Department of Experimental Biology, Center for Advanced Studies in Olive Groves and Olive Oil, Faculty of Experimental Sciences, University of Jaén, Jaén, Spain. HONg li · Center for Advanced Proteomics Research, Rutgers and Department of Microbiology, Biochemistry and Molecular Genetics, New Jersey Medical School, Newark, NJ, USA.
NO Treatment and Detection
Abstract
1 Introduction
A simple and useful method to apply exogenous NO gas to plant systems: Peppers as a model. Therefore, in this chapter we report an easy and inexpensive method, free of side effects, to apply NO gas to plant systems using peppers as a model.
2 Materials
To date, most information available on the effect of NO in plant tissues/organs, including fruit, has been obtained from studies conducted with NO-releasing substances as referred to above, although there are also some reports that have used NO gas fumigation to determine the plant response. to study [22–25]. Recently, the involvement of exogenously applied NO gas in the delay of the ripening of sweet pepper fruits was described together with some post-translational modifications undergone by proteins as a result of the treatment [26].

3 Methods
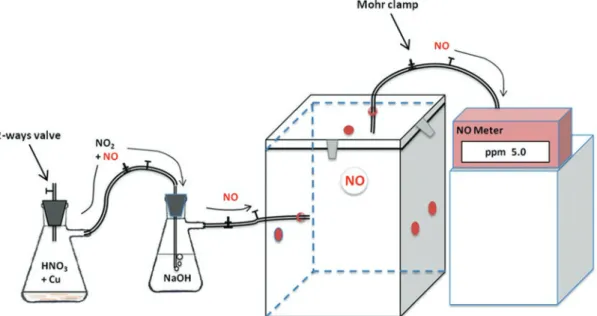
To apply the treatment of NO gas to the other fruits, the following steps are developed, and a schematic of the whole apparatus can be viewed in Fig. Then connect the cover of the IC to the inlet of the NO meter and open the 2-way valve.
4 Notes
Aliquots of fruit for further tests, which may include either determination of NO using different methods [12], tests of protein nitration and protein S-nitrosylation [26–28], among others, were taken after 7–10 days at room temperature after the start of the experiment . connection holes for the supply of a set of gases. Under conditions of saturated NO at the beginning of the treatment, nitrogen dioxide is formed by reaction 1. When the concentration of NO decreases after the initial reaction with copper, NO is formed according to reaction 2. Reaction 1) Cu (s) + 4HNO3 (aq) → Cu (NO3) 2 (aq) + 2 NO2 (g) + 2 H2O.
Acknowledgment
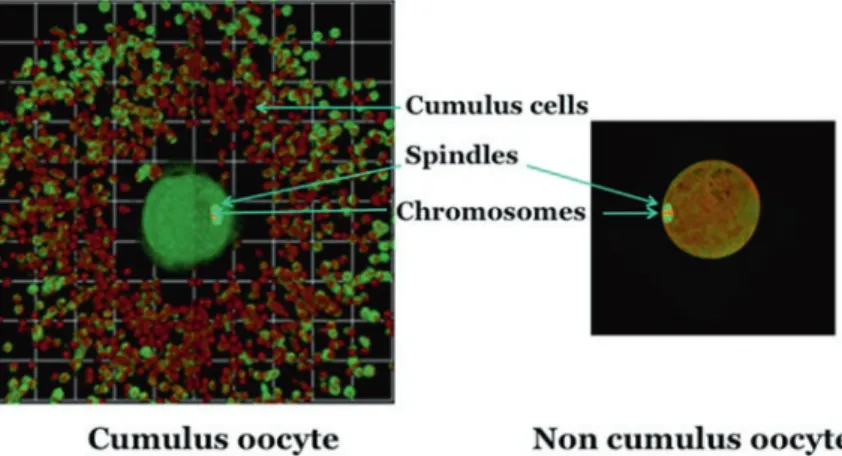
Nitric oxide is produced enzymatically mainly by three different isoforms of NO synthase (NOS): neuronal (nNOS), inducible (iNOS) and endothelial (eNOS) by converting L-arginine (L-Arg), molecular oxygen (O2) and NADPH into NO and citrulline [10, 12]. Goud PT, Goud AP, Diamond MP, Gonik B, Abu-Soud HM (2008) Nitric oxide extends the oocyte time window for optimal fertilization.
