehlen (z. B. bei CARR 1952, PRITCHARD 1971, 1979, 1980, ERNST und BARBOUR1972, PRITCHARD und TREBBAU 1984), ist falsch. Tatsachlich besitzen Lederschildkroten namlich an den Kiefern Hornschneiden, die allerdings recht diinn sind (DERANIYAGALA 1930, 1932, 1939, RAINEY 1981, FRAZIER 1987).
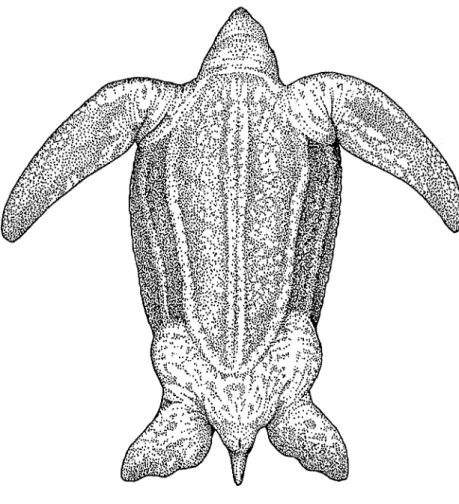
Abb. 47b: Adulte Dermochelys coriacea in Ventralansicht; Original J. MORAVEC.
254 8. Dennochelyidae - Lederschildkroten
Embryonen und teilweise auch Schliipflinge weisen nodi Krallen auf, und bei adulten D. coriacea kommen haufig auf den Augenlidern, am Hals und auf dem Kamm auf der Sclrwanzoberseite rudimentare Schuppen vor (DERANIYAGALA
1930,1932, 1934, 1936, 1939, 1953, BRAZIER 1987).
Der Kopf von Lederschildkroten (Abb. 48) ist in Relation zum Panzer grofi.
Die Hornschneiden des Oberkiefers sind in der Mitte stark eingekerbt, wobei diese Kerbe beiderseits von je einer auffalligen, nach hinten gerichteten Zacke eingefafit wird, auf die weiter hinten wiederum eine deutliche Kerbe im Oberkie- fer folgt. BRONGERSMA (1970) und RAINEY (1981) wiesen nach, dafi diese Zak- ken bei Schlupflingen in einem spitzen Dorn enden. Der Vorderrand des Unter- kiefers lauft mittig in eine hakenschnabelartige Spitze aus, die hochgezogen und etwas nach hinten gebogen ist. Diese Spitze pafit im Oberkiefer exakt in eine Vertiefung vor den Choanen. Auf den beiden Oberkieferasten liegt innen paral- lel zu den Hornschneiden ein leichter, aber deutlich erkennbarer Kamm. Zwi- schen diesem Kamm und der Hornschneide befindet sich eine flache Mulde, die die Schneidekanten des Unterkiefers aufnimmt, wenn das Maul geschlossen ist (DERANIYAGALA 1932; siehe die Abbildung bei BRONGERSMA 1970). Der Kopf ist vom Hals nur undeudich abgesetzt. Die Augenlidschlitze verlaufen beinahe
Abb. 48t Kopf einer Dermocbelys coriacea mit geoffhetem Maul. Man beachte die zahlrei- chenhakenformigen Papillen am Gaumendach und Mundboden; aus SCHUMACHER (1973).
vertikal, und die Nasenlocher sind etwas nach oben verschoben. Ein aufieres Tympanum fehlt. Kopf und Hals sind nur wenig beweglich.
Aufdem von oben gesehen leierfbrmigen Carapax befinden sich sieben longi- tudinale Kiele oder Leisten, die unregelmafiig fein gezahnt sind (BRONGERSMA 1969). Der Mittelkiel ist am hochsten und verlauft auf der Langsachse des Kor- pers; seitlich vom Mittelkiel liegen links und rechts achsensymmetrisch jeweils drei weitere Kiele. Zwischen den Kielen bildet die Panzeroberflache leicht ein- gesenkte „Tr6ge". Der Mediankiel ist bei manchen Tieren aufdem hinteren Panzer- drittel teilweise in einzelne erhabene Dornen aufgelost. An dem iiber dem Schwanz zipfelfbrmig ausgezogenen Carapaxhinterrand weisen Lederschildkroten haufig Verletzungen bzw. Narben auf (BRONGERSMA 1969,1972, FRETEY 1982, PRITCHARD und TREBBAU 1984, ERNST et al. 1994, 2000; siehe auch den Ab- schnitt „Mortalitat und Gefahrdung", S. 292). Die Panzerober- und Unterseite wird durch das am weitesten seitlich gelegene Kielpaar, die sogenannten Marginal- kiele, begrenzt. Das Plastron weist drei schwache Langskiel-Paare auf, wobei zwei Kiele quasi die Aufienkanten des Bauchpanzers darstellen, zwei Kiele auf der Briicke liegen und zwei weitere Kiele langs der Medianlinie so eng zusammen- rucken, dafi sie manchmal fur nur einen Kiel gehalten werden. Insgesamt ist das Plastron recht flexibel (BURNE 1905, DERANIYAGALA 1930,1939, BRONGERSMA 1969, 1970, PRITCHARD und TREBBAU 1984, ERNST et al. 1994, 2000).
Die Vorderflossen sind aufierordendich lang und breit. Die Hinterextremitaten sind durch eine dicke Membran (Patagium=Crurocaudalfalte) mit dem Schwanz verbunden. Die breiten, paddelahnlichen Hinterflossen sind nach hinten gerich- tet. Auf dem auffallig seitlich komprimierten Schwanz befindet sich auf der Oberseite ein hautiger Kamm. Die Kloake liegt bei beiden Geschlechtern weit vom Plastronhinterrand entfernt. Moglicherweise dient die Crurocaudalfalte der Steuerung und Balance (DERANIYAGALA 1939).
Auf dem Carapax liegen bei adulten Tieren auf jeder Korperseite zwischen den Marginal- und den Supramarginalkielen drei oder vier Poren, die bereits bei Schlupflingen vorhanden sind. Bei kleinen Jungtieren ist jede Pore von vier oder funfSchuppen umgeben. Ausgewachsene Lederschildkroten weisen statt dessen um die Poren nur vier oder funf leichte Vertiefungen auf, die strahlenformig von jeder Offhung wegziehen (DERANIYAGALA 1932,1936,1939, BRONGERSMA 1970).
Diese Poren konnten vielleicht die Exkretionsofrhungen der Rathkeschen Dru- sen darstellen (RAINEY 1981); Rathkesche Drusen wurden allerdings bislang von Dermochelys coriacea noch nicht nachgewiesen (WYNEKEN 2001).
Die Korperoberflache ist glatt, zah, lederartig und wenig flexibel. Die Haut setzt sich aus einem ziemlich lockeren, faserigen Gewebe zusammen und ent- halt kein Knorpelmaterial (DUNLAP 1955). Der Ruckenpanzer wird durch eine Art Schwarte gebildet, die aus bis zu 3,5 bis 4,0 cm dickem, kraftigen Bindegewe- be besteht, in die ein Mosaik von kleinen polygonalen Hautknochen (Osteodermen) eingelagert ist. Diese Osteodermen bilden eine nur knapp 5 mm
256 8. Dermochelyidae - Lederschildkroten
starke Schicht (DERANIYAGALA1932,1936,1939,1953). Das Plastron von Leder- schildkroten ist gelegentlich aufgrund einer starken Durchblutung rosa gefarbt (PRITCHARD 1979, PRITCHARD und TREBBAU 1984). In diesem Zusammenhang vermutet PRITCHARD (1979), dafi D. coriacea einen Teil der Atmung iiber die hautbedeckte Panzeroberflache decken kann, ahnlich wie Weichschildkroten (Trionychidae). Bislang ist diese Vermutung aber unbestatigt geblieben.
Adulte Lederschildkroten sind auf dem Carapax matt- oder grauschwarz ge- farbt, mit unterbrochenen weifien Linien auf den Kielen; besonders zwischen den Kielen befinden sich weifie Tupfen, die haufig in drei oder vier Langsreihen angeordnet sind. Der Kopf weist grofie weifie Flecken auf, von denen einige bis zum Kiefer reichen. Auf der Halsoberseite konnen die Flecken in bis zu runf Langsreihen angeordnet sein. Die Flossenansatze zeigen viele weifie Flecken.
Der Hautkamm auf der Schwanzoberseite tragt eine weifie Langslinie. Die Un- terseite von Lederschildkroten ist iiberwiegend hell gefarbt, insbesondere an den Kielen. Von der Inguinalregion bis zur Kloake kann sich ein schwarzes Band erstrecken.
Ein lebender, frischgeschliipfter Lederschildkroten-Albino wurde in Trinidad gefunden (GODLEY et al. 1993).
Weitere, detaillierte Angaben zur Farbung von D. coriacea finden sich bei DERANIYAGALA (1930,1932,1936,1939) und PRITCHARD und TREBBAU (1984).
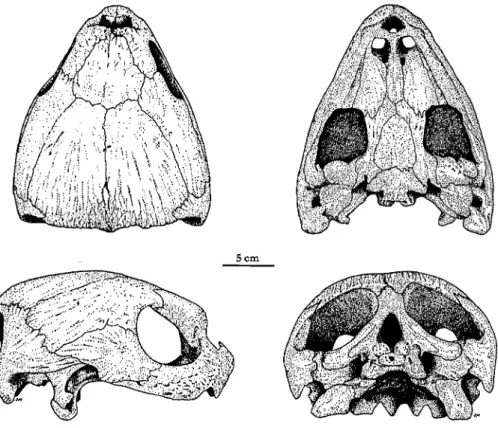
Schadel: Zum Schadel von Dermochelys coriacea (Abb. 49) haben NlCK (1913), WEGENER (1959) und GAFFNEY (1979) bedeutende Arbeiten vorgelegt; weitere Angaben finden sich bei DERANIYAGALA (1939) und im Kapitel liber Dermochelyidae von BEVER und JOYCE (2005) in diesem Band. Der Schadel von D. coriacea gehort aufgrund seiner zahlreichen Besonderheiten zu den am besten untersuchten Schildkrotenschadeln iiberhaupt. Im Gegensatz zu den weitaus meisten anderen Schildkrotenarten sind viele Schadelelemente reduziert oder paedomorph. Trotz der beeindruckenden Schadelgrofie sind die Knochen diinn und nur schwach miteinander verbunden. Es gibt keine starke temporale Emargination, wie sie bei den meisten rezenten Schildkroten vorkommt, und der Fortsatz des Supraoccipitales ist dorsal fast vollstandig vom Schadeldach bedeckt. Der Oberkiefer ist mittig eingekerbt. Die Kerbe wird links und rechts durch je eine Zacke flankiert, die hinten wiederum eingekerbt ist. Sowohl Praemaxillare als auch Maxillare sind an der Bildung der Zacken beteiligt.
