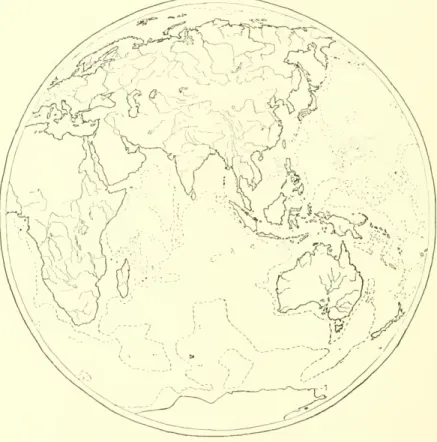
A single example of the use of host-parasite data may be given in the introduction to make the topic more concrete. In South America and Australia, the frogs of the genus Rana are missing, except in R. The place of true frogs in these southern lands is taken by the Leptodactylidae, a dominant group that might be called "southern frogs". This family is found in
Here again the parasites of the southern frogs provide evidence that can be used in connection with other considerations. If Zclleriella was once in the southern frogs in Arctogea, it should still be in some of the other Anuras of these lands. The absence of Zclleriella from Arctogea thus highlights the absence of the southern frogs, indicating that neither was ever at home in the north.
The hypothesis of a northern origin and southward spread of the southern frogs becomes grotesque in the light of the evidence furnished by Zclleriella. and we shall see later that the evidence of the southern frogs and. Similarly, the current occurrence of Hylidae (tree frogs), interpreted in the light of their parasites, indicates a southern origin and a southern spread. Kellogg repeatedly emphasized this idea of the interpretive value in avian taxonomy of evidence from their parasites.
NO. 8 PARASITES— METCALF 9
At least two other students independently came to the same realization of the importance of host-parasite data, with Launcelot Harrison being the next. In 1911 he discussed genetic relationships between hosts on the basis of their parasites, and curiously it was the birds and their Mallophaga that first brought him, as for Kellogg, this view of the use of parasite data. Among other findings, a study of Mallophaga would suggest the inclusion of penguins with birds, pigeons and tinamous, a relationship never before suggested.
From the greater variety of hosts on which they are found in South America, it appears that Temnocephaloidea evolved there and became parasites on the ancestors of Parastacus, and were carried with the migrating crustaceans to Antarctica, New Zealand, Australia and Madagascar (perhaps via the Molde Islands and the Molde Islands). has been suggested for many other animals). He also failed to realize the extent of Kellogg's appreciation of the broad applicability of the host-parasite method. But his understanding of the importance of parasites as signs of host relationships was reached independently of von Ihering and Kellogg and much in advance of my own.
The following quote demonstrates Harrison's understanding of the broad scope of the utility of the host-parasite method. In 1926, Harrison discussed before the Australian Association for the Advancement of Science "The Composition and Origin of the Australian Fauna, with Special Reference to the Wegener Hypothesis." Johnston, of Sydney, Australia, had heard Harrison present before the University of Sydney Science Association. .The first discussion of biting lice (Allalophaga) of birds as reassuring evidence of the genetic relationships of their hosts (Harrison 1911) and two years later Johnston (1913) wrote about the frogs of Europe, Amer-.
Kellogg also seems to have realized the applicability of the host-parasite method to problems other than genetic relationships of hosts, although he applied it little, if any, to them. Metcalf studied the opaline parasites found in the preserved Anura. frogs and toads) in the United States National Museum, including species from all parts of the world. Before applying the data from the study of the opalinid parasites, he tabulated the available data from both hosts and parasites under six items as follows: "Species of opalinid; Host species; Family or subfamily of host; Known geographic occurrence of opalinid in the species of host named; Known occurrence of host; Known.
Where data on host or parasite fossils are known and sufficiently comprehensive, they should be tabulated, for example under items such as these: Geographical location of host family fossils; Geological period of such fossils. Of course, the preservation of parasites as fossils will be rare, but their traces can be found and can be very specific, as for example in the case of the Peridermiums of pines. Let me mention here just a few things that seemed to be indicated with more or less probability by these earlier studies of Metcalf.
Group III before the separation of Australia from Asia in Jurassic or early Cretaceous times, in Australia or southeast Asia, spread to Europe during the Cretaceous or early Tertiary by a route north of the Himalayas, and to Africa in the late Tertiary, which from entering the northeast.
NO. 8 PARASITES METCALF I?
Metcalf subsequently published several papers discussing the host-parasite method or host-parasite data, as noted in the attached bibliography. It therefore seems likely that Darling may have been another independent discoverer of the wide significance of such data from parasites. Possibly, the primate ancestral tree may be revised after a study of the host relationships of their respective obligate nematode parasites.