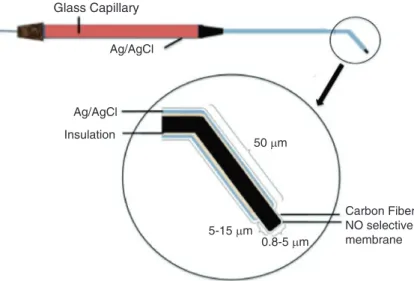
Emrah Eroglu, Helmut Bischof, Suphachai Charoensin,
Binding of NO• to the non-heme iron(II) center directly quenches the fluorescence intensity of geNOps in a highly sensitive, selective and concentration-dependent manner [ 20 , 35 ]. Correctly performing the incubation procedure with the iron(II) booster solution provides full activation of the NO• sensors.
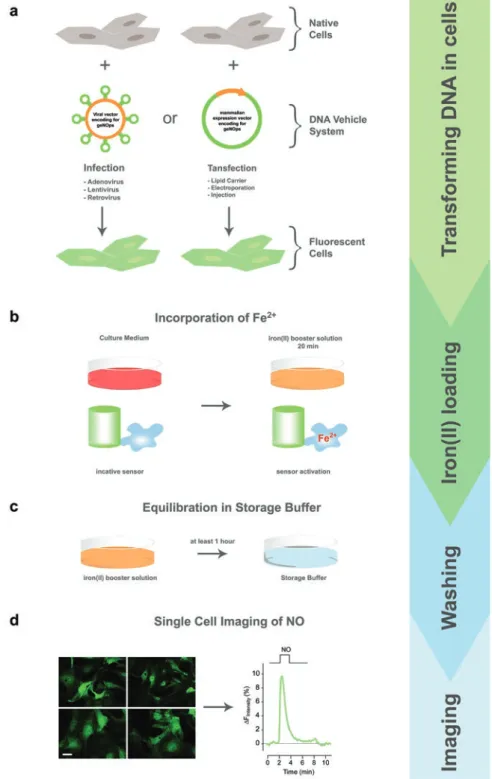
Acknowledgments
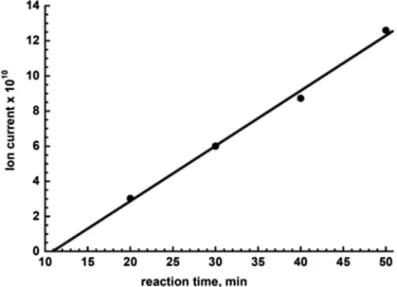
Nitric oxide synthases (NOS, EC catalyze the oxygen- and NADPH-dependent oxidation of L-arginine to citrulline and NO via two monooxygenation reactions [2]. Rinse the reaction vessel and establish an m/z 30 ion current baseline with fresh 1400 μl of calibration solution.
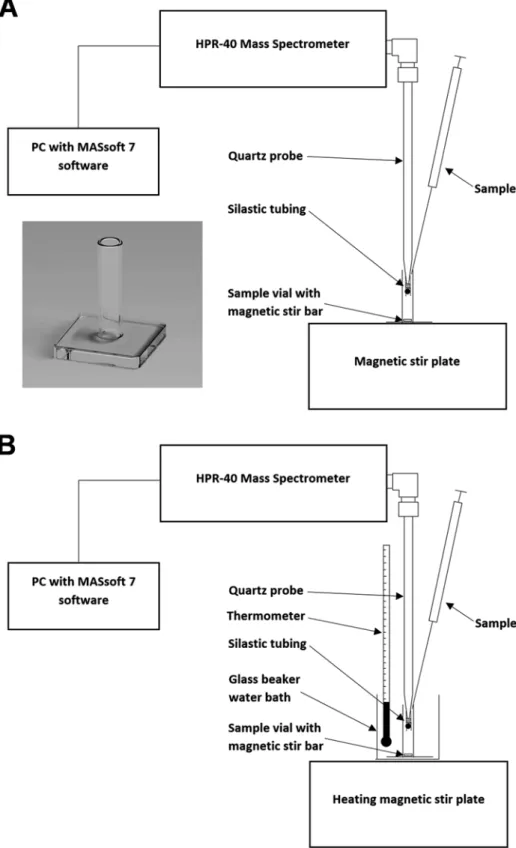
Paula Mariela González and Susana Puntarulo
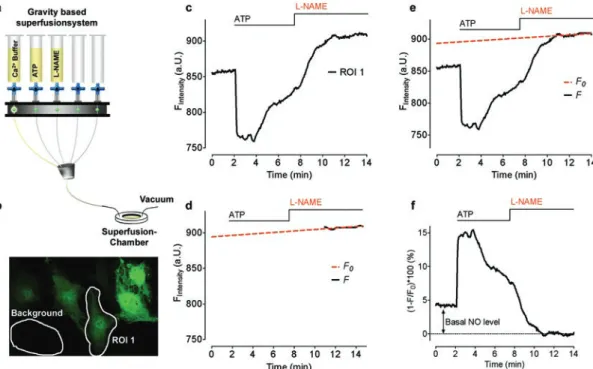
Homogenize the isolated DG with a glass nail cloth mill in 1:1.5 (w/v) spin trap solution. Use the increasing height of the first peak to estimate the rate of NO generation. Estimate NOS-like activity from the slope of the line as shown in Fig.
Inset: Calculating the NO generation rate requires determining the slope of the graph obtained over time. The concentration and incubation time of the NOS-like inhibitor (l-NAME) with the tissues should be tested before performing the experiments. Depending on the level of l-Arg present in the sample, the effectiveness of the inhibitors may change.
NO Modifications
TMT- Switch Assay
For acetone precipitation, incubate the samples with 4 x volume of pre-cooled acetone for 20 min at -20 °C and gently wash the resulting pellets with an additional volume of the cold acetone (see Notes 7 and 14). Incubate the nitrocellulose membrane with the primary mouse anti-TMT antibody (1:2000) in the blocking solution at room temperature for 1 h. The sample is then incubated with 8 mM IAA for 30 minutes at room temperature in the dark.
Resuspend it with TBS up to 2× volume of the immobilized anti-TMT™ antibody resin. Collect quantitative values for the TMT reporter ions with the Libra module of TPP using a custom state file [15]. In the absence of reduction/alkylation steps prior to trypsin digestion, resolubilization of the pellet can be aided by vortexing.

Lili Zhang, Pengpeng Shang, Changbao Chen, Jie Zhou, and Shuhua Zhu
FT-SPR Experiments
Rinse successively with approximately 10 mL of deionized water, anhydrous ethanol, and double-distilled water (see Note 3). Place the slide in 20 ml of 1.0 mM 11-mercaptodecanoic acid in ethanol, and incubate overnight at 4 °C. To activate the carboxyl group, place the slide in 20 ml of EDC/NHS solution for 20 min.
Place the chip in 20 mL of biotin-HPDP solution at room temperature overnight, thereby forming a surface-grafted film on the chip. According to the method of Jaffrey [6], block free protein thiols with 4 mL of blocking buffer and mix for 20 min in a water bath at 50 °C. To remove unreacted MMTS, incubate the protein mixture with 10 mL of pre-cooled acetone for 20 min at -20 °C.
FT-SPR Experiments
Adjust the angle of incidence to 9000 cm−1 to be in the best sensitivity range of the detector. Pump different analytes into the flow cell until the wavenumber shifts reach stable values, which can be recorded in real time with the FT-SPR (see Notes 9 and 10). During the course of the experiment, it is best not to stop the peristaltic pump that may be used to pump the analyte solution from a reservoir into the flow cell.
If there is little residual sample in the reservoir, slowly pour the buffer into the reservoir, rinse it for a while and then pour the next sample into the reservoir. Wang S, Circu ML, Zhou H, Figeys D, Aw TY, Feng J (2011) Highly sensitive detection of S-nitrosylated proteins by capillary gel electrophoresis with laser-induced fluorescence. MB (2017) Graphene-coated fiber optic surface plasmon resonance biosensor for the detection of DNA hybridization: simulation analysis.
Dimitrios Tsikas and Erik Hanff
Inorganic nitrite is reabsorbed in the proximal tubule of the nephron by anion transporters and by renal carbonic anhydrase (CA) [15]. The autosampler of the LC-MS/MS system should be cooled (e.g. at 4 °C) to minimize thermal decomposition of GSNO until injection. 5 Scheme of the main steps of the UPLC-MS/MS method for the quantitative determination of GSNO in human plasma using S-[15N]nitroso-glutathione (GS15NO) as an internal standard.
6 Results of a study on GSNO in the thermostatic autosampler (4 °C) of the LC-MS/MS apparatus model XEVO TQ MS. QC samples are used to determine the precision (RSD, %) and precision (recovery, %) of the method. Mansoor MA, Svardal AM, Ueland PM (1992) Determination of the in vivo redox status of cysteine, cysteinylglycine, homocysteine and glutathione in human plasma.

Sébastien Aimé, Siham Hichami, David Wendehenne, and Olivier Lamotte
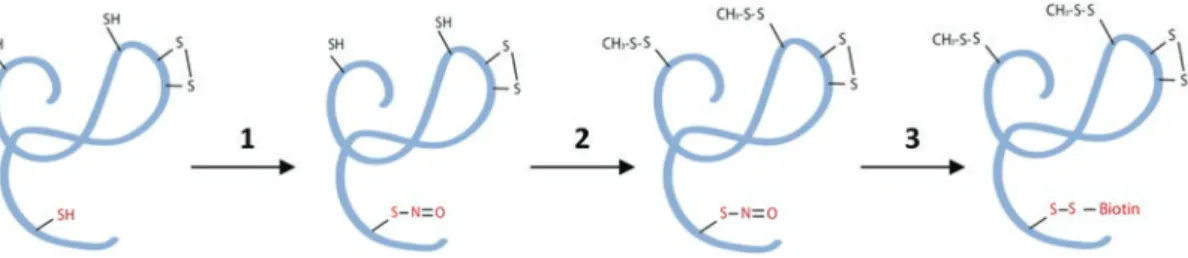
This progress has been made possible by the development of the Biotin-Switch technique (BST), a method dedicated to the identification of S-nitrosylated proteins in which the NO group of S-nitrosothiol is selectively replaced by biotin-HPDP [8. ]. Its S-nitrosylation inhibits most of the ATPase activity of the protein and causes conformational changes in the vicinity of Cys526. Cut a nitrocellulose membrane to gel size and 2 pieces of Whatman 7CHR chromatography paper.
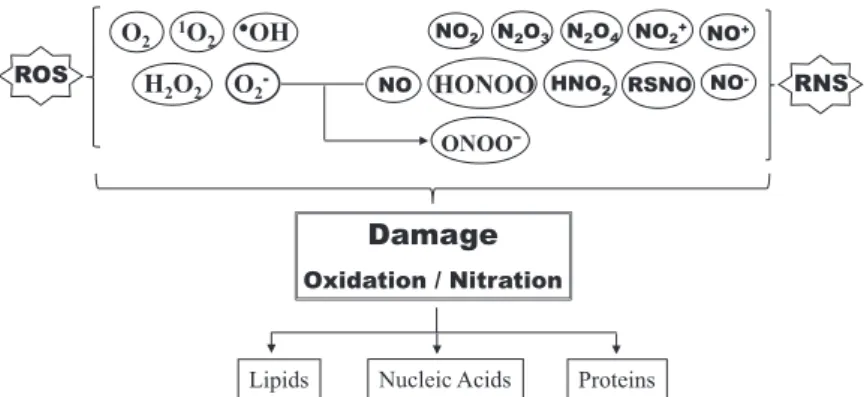
The percentage of acrylamide in the separating gel depends on the molecular size of the protein being separated. Consist in the addition of a nitro group (-NO2) to one of the two equivalent ortho-carbon atoms in the aromatic ring of tyrosine residues, leading to the formation of 3-nitrotyrosine. Meanwhile, the PVDF membrane is cut to the size of the resolving gel and it is activated by immersion in methanol (100%) for 10 s.
CLSM analysis of the distribution of tyrosine-nitrated proteins in seedling cotyledon cross-sections shows an increase in the rate of protein tyrosine nitration in the presence of salt stress (Fig. 2) [11]. The membrane must be rinsed repeatedly with distilled water to completely remove the Ponceau S stain.
Identification of NO-Sensitive Cysteine Residues Using Cysteine Mutants of Recombinant Proteins
Azam Shekariesfahlan and Christian Lindermayr
SDS–
Incubate the reaction at 25 °C overnight to recombine the entry clone into the destination vector pDEST™17, which represents a 6X His-tag in the N-terminal region of the produced proteins in E. Cover the column lid with a Parafilm strip carefully and incubate at 4 °C for 30 minutes with overhead mixing at 5 minute intervals. Take a 25 μL aliquot of the eluted sample and store it for analysis on a 12% self-cast gel (see subsection 3.11).
Detect the formation of the target protein by Coomassie staining of the gel (see Subheading 3.12) (see Figure 1) and the presence of a histidine tag on the recombinant protein by western blotting using a mouse-produced polyhistidine monoclonal antibody (see Subheading 3.13) (see Figure 1). Fill the remaining gap with isopropanol so that the top of the gel is horizontal. Pipet the stacking gel on top of the separating gel until overflow occurs.
SDS Polyacrylamide Gel
The Bradford assay is based on the binding of Coomassie Blue G250 dye to lysine and arginine residues in proteins, resulting in a shift of the maximum absorbance of this dye to 595 nm (see Note 14). To evaluate the concentration of protein extracts and recombinant proteins; perform sample preparation, incubation, and measurement in a similar manner and apply the standard curve equation to the data. Remove the glass plates from the casting frames and place them in the liquid module inside the electrophoresis tank.
Run the electrophoresis until the bromophenol blue dye front reaches the bottom of the gel (this takes about 1 hour). To check S-nitrosation of wild-type and mutant recombinant proteins, take 5% samples, separate proteins by 12%. So, download the required cDNA sequences of the genes of interest from The Arabidopsis Information Resource (TAIR) or the National Center for Biotechnology Information (NCBI) database.