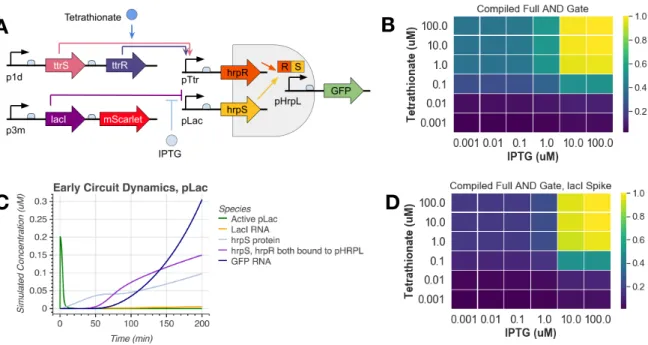
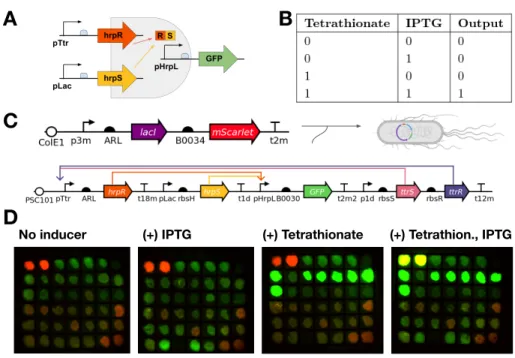
In this work, we implement a previously characterized split activator AND logic gate in the probiotic E. In the cases outlined here, engineered microbes constitutively express target molecules, without taking advantage of bacteria's ability to sense and respond to their environment . Optimization of the validated tetrathionate system was then performed in the context of the split activator AND gate in E.
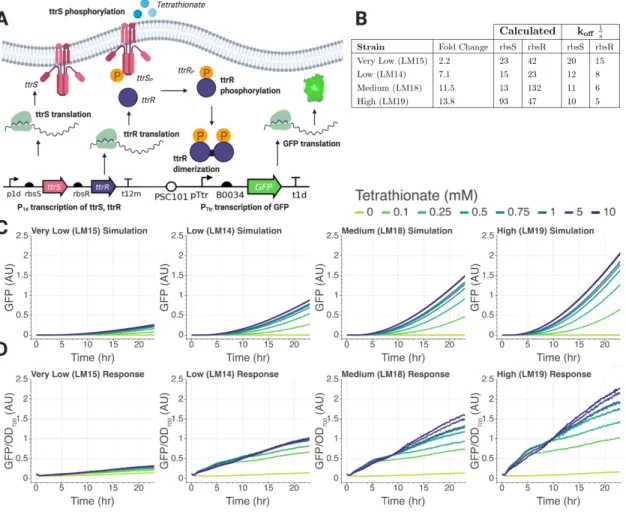
Modeling the Tetrathionate Two-Component System
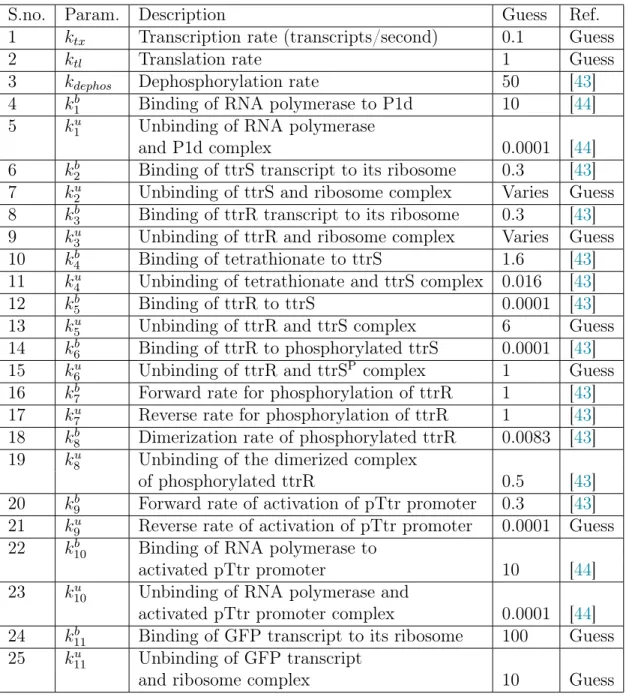
Since the ribosome binding strength is a tunable parameter in the experimental design, we keep this parameter free to change during the simulations to observe its effects. 7 k2u Binding of ttrS and ribosome complex Varies Guess 8 k3b Binding of ttrR transcript to its ribosome 0.3 [43].
