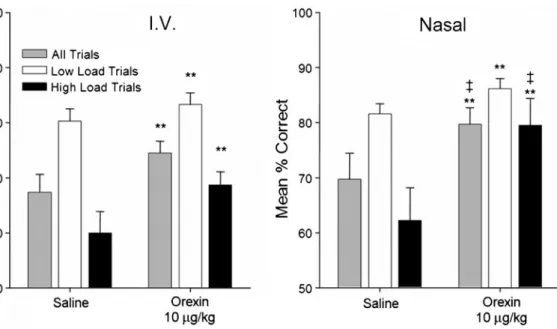
For example, narcolepsy appears to be caused by a selective loss of hypocretin-producing neurons, yet the target antigens are expressed by many non-hypocretin neurons. It is possible that hypocretin/orexin stabilizes the neural pathways that regulate the sleep/wake transition; so a loss of hypocretin neurons would destabilize this mechanism, leading to frequent transitions between wakefulness and sleep.
The Diagnosis of Narcolepsy and the Assessment of Fitness to Drive
Treatment of Narcolepsy
Contributors
Fronczek, PhD
Sleep Disorders Center and Department of Neurology, Inselspital, University Hospital Bern and University of Bern,. Department of Neurology, Donders Institute for Neuroscience, Radboud University Medical Center Nijmegen, Nijmegen, The Netherlands.
Etiology of Narcolepsy
Low levels of the neuropeptide hypocretin-1 (hcrt-1, also called orexin-A) are found in the cerebrospinal fluid (CSF) of most people with narcolepsy with cataplexy and in some without cataplexy [1-6]. As a result, narcolepsy with cataplexy and narcolepsy without cataplexy have been separated in the latest revision of the International Classification of Sleep Disorders (ICSD).
Prevalence Studies
Etiology and Genetics of Human Narcolepsy
The prevalence of narcolepsy without cataplexy is largely unknown, as a proper population study would require MSLT of all subjects. The idea is also supported by the slightly increased frequency of HLA observed in subjects with narcolepsy without cataplexy with normal CSF hcrt [21], although in this case it is difficult to rule out that some patients were diagnosed after HLA positivity, thus creating a bias .
Twin Studies and Environmental Factors in Narcolepsy
It is likely that registry-based estimates of the prevalence of diagnosed cases, while population-based epidemiological studies that do not rule out other confounding factors, overestimate the true population prevalence of narcolepsy without cataplexy, explaining the 300-fold range. . It is therefore possible that some subjects without cataplexy may have less pronounced hcrt cell loss, as evidenced by average (110–200 pg/ml) or normal CSF hcrt-1 [19, 20].
Familial Aspects of Human Narcolepsy
Indeed, HLA DQB1*0602 positivity was significantly lower in familial cases (70%) than that in sporadic narcolepsy cases (87%), especially in families with more than two affected individuals (56%). This suggests that in these cases, multiple DQB1*0602 haplotypes (if not all DQB1*0602 alleles in the general population) in the family were equally susceptible to narcolepsy [10].
Hypocretin (Orexin) Deficiency and Human Narcolepsy–Cataplexy
HLA typing data in these non-DQB1*0602 families did not support the concept of association with other HLA subtypes [10]. This strongly suggests that non-HLA genetic factors may be involved in a subset of non-HLA-DQB1*0602 cases.
HLA-DR2, Narcolepsy, and Autoimmunity
DQB1*0602 and DQA1*0102 Are the Main HLA Narcolepsy
A number of other DR-DQ haplotypes in the population carry DQA1*0102 without DQB1*0602 and these do not predispose to narcolepsy [51]. Recent findings in families and in unrelated cases also suggest that most, if not all, of the DQB1*0602/DQA1*0102 alleles present in the population equally predispose to narcolepsy.

Usefulness of HLA Typing in Clinical Practice
9 Etiology and genetics of human narcolepsy. DQA1*0102) rather than an unknown genetic factor in the region predisposes to narcolepsy.
Genetic Factors Other than HLA
Linkage analysis in HLA-DQB1*0602-positive Japanese families has suggested the existence of a susceptibility gene on 4q13-23 [57]. Recently, a larger GWAS compared 807 Caucasian subjects with narcolepsy-cataplexy/hcrt deficiency with 1,074 HLA-DQB1*0602-positive controls using the Affymetrix 500K and 6.0 platforms (Fig. 2).