Das Frontale reicht nicht bis zum Rand der Augenhohle. Das Postorbitale ist sehr grofi und bedeckt den grofiten Teil des Schadels in der Temporalregion. Der medialgerichtete Fortsatz des Jugale ist reduziert und steht weder mit Palatinum noch Pterygoid in Verbindung. Die Fortsatze des Palatinums sind aufierordent- lich schmal. Das Palatinum reicht lateral zum Labialrand des Maxillares. Es ist kein sekundarer Gaumen ausgebildet. Die Crista supraoccipitalis, die die An- satzstelle fur Sehnen und den Musculus adductor mandibulae externus darstellt,
5 cm
Abb. 49: Oberschadel einer adulten Dermochelys coriacea (2MB 64820, portugiesische Atlantikkuste). Oben links Dorsal-, rechts Ventralansicht Unten links Lateral-, rechts
Occipitalansicht; Original J. MORAVEC.
ist relativ klein. Ebenso wie bei den Cheloniiden fehlt auch D. coriacea das Foramen palatinum posterius. Die Incisura columellae auris, die den Ohrknochen ent- halt, ist relativ weit ofFen. Maxillare und Pterygoid beriihren sich nicht. Das Vomer erreicht die Praemaxillaria und trennt so die Palatina. Das Basioccipitale liegt dorsal zwischen den Exoccipitalen, die dem Condylus occipitalis in der Lange entsprechen (GAFFNEY 1979). Der Processus trochlearis oticum des Prooticum ist stark reduziert. Bin Parasphenoid ist vorhanden (VERSLUYS 1907, 1909). Das Squamosum reicht nicht bis zum Processus paroccipitalis des Opistho- ticums (GAFFNEY 1979). Die Art ist die einzige Cryptodire, die kein verknocher- tes Epipterygoid besitzt (NICK 1913, GAFFNEY 1975,1979). Weder das Prooticum noch das Pterygoid beriihren den rudimentaren Processus inferior parietalis, auch das Pterygoid und dem Prooticum beriihren sich nicht (GAFFNEY 1979).
Der Hirnschadel besitzt stark reduzierte Knochenwande und ist sekundar mit Knorpelgewebe umgeben (NICK 1913). Der nasale Knorpel, eine Verlangerung
258 8. Dermochelyidae - Lederschildkroten
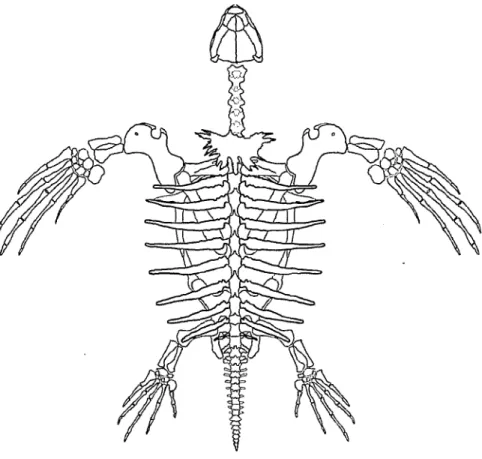
Abb. 50: Skelett einer adulten Dermochelys coriacea. Links Dorsal-, rechts Ventralansicht.
Epithecaler Panzer, Unterkiefer und Zungenbein sind weggelassen. Man beachte das ein- zige thecale Carapaxelement (Proneurale, grau); Original I. DANILOV, kombiniert nach Abbildungen in GERVAIS (1872). Im vorliegenden Band wird dasselbe Skelett mit
epithecalem Panzer auf S. 240-241 gezeigt.
der Nasenscheidewand, entwickelt sich bereits beim Embryo (PEHRSON 1945).
Der Condylus occipitalis bleibt zeitlebens knorpelig (HAY 1908).
Am Auge sind im allgemeinen 14 Sclera vorhanden, manchmal kann ihre Zahl aber auch auf 7 reduziert sein, wenn eine Liicke im anterodorsalen Teil des Ringes vorhanden ist. Normalerweise sind an beiden Augen gleich viele Sclera vorhanden. Einzelne Sclera konnen verbreitert sein, wodurch Liicken im Ring geschlossen werden, und nebeneinander liegende Sclera konnen sich xiberlap- pen oder sogar miteinander verschmelzen (DERANIYAGAIA 1932,1939).
Der Unterkiefer von Lederschildkroten ist ebenfalls sehr ungewohnlich. Das Dentale beriihrt nur das Surangulare und das Angulare und nicht funf verschie- dene Knochen3 wie es normalerweise der Fall ist. Auf dem Dentale ist nur ein
Labialkamm vorhanden, ein Linguinalkamm fehlt (GAFFNEY 1979). Auf der la- teralen Seite des Dentales findet sich fur den Ansatz des Musculus adductor mandibulae externus keine Vertiefung. Das Coronoid fehlt. Das Articulare ist nicht ossifiziert. Das Praearticulare beriihrt keine anderen Knochen.
Postcranialskelett: Obwohl der Panzer stark reduziert ist, bilden Carapax und Plastron etwa ein Drittel der Korpermasse. Ebenfalls ein Drittel wird von den Gliedmafien beansprucht, wobei die Vorderextremitaten doppelt so viel wiegen wie die hinteren.
Das Skelett einer Lederschildkrote zeigt Abbildung 50. Es sind acht Cervical- wirbel und 10 Dorsalwirbel vorhanden. Daran schliefien sich zwei Sacralwirbel und 18 Caudalwirbel an, die allerdings auch als drei Sacral- und 17 Caudalwirbel aufgefafJt werden konnen. Bis auf die letzte Dorsalrippe artikulieren alle ande- ren an einem Neuralbogen und zwei angrenzenden Wirbelzentren. Die Dorsal- rippen sind in eine schwartenartige Knorpelmasse eingebettet (GERVAIS 1872, VOLKER 1913, DERANIYAGALA 1939).
260 8. Dermochelyidae - Lederschildkroten
Die Langknochen von Dermochelys coriacea unterscheiden sich von alien ande- ren rezenten Reptilien, da ihre Epiphysen zeidebens knorpelig und unverknochert bleiben. Die Knochen werden von zahlreichen, auffalligen perichondralen und transphysealen Gefafien versorgt. Diese Verhaltnisse ahneln Meeressaugern und weisen aufein schnelles Wachstum und einen raschen Stoffwechsel hin (RHODIN et al. 1981, RHODIN 1985).
Der Hals der Lederschildkrote ist kurz; der Kopf kann nicht in den Panzer eingezogen werden. Obgleich einige Wirbel durch dicke Knorpelpolster und faseriges Gewebe miteinander verbunden sind, zeigen sie Artikulationen, wie sie fur Cryptodiren charakteristisch sind (VOLKER1913, VERSLUYS 1913). Der vier- te Halswirbel ist bikonvex, die davor gelegenen Wirbel sind opisthocoel, die dahinter gelegenen procoel. Die Verbindung zwischen dem sechsten und sieb- ten Cervicalwirbel ist oft rest und manchmal massiv verschmolzen. Die Artiku- lation zwischen dem siebten und achten Cervicalwirbel ist ziemlich variabel und gelegentlich bikonvex (WILLIAMS 1950).
Von den zehn Dorsalrippen ist das erste Paar sehr kurz und das letzte nur rudimentar ausgepragt. Die anderen haben dunne proximale und distale Enden und sind mittig am dicksten. Verglichen mit den Pleuralknochen anderer Schild- kroten sind die Rippen der Art sehr schmal und schwach. HAY (1898, 1908) nahm an, da6 ihre abgeflachte Form mit den rauhen Kanten darauf hindeutet, dafi sie einst mit Pleuralplatten verschmolzen waren. Die Caudalwirbel sind procoel und besitzen keine Haemalbogen (DERANIYAGALA 1939).
Der Humerus zeigt mehrere Besonderheiten. Anders als bei Cheloniiden bleibt das Ectepicondylar-Foramen wahrend des ganzen Lebens geschlossen und ist nicht zu einer Rinne umgebildet. Der Deltopectoralkamm reicht weit seitwarts und ist auf der ventralen Seite des Scharts mit einer transversalen Reihe von Muskelansatzpunkten versehen (vgl. Abb. 2 auf S. 18). Bei der Hinterextremitat sind Femur, Tibia und Fibula dorsoventral ein wenig abgeflacht und relativ kurz (ROMER 1956). Die Phalangen sind lang und Condylen fehlen. Der Carpus hat nur ein Centrale, obgleich ein zweites Centrale als Rudiment bei jungen Indivi- duen vorkommen kann (VERSLUYS 1913, VOLKER 1913).
Die Elemente des Schultergiirtels sind kraftig und besitzen ein massives Coracoid. Die starksten Knochen sind die Scapulae und die ziemlich kurzen, aber breiten Humeri. Am Beckengurtel fehlt das Thyroid-Fenster. Start dessen ist ein Paar kleiner Foramina vorhanden. Die puboischiadische Platte bleibt weit- gehend verknorpelt und verknochert nicht. Das Epipubis besitzt ein mediales Fenster (VOLKER 1913).