Tetrathionate Sensor Validation
Since the various parameters in the model are context-dependent, parameter tuning of the validated in silico model can provide useful insight into the implementation of this circuit in the intestinal environment. Similarly, the effects of resource sharing and high burden due to protein expression can be quantified using this model. To this end, in the next section we discuss the results of preliminary data collected for the two-component system.
Engineering Two-Input AND Gate
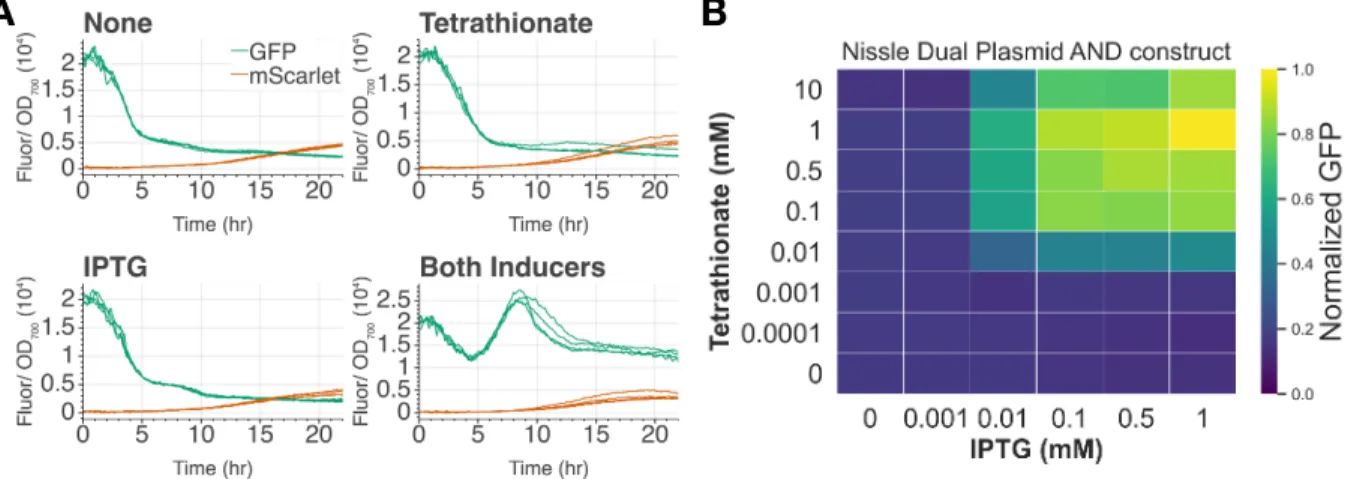
Should the AND gate function correctly, we expect GFP expression only when both inducers are present. In our experimental results, we observed a 6-fold induction when tetrathionate and IPTG are both added and minimally expressed in one or no inducer conditions. We further show the AND gate shows digital-like activation across a series of both inputs Figure 5-B.
To the best of our knowledge, this is the first functional inflammatory sensing EN gate in E. For example, if the EN gate senses epithelial proximity and tetrathionate, it is likely that tetrathionate could be detected first, as it is a small biomarker that may diffuse some distance away from the site of inflammation. Values displayed are GFP fluorescence normalized to OD, divided by the maximum value, which was reached at 1 mM IPTG and 1 mM tetrathionate.
We found that cells grown in IPTG and then added to tetrathionate reached their maximum level of induction faster than those grown in tetrathionate and then added to IPTG. Note that peak height may not be informative due to differences in Biotek fluorescence levels.