CSF Hypocretin-1 as a Diagnostic Tool for Narcolepsy
Although the diagnostic value of hcrt-1 in the low CSF (<110 pg/ml) is well established, it is interesting to note that healthy control levels have been shown to be above 200 pg/ml [4]. However, such values should be interpreted with caution, as in a large series of individuals with various neurological disorders, we found that up to 15% had CSF hcrt-1 values within this intermediate range; most of these patients had severe brain pathology, especially head trauma, encephalitis, and subarachnoid hemorrhage [37].
Secondary Narcolepsy
CSF hcrt-1 has been measured in cases of Niemann-Pick disease type C, a condition in which oculomotor symptoms are frequent, and intermediate levels have been found in some cases with cataplexy. We have examined CSF hcrt-1 levels in such cases and have found that some, but not all, of these patients have very low CSF hcrt-1 levels (<110 pg/ml), suggesting hcrt -lack.
Conclusion and Perspectives
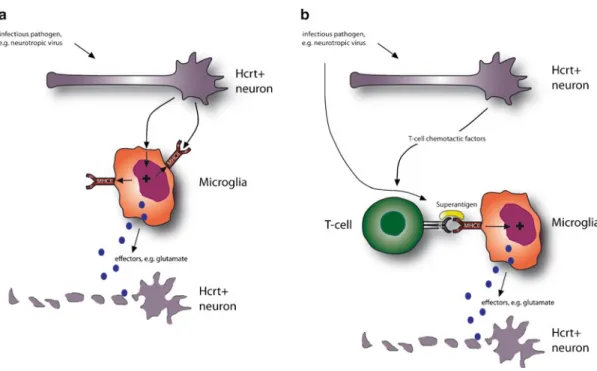
Interestingly, superantigens are known to be involved in mediating toxic shock, and DRB1*1501-DQB1*0602 is protective against septic shock caused by Streptococcus pyogenes. The possibility that the unique narcolepsy DQB1*0602-TCR interactions occur at the brain level, independent of autoimmunity, is, however, still possible, as TCRB localization has been shown to be expressed in the brain [85].
Medical exposure in young people and the frequency of narcolepsy with cataplexy: a population-based case-control study in genetically predisposed people. Complex HLA-DR and -DQ interactions confer the risk of narcolepsy-cataplexy in three ethnic groups.
Narcolepsy: Autoimmunity or Secondary to Infection?
TNF and Its Receptors: Essential in the Pathogenesis of Narcolepsy?
When discussing the potential role of TNF in sleep disorders, studies on TNF overexpression in the CNS are important. In addition to its role in inflammation and its response to infection, much interest has been gained in the role of TNF in sleep regulation (see review [4]).
Anti-self T Lymphocytes
As noted above, genetic polymorphism in the Tnf promoter may also affect serum TNF concentrations. The T cell allele of the C-857T polymorphism was strongly associated in the subset of DRB1*15/16 negative patients (HLA-DR2 type) [ 16 ].
Our laboratory has shown that subcutaneous infusion of TNF impairs the locomotor activity of mice and reduces the expression of clock genes in the liver. One of the clock gene polymorphisms was found to be associated with sleep genotypes [23].
Microglial Cells: Key Factors in Narcolepsy?
The data revealed an association between narcolepsy and polymorphisms in the T-cell receptor alpha [TRAa] locus. This is remarkable, as the upregulation of HLA-DR and the transition from quiescent branched microglia are hallmarks of immune-mediated inflammation in the CNS.
Are Anti-neuronal Antibodies Involved?
In the context of the aforementioned discussion on dysregulated TNF expression, it is interesting that in situ hybridization of TNF did not produce significant signal in control and narcoleptic tissue [29]. Histological analysis and HLA-DR staining in four adult and three juvenile narcoleptic Dobermans did not reveal lymphocyte infiltration or inflammation in the CNS in canine narcolepsy.
Conclusion and Hypothesis
Furthermore, in another recent study in narcolepsy patients, autoantibodies against Tribbles homolog 2 (Trib2), which is expressed by hypocretin neurons and by many other neurons, were detected in a subset of patients [40]. Haplotype analyzes with the human leukocyte antigen and tumor necrosis factor alpha genes in narcolepsy families.
Introduction
Is Narcolepsy a Neurodegenerative Disorder?
Accumulation of aberrant or misfolded proteins, ubiquitin-proteasome dysfunction, excitotoxic damage, autoimmune disease, oxidative and nitrosative stress, mitochondrial damage, synaptic failure, altered metal homeostasis, and failure of axonal and dendritic transport are events that have been described in progressive studies. neurodegenerative disease. Here we discuss the arguments for and against the idea that NC could be a neurodegenerative disease, considering only the NC/HLA+/Hcrt−.