Auf die Besonderheiten des Panzers von Dermochelys coriacea wurde im vorlie- genden Band bereits von BEVER und JOYCE (2005) ausfuhrlich eingegangen, so dafi hierauffur Naheres verwiesen werden kann (siehe S. 235-248). Der thecale Ruckenpanzer ist extrem reduziert. Es ist nur noch ein schmetterlingsformiges Proneurale erhalten, das von der Panzeraufienschicht durch Bindegewebe ge-
trennt ist. Das Proneurale ist mit dem Neuralbogen des achten Halswirbels ver- bunden (VERSLUYS 1913).
Die thecalen Elemente des Plastrons sind ebenfalls stark reduziert. Anstelle der iiblichen festen Knochenplatten ist nur noch ein diinner Rahmen entlang der Plastron-Aufienkanten vorhanden. Dieser Rahmen besteht aus acht spangen- artigen Knochenelementen. Das Entoplastron fehlt. Es ist lediglich bei manchen Embryonen als knorpeliges Rudiment angelegt (DERANIYAGALA 1939).
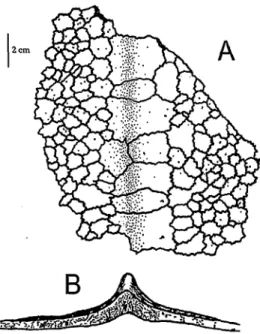
Wahrend auf dem Carapax groSflachig epithecale Knochenelemente vorhan- den sind, die aus einem Mosaik von Tausenden von Ossikeln bestehen, sind epithecale Knochen auf dem Plastron vor allem an den sechs Langskielen kon- zentriert und auf diese beschrankt. Nach aufien werden die Osteodermata auf dem Carapax haufig grofier, dafiir nimmt ihre Zahl ab. Audi unter den Carapax- kielen sind die Ossikeln grower als dazwischen. Die Ossikel unter den Kielen sind dachformig aufgeworfen und tragen so zur Bildung der Kiele bei (PRITCHARD und TREBBAU 1984, WOOD et al. 1996; Abb. 51). BRONGERSMA (1969) stellte bei zwei von drei untersuchten Exemplaren (zwei adulte cfcf und eine subadulte Schildkrote unbekannten Geschlechts) fest, daf? die Osteodermen am Plastron
Abb. 51: Teil des epithecalen Panzers einer Dermochelys coriacea in Ventralansicht (A). Die grofien Ossikeln liegen unter einem der Panzer-Langs kiele. (B) Querschnitt durch einen Teil des epithecalen Panzers an einem Langskiel. Umgezeichnet nach Fotografien in WOOD
et al. (1996); Original U. FRITZ.
262 8. Dermochelyidae - Lederschildkroten
leicht abgescheuert waren. Bei alien Schildkroten waren Knochenplattchen her- ausgefallen.
Viscerale Anatomic:DasGehirnvonDmnochelyscoriacealiegtmitAus- nahme des Pinealorgans tief im Schadel verborgen. Das Pinealorgan befindet sich direkt unter dem charakteristischen rosafarbenen Fleck auf der Kopfoberseite.
Wie bei Cheloniiden ist das Gehirn langgestreckt. Fur Abbildungen und einen Vergleich mit Cheloniiden siehe WYNEKEN (2001). Die durch den fehlenden sekundaren Gaumen gut sichtbaren Choanen sind bei Lederschildkroten beson- ders grog und weit vorne gelegen (PARSONS 1968). Der seitliche Choanalrand tragt keine kleinen warzenartigen Erhebungen wie bei Cbelonia mydas (DERANIYAGALA 1939, PARSONS 1968, BRONGERSMA 1970, WYNEKEN 2001).
Am Gaumendach, dem Mundboden und im grofien Schlund fallen zahlreiche hakenfbrmige, verhornte Papillen auf (Abb. 48, S. 254), die eine Rolle beim Fressen und Festhalten der glitschigen Hauptnahrung spielen konnten. Bei Cheloniiden beschranken sich derartige Papillen auf den Oesophagus (SCHUMACHER 1973, WYNEKEN 2001). Die Speiserohre von D. coriacea ist weit- aus langer als bei Seeschildkroten und erstreckt sich iiber beinahe die Halfte der Korperlange und biegt dann links wieder nach anterior ab, wo er auf Achselhohe mit einer erneuten Linksbiegung schliefilich in den Magen mundet. Verglichen mit Seeschildkroten ist der Magen grofi und voluminos, der Darmtrakt dagegen auffallig kurz. Oberhalb und hinter dem Auge sind wie bei Cheloniiden Salzaus- scheidungsdriisen vorhanden, die allerdings wesentlich grofier als bei See- schildkroten sind (WYNEKEN 2001).
Zungenbeinapparat und Kehlkopf wurden von SlEBENROCK (1898,1899) de- tailliert untersucht, worauf hier verwiesen sei.
Das Herz ist fur eine Schildkrote ungewohnlich lang und schmal, da die Ven- trikel ein langes und dickes Gubernaculum cordis bilden. Die posteriore Halfte des Ventrikels ist praktisch ein kompakter Muskelkorper ohne jede Aushohlung.
Die Auricularwande sind relativ diinn (BURNE 1905). Weiter vorne ist der Ven- trikel „schwammartig" mit vielen Trabeculae. Die Arteria coronaria ist relativ eng, die Vena coronaria grofi. Die Herzanatomie der Lederschildkrote erlaubt eine weitgehende Trennung der Blutzirkulation in einen weitgehend separaten Herz- und Lungenkreislauf (DUNLAP 1955, WYNEKEN 2001).
Lederschildkroten verfugen iiber einige anatomische und physiologische Beson- derheiten, die vermutlich spezielle Anpassungen an das Tieftauchen darstellen. So ist das Atemvolumen von Lederschildkroten auffallig klein, woraus man auf relativ Heine Lungen schliefien kann. Allerdings weist das Blut einen extrem hohen Haematokrit und eine sehr hohe Haemoglobm-Konzentration auf, wodurch eine hohere Kapazitat fur den Sauerstoff-Transport erreicht wird als bei den See- schildkroten. Lederschildkroten speichern SauerstofFetwa zu gleichen Teilen in den Lungen, im Blut und im Gewebe. Im Pectoralmuskel ist doppelt soviel Myoglobin
vorhanden wie bei Cheloniiden. Dies deutet klar darauf hin, daft bei Leder- schildkroten eine andere Strategic zur SauerstofFspeicherung verwirklicht ist als bei Seeschildkroten. Wahrend bei Cheloniiden der Sauerstoffiiberwiegend in der Lun- ge vorgehalten wird, ist bei D. coriacea ein hochentwickelter Mechanismus vorhan- den, der erlaubt, eine grofiere Sauerstofrmenge zusatzlich im Blut und Gewebe zu speichern. Dadurch werden langere (und somit tiefere) Tauchgange ermoglicht. Die kleinen Lungen diirften auch dem mit der Tauchtiefe zunehmenden hydrostati- schen Druck besser standhalten als die relativ grofieren Lungen der Seescbildkroten.
Aufierdem verringert das relativ kleine Volumen der Lungen von D. coriacea den Auftrieb, wodurch das Abtauchen erleichtert wird (LUTCAVAGE et al. 1990, 1992, LUTCAVAGE und LUTZ 1997). Die Lungen von Lederschildkroten zeichnen sich im Vergleich zu Cheloniiden auch durch wesentlich dichter gepackte Lungenblaschen (WYNEKEN 2001) und somit eine groCere innere Oberflache aus.
Dermochelys coriacea besitzt aufierdem eine spezielle „Arterienweiche", die durch einen Schliefimuskel (Sphinkter) kontrolliert wird. Vermutlich kontrahiert dieser Sphinkter wahrend des Tauchens und schliefit so den Blutkreislauf unter Umge- hung der Lunge kurz (KOCH 1934, DUNLAP 1955, SAPSFORD 1978).
Fur die Extremitatenansatze der Lederschildkrote wurde ein Gegenstrom- Warmeaustauscher-Prinzip beschrieben (GREER et al. 1973). Hier gibt es dichte GefaEbiindel mit bis zu vier Venen pro Arterie, die wie ein Warmetauscher wir- ken. Dieses System findet sich bei Schlupflingen und Adulti (MROSOVSKY" 1980).
Moglicherweise bilden auch die Blutgefafie im Bereich der Nares einen analogen Warmeaustauscher (SAPSFORD und HUGHES 1978; siehe auch den Abschnitt
„Thermoregulation").
Beschreibungen der Anatomic der Muskulatur finden sich bei FURBRINGER (1874), BURNE (1905) und WYNEKEN (2001), des Verdauungssystems bei
VAILLANT (1896), BURNE (1905), DUNLAP (1955), RAINEY (1981), HARTOG und NIEROP (1984) und des Urogenitalsystems bei BURNE (1905), DUNLAP (1955) und WYNEKEN (2001). Bei der Muskulatur sind die ausgesprochen kraftig ent- wickelten Muskelstrange im Bereich des Schulter- und Beckengurtels zu erwah- nen, die im Zusammenhang mit dem hervorragenden und ausdauernden Schwimmvermogen von Lederschildkroten zu sehen sind.
Schlupflinge der Lederschildkrote besitzen in der Axillar- und Inguinalregion aufFallige linsenfbrmige Fettkorper, die gelblich-weifi gefarbt und verhaltnisma- fiig grofier sind als bei Cheloniiden (RAINEY 1981). Unter der Panzerepidermis hat D. coriacea zwei Fettschichten, die durch ein faseriges Septum voneinander getrennt werden. Die aufiere Schicht ist weifi gefarbt, von fester Konsistenz und unter dem Carapax und Plastron vorhanden. Sie ist etwa um ein Drittel dicker als die darunter liegende Schicht. Diese ist weicher, braun gefarbt und stark mit Blutgefafien durchzogen. Die untere Schicht erstreckt sich iiber fast den ganzen Carapax, besonders entlang der Wirbelsaule und auch iiber einen Teil des Plastrons. Nur zwischen den Fettzellen der aufieren Schicht tritt etwas dirrus
264 8. Dermochelyidae - Lederschildkroten
verstreutes Bindegewebe auf (GOFF und STENSON1988). Die innere braune Fett- schicht dient wohl ahnlich wie bei Saugetieren der Warmeproduktion (ROTHWELL und STOCK 1979); der aufSeren weifien Schicht wird neben Isolation auch eine strukturelle Stiitzfiinktion fur den Panzer zugesprochen (GOFF und STENSON 1988). Interessanterweise besitzt das Flossenfett einen niedrigeren Gefrierpunkt als das Fett aus dem Korperinneren von Lederschildkroten und diirfte so mit zu der Kaltetoleranz der Art beitragen (DAVENPORT et al. 1990b).