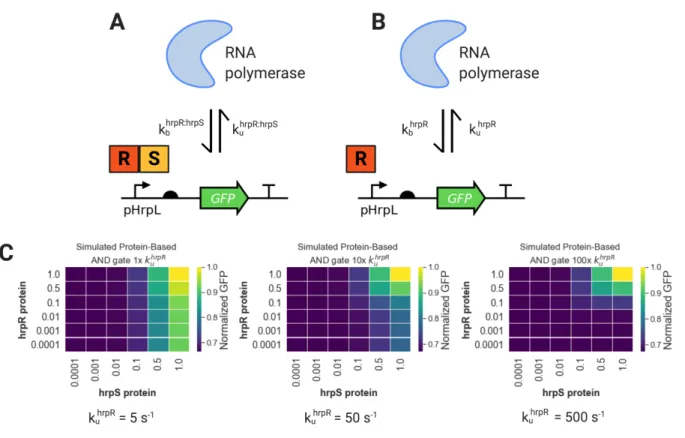
Design Space Exploration with an AND Gate Model
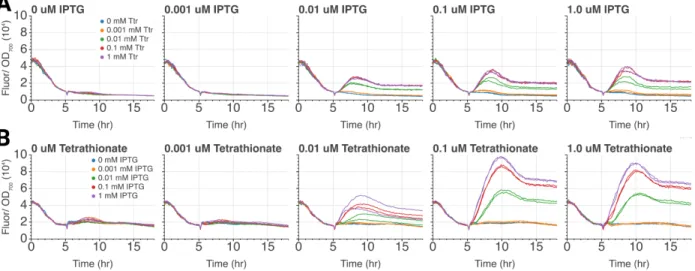
Indeed, we see that even small amounts of hrpR are now sufficient to activate the EN gate. To further analyze the system behavior, we extended the coarse grain model of the AND gate to also include the expression of the two activators, hrpR and hrpS. When we introduce 0 mM IPTG and 1 mM tetrathionate, we see that there is still a burst of active pLac at the beginning of the time course.
This led us to increase 1 mM lacI at the start of the experiment, which prevented the initial jump of unrepressed pLac. A key design point that emerges from this analysis is that if we start without LacI present, we get leaky IN gate activation independent of IPTG levels, as shown in Figure 8-B. If we look closely at the dynamics of the time course of the system, we find that this is indeed the case.
This induces small amounts of hrpR transcription, which is sufficient to activate the AND gate. To account for this in our model, we simply specified low concentrations of LacI as an initial condition, thereby recapitulating the expected AND gating dynamics as shown in Figure 8-D .
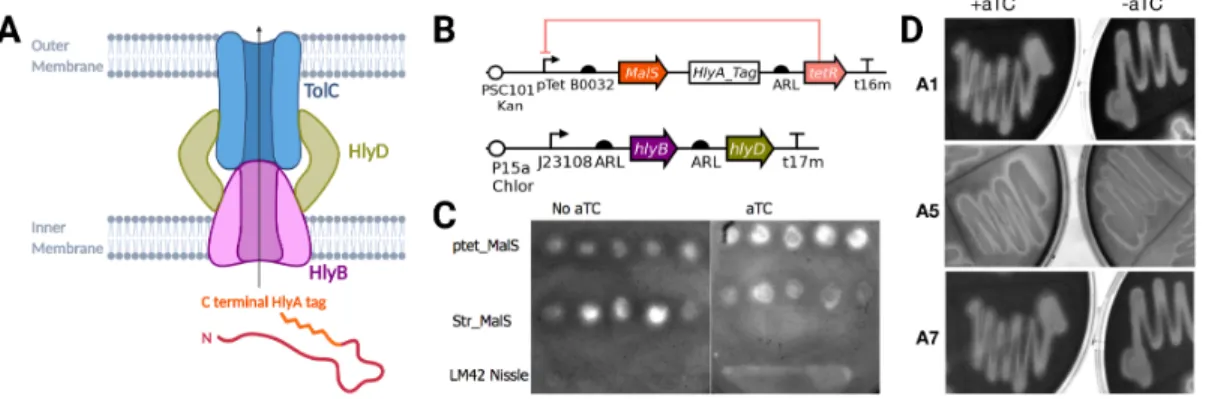
Secretion System Optimization
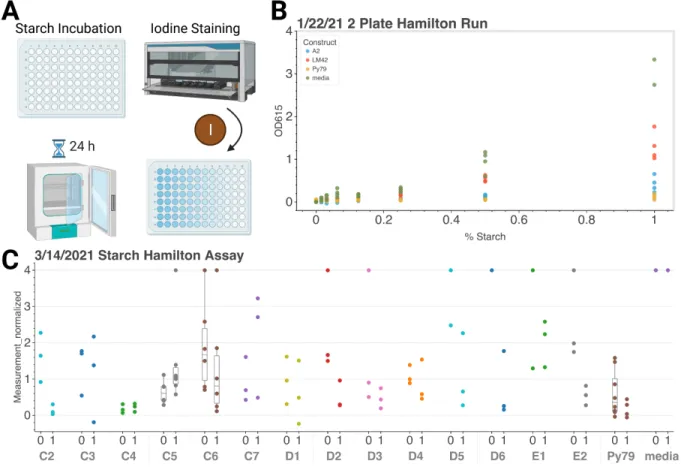
Addition of aTC appears to activate the Tet-inducible MalS construct, which has less starch present than in the no-inducer state. As shown in Figure 9-C, the starch agar plate test was first used to identify the potential success of this system. An increase in cleared (light) areas on the plate means increased starch clearance, and thus an increase in extracellular amylase MalS.