Tracking Signs of
Neurodegeneration In Vivo
Are Hypocretin Neurons Missing in Patients with Narcolepsy-Cataplexy?
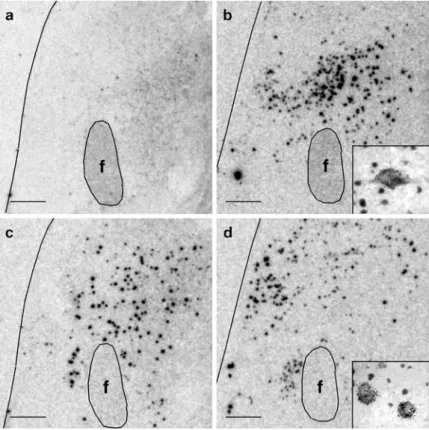
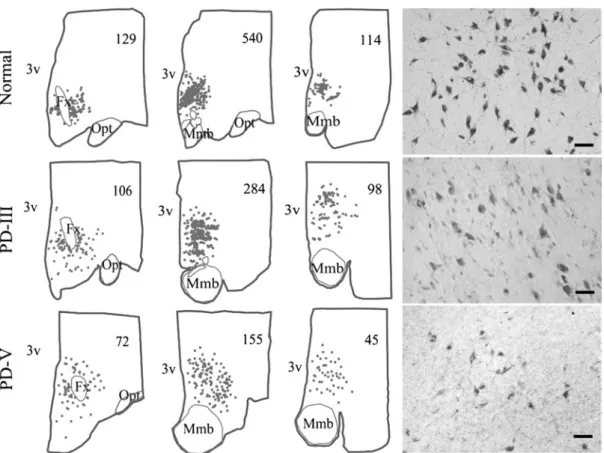
No change in NARP staining was observed in brain regions lacking hypocretin neurons, such as the paraventricular and supraoptic nuclei of the hypothalamus. These are the strongest arguments to date to support the hypothesis that NC is a neurodegenerative disease that damages hypocretin neurons.

Looking for Signs of Inflammation or Gliosis
However, an increase in GFAP staining density was demonstrated in the hypocretin-hypothalamic area (n = 3) by Thannickal and colleagues [ 25 , 26 ]. Furthermore, an increase in the number of GFAP-stained astrocytes was observed in the locus coeruleus and raphe nuclei (n = 4), but not in the dorsomedial thalamic nucleus (n = 3) in NC versus control brain samples [25, 26].

Testing the Neurodegenerative Hypothesis: Do the Remaining
Progression of the Disease
Conclusion
The role of cerebrospinal fluid hypocretin measurement in the diagnosis of narcolepsy and other hypersomnias. A mutation in a case of early-onset narcolepsy and a generalized absence of hypocretin peptides in human narcoleptic brains.
Neurochemistry of Narcolepsy
The Roles of Hypocretin/Orexin in Narcolepsy, Parkinson’s Disease,
Therefore, increased inhibition by glycine and GABA [9, 10] and decreased facilitation by norepinephrine contribute to the loss of muscle tone in cataplexy and ReM sleep. Loss of consciousness is associated with disruption of activity in serotonergic and histaminergic neurons in slow wave sleep (SWS) and ReM sleep.
Human Narcolepsy with Cataplexy and Hypocretin
It has been shown that the loss of norepinephrine signaling is more important in causing the loss of muscle tone in ReM sleep than the increased release of GABA and glycine [21–23] . These studies elucidate the mechanisms responsible for muscle tone suppression in both normal ReM sleep and cataplexy.
Parkinson’s Disease and hcrt
The most likely cause of such gliosis and specific loss of hcrt cells is an autoimmune process [34-36]. We found that the whole brain had gliosis and a loss of hcrt cells in the posterior hypothalamus, but normal levels of glial cells and hcrt in more anterior regions and that the partial brain showed a similarly intact rostral region [ 30 ].
