In the main part of the thesis, the sequence of the BALB/c H-2Dd transplant antigen gene is presented. Appendix B contains the sequence of the H-2Ld gene, which was the first transplant antigen gene to be sequenced. DNA sequence of the mouse H-2Dd transplant antigen gene (transplant antigens/class I genes/H-2Dd protein sequence).
The mouse major histocompatibility complex (MHC), located on chromosome 17, has four regions containing class I genes: _ H-20, H-2K, Qa, and the Tla complex (1-4). In this publication, we present the substantial new sequence of the H-2Dd protein and the complete DNA sequence of the H-2Dd gene. A second region of the H-2Dd gene, the seventh intron, also contains potential alternative splicing sites.
It is interesting to note that all of the potential alternative splice acceptor sites listed in There is a third eDNA whose DNA sequence is >99% homologous to the coding sequence of the H-2Dd gene. Comparisons between the H-2Dd gene sequence and the sequences of the other H-2d haplotype genes, the H-2Kd and H-2L d genes, reveal several interesting points.
The only region of the H-2Dd gene that shows significantly higher homology to the H-2L d gene than to t~e. As noted in the discussion of the pAG64 eDNA clone ( 34 ), divergence between these two sets of sequences results in the insertion of a sequence that is 84% homologous to the consensus sequence of the 82-repeat sequence family ( 39 ). In general, the H-2Dd sequence is more homologous to the transplant antigen sequences than to the sequences of the Qa region molecules.
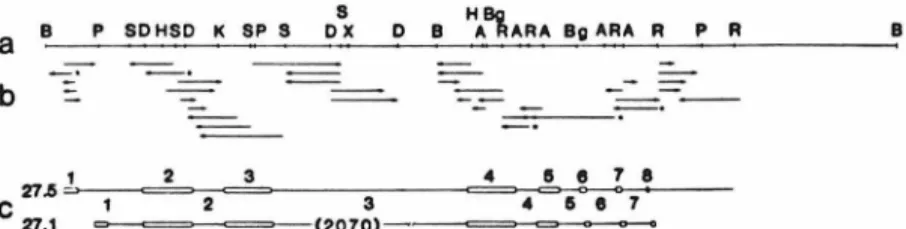
The restriction maps of the two types of clones differ only in the 3' sequence flanking the H-2Dd gene. This figure shows the structure of the H-2Dd gene and the sequencing strategy used to determine its sequence. The major histocompatibility complex plays a fundamental role in the regulation of the vertebrate immune response.
This gene is a pseudogene that maps to the Qa-2,3 gene cluster of the Tla complex. The eight exons of Gene 27.1 CorTeepond to the structurally defined protein domains of the TranaplanutJon antigen. Sequences of the extracellular domains of gene 27.1 are 73-85% homologous to the Kb molecule at the protein level (Table 2).

Gene transfer studies and comparison of the translated sequence with the partial amino acid sequence of the Ld transplant antigen establish that gene 27.5 encodes an L" polvpeptide. A comparison of the gene 27.5 sequence with various complementary DNA sequences indicates that the BALB /c mouse may contain a number of closely related L-like genes. The graft or class I antigens of the major histocompatibility complex (MHC) are present on the surface of all mammalian somatic cells and play a key role in T cell -immunosurveillance (/, 2).
We cloned 30 to 40 genes of BALB/c mice (haplotype H-2d) that are homologous to complementary DNA (eDNA) probes for transplantation antigens (4) and demonstrated through gene transfer experiments that one of these clones (27.5) encodes the transplantation antigen L d (5). All exon-intron boundaries have consensus signals for upstream or downstream RNA splicing (12). The translated sequence of the 27.5 gene is identical to the amino acid sequence of the Ld molecule at 77 out of 77 comparable positions (Table I) (13).
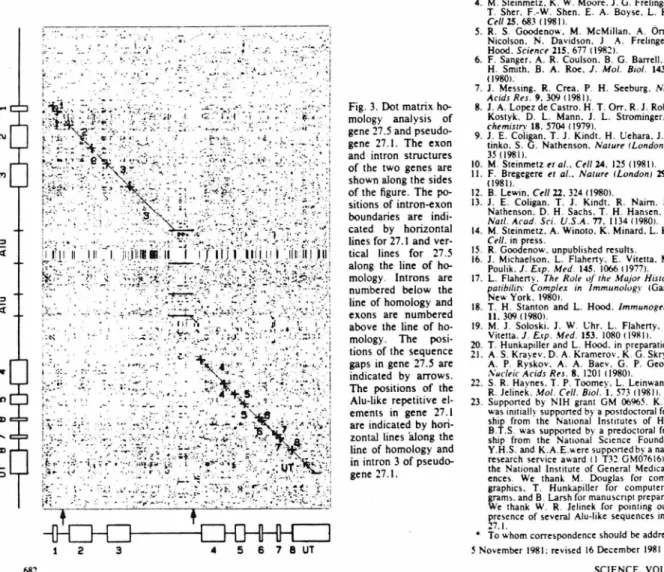
Two of the sequence differences are in the first 60 nucleotides of the found exon and lead to one codon substitution. The differences are gathered in the first 45 nucleotides of the found exon and lead to five codon substitutions. The homology of the translated part of each ex to the corresponding part of the Kb and Ld sequences. as well as to the translated 27.1 pseudogene exons 14).
Therefore, mice of the H-2d haplotype may have three closely related class I genes encoding L d -like polypeptides. I and the L d 27.5 gene with the amino acid sequence of the mouse Kb molecule (Table I) shows that the 27 .I pseudogene appears to have deviated significantly more from the translated Ld gene than has the Kb molecule (Table I). 27 .I is a Qa-2.3 pseudogene [see (4) for discussion), then this homology suggests that the Qa antigens are class I molecules and that the Tla complex should be considered part of the H-2 complex.
The exonic and intronic structures of both genes are shown on the sides of the figure. An unrelated error was in the misidentification of the location of the murine B1 and B2 repetitive sequences (referred to in the paper as Alu-1 ike sequences). This B1 sequence is in a position analogous to the position of the B1 sequence in the third intron of pseudogene 27.1.
![Table I. Homology of the exons of the translated L d gene (27.5) to the corresponding regions of the translated pseudogene 27.1 (4) and to the K" (/])and Ld (JJ) molecules](https://thumb-ap.123doks.com/thumbv2/123dok/10405466.0/58.922.158.615.611.769/table-homology-translated-corresponding-regions-translated-pseudogene-molecules.webp)
TL TL TL TL Mouse L tk- cells were co-transformed with -I 1-
ON A from each of the A clones that yielded Kd-positive transformants was restriction-mapped with the indicated enzymes by single and double digestion as previously described24. Transformants of c50.2 DNA appear to express low levels of the Qa-2,3 antigen as detected by radioimmunoassay with monoclonal antibodies specific for these products !Fig. 4d. 132-microglobulin !see below) does suggest that significant amounts of the antigen may be expressed in an altered form associated with 132-microglobulin.
The origin of the H-2 and Tla regions for each of the mice is shown in the panel. . Al7.3 or A24.8 Dl':A destroys the ability of these genes to transform L cells to a TL-positive phenotype. Therefore, expression of foreign class I molecules on the mouse L cell surface should result in an increase in the amount of 1'2-microglobulin expressed on the cell surface. Moreover, the upregulation of 132-microglobulin expression appears to be a stable phenotype observed after SO passages of some cell lines in culture.
The ability to measure an increase in 132-microglobulin expression by the co-transformants is related to the fact that under the conditions used for transformation, almost 100% of the co-transformants express significant and comparable levels. 7 Fluorographs of two-dimensional gels of the products of the c2.1 gene, immunoprecipitated from KB-1 cells Ia and tk • transformants lbi using C3H antibodies against K8-1 transformants. The antibodies prepared in C3H mice against c2.1 transformants tKS-1 cells were used to immunoprecipitate radiolabeled antigens for two-dimensional gel electrophoresis as previously described18. The insets contain longer exposures and schematic representations of the 45,000 molecular weight region of each gel. .
The products of the c2.1 gene appear to have a molecular weight of ~45,000 and to consist of a number of spots of the right pi in a consecutive array somewhat characteristic of class I molecules analyzed on two-dimensional gels ( see figure). new genes 1c2.1 l may encode a differentiation antigen based on the pattern of tissue distribution determined with the alloantibodies. For example, the third external domain of the H-2Dd molecule contains 95 amino acids, but only the 92 amino acids corresponding to the 92 amino acids.