It is interesting to note that LM42, a Nissle strain without the secretion machinery or MalS does not have high starch clearance. While the level of the tetR repressor was optimized with a fluorescence screen, it appears to still leak in the aTC-free state. Initial tests shown in Figure 10-B show that the positive control, PY79, had only slightly greater starch degradation than the pTet-induced MalS construct, A2, which also has the HlyBD plasmid.
Some constructs show a decrease in starch in the activated case, suggesting that large amounts are leaking from the membrane or that this assay is flawed. Given that AND-gated GFP constructs also had a reduction in starch compared to media alone, it is likely that cell density is confounding the readout and altering the result.
Successful Secretion in E. coli Nissle and HB2151
We created a Hamilton STARlet protocol to add 30 µL of iodine to 100 µL of varying amounts of starch media.
Replacing Backbone and Promoter Maintains Secretion Efficacy
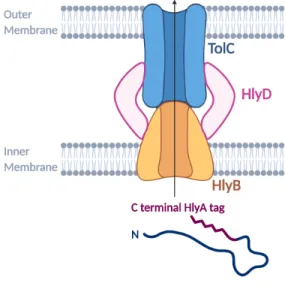
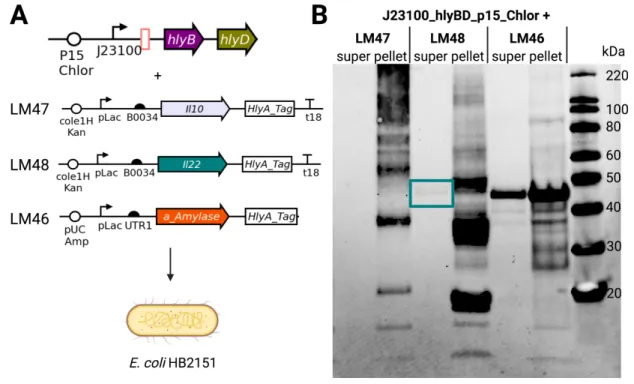
This choice was also made to allow the expression of the secretion machinery plasmid and the protein expression plasmid, which to. The nanobody to be secreted, anti-α-amylase, is under pLac expression on the pUC Amp backbone. Results show that the hlyA-tagged nanobody, which is 43.5 kDa, is present in the supernatant in all conditions, suggesting some leakage into the pLac promoter for HB2151.
We co-transformed the new backbone secretion plasmids with the VHH-containing plasmid against hlyA-tagged α-amylase. The new backbone, p15A, retains active secretion with plasmid pEHLY in Figure 12-C, as shown by the correct size band present in IPTG positive samples. It is possible that these bands correspond to early termination of translation, giving rise to a shorter hlyA tag while maintaining effective secretion.