Normal Role of hcrt
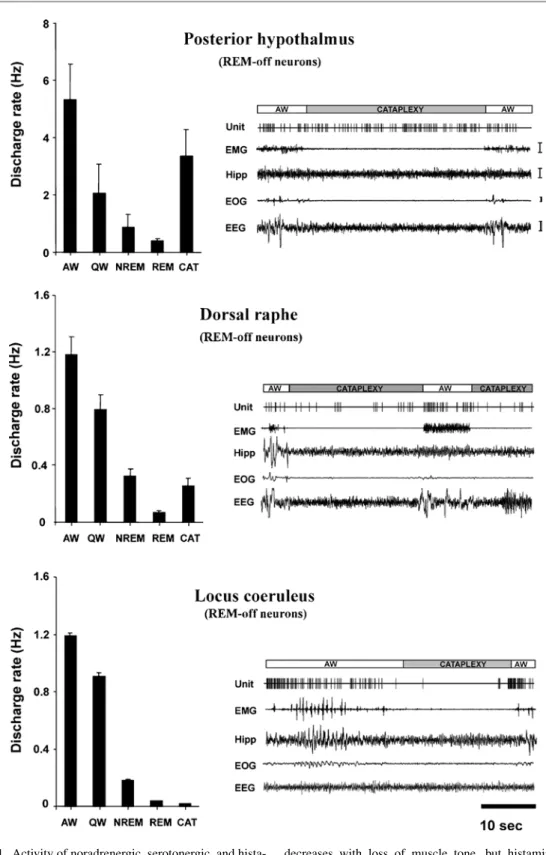
However, we found that these cells can be completely inactive even during periods of high arousal, as shown by eeG and behavioral criteria (Fig. 4). Other systems must mediate arousal in aversive situations, as hcrt cells are silent during this time.
Summary
Most human narcolepsy with cataplexy is caused by loss of hcrt cells, and at least one case of narcolepsy without cataplexy was characterized by loss of posterior hypothalamic hcrt cells. Medial mesopontine unit activity during cataplexy and sleep-wake states in the narcoleptic dog.
Histamine in Narcolepsy
The chapter also discusses the development of histamine-mimetic compounds for the treatment of narcolepsy and other hypersomnias in humans.
Physiology
2 Fluctuation of extracellular hypocretin and histamine levels in relation to changes in the amount of wakefulness. Two microdialysis probes were implanted in the brain (lateral hypothalamus for hypocretin measures and anterior pre-optic area for histamine measures) of each rat.

Pathophysiology Decreased Histaminergic
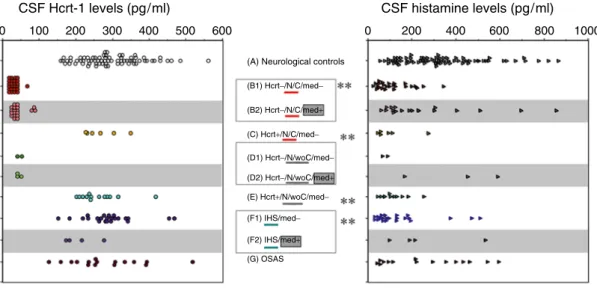
Thus, altered histamine levels may play critical roles in the phenotype expression of the narcoleptic mice. Unmedicated subjects tended to have low CSF histamine levels in hypocretin-deficient narcolepsy without cataplexy (D pg/ml) (adapted from [39]).
Pharmacology
To our surprise, we did not observe any changes in sleep parameters after administration of a-FMH [31]. These results (in contrast to those obtained in rats) may suggest that there are likely to be important species differences in the roles of histamine in the control of sleep and wakefulness.
Discussion
Innervation of histaminergic tuberomammillary neurons by GABAergic and galaninergic neurons in the rat ventrolateral preoptic nucleus. The connections between the septum-diagonal band complex and histaminergic neurons in the posterior hypothalamus of the rat.
Dopaminergic Substrates Underlying Hypersomnia, Sleepiness, and REM
Physiological Effects of Dopamine
Anatomy of Mesotelencephalic Dopamine Neurons in Relation
Indirect effects of midbrain dopamine neurons on thalamocortical arousal state may also occur via output pathways to the ventral forebrain, perifornical hypocretin/orexin, and adjacent hypothalamic neurons ( Fig. 2 ) and LC (among others). Dopaminergic innervation of the LC originates from hypothalamic cell groups A11 and A13 and midbrain VTA-A10 and has.
Circadian, Homeostatic, and State Influences upon Dopamine
The physiological and behavioral effects of these interactions are incompletely defined and include putative direct and indirect, excitatory and inhibitory effects on magnocellular cholinergic and perifornical hypocretin/orexin neurons. Although there is experimental in vivo evidence from the preoptic area that dopamine can act via adrenoreceptors, the concentrations required appear to be supraphysiological and are unlikely to occur in nature [7].
Signaling
Sleep and REM sleep deprivation also lead to strong changes in dopamine availability and the density and affinity of its various receptors in mesotelencephalic circuits. In the rhesus monkey putamen and limbic regions of the human brain, apparent dopamine fluctuations related to specific sleep stages are undetectable by microdialysis.