In a study of biting lice of the family Gyropidae, Ewing (1924) discusses the importance of their geographic and host distribution and argues in favor of a cross between rodent and primate hosts. In a second paper (1924a) Ewing discusses the host-parasite relations of human and aphid races and the hybridization of both and he includes in this discussion prehistoric races of men and of their head lice, and he again mentions the probability that the tropical American spiders {Atheists) their head lice (Pediculus) "originally of man, but not of recent man." Two years later, the same author (Ewing, 1926) further discusses the importance of the geographical and host distribution of the genus Pcdiculus. Ward (1926), in a presidential address before the American Society of Parasitologists, mentioned the importance of such uses of data from parasites and referred in this connection to some of the work reviewed in the present paper.
As far as I can tell, the papers mentioned cover the previous use of data from parasites in connection with the three problem classes mentioned here. Harrison (1928) has, from this point of view, reviewed various groups of animal parasites in view of their accessibility for host-parasite studies. I know of no use of data from these charts for studying such general problems as we have had in mind.
Our knowledge of the taxonomy of these parasites, their host occurrence, and the geographic distribution of other parasites and hosts is insufficient, but there appears to be material for such host-parasite studies in these groups. There is a considerable degree of specificity in the host relationships of Endaniocbac and they are found in many groups of animals. Masses of flagellates are parasitic, and probably no other group presents more favorable material for host-parasite studies.
Although knowledge of learned parasites is extensive, it is ver^^ fragmentary, and is almost nil for many regions of the earth and far from complete leading regions and leading hosts. In some groups we have enough records to tabulate the host occurrence and geographic occurrence and examine the tables for what the}- may indicate.
NO. 8 PARASITES METCALF 21
But the important results that have already been obtained with the help of flatworm evidence are only a very small part of the harvest that could be. Darling's studies on the origin and spread of human races in the light of their hookworm parasites are an example of the use of data on roundworms (Nematoda). Nematodes compete with the trichonymphs of termites as a source of data for such applications, because, because of their universal abundance and the large number of their species, they must surpass the trichonymphs in the number and variety of problems that their evidence will help to solve.
Plants and their parasites, when studied by von Ihering's method, are sure to yield very important results, but such a study must be accompanied by further and painstaking study of the structure, life history, and taxonomy of the parasites. The fossil record of hosts is of particular interest to biogeographical problems and whether they can be reconciled with the fossil record of parasites. Such studies should ultimately determine not only the question of the former existence of such east-west migration routes, but also their position, connections, and geological time.
The most characteristic feature of the Chilian ant fauna is the occurrence of peculiar species of Monomosium, such as those inhabiting Australia and New Zealand, and of the genus Melophorus which occurs only in Australia and New Zealand. Probably Chile can be considered a part of ancient Archiplata, secured from Guyana and Brazilian immigrants by the heights of the Cordillera, but with only an incomplete set of the original Archiplatean fauna. Myro (aspider) with species—2 in the Antarctic Islands of New Zealand, I on Kerguelen Island, i at the Cape of Good Hope.
Many of the most common New Zealand polychaetous annelids are identical to the Strait of Magellan, Fuegia and Chili. Chilton (1909) states "of 13 species of the subantarctic islands of New Zealand, only 2 are endemic to New Zealand, 8 are found in South America or the Falkland Islands, and 2 extend to the Kerguelen". Two paragraphs from Cheesman (1909) may be quoted: .. Of 27 species of flowering plants and ferns known from the Kerguelen-South Georgia region, 20 are also found in the subantarctic islands of New Zealand, while 27 are found in Fuegia and the Falkland Islands.
The total number of Fuegian plants found in the subantarctic islands of New Zealand is 29, of which 14 also extend to the Kerguelen and South Georgia archipelagos. Eleven species of plants found on the subantarctic islands of New Zealand occur either in the Tristan da Cunha Group of the South Atlantic or in The last few pages have noted a few sources of data for but one series of problems associated with the biogeography of the southern Peninsula.
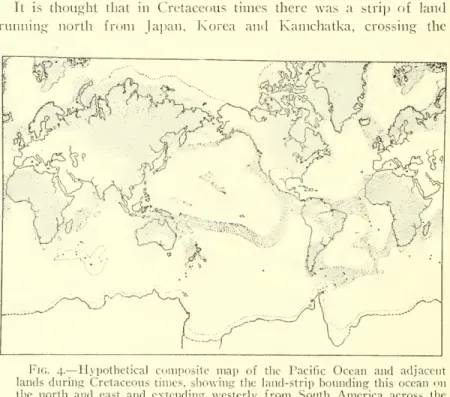
4.—Hypothetical composite map of the Pacific Ocean and adjacent lands during the Cretaceous period, showing the belt of land bounding that ocean on the north and east, and extending westward from South America across the South Pacific to Papua and Australia.
NO. 8 PARASITES METCALF 33
LITERATURE CITED
Opalinid parasites and the geographical distribution of bellfish (Discoglossidae), with a critique of the age-range hypothesis.