Note that the nanobody on pEHLY is driven by pLac, so no shedding is expected in the absence of IPTG. Expression of VHH against α-amylase driven by pLac and J23100 show strong bands where expected.
Secreting Anti-Inflammatory Molecules
The successful constitutive secretion machinery construct from Figure 12-A was isolated and co-transformed with inducible IL-10 or IL-22. There are bands of the correct size in the supernatant of both IL-22 and positive secretion control. These three bands can be seen in the pellet of the IL-22 construct, indicating that these three products are present inside the cell.
This is the same size as the secreted band present in the supernatant, supporting the conclusion that secretion of the HlyA-tagged IL-22 monomer was successful. To further increase the amount of secreted anti-inflammatory product, B0034 was replaced with UTR1 as the RBS preceding both IL-10 and IL-22. A Western blot with anti-His antibody showing IL-22 secretion only when aTC and tetrathionate are present.
This could represent a leak in the pLac promoter, especially given the high copy number and strength of the RBS preceding the IL-22 sequence. With IL-22 secretion confirmed in the case of single-promoter induction, we proceeded to test secretion in the AND gate.
AND Secretion of IL-22
Interestingly, we see a peak induction at about 5 hours, which is faster than the 9-hour peak seen in the gates of the two IN plasmids shown in Figure 5-A. We amplified the backbone of the LM63 construct and replaced the GFP portion of the AND gate promoter with the IL-22 AND gate promoter construct. This would allow IL-22 to be secreted only when the AND promoter is activated, in the presence of both inducers.
Note that since the successful IL-22 secretion construct was on the ColE1 backbone, the copy number on the AND PSC101 plasmid would be lower. We co-transformed with a constitutive secretion machinery plasmid, as this was successful in previous IL-22 secretion experiments. We tested the first colony, LM83, to determine whether the AND knockout construct was active only in the presence of the two inducers shown in Figure 16-B.
We see the correct size band for the IL-22 monomer labeled with HlyA in the supernatant when both inducers are present. IL-10 and IL-22 sequences were ordered as gBlocks from IDT and resuspended in IDTE buffer.
Bacterial Strains
The tetrathionate construct was optimized by plate reader screening, while AND gates were first screened on inductor plates. Stereoscopic images were taken and colonies that were OFF in all cases except for both inducers were then selected for further plate reader assays. Note that in the full IN gate, mScarlet is constitutively expressed, so that one or none of the inductor plates is still red fluorescent.
The HlyBD genes were amplified from pVDL9.3 and inserted into vectors compatible with AND gate plasmids.
In vitro Aerobic Experiments
SDS-PAGE
Western Blotting
After completion of the protocol, the membrane was soaked in Tris-buffered saline (TBS), pH 7.6, for five minutes at room temperature with gentle shaking. During the TBS wash, 20 ml of the SuperSignal West Pico PLUS chemiluminescent substrate was prepared. We experimentally varied the ribosome binding sites, demonstrating that the stronger response of the tetrathionate two-component system could be recapitulated by varying the ribosome binding rates in our model.
We incorporated sensor logic by placing input from the split activator AND gate under the regulation of the tetrathionate and IPTG response promoters. We demonstrated successful secretion of IL-22 through the hemolysin pathway for the first time in E. Thanks to all members of the Murray lab for being supportive and providing excellent feedback during group meetings.
Zhou, Z. et al. Engineering probiotics as living diagnostics and therapeutics to improve human health. Microbial Cell Factories19,56.issn https://doi.org/. Analysis of the hemolysin transport process through secretion of PCM, CAT or beta-galactosidase from Escherichia coli fused to the C-terminal Hly signal domain. Su, L. et al. Extracellular overexpression of a recombinant Thermobifida fusca cutinase with an alpha-hemolysin secretion system in E.
Rao, S. et al. Against a living microbial microbicide for HIV: commensal bacteria that secrete an HIV fusion inhibitor peptide.