Auffallig sind bei D. coriacea auch die grofien Mengen von Iran, die im Kor- pergewebe, Knochen und Panzerschwarte vorhanden sind. Dieses Ol spielt si- cherlich eine Rolle bei der Thermoregulation und hat moglicherweise auch eine antibiotische Funktion. Vielleichtist dadurch zu erklaren, dafi Lederschildkroten nur sehr schwach von Epizoen besiedelt werden (BLEAKNEY 1965). Moglicher- weise kommt dem Korperfett auch eine wichtige Aufgabe beim Druckausgleich wahrend des Tauchens zu (PRITCHARD 1979).
Penis und Kloake: Der Penis ist relativ einfach gebaut. Auf der Glans befindet sich eine U-fbrmige Falte, die anscheinend eine vergrofierte Verlange- rung der Samenrinne ist. Die Samenrinne endet auf der Innenflache der Falte.
Diese Struktur ist emfacher als bei den meisten anderen Cryptodiren und ahnelt den Seeschildkroten (BURNE 1905).
Blutmerkmale: Dermochelys coriacea weist verglichen mit anderen Schild- kroten einen aufrallig hohen Haematokritwert auf, und das Blut zeichnet sich durch eine extrem hohe Haemoglobin-Konzentration aus (LUTCAVAGE et al.
1990,1992). Die Anzahl der Erythrocyten variiert zwischen 447.000 und 547.000 pro mm3 und betragt im Durchschnitt 503.000 pro mm3 (FRAIR 1977a, b). Der Gesamtproteinanteil im Blutserum der Art betragt 3,70 g/100 ml und der Albuminanteil 1,42 g/100 ml (COHEN 1954).
Immunopraezipitationstests mit Antiseren zeigen, dafi sich D. coriacea deut- lich von Cheloniiden unterscheidet, ihnen aber naher steht als anderen Schild- kroten (FRAIR 1979). Vergleichbare Resultate wurden durch Elektrophorese und Immunoelektrophorese erzielt (FRAIR 1982).
Karyotyp: 2n = 56 Chromosomen, wovon 14 meta- oder submetazentrisch, 10 telo- oder subtelozentrisch und 32 Mikrochromosomen sind. Geschlechts- chromosomen sind keine vorhanden. Der Karyotyp gleicht von der Chromoso- menzahl und -morphologie dem von Cheloniiden (MEDRANO et al. 1987).
Verbreitung. Dermochelys coriacea besitzt ein gewaltiges Verbreitungsgebiet, mit dem sich kaum eine andere Tierart und keine andere Reptilienart messen kann (Abb. 52). Bemerkenswert sind die weit nach Nord und Slid vorgeschobenen
%T\ JJ (H I ,25 <H \D
. &* 5 13 a c: N
a 3
! T3 12 -s
9.280- so |
i
a °
M-8
i a-a a
8^
S3.3 g <u
•S S '•{ ^ -o >-) .a
£
w1 * i
a a SP^ • uu w
m j "O w O aj ki
35 53 5 S I I lo
3
8ia%
a
266 8. Dermochelyidae - Lederschildkroten
Nachweise. So existieren zweifelsfreie Meldungen fur die Gewasser um die nor- wegischen Versteral-Inseln (69°18'N 16°10'O und 68°37'N 14°17'0;
BRONGERSMA 1972), also bereits nordlich des Polarkreises! 1992 wurde eine an- dere Lederschildkrote mit 330 kg Gewicht vor der Kiiste Norwegens unweit des Polarkreises gefangen (ca. 66°30'N; ANON. 1993), und ein weiterer Nachweis stammt aus den Gewassern vor Island (ca. 64°N; PETERSEN 1984). Eine Mel- dung fur die Barentssee von BANNIKOW et al. (1971) erscheint allerdings in einer uberarbeiteten Neuauflage desselben Werkes nicht mehr (BANNIKOW et al. 1977) und diirfte zweifelhaft sein, da die Lederschildkrote im Unterschied zu Caretta caretta auch von BRONGERSMA (1972, 1982) fur die Barentssee nicht genannt wird. Imwesdichen Atlantik dringenLederschildkroteniiber Sommer regel- mafiig nach Norden in den Golf von Maine und bis nach Neufundland (48°N) vor (u. a. BLEAKNEY 1965, BRONGERSMA 1972, LAZELL 1980, GOFF und LIEN 1988, SHOOPundKENNEY 1992). Es gibt aufierdem in derLabradorsee (56°45'N) einen Lederschildkroten-Nachweis (THRELFALL 1978) und Indizien fur ein gele- gentliches Vorkommen bis Baffin Island (SHOOP 1980). Im Suden erstreckt sich das Verbreitungsgebietbis vor Kapstadt (34CS; HUGHES 1974a). Aus Siidamerika sind zahlreiche Nachweise von weit im Suden gelegenen Fundorten bei Rio de la Plata und Mar del Plata (38°S) in Argentinien bekannt (FREIBERG 1945, FRAZIER
1984).
ImBereich des westlichenIndischen Ozeans und des Roten Mee- res gibt es Lederschildkroten-Nachweise vom nordlichen Roten Meer (28°N;
FRAZIER und SALAS 1984) bis zu den Gewassern vor der Kap-Provinz, Sudafrika (34°S; HUGHES 1974a). Aus Siidostasien sind viele Funde bekannt (POLUNIN 1975), genauso aus Siidwestaustralien und Tasmanien (LlMPUS und McLACHLAN 1979) sowie von Neuseeland. Hier liegen die siidlichsten Funde im Bereich der Foveaux-Strafie an der neuseelandischen Sudinsel (47°S; PRITCHARD 1982a).
Aus dem nordwesdichen Pazifikraum kennt man von der russischen und japanischen Kiiste Nachweise bis zum 44. nordlichen Breitengrad (NlSHIMURA 1964a, b). Der nordlichste Nachweis liegt bei Kap Nawarin im Beringmeer (etwa 62°N; BANNIKOW et al. 1971,1977). Funde aus dem ostpazifischen Raum reichen im Norden bis nach British Columbia (MACASKIE und FORRESTER 1962) und zum Golf von Alaska (61°N; HODGE 1979, STINSON 1984), im Suden gehen sie bis zur Isla Chiloe, Chile (ca. 43°S; PRITCHARD 1980, BROWN und BROWN 1982).
Die Hauptnistgebiete der Lederschildkrote liegen ausschlieGlich in den Tropen und konzentrieren sich besonders auf Mittel- und Siidamerika sowie Siidostasien (Abb. 52; siehe auch unter „Biotop", S. 284 und „Populations- dynamik", S. 291). Zusammenstellungen der Nistgebiete und -zeiten finden sich in MARQUEZ (1990) und ERNST et al. (1994,2000); eine tJbersicht iiber sporadi- sche Eiablagen und Nistversuche an der US-amerikanischen Atlantikkiiste ge- ben RABON et al. (2003).
Fruher nahm man an, dafi die Lederschildkrote an der europaischen Atlan- tikkiis te eherseltenist(z. B. MERTENSundMOLLER1928). SCHREIBER(1912) hielt Cornwall (Grofibritannien) fur das nordlichste Vorkommensgebiet in euro- paischen Gewassern. Die 188 von BRONGERSMA (1972) zusammengestellten, zweifelsfreien Lederschildkroten-Nachweise aus dem europaischen Atlantik, der Nord- und Ostsee belegen jedoch, daiS die Art besonders im Atlantik wesentlich haufiger vorkommt, als ursprunglich vermutet wurde und dafl die Lederschildkrote nach Norden bis in skandinavische Gewasser vordringt. Die Nachweise verteilen sich wie folgt auf die Hoheitsgewasser der einzelnen Lander (von West nach Ost und Nord nach Slid; BRONGERSMA 1972): Island - 1, Britische Inseln - 66, Spanien - 11, Portugal - 10, Frankreich - 83, Norwegen - 9, Schweden - 1, Niederlande - 3, Danemark - 2 und Deutschland - 2. Dabei ist die grofie Nach- weisdichte in der Bucht von Biskaya auffallig (siehe auch unten).
In die O s t s e e verirren sich Lederschildkroten nur ausgesprochen selten. Am 20.10.1965 wurde ein lebendes 2 mit einer GL von 215 cm im Strelasund 5 km nordosdich von Stralsund in einer Fischreuse gefangen (SCHUMACHER 1973). In der Nordsee werden Lederschildkroten etwas haufiger nachgewiesen. Unter den oben genannten Nachweisen von BRONGERSMA (1972) gibt es eine ganze Reihe von Meldungen, die sich besonders auf den nordlichen Teil der Nordsee und die britische Ostkuste beziehen. Auf das Wattenmeer und das Kattegat ent- fallen allerdings nur wenige Nachweise.
Im folgenden sind einige neuere Nachweise fur den europaischen Atlantik und die Nordsee zusammengestellt, wobei kein Anspruch auf Vollstandig- keit erhoben wird.
Spanien und Portugal: Fur die iberische Adantikkuste wird Dermochelys coriacea regelmafiig gemeldet. Die Nachweise verteilen sich iiber die gesamte Kuste, konzentrieren sich aber besonders an der Nordkiiste und in Galicien (LOPEZ-JURADO et al. 1997, LOPEZ und ANDREU 1997).