Dopamine’s Modulation of Wake–Sleep State
Dopamine by way of D2 receptor modulation of nonvisual photic responses in the retina is necessary for light to disrupt circadian resting activity rhythms (i.e., light “masking” of endogenous circadian rhythms) [21]. Wakefulness attributable to dysfunctional D3 receptors may alternatively be an epi-phenomenon of an increased response to novelty, or decrease in the gating of spinal sensorimotor and autonomic excitability.
Relevance of Dopamine to Arousal Disorders and Hypersomnia
Where you least expect it: dopamine in the pons aion of sleep and REM sleep. Prominent bursting of dopaminergic neurons in the ventral tegmental area during paradoxical sleep.
![Fig. 4 Rat hypnogram observed 1 week after bilateral lesioning of dopaminergic ventral tegmental pathways made by direct injection of the dopaminergic toxin 6-hydroxydopamine into the nucleus accumbens [29]](https://thumb-ap.123doks.com/thumbv2/azdoknet/10579369.0/84.756.81.678.94.307/hypnogram-bilateral-lesioning-dopaminergic-tegmental-injection-dopaminergic-hydroxydopamine.webp)
The Serotoninergic System in Sleep and Narcolepsy
Serotonin and Sleep–Wake Regulation
The DRN contains about half of all 5-HT neurons in the rat, and the caudate nuclei contain many fewer 5-HT neurons. Most putative 5-HT neurons in the DRN are awake active, slow their firing during NREM sleep, and are silent during REM sleep [ 34 – 39 ].

Interactions of the 5-HT and Orexin Systems
Effects of 5-HT on Orexin Neurons Orexin neurons are innervated by multiple serotonergic terminals. Perhaps, negative feedback from the 5-HT system inhibits orexin neurons to avoid excessively high levels of arousal.
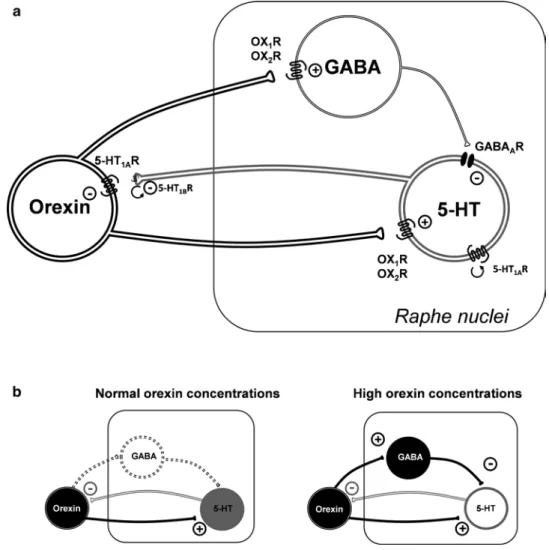
5-HT and Narcolepsy
After a period of sleep, NE excites the orexin neurons, but after 2 hours of waking, NE inhibits the orexin neurons [84]. NE also inhibits the orexin neurons of mice killed during the day, just after the nocturnal active period [85].
Conclusions
Sleep Homeostasis, Adenosine, Caffeine, and Narcolepsy
Sleep Homeostasis Can Be Reliably Tracked in the
The Neuromodulator Adenosine Is Involved in Sleep Homeostasis
The data show that genetic variation in ADA contributes to interindividual differences in subjective and objective markers of sleep homeostasis in humans. Studies are currently underway to investigate the consequences of sleep deprivation on awake performance, sleepiness, sleep, and sleep EEG in healthy individuals with different ADA genotypes.
Adenosine A 1 and A 2A Receptors May Mediate Effects of Adenosine
Recent studies in rodents, including experiments in knockout mice, suggest that A2A receptors also contribute to the effects of adenosine on sleep. Direct activation of sleep-inducing VLPO neurons by stimulation of A2A receptors may underlie this effect [21].
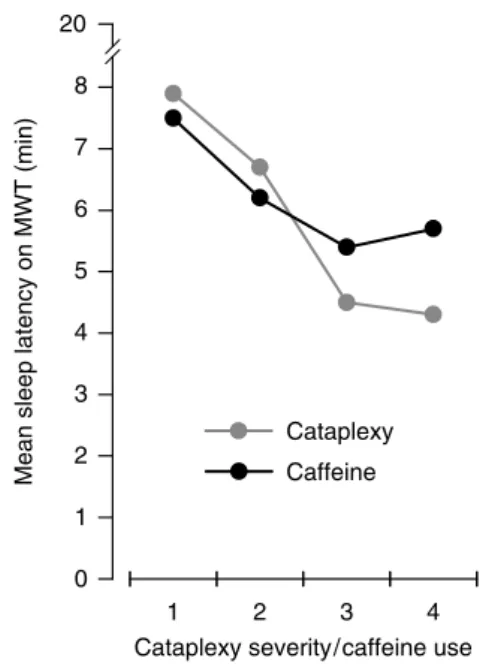
Caffeine Attenuates Markers of Sleep Homeostasis in Sleep
Together, pharmacological blockade of adenosine A1 and A2A receptors with caffeine interferes with the wakefulness-induced rise in sleep pressure. Nevertheless, the state-specific changes in predefined EEG markers of sleep homeostasis in sleep and wakefulness are highly unlikely to reflect a nonspecific action of caffeine on EEG generation.
