Basant Kumar Patel for his indispensable guidance and encouragement throughout the period of his master's thesis work. Parag Pawar for their thoughtful and valuable comments, constructive criticism and tireless review of the work. The cause of sporadic ALS is unknown, but there are associations with occupations that involve heavy work, exposure to heavy metals, or a history of traumatic injury.
This model exhibits some of the pathological features associated with ALS in humans, such as cytoplasmic protein aggregation and cytotoxicity. TDP43-YFP when expressed in yeast forms toxic aggregates that inhibit cell growth. Aβ42 fused to the functional domain of Sup35, when expressed in yeast acts as a disease model for AD.
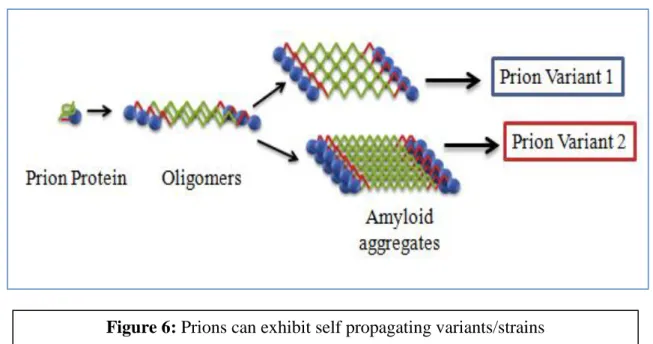
Prions can reproduce as different strains due to the different aggregated conformation of the same protein. It is proposed here that several Aβ prion variants, even from wild-type Aβ sequences, can also be acquired in yeast, which may act as a disease model for AD.
Introduction
- Amyloids and Prions
- Yeast: Model for prion and Amyloid diseases
- Prion strains
- Amyotrophic Lateral Sclerosis
- Alzheimer’s Disease
Prions can reproduce as different strains due to different conformation of the protein in the protein aggregates. Yeast has been used as a model system to study the disease mechanism of various neurodegenerative diseases such as Alzheimer's [15], Parkinson's [16] and Huntington's [17] by expression of the pathological protein. Prion Domain of Pathogenic protein can be fused with Sup35MC and the fusion protein can be expressed in yeast to study the mechanism of the disease.
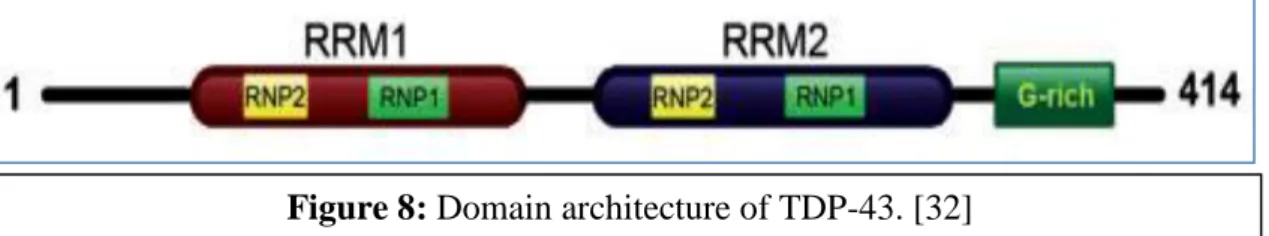
The prevalence of the disease is 5 individuals per 100,000 and the risk increases after the age of 60. The pathological TDP-43 is hyperphosphorylated, ubiquitinated and cleaved to generate C-terminal fragments of the protein [32]. TDP-43 aggregates are non-amyloid-like and mutation of the protein in the C-terminal region increases its aggregation and cytotoxicity [34].
This model exhibits some of the pathological features associated with ALS in humans, such as nuclear localization and cytoplasmic aggregation of the protein. And it has been suggested that the toxicity is caused by loss of function of TDP43 protein due to aggregation rather than due to accumulation of the protein. Aβ42 fused to the functional domain of Sup35 when expressed in yeast produces Aβ42-Sup35MC fusion proteins, which in the [PSI+] ade1-14 strain will give rise to red colonies on rich growth medium (YPD) due to functional C-terminal region of Sup35MC and will not survive on SD-Ade media as the Ade protein produced is truncated.
Prions can propagate as different strains due to the different conformation of the protein aggregates and it has been observed that at least two different strains of Aβ prions, i.e.