Auch fur die Strafie von Gibraltar existieren zahlreiche Lederschildkroten- Meldungen. Fur diejahre 1980 bis 1982 fuhren FERNANDEZ und MORENO (1984) z. B. 13 Nachweise an. Einer der bemerkenswertesten Berichte stammt von GARCIA und CHAMORRO (1984), die angeben, dafi im November und Dezember ganze Gruppen von Lederschildkroten beobachtet werden konnen. Auch DELAUGERRE
(1987) erwahnt Gruppen von zwei bis elf D. coriacea (insg. 24 Exemplare), die im Juli 1984 auf der mediterranen Seite der Strafie von Gibraltar gesichtet wurden.
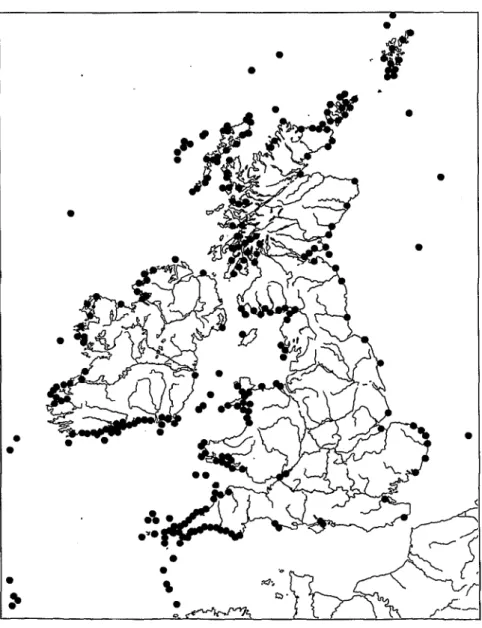
FurdieBritischen Inseln verdankenwireinemErfassungsprogrammftir Meeresschildkroten-Sichtungen und -Strandungen ein sehr exaktes Bild. Unter alien aus britischen und irischen Gewassern bekannten Meeresschildkrotenarten werden Lederschildkroten bei weitem am haufigsten nachgewiesen. Bis 2000 wur- den 461 eindeutige Sichtungen und Strandungen von Dermochelys coriacea regi-
268 8. Dermochelyidae - Lederschildkroten
striert, gefolgt von 94 Nachweisen fur Caretta caretta, 25 Nachweisen fur Lepidochelys kempii, vier Nachweisen von Chehnia my das und einem fur Eretmochelys imbricata. Die meisten Lederschildkroten-Nachweise stammen aus dem 20. Jahr- hundert, und hier wiederum - vermudich aufgrund einer zunehmend systema- tischen Erfassung - aus den letzten beiden Jahrzehnten (PlERPOINT und PENROSE
Abb. 53: Lederschildkroten-Nachweise um die Britischen Inseln; nach PlERPOINT und PENROSE (2002).
2002). Zwischen 1983 und 1993 wurden allein 167 D. coriacea nachgewiesen, wovon 67 auf England, 61 auf Schottland, 29 auf die Republik Hand, eine auf Nordirland und neun auf Wales entfallen. Zwischen 1993 und 2003 waren es sogar 349 Lederschildkroten (England: 145, Schottland: 78, Hand: 64, Wales:
62). An den West-, Sudwest- und Sudkiisten Hands und der britischen Hauptin- sel ist die Nachweisdichte besonders hoch (Abb. 53). Mit Ausnahme von 1998 (37) und 2002 (57) wurden zwischen 1993 und 2003 jahrlich zwischen 10 und 26 lebende Lederschildkroten gesichtet (PENROSE 2004). 2001 wurden neun Leder- schildkroten-Kadaver angespiilt (Mai: 1, August: 1, Oktober und November:
jeweils 3, Dezember: 1; PENROSE 2002), davon auch ein Exemplar an die irische Ostkuste. 2002 wurden vier (Februar: 2, Juni: 1, Oktober: 1; PENROSE 2003) und 2003 sechs tote D. coriacea gefunden (Januar: 1, August und September: jeweils 2, November: 1; PENROSE 2004).
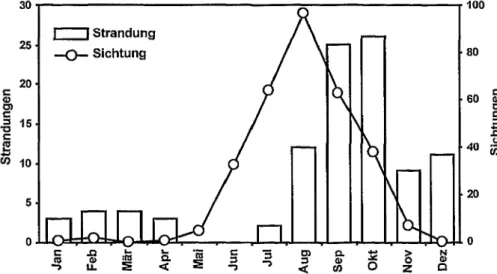
Lederschildkroten werden in britischen und irischen Gewassern ganzjahrig nach- gewiesen. Die meisten Tiere werden aber zwischen Juni und Oktober beobach- tet, mit einem klaren Nachweisgipfel im August. Interessanterweise stranden die meisten geschwachten oder toten Tiere etwas spater, namlich im September und Oktober (Abb. 54). Die jahreszeidich fnihesten Nachweise erfolgen meist an der irischen Slid- und der englischen Siidwestkuste, die relativ seltenen Nachweise an der englischen Nordseekiiste wesentlich spater, namlich im Winter oder zeiti- gen Fruhjahr. Haufig wird das Auftreten von Lederschildkroten in einen Zusam- menhang mit Quallenvorkommen gebracht (PlERPOINT und PENROSE 2002).
100
Abb. 54: Jahreszeidiche Verteilung von Lederschildkroten-Strandungen und -Sichtun- gen fur die Britischen Inseln; nach PlERPOINT und PENROSE (2002).
270 8. Dermochelyidae - Lederschildkroten
Frankreich: Die Lederschildkrote wird an der Atlantikkuste ausgesprochen haufig, im Armelkanal erheblich seltener nachgewiesen. Zwischen 1979 und 2003 wurden hier insgesamt 1.366 Lederschildkroten registriert (Tab. 27). Die auffalli- gen Haufigkeitsunterschiede hangen offenbar teilweise mit der Wassertempera- tur und der Haufigkeit von Quallen zusammen. In Jahren mit erhohter Wasser- temperatur und reichen Quallenvorkommen sind Lederschildkroten wesentlich haufiger als sonst (DUGUY 1996). Der enorme Anstieg im Jahr 1997 geht aller- dings wohl eher auf ein spezielles Erfassungsprogramm zuriick (DUGUY et al.
1998).
Tabelle 27. Nachweise von Dermocbelys coriacea an der franzosischen Atlantikkuste und im Armelkanal in den Jahren 1979-2003; nach DUGUY et al. (1980, 1997, 1998, 1999, 2000a, 2001, 2002, 2003, 2004), DUGUY und DURON (1981, 1982, 1983, 1984, 1985),
DUGUY (1986,1987, 1988, 1989, 1990, 1991, 1992, 1993, 1994, 1995, 1996).
Jahr Lebend Tot Gesamt
1979 > p 57
1980 ? > 42
1981 29 3 32
1982 26 3 29
1983 28 6 34
1984 24 6 30
1985 23 7 30
1986 12 4 16
1987 8 1 9
1988 15 16 31
1989 8 8 16
1990 15 9 24
1991 14 9 23
1992 4 6 10
1993 4 9 13
1994 2 7 9
1995 22 61 83
1996 79 25 104
1997 223 55 278
1998 158 32 190
1999 149 25 174
2000 22 23 45
2001 27 16 43
2002 19 5 24
2003 16 4 20
Summe 927 340 1366
Die Nachweise konzentrieren sich langs der Atlantikkuste besonders vox den Departements Vendee und Charente-Maritime, also am mittleren Kiistenab- schnitt, wahrend weiter nordlich und siidlich betrachtlich weniger Leder- schildkrotenfUnde und -sichtungen verzeichnet werden. Der Grofiteil lebender Tiere wird von Juli bis September beobachtet, wahrend die meisten Kadaver von Oktober bis Januar angespult werden (DUGUY 1983, 1997; Abb. 55).
Offenbar kommt es sogar gelegentlich zu Nistversuchen. Im Sommer 1938 wurde auf der lie d'Oleron (Charente-Maritime) ein 2 aufdem Strand von Vert- Bois in der Abenddammerung beobachtet. Ein weiterer Nistversuch wurde am 17. August 1978 bei Pointe d'Arcay (Vendee) verzeichnet. Das 2, das um 18:00 Uhr auf den Strand kroch, wurde durch eine Menge Schaulustiger, die zusam- mengelaufen war, rasch wieder vertrieben PURON-DUFRENNE 1978).
Belgien : Von der belgischen Kiiste werden nur selten Lederschildkroten-Nach- weise bekannt. COMPEL (1989) berichtet von der Strandung eines Kadavers (CL 135 cm) am 24. Dezember 1988 bei Middelkerke (51°12'N 02°49'O). Ein weite- rer Fund betraf ein Exemplar, das wahrscheinlich durch eine Schififsschraube zerrissen wurde. Ein Teil wurde am 10. September 1998 bei De Haan (51°17'N 03°02'O) aufgefischt, ein anderer am nachsten Tag bei Wenduine angespult (HAELTERS und KERCKHOF 1999). Es handelte sich vermutlich um ein cf mit einer GLvon etwa 165-175 cm (Stockmafi) und einer CL von etwa 135 cm. Eine dritte D. coriacea, ebenfalls ein <f (CL 194 cm, Gewicht 370 kg), strandete am 19.
Dezember 2000 bei Oostende (51°13'N 02°53'O; HAELTERS et al. 2001).
160
|12CH
"g 80 c
40
0 Strandungen
• Sichtungen
n
MAbb. 55: Jahreszeitliche Verteilung von 655 Lederschildkroten-Strandungen und -Sich- tungen im Golf von Biskaya (Frankreich); nach DUGUY (1997).
272 8. Dermochelyidae - Lederschildkroten
*£» fV
i
Abb. 56: Lederschildkroten-Nachweise im Mittelmeer; aus CASALE et al. (2003). Kleine Kreissymbole: einzelne Nachweise, grofie Kreissymbole: 13-25 Nachweise. Die Fundorte sind in CASA1E et al. (2003) aufgeschliisselt. Die gestrichelten Linien trennen das Mittel- meer in einen westlichen und einen osdichen bzw. einen nordlichen und siidlichen Teil
(vgl. Abb. 60).
274 8. Dermochelyidae - Lederschildkroten
Bereits friihe Naturforscher gingen von einem Vorkommen der Lederschildkrote imMittelmeer aus. So stammtbeispielsweise das Exemplar, anhanddessen VANDELLIUS (1761) die Lederschildkrote als „Testudo coriacea" beschrieb, aus dem Mittelmeer. Im 20. Jahrhundert glaubte man allerdings, dafi D. coriacea im Mit- telmeer selten ist (SCHREIBER 1912) und als Irrgast angesehen werden mug (MERTENS 1975). Tatsachlich ist die Art jedoch durchaus regelmafiig bier anzu- trefFen, wenngleich deutlich seltener als im europaischen Atlantik. Schon CAPOCACCIA (1968) fuhrte 54 Meldungen fur das Mittelmeer auf; davon stam- men allein 34 aus europaischen Gewassern (Spanien: 3, Frankreich: 6, Italien:
23, Ex-Jugoslawien: 2). In einer kiirzlich veroffendichten Obersicht sind nicht weniger als 411 Nachweise fur das gesamte Mittelmeergebiet zusammengestellt.
Sie zeigen, dafi die Lederschildkrote besonders im wesdichen und zentralen Mittelmeerbecken haufig vorkommt. Im Ostmediterran sind die Nachweise we- sendich sparlicher und konzentrieren sich auf den Bereich zwischen der Nil- miindung und Israel. Die 411 Nachweise betreffen, soweit bekannt, gestrandete (n = 52), zur Pflege aufgenommene (n = 9) und gefangene Lederschildkroten (n
= 170) sowie Sichtungen (n = 53; CASALE et al. 2003; Abb. 56).
Spanien: Von 1808 bis 2000 liegen insgesamt 70 Meldungen vor (CASALE et al. 2003), die sich uber fast die gesamte spanische Mittelmeerkuste sowie die Gewasser um die Balearen verteilen (Abb. 56). Erganzend zu den Nachweisen bei CASALE et al. (2003) sei erwahnt, dafi am 18. September 1993 ein Tier mit 162 cm BCL mit einem Schleppnetz ostlich des Ebro-Deltas in einer Tiefe von 35 m gefangen wurde (PEREZ et al. 1994). Im November 2001 strandete eine adulte D. coriacea m Cullera (39°10'N 0°14'W), Provinz Valencia (TOMAs et al.
2003).
Auf die Nachweise im Bereich der Strafle von Gibraltar wurde bereits oben eingegangen.
Frankreich : Zwischen 1520 und 1997 wurden exakt 50 Lederschildkroten im franzosischen Mittelmeergebiet nachgewiesen, wovon der Lowenanteil (39) auf den Zeitraum 1950-1997 entfallt. Unter diesen 50 Nachweisen betreffen sie- ben Lederschildkroten, die vor Korsika gesichtet oder gefangen wurden bzw. auf Korsika gestrandete Tiere. Ein zusatzlicher Nachweis bezieht sich auf eine im Juli 1989 vor Monaco gefangene Lederschildkrote (CASALE et al. 2003).
Italien: Aus Italien und italienischen Gewassern liegen aus dem Zeitraum 1743-2001 insgesamt 152 Nachweise vor, die sich langs der Tyrrheniskuste der Apenninen-Halbinsel, um Sizilien, besonders in der Storage von Messina, um die Pelagischen Inseln sowie im Golf von Tarent besonders konzentrieren. Langs der mittleren Adriakuste gibt es gar keine und an der nordlichen Adriakiiste nur sehr wenige Nachweise (CASALE et al. 2003; Abb. 56). Zusatzlich zu diesen Nach-
Tabelle 28. Nachweise von Dermochelys coriacea in maltesischen Gewassern in den Jahren 1970-1980; nach GRAMENTZ (1989).
Datum Ort Bemerkungen
22.03.1970 05.08.1970 09.10.1975 13.05.1976 03.07.1976 August 1976 09.09.1976 03.06.1977 13.07.1977 November 1978 Juli 1979
10 Seemeilen vor Zurrieq Zurrieq
Sliema Marsaxlokk Filfla
8 Seemeilen SO von Delimara Point Gozo
Spinola Bay Marsaxlokk
70 Seemeilen S von Malta 10 Seemeilen SO von Malta Ende Juli 1980 81 Seemeilen SO von Malta
Gespeert (starb am 6.8.1970 im ,,Maltaquarium''),
CL ca. 195 cm CL ca. 135 cm, 101 kg
Getotet, CL ca. 195 cm, ca. 300 kg Sichtung, entkam beim Fang- versuch
Getotet, CL ca. 185 cm Tot angespiilt, CL ca. 185 cm Getotet, CL ca. 180 cm Nach Fang gestorben, CL ca. 210 cm (?)
Langleinenfang, CL ca. 180 cm
weisen sei erwahnt, dafi Anfang der 1980er Jahre vor Sizilien einige weitere Leder- schildkroten gefangen wurden (D. GRAMENTZ unpubl.). Ihre Ruckenpanzer wurden noch im September 1985 in sizilianischen Pischhallen aufbewahrt. Die Tiere wurden an folgenden Orten gefangen: Spadafora (1983), 30 Seemeilen nordlich von Capo San Vito (1983), Ganzirri (1984) und Catania (1984).
Malta: Mitunter werden Lederschildkroten beim Fischen von Grofien Gold- makrelen {Coryphaena hippurus) und Schwertfischen (Xiphiasgladius) in maltesi- schen Gewassern gefangen (Tab. 28).
Kroatien und Montenegro :Am24. September 1894 wurde vor Budva eine Lederschildkrote gefangen. Am 12. September 1920 ging bei Kraljevica ein weiteres Exemplar in eine Thunfischfalle. Am 5. September 1956 wurde bei Omisalj auf der Insel Krk, am 6. September 1956 aufder Halbinsel Peljes'ac und im Juli 1990 bei Valbiska auf Krk jeweils eine Lederschildkrote nachgewiesen (LAZAR und TVRTKOVic 1995).
A1 b a n i e n: Nach HAXHIU (1995) befindet sich im Naturkundemuseum von Tirana eine Lederschildkrote, die wohl aus albanischen Gewassern stammen durfte.
276 8. Dermochelyidae - Lederschildkroten
Griechenland: Zwischen Juni 1968 und November 1984 wurden 14 D.
coriacea in der Agais nachgewiesen, wobei die meisten Tiere in Netze gingen
(MARGARITOULIS 1986). Am 29. Juli 2000 wurde ein Exemplar an einer Lang- leine vor den Cycladen gefangen (KAPANTAGAKIS 2001). BRUNO und MAUGERI (1976) fiihren zwei Lederschildkroten-Nachweise in den Gewassern vor Kreta auf. An der griechischen Westkiiste wird D. coriacea selten angetroffen. 1993 wurden zwei und 1995 ein Exemplar vor Kefallinia mit Langleinen gefangen (PANOU et al. 1999).
Tiirkei: Konkrete Nachweise aus tiirkischen Gewassern liegen erst seit weni- gen Jahrzehnten vor. 1985 brachten Fischer erne Lederschildkrote in den Hafen von Antalya (BARAN und KASPAREK 1989). An der tiirkischen Siidkiiste wurde 1994 ein Exemplar zwischen Anamur und Bozyazi angespiilt (CL 172 cm, CB 133 cm), und 1995 ging nicht weit entfernt, bei Karata§, eine D. coriacea in ein Fischernetz (TASKAVAK und PARKAS 1998). Am 4. Oktober 1997 wurde an der tiirkischen Agaiskiiste bei Oren (Provinz Balikesir) ein adultes Exemplar gefan- gen (GL ca. 180 cm, SCL 129 cm; BARAN et al. 1998). Nur einen Tag sparer wurde ganz in der Nahe, in der Bucht von Edremit (Balike§ir), in 10-13 m Tiefe ein subadultes c? mit einem Netz gefangen (SCL 129 cm BCL136 cm; TA§KAVAK
et al. 1998, TASKAVAK und FARKAS 1998). Ende November 1997 wurde zwi- schen Foca und Karaburun in der Bucht von Izmir (Izmir Korfez) eine dritte D.
coriacea in einem Schleppnetz gefangen, die wieder freigelassen wurde (TA§ KAVAK und FARKAS 1998).
Zypern : DEMETROPOULOS und HADJICHRISTOPHOROU (1981) fiihren zwei Nachweise auf.
Syrien: Am 4. August 2004 wurde etwa 2,5 km vor der Kiiste von Jableh eine subadulte Dermochelys coriacea (BCL ca. 140 cm) von Fischern gefangen (REES et al. 2004).
Israel: CAMINAS (1998) fuhrt insgesamt 12 Nachweise zwischen 1956 und 1987 auf, wovon die Halfte auf den Gaza-Streifen entfallt. Zwischen 2002 und 2005 strandeten mindestens drei Lederschildkroten in Israel, und am 6. Oktober 2001 wurde ein Exemplar vor Jaffa von einem Trawler gefangen (LEVY et al. 2005).
Fur Agypten gibt es nur relativ wenige Lederschildkroten-Nachweise, wobei dies vermudich auf ein Untersuchungsdefizit zuriickgehen durfte. FLOWER (1933) fand ein Exemplar auf dem Fischmarkt von Alexandria, und BAHA EL DlN (1992) berichtete von drei gestrandeten Tieren an der Kiiste der Sinai-Halbinsel. Zwi- schen 1998 und 2000 wurden hier vier weitere Kadaver angespiilt (CAMPBELL et al. 2001).
Auch die geringe Zahl von nur drei Nachweisen fur Li by en konnte auf ein Untersuchungsdefizit zuriickzufuhren sein. 1927 und 1928 ging je eine Leder- schildkrote bei Bengasi und Tripolis in eine Thunfischfalle (DELAUGERRE 1987);
1996 wurde bei Tajura, etwas ostlich von Tripolis ein weiteres Tier gefangen (CASALE et al. 2003).
Tunesien: Fur den Zeitraum 1907-1996 sind 36 Nachweise langs der gesam- ten tunesischen Kuste bekannt, wovon mehr als die Halfte aus dem Zeitraum 1983-1996 stammt. GrofStenteils handelt es sich dabei um Schildkroten, die in Thunfischfallen oder Fischernetze gingen (HELDT 1933, BRADAI und EL ABED
1998).
Algerien: Vom mittleren und westlichen Teil der Kuste liegen aus dem Zeit- raum 1867-1998 13 Nachweise, meist ohne nahere Details vor (LAURENT 1990, CASALE et al. 2003).
Marokko: Am 15. Dezember 1980 strandeten bei Restinga sechs Leder- schildkroten. Aus den 1980er und 1990er Jahren liegen drei weitere Nachweise fur die Mittelmeerkuste vor (DELAUGERRE 1987, LAURENT 1990, CAMINAS 1998).
Auf Nachweise aus der Strafie von Gibraltar wurde bereits oben unter „AtlantikK
bei Spanien eingegangen.
Fortpflanzung im Mittelmeergebiet: OfFenbar kommt es nur extrem selten zu einer Vermehrung von Dermochelys coriacea im Mittelmeergebiet (LESCURE et al. 1989), so dafi man im Unterschied zu Caretta caretta und Chelonia mydas nicht von einer indigenen mediterranen Population ausgehen kann. Die weitaus meisten Lederschildkroten im Mittelmeer stammen wahrscheinlich aus Nistgebieten im tropischen Atlantik (vgl. „Migrationen", S. 305).
Anscheinend kam es Ende des 18. Jahrhunderts zu vereinzelten Eiablagen an der nordwestafrikanischen Mittelmeerkuste, und sparer wurden hin und wieder 92 mit legereifen Eiern im Mittelmeergebiet gefunden (CAPOCACCIA 1968, LESCURE et al. 1989). Auch die sehr seltenen Fange von kleinen Lederschildkroten im Mittelmeer deuten auf eine gelegentliche Reproduktion hin. So soil ein Jung- tier (MNHN 632) mit einer SCL von nur 69,1 mm aus dem Pariser Museum National d'Histoire naturelle aus dem Mittelmeer stammen (DELAUGERRE 1987, LESCURE et al. 1989). Dieses Praparat gelangte wohl zwischen 1810 und 1830 in die Museumssammlung und wurde bereits von DUMERIL und BlBRON (1835, 1854) beschrieben und abgebildet (LESCURE et al. 1989). In Gefangenschaft auf- gezogene Lederschildkroten erreichen nach 22 Tagen eine vergleichbare Grofie
(DERANIYAGALA 1936), so dag man in diesem Fall von einer Fortpflanzung im Mittelmeerraum ausgehen mufi - wenn es zu keiner Fundortverwechslung gekom- men ist (vgl. GROOMBRIDGE 1990). Ein weiteres kleines Jungtier SCL 47 mm,
278 8. Dermochelyidae - Lederschildkroten
BCL 66 mm) wurde am 10. April 1896 in der Strafie von Messina gefangen. Es befindet sich heute in der Sammlung des Zoologischen Museums von Florenz (MZUF 12142) und wurde mit dem Vermerk „rarissima" inventarisiert (BRUNO 1978, LESCURE et al. 1989). Eine mittelgrofie Dermochelys coriacea von 50-60 cm Lange, die Mitte des 19. Jahrhunderts vor der algerischen Kuste bei Sidi Ferruch gefangen wurde (LALLEMANT 1867), muf? wohl gleichfalls auf eine Fortpflan- zung im Mittelmeergebiet zuriickgefuhrt werden (LESCURE et al. 1989), da Leder- schildkroten unter einer Panzerlange von 100 cm offenbar kaum aus tropischen Gewassern abwandern (ECKERT 2002).
Eine ganze Reihe von angeblichen jungtierfunden und Nistungen fur die sizi- lianische Sudkuste (BRUNO 1969, 1970, 1978, BRUNO und MAUGERI 1976) sind dagegen sehr fraglich, da es sich lediglich um Horensagen-Informationen han- delt. Dasselbe gilt fur mehrere Meldungen von Eiablagen in Korsika, Libyen und der Turkei (LESCURE et al. 1989). Audi ein angeblicher Nistversuch in Israel ist zweifelhaft. Am 30. Juni 1963 wurden am Strand von Palmachim (sudlich von Tel-Aviv) sehr grofie Spuren und eine unvollstandige Korpergrube (Durch- messer ca. 2 m) einer Meeresschildkrote gefunden (SELLA 1982). Auf eine Leder- schildkrote wurde jedoch nur aufgrund der Dimensionen der Spuren und der Korpergrube geschlossen (LESCURE et al. 1989).
Auf einen abgebrochenen Nistversuch auf Lampedusa (Italien) deutet aller- dings die Beobachtung einer Lederschildkrote mit einer Gesamtlange von 180- 200 cm am Conigli Beach hin. Das Tier wurde am 14. Juli 1985 nachts gegen 3:00 Uhr direkt an der Wasserlinie gesichtet. Es blieb fur etwa 2-3 min mit dem Kopfzum Strand gewandt und drehte dann langsam in siidostliche Richtung ab (GRAMENTZ 1989).
Loci typici: Die Nomenklaturgeschichte der Lederschildkrote ist verwir- rend und kompliziert. Lange Zeit wurde LINNAEUS (1766) als Erstbeschreiber von Testudo coriacea angesehen. FRETEY und BOUR (1980) fuhrten jedoch Argu- mente ins Feld, wonach VANDELLIUS (1761), auf den sich LINNAEUS (1766) be- zieht, als Autor gelten mufi. Hieriiber entziindete sich ein Disput (RHODIN und SMITH 1982, BOUR und DUBOIS 1983, SMITH und RHODIN 1986), der in der formalen Anerkennung von VANDELLIUS (1761) als Erstbeschreiber der Art mundete. FRETEY und BOUR (1980) wiesen nach, dafi das der Beschreibung von VANDELLIUS (1761) zugrunde liegende Exemplar bis heute im Museum der Universitat von Padua erhalten geblieben ist. Es mufi als Holotypus von Testudo coriacea Vandellius, 1761 gelten.
Ausfuhrliche Synonymien von Dermochelys coriacea finden sich u. a. in WERMUTH und MERTENS (1961,1977), PRITCHARD (1980) und PRITCHARD und TREBBAU (1984). In Abbildung 57 sind die Typuslokalitaten der folgenden auf die Lederschildkrote zu beziehenden Namen zusammengestellt:
280 8. Dermochelyidae - Lederschildkroten
A Testudo coriacea Vandellius, 1761 (Laurentum, zwischen Lido di Ostia und Tor Palermo, Tyrrheniskiiste, Italien)
B Testudo arcuata Catesby, 1771 (Locus typicus designatus MERTENS und WERMUTH 1955: Kiistengebiete von Carolina und Florida)
C Testudo lyra Lacepede, 1788 (Mittelmeer) D Chelonias lutaria Rafinesque, 1814 (Sizilien)
E Sphargis mercurialis Merrem, 1820 (Mittelmeer und Atlantik) F Sphargis angusta Philippi, 1889 (Tocopilla, Chile)
In der Karte sind die Typuslokalitaten fur drei weitere Namen nicht verzeichnet.
Am wichtigsten ist davon zweifellos Sphargis coriacea var. schlegelii Garman, 1884.
Dieser Name wurde haufig auf Lederschildkroten aus dem Pazifik und Indi- schen Ozean bezogen. Tatsachlich ist dies jedoch problematisch, da zuerst ein Lectotypus festgelegt werden miifste, um diese geographische Zuordnung recht- zufertigen (BRONGERSMA 1996). Nicht verzeichnet sind ebenfalls die Typus- lokalitaten von Testudo marina Wilhelm, 1794, die aus alien Weltmeere stammen soil sowie von Testudo tuberculata Pennant in Schoepf, 1801, die ohne Angabe einer Typuslokalitat beschrieben wurde.
Merkmalsvariation. Geschlechtsdimorphismus: Der Schwanz adulter cfcT ist etwa doppelt so lang wie der von 22 und reicht deutlich iiber den hinte- ren Carapaxrand hinaus. Bei c?d soil das Plastron konkav und bei 29 plan sein.
Aufierdem sollen sich o*cf von 22 noch durch eine schmalere „Huftregion", einen flacheren und schmaleren Panzer, ein konkaves Profil in der Nasenregion, einen starker gewolbten Schadel und endstandige Osteodermata, die an jedem Ventralkamm kraftige Erhebungen bilden, unterscheiden (DERANIYAGALA1939, PRITCHARD und TREBBAU 1984). Allerdings wurden bislang erst wenige cfcf eingehend untersucht.
Ein aufFalliges Farbungsmerkmal, das bislang nur von 29 bekannt wurde, ist der charakteristische rosafarbene Fleck auf der Kopfoberseite. Dieser Fleck wur- de z. B. fur nistende 29 aus Surinam und Franzosisch Guayana (PRITCHARD 1969,1971,1979, FRETEY 1978,1981, 1982), Trinidad (BACON 1970), Kolumbi- en (KAUFMANN 1973), den U.S. Virgin Islands (MCDONALD und DUTTON 1996), Mexiko (Michoacan; BENABIB 1983) und Siidafrika (Tongaland; HUGHES 1974a) beschrieben. Es ist unklar, ob dieser Fleck nur zur Nistzeit auftritt. Immerhin erwahnt CAPOCACCIA (1968), dafs auch ein adultes 9, das im Golf von Genua im Mittelmeer gefangen wurde, einen solchen Fleck hatte. Mitunter wird angenom- men, dafi dieser Fleck eine Narbe oder eine Scheuerstelle darstellt, die vom <f bei der Kopulation verursacht wird (PRITCHARD 1969, HUGHES 1974a). Wahr- scheinlicher ist allerdings, dafi der Fleck zur normalen Farbung von 99 gehort
(vgl. auch PR1TCHARD 1971, PRITCHARD und TREBBAU 1984). Unter dem rosa- farbenen Fleck liegt das Pinealorgan, das als Hormondriise eine wichtige Rolle fur zahlreiche biologische Zylden spielt (WYNEKEN 2001).
Makroskopische und histologische Unterschiede der mannlichen und weibli- chen Gonaden sowie der Miillerschen Gange von Embryonen und Schliipflingen wurden von BENABIB (1984), DUTTON et al. (1985) und RlMBLOT et al. (1985) beschrieben.
Altersbedingte Variation : Die wohl auffalligste ontogenetische Ver- anderung betrifft das Mosaik aus kleinen Schuppen, die bei Schliipflingen nodi den ganzen Korper bedecken (Abb. 58). Bald nach dem Schlupf beginnen sich die Schildkroten stiickchenweise zu hauten, so dafi allmahlich fast alle Schup- pen verschwinden. In einem Alter von ungefahr funf Monaten beginnen sich dann zunachst in den am Panzerrand liegenden Riickenpanzer-Kielen Ossikeln zu entwickeln. Spater bilden sich auch an den anderen Panzerkielen Ossikeln, und schliefilich werden aufdem Carapax auch die Flachen zwischen den Kielen mit solchen Knochenplattchen geschlossen. Der daraus allmahlich entstehende epithecale Panzer ist nach zwei Jahren vollstandig ausgebildet (DERANIYAGALA 1939, siehe auch unten unter Jugendentwicklung").
Wahrend des Wachstums nimmt die Korperlange von Lederschildkroten um etwa das 25-fache zu. Anhand einer in Gefangenschaft aufgezogenen Leder- schildkrote konnte DERANIYAGALA (1939) eine allometrische Reduktion der Kopflange, der Carapaxbreite und der Flossenlange zeigen. Besonders kompli- ziert sind die allometrischen Anderungen des Langenverhaltnisses von Rucken- und Bauchpanzer. Wahrend bei frischgeschliipften Dermochelys coriacea das Plastron zunachst kiirzer als der Carapax ist, iiberragt das Plastron ab dem Alter von etwa drei Wochen den Carapax. Im Alter von einem Jahr sind Rucken- und Bauchpanzer dann etwa gleichlang; spater ist der Carapax dann langer das Plastron (DERANIYAGALA 1939).
Auch die Korperfarbung verandert sich wahrend des Heranwachsens. So losen sich nach und nach die bei Schliipflingen vorhandenen weifien Langsstreifen auf dem Carapax auf, wahrend unregelmafiig verstreute weifie Flecken hinzutreten.
Diese Flecken werden bis zu einem Alter von knapp zwei Jahren grofier, an- schliefiend wieder kleiner (DERANIYAGALA 1939). Die zunachst iiberwiegend dunkle Unterseite hellt mit fortschreitendem Wachstum immer mehr auf, bis das Plastron und die Extremitatenunterseiten fast weifi sind (DERANIYAGALA 1939, BRONGERSMA 1970). Wie ein Vergleich der in BRONGERSMA (1972) und PRITCHARD und TREBBAU (1984) veroffentlichten Fotografien zeigt, existieren allerdings bei adulten Lederschildkroten erhebliche individuelle Variationen bei der Ventralfarbung. Das bei BRONGERSMA (1972) abgebildete 5 aus Cornwall hat ein iiberwiegend helles (weifies) Plastron mit vier zu Langsstreifen zusam- menfliefienden dunklen Makeln. Das Plastron des von PRITCHARD und TREBBAU
282 8. Dermochelyidae - Lederschildkroten
Q o
(1984) gezeigten 5 von der mexikanischen Pazifikkuste ist dagegen hell-dunkel meliert und nur die Langskiele treten rein weifi in Erscheinung.
Geographische Variation und Unterarten: Obwohl bei der Lederschildkrote von manchen Autoren zwei Unterarten anerkannt werden (sie- he unten), gibt es kaum quantitative Untersuchungen zur geographischen Varia- bilitat. Hinsichtlich der Schuppenzahl von Schliipflingen lassen sich keine Un- terschiede zwischen den Populationen von Sri Lanka (DERANIYAGALA 1939), Costa Rica (CARR und OGREN 1959) und Siidaftika (Tongaland; HUGHES et al.
1967, HUGHES und MENTIS 1967, HUGHES 1970, 1974a) feststellen, da sich die
"Werte stark iiberlappen. Nistende 99 im Ostpazifik (CORNELIUS 1978) und aus den Gewassern um die Philippinen (ERNST et al. 1994,2000) sollen durchschnitt- lich kleiner sein als bei anderen Populationen (siehe auch PRITCHARD und TREBBAU 1984).
Hinsichtlich der Farbung ist selbst bei ein- und derselben Brutkolonie eine erhebliche Variabilitat zu beobachten (PRITCHARD und TREBBAU 1984). Bei 99 aus Trinidad und Michoacan (Mexiko) variiert etwa der Anteil weifier Farbungs- elemente so stark, dafi fast ganz schwarze Exemplare neben deudich gefleckten vorkommen (BACON 1970, BENABIB 1983). Gelegendich in der Literatur erwahnte Lederschildkroten mit braun-gelblicher Zeichnung (z. B. DUMERIL und BlBRON 1835, YANEZ 1951) lassen sich allerdings auf Artefakte zuruckfuhren. Diese Lite- raturangaben beruhen auf praparierten Exemplaren, bei denen sich im Laufe der Zeit die Farbung unter LichteinfluC verandert.
Eine Untersuchung der mitochondrialen Kontrollregion deutet darauf hin, dafi die verschiedenen Brutkolonien der Lederschildkrote genetisch wenig difre- renziert sind. D. coriacea iiberlebte das Pleistozan moglicherweise nur in einem einzigen Refugialgebiet, das im Bereich des Indischen Ozeans oder des West- pazifiks gelegen haben durfte. Von dort aus wurden innerhalb der letzten 900.000 Jahre alle heutigen Brutkolonien begriindet (DUTTON et al. 1999).
Es ist offensichtlich, dafi sich von der Datenlage her kaum Argumente fur eine subspezifische Gliederung finden lassen. Dennoch werden von manchen Auto- ren zwei Unterarten unterschieden, deren Validitat allerdings umstritten und keinesfalls allgemein anerkannt ist (PRITCHARD 1980, PRITCHARD und TREBBAU
1984, ERNST et al. 1994, 2000):
(1) Dermochelys coriacea coriacea (Vandellius, 1761)
Verbreitung: Atlantik, Golf von Mexiko und Karibik von Labrador und Norwegen im Norden bis Argentinien bzw. zum Kap der Guten Hofmung im Suden; Mittelmeer.
(2) Dermochelys coriacea schlegelii (Carman, 1884)
Verbreitung: Pazifik und Indischer Ozean von Alaska bis Chile und vom Fernen Osten Rufilands und Japan bis Neuseeland, Tasmanien und OstafHka.
284 8. Dermochelyidae - Lederschildkroten
Morphologisch sind beide - wenn uberhaupt - wenig differenziert. Dermochelys coriacea scbkgelii soil im Vergleich zur Nominatform kiirzere Vorderflossen, einen grofieren Kopf und eine weniger kontrastreiche Farbung mit hellerer Fleckung besitzen (ERNST et al. 2000). Problematisch ist aufierdem, dafi der Name Sphargis scblegeln Carman, 1884 wohl gar nicht auf die pazifische Population zu beziehen ist (BRONGERSMA 1996).
Philippinische Lederschildkroten sind besonders klein. ERNST et al. (1994,2000) deuten an, dafi es sich dabei um ein eigenes Taxon handeln konnte, fur das der Name Dermochelys coriacea aiigtsta (Philippi, 1899) verfugbar ware. Da dieser Name mit derTypuslokalitat „Tocopilla, Chile" aufgestellt wurde, ist die Beschrankung auf Lederschildkroten aus philippinischcn Gewassern nicht unproblematisch.
Aufierdem ist unklar, zu welehen Brutkolonien im Westpazifik diese Tiere geho- ren.
£)kologie. Biotop: Haufig wird Dermochelys coriacea als die am starksten pelagisch Iebende Meeresschildkrotenart bezeichnet und angenommen, dafi sich Lederschildkroten mit Ausnahme der Nistsaison weit vom Land entfernt aufhal- ten und in grofie Wassertiefen vordringen (DERANIYAGALA 1930, 1932, CARR 1952, VlLLIERS 1958, BUSTARD 1972, SCHULZ 1975, PRITCHARD 1976, 1979, HENDRICKSON 1980, MARQUEZ1990). Neuere Beobachtungen relativieren die- ses Bild. Offenbar lebt D. coriacea namlich wahrend der warmen Sommer- und der ersten Herbstmonate regelmafiig in den flachen Schelfmeeren.
Auf das regelmiiEige Vorkommen von Lederschildkroten in Kustennahe wies DUGUY (1968) erstmals fur die franzosische Charente-Maritime hin.
BRONGERSMA (1972) bescatigte wenige Jahre spater, dafi D. coriacea ein regelma- fiiger Cast vor der gesamten europiiisehen Atlantikkuste ist, wobei sich insbeson- dere vor der franzosischen Atlantikkuste viele Lederschildkroten einfinden (sie- he oben unter „Verbreitung").
Regelmafiige kustennahe Nachweise liegen auch aus dem nordlicheren West- atlantik vor (vgl. auch BRONGERSMA 1972), u. a. aus North Carolina (LEE und PALMER 1981), Virginia (ERNST und GiLROY 1979), der Chesapeake Bay (HARDY 1969), New England (LAZELL1980) und Connecticut bis Neufimdland (BLEAKNEY
1965). Dasselbe gilt auch fur den Pazifik um Australien (LlMPUS und McLACHIAN 1979, LlMPUS 1984).
Beobachtungen aus Flugzeugen lassen darauf schliefien, dafi sich grofie Leder- schildkroten langs der nordamerikanischen Atlantikkuste eher uber dem Kontinentalschelf als uber Tiefwasser aufhalten (HOFFMAN und FRITTS 1982, FRITTS et al. 1983, SHOOP und KENNEY 1992). Systematische Zahlungen im Golf von Mexiko zeigten, dafi im August die grofiten Individuenzahlen in Ku- stennahe auftreten. Die Schildkroten halten sich dann in nur etwa 20 m tiefem Wasser auf. Im April werden die Tiere dagegen moist in Gebieten mit einer Was-