Dysregulation of Sleep Homeostasis Involving Adenosinergic
Thus, the presumed residual level of caffeine cannot immediately explain the delayed decrease in SWA during recovery sleep. Caution is needed when comparing physiological mechanisms of sleep and wakefulness regulation with pharmacological interventions.
Mechanisms in Narcolepsy?
In both groups and in baseline and recovery nights, SWA was highest in the first nonREM sleep episode and decreased over the course of sleep. Although genetic studies in dogs suggest that narcolepsy may be transmitted as an autosomal recessive trait caused by a mutation in the canine hypocretin (orexin) receptor 2 gene, there are no pathogenic mutations or polymorphisms in the genes that secrete hypocretin and its two receptors do not encode (hypocretin-1 and hypocretin-2) have been identified to date in human patients with narcolepsy [31].
Summary and Conclusions
A1 receptor and adenosinergic homeostatic regulation of sleep-wake: effects of antisense to the A1 receptor in the cholinergic basal forebrain. The endogenous somogen adenosine excites a subset of sleep-promoting neurons via A2A receptors in the ventrolateral preoptic nucleus.
Sleep Induction by PGD 2
Thus, these scientists became the pioneers of sleep research, demonstrating the existence of endogenous sleep-promoting substances. Among them, prostaglandin (PG) D2 (Fig. 1) is now recognized as the most potent endogenous sleep-promoting substance; and its mechanism of action to induce sleep is best characterized.
3] i.e., microinjection of nanomolar amounts of PGD2 into the rat brain increased non-rapid eye movement (non-REM, NREM) and REM sleep. Essentially the same sleep induction was demonstrated in the rhesus monkey Macaca mulatta during i.c.v.
PGDS and PGD 2 Receptors in the CNS
Using a sleep bioassay system based on electroencephalogram (EEG) and electromyogram (EMG) recordings during continuous intracerebroventricular (i.c.v.) infusion of drugs in freely moving rats, the somnogenic activity of PGD2 was demonstrated to be dose-dependent and time-dependent [4]. In addition, PGD2 concentrations were selectively and time-dependently elevated in the cerebrospinal fluid of patients with African sleeping sickness caused by Trypanosoma infection [9].
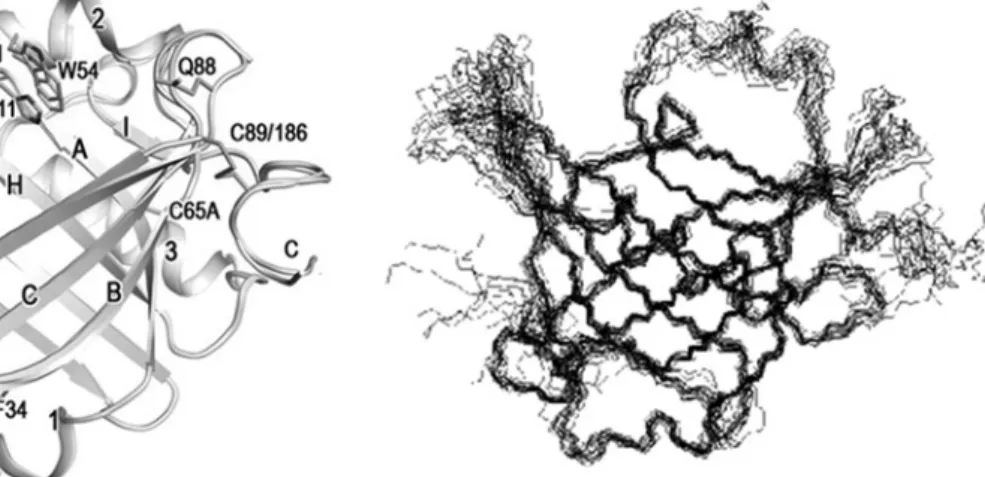
The Molecular Mechanisms of PGD 2 -Induced Sleep
In contrast, the number of Fos-positive neurons was markedly decreased in the tuberomammillary nucleus (TMN) of the posterior hypothalamus. The flexibility of the EF loop is clearly seen in the superposition of 15 lowest energy backbone conformers of L-PGDS determined by NMR [25].