![Table 1: List of examples of neurodegenerative protein misfolding diseases [1, 10, 11]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/9.892.160.783.118.582/table-list-examples-neurodegenerative-protein-misfolding-diseases-10.webp)
Materials and Methods
Materials
Methods used for studying TDP- 43
- Preparation of Media
- Plasmid Isolation and Yeast Transformation
- Spot Assay to study the toxicity of TDP- 43 expression in
- Spot Assay to study the effect of 3- Aminophenol on toxicity of
- Selection of carbon source to study the effect of zinc/copper
- Effect of 1% Galactose and 10mM zinc chloride/5mM copper sulphate 15
- Optimization of copper concentration
- Optimization of galactose concentration
- Fluorescence Microscopy to check for different levels of expression
- Effect of 0.1% and 1% Galactose and 3mM zinc chloride
While checking the effect of metal ions and small molecules, these compounds were added together with SRaf (1%) +0.1% Gal-Ura medium. Protein expression can be induced by galactose, and the level of expression can be controlled by varying the concentration of galactose. 500 l of PB buffer was added and allowed to stand for 1 minute, then centrifuged for 2 minutes at 10,000 revolutions per minute and the flow-through was discarded.
700 l of PE buffer was added and was centrifuged at 10000 rpm for 2 minutes and the flow-through was discarded. Residual ethanol in the column was allowed to dry and the column was centrifuged for 2 minutes at 10,000 rpm. The QIA preparation column was placed in a clean eppendorf and 50l of autoclaved water was added to the center of the column.
This was allowed to stand for 5 minutes and centrifuged at 14000 rpm for 2 minutes and the concentration was found using nanodrop. The culture was grown overnight on a rotary shaker at 200 rpm and 30°C; 2 ml of the culture was centrifuged and washed with 1 ml of autoclaved water to remove traces of media. The culture was grown overnight on a rotary shaker at 200 rpm and 30°C; and absorbance was measured at 600 nm.
2 ml of the culture was centrifuged and washed with 1 ml of water to remove traces of medium. 5 ml of the culture was centrifuged and washed with 1 ml of autoclaved water to remove traces of medium. Inv Sc1+ control plasmid cells were grown overnight in 5 ml SRaf-Ura in a rotary shaker at 200 rpm and 30°C.
3 ml of the culture was centrifuged and washed with 1 ml of autoclaved water to remove traces of media. Then, a drop of immersion oil was placed on the coverslip and the cells were observed under oil immersion 63x and 100x magnification. The culture was allowed to grow overnight and 2 ml of the culture was centrifuged and washed with 1 ml of autoclaved water to remove traces of media.
Methods used for studying Aβ42-Sup35MC fusion
- Preparation of Media
- Toxicity of Aβ42-Sup35MC expression in yeast
- Search for Aβ42-Sup35MC amyloid variants
Inv Sc1 + control plasmid and Inv Sc1 + TDP43-YFP were grown overnight in 5 ml of SRaf-Ura. 2% glycerol instead of 2% dextrose and prohibits the growth of small cells with respiratory defects that cannot survive on non-fermenting carbon sources and allows the growth of healthy cells [52]. The yeast strain was revived at -80°C on a YPD plate and subcultured on YPD plates.
The cells were plated on YPG plates and incubated at 30°C for 5 days for the selection of healthy cells. Cells from the YPG plate were inoculated into 10 ml YPD broth and into 10 ml YPD+50 µM CuSO 4 broth. 5 µl of the culture was spotted in triplicate on YPD and YPG plates and incubated at 30°C for 2 days (YPD) and 5 days (YPG).
Cells from YPD were plated on YPG plates and incubated for 5 days at 30°C to select healthy cells.
Results and discussion
- Toxicity of TDP-43 expression in yeast cells
- Spot Assay to study the effect of 3-Aminophenol on toxicity of TDP-43 expression
- Selection of Carbon source
- Effect of 1% Galactose and 10mM zinc chloride/5mM copper sulphate
- Optimization of Zinc Concentration
- Optimization of Galactose Concentration
- Fluorescence microscopy to check for different levels of
- Effect of 0.1% Galactose and 3mM zinc chloride
- Effect of 1% Galactose and 3mM zinc chloride
- Concentration optimization of copper sulphate
- Toxicity of Aβ42-Sup35MC expression in yeast
- Search for Aβ42-Sup35MC amyloid variants
In situ assay to study the effect of 3-Aminophenol on the toxicity of TDP-43 expression in yeast cells. When cells are exposed to 10 mM zinc chloride or 5 mM copper sulfate [ 47 ], cell growth is further inhibited in the SD-Ura sample due to zinc/copper toxic effects and glucose suppression. 1% galactose induction causes high expression of the TDP43-YFP fusion protein, and the concentration of zinc chloride and copper sulfate used was determined from a previous work [ 47 ].
High expression of TDP43-YFP together with zinc chloride causes toxicity compared to 1% galactose sample. Copper sulfate in molar concentration of the mill causes very high toxicity compared to 1% Galactose sample. To find the optimum concentration of zinc, different concentrations of were checked The optimum concentration of zinc was found between 2 mM and 4 mM i.e.
If the zinc concentration is more than 3 mM, zinc itself is toxic to the cells because growth is stunted and the effect of TDP43-YFP expression cannot be studied. TDP43-YFP is under the Gal promoter and when the promoter is induced with a high concentration of galactose, the protein is overexpressed and the protein tends to misfold. The optimal galactose concentration at which a balance between aggregation and toxicity was observed was 0.1% galactose.
To check the time required for the formation of fluorescent foci, the cells were incubated for different time intervals of 24 hours, 48 hours and 72 hours. 0.1% and 0.01% galactose were found to be the optimal concentrations for maintaining TDP43-YFP foci. 0.1% galactose was used to maintain the balance between aggregation and toxicity caused by the protein.
Results obtained from the previous experiments indicate that molar concentration of copper is toxic to the cells. Growth of cells expressing Aβ42-Sup35MC (YPD+50 M copper sulfate) is less than the cells not expressing Aβ42-Sup35MC (YPD). The cells expressing Aβ42-Sup35MC appear to have mitochondrial dysfunction as the cell growth on YPG plate is inhibited compared to the cells not expressing Aβ42-Sup35MC.
Aβ42 fused to the functional domain of Sup35 when expressed in yeast produces Aβ42-Sup35MC fusion proteins, which in the [PSI+] ade1-14 strain will give rise to red colonies on rich growth medium (YPD) due to functional C-terminal region of Sup35MC. If the protein is insoluble, it will give white colonies indicating prion formation, leading to loss of function of the C-terminus of Sup35MC.

Conclusion
Outeiro, Tiago Fleming and Paul J. Park SK, Pegan SD, Mesecar AD, Jungbauer LM, LaDu MJ, Liebman SW. 34; Development and validation of a yeast high-throughput screen for inhibitors of Aβ₄₂ oligomerization". Michael McCaffery, Susan Lindquist and Aaron D A yeast TDP-43 proteinopathy model: Exploring the molecular determinants of TDP-43 aggregation and cellular toxicity". Deletion analysis of the SUP35 gene from the yeast Saccharomyces cerevisiae. reveals two non-overlapping functional regions of the encoded protein."
The yeast non-Mendelian factor [ETA+] is a variant of [PSI+], a prion-like form of release factor eRF3”. Exposure to environmental toxicants and pathogenesis of amyotrophic lateral sclerosis: state of the art and research perspectives”. Elman, Dana Clay et al. TARDBP mutations in amyotrophic lateral sclerosis with TDP-43 neuropathology: a genetic and histopathological analysis.
Doudeva, Yi-Ting Wang, Che-Kun James Shen and Hanna S. Johnson, Brian S., David Snead, Jonathan J. Michael McCaffery, James Shorter and Aaron D. and amyotrophic lateral sclerosis-linked mutations accelerate aggregation and increase toxicity. " Journal of Biological Chemistry 284, no. 34; TDP-43, the signature protein of FTLD-U, is a neuronal activity-responsive factor." Journal of Neurochemistry 105, no. H., Foon Wu, David Harrich, Leon F Cloning and characterization of a novel cellular protein, TDP-43, that binds to human immunodeficiency virus type 1 TAR DNA sequence motifs." Journal of virology 69, no. (ALS)-associated TDP-43 aggregation by the proteasome targeting factor, ubiquilin 1”.
![Figure 1: Fluorescence spectra of ThT in the absence (solid line) of peptide, and in the presence of preaggregated (1-28) (short dashed line), or (1-40) peptide (long dashed line) A) Excitation spectra B) Emission spectra [2]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/8.892.182.780.461.755/figure-fluorescence-spectra-absence-presence-preaggregated-excitation-emission.webp)
![Figure 4: The [PSI+] yeast prion. [21]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/11.892.96.777.80.1104/figure-4-the-psi-yeast-prion-21.webp)
![Figure 5: The [RPS+] and [rps-] prion in yeast. Prion domain of Rnq1 when fused with Sup35 mimics [PSI+] prion](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/11.892.152.763.101.556/figure-prion-yeast-prion-domain-rnq1-sup35-mimics.webp)
![Figure 7: The [PSI+] yeast prion exhibits three variants [26]: Strong [PSI+] (White), Weak [PSI+] (Pink) and [psi–] (red)](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/12.892.186.752.752.969/figure-yeast-prion-exhibits-variants-strong-white-weak.webp)
![Figure 8: Domain architecture of TDP-43. [32]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/14.892.140.804.78.1128/figure-8-domain-architecture-of-tdp-43-32.webp)
![Figure 11: Amyloid Precursor Protein (APP) showing the N-Terminal signal peptide, a serine protease inhibitor domain of the Kunitz type (KPI) and amyloid β-peptide (Aβ) including the transmembrane domain (TM) [43]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/15.892.172.783.447.843/amyloid-precursor-protein-terminal-protease-inhibitor-including-transmembrane.webp)
![Figure 10: Hallmarks of AD: Amyloid plaque and neurofibrillary tangles [42]](https://thumb-ap.123doks.com/thumbv2/azpdfnet/10552072.0/15.892.159.791.165.1116/figure-10-hallmarks-ad-amyloid-plaque-neurofibrillary-tangles.webp)