Involvement of the Histaminergic Neurons in Sleep–Wake Regulation
These results indicate that orexin is a potent arousal agent that acts on its receptor in the TMN and that the excitatory effect of orexin A depends on histaminergic neurotransmission mediated by H1 receptors. These results suggest that endogenous adenosine in the TMN inhibits the histaminergic system via A1 receptors to promote NREM sleep.
Sleep Abnormalities in Mice with Manipulations of the Genes Coding
Adenosine deaminase, an enzyme that catabolizes adenosine to inosine, is strongly expressed in TMN neurons, and the TMN is also enriched in A1 adenosine receptors. Bilateral injection of adenosine or an adenosine deaminase inhibitor, coformycin, into the rat TMN also increased the amount of NREM sleep.
PGD2 content in the brains of wild-type mice 2 hours after an intraperitoneal injection of SeCl4 at a dose of 1.25-5 mg/kg body weight during the light period (when mice sleep normally). The H-PGDS KO mice showed strong NREM and REM sleep induction and suppression of wakefulness during the following dark period, similar to the wild-type mice.
Contribution of Adenosine to Physiological Sleep
In the case of 1.25 mg/kg SeCl4, the sleep-wake pattern was almost identical before and after the injection, similar to the pattern of PGD2 content in the brain. Cellular localization of lipocalin-type prostaglandin D synthase (beta-trace) in the central nervous system of the adult rat.

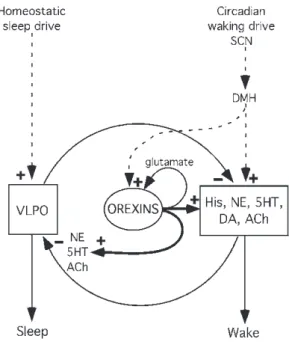
The Role of the Hypocretins
The Neurobiology of Sleep–Wake Systems: An Overview
Neuronal Networks Responsible for Sleep Onset and Maintenance
These studies showed that the number of Fos + neurons in the VLPO and MnPn was positively related to the amount of sleep and sleep consolidation during the last hour before sacrifice. Indeed, infusion of an A2AR agonist into the subarachnoid space rostral to the VLPO increases SWS and induces Fos expression in VLPO neurons [ 19 ] ( Fig. 2 ).
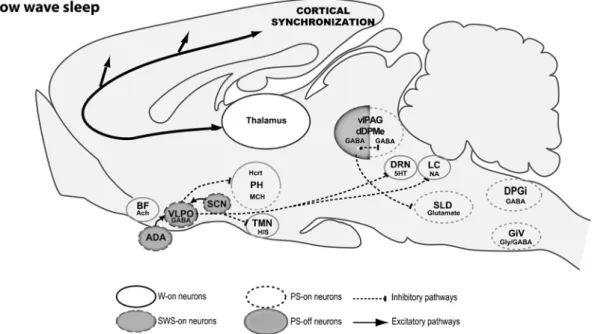
Neuronal Network Responsible for Paradoxical Sleep
We propose that PS induction results from the activation of glutamatergic PS-na neurons in the SLD. Localization of neurons active during paradoxical (REM) sleep and projection to noradrenergic neurons of the rat locus coeruleus.

The Hypocretins/Orexins
Master Regulators of Arousal and Hyperarousal
The Hcrts and their Receptors
In vitro electrophysiology studies show that several neurotransmitters/neuromodulators excite Hcrt neurons (including corticotropin-releasing factor, ghrelin, neurotensin, oxytocin, and vasopressin) or inhibit Hcrt neurons (including serotonin, norapamine, neuronetin, noradrenaline, and leptin) [22]. These anatomical and electrophysiological studies suggest that Hcrt neurons integrate a variety of homeostatic signals from the central nervous system and the periphery, and project to multiple brain regions, many of which express other neuromodulators and are capable of regulating various physiological functions. and behaviors.
Hcrts and Wakefulness
Finally, electrophysiological studies suggest that Hcrt neurons are actually more active during wakefulness and arousal-related behaviors compared to sleep. Hcrt neurons are activated by excitatory neurotransmitters, including glutamate [ 44 , 45 ], corticotropin-releasing factor [ 46 ], ATP [ 47 ], norepinephrine [ 48 ], and carbachol [ 48 ].
Hcrts in the Context of Allostasis
Furthermore, the number of c-Fos-positive Hcrt neurons induced by calorie restriction strongly correlates with the improvement on the social interaction test [61]. This suggests that Hcrt neurons mediate an allostatic generalized stress response to caloric restriction, allowing an animal to overcome maladaptive depressive symptoms induced by chronic stress.
Stress
Similarly, although Hcrts do not necessarily stimulate food intake under normal conditions, in situations of caloric restriction, Hcrts are required for adaptive increases in food anticipatory behavior [62]. This study further demonstrates that Hcrt neurons mediate allostatic changes in behavior, in this case ensuring that animals will be alert and motivated to obtain food during the limited times it is available.
Addiction
While more research is needed to understand the functions of Hcrts in various environmental challenges, recent work has illuminated important roles for Hcrts in arousal-related behaviors, including brain reward and motivation, increased arousal following acute stressors, behavioral sexual. , and attention.
Hcrts in Other Arousal-Related Behaviors
New Methods to Study the Hcrt System
Peyron C, Tighe DK, van den Pol AN, de Lecea L, Heller HC, Sutcliffe JG, et al. Neuronal activity of arousal-active orexin and non-orexin neurons during awake sleep states in the rat.
Optogenetic Probing
The Hcrt System as a Modulator of the Sleep–Wake Cycle
Correlation Studies Between Hcrt Neural Activities and Behavioral
Gain-of-Function Studies
Local injections of Hcrt-1 into cholinoceptive neurons of the pons (nucleus pontis oralis) induced arousal and suppressed NREM sleep and REM sleep, while directly inhibiting REM sleep when injected into the ventral part of the NPO. The injected pharmacological compound remains active in the brain for minutes to hours before clearing and can act on various types of cells near the injection site.
Lack-of-Function Studies
Narcoleptic patients with cataplexy have a marked reduction in prepro hcrt gene transcripts in the hypothalamus and barely detectable levels of Hcrt peptides in the cerebrospinal fluid [7]. Doberman narcoleptic dogs with a mutation in Hcrt-R2 and all transgenic rodents with either a deletion of the Hcrt gene, the Hcrt-R2 gene, or Hcrt cells exhibit behavioral arrest similar to the cataplexy characteristic of narcolepsy [7].
Manipulating Hcrt Neuron Activity
1 Comparison between electrical/pharmacological activation or inhibition and optogenetic activation of Hcrt neurons in the lateral hypothalamus. We found that direct, unilateral deep brain optical stimulation of Hcrt neurons in the hypothalamus increases the probability of transition to wakefulness from NREM or REM sleep [ 49 ].
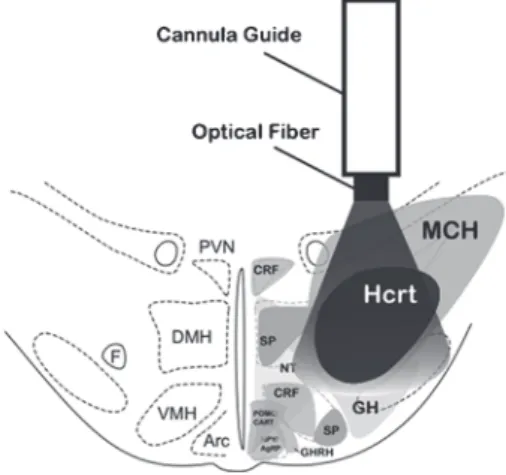
Perspectives
Effect of lateral hypothalamic lesion on sleep-wake pattern and EEG power spectra in rats. Blockade of orexin-1 receptors attenuates orexin-2 receptor antagonism-induced sleep promotion in rats.
Hypocretin/Orexin Receptor
Clues about the relevant neuronal populations come from the projection pattern of orexin neurons and the distribution of orexin receptors [1, 2], although it is likely that key targets remain to be identified. Neighboring structures, including the dorsal raphe (DR), the laterodorsal tegmental (LDT) nuclei, and the pedunculopontine tegmental (PPT) nuclei also receive significant numbers of orexin fibers and express both OX1R and OX2R receptors.
Functions in Mesopontine Systems Regulating Sleep, Arousal,
However, it remains completely unclear how the absence of signaling by these peptides causes the symptoms of narcolepsy. Either a simple loss of orexin excitatory action causes narcolepsy, or there are more subtle aspects of the loss of orexin signaling that result in plastic or trophic changes that result in the symptoms of narcolepsy and cataplexy.
These mesopontine regions and especially the noradrenergic neurons in the LC, the cholinergic neurons in the LDT/PPT and the serotonergic neurons in the DR have long been associated with regulation of global aspects of behavior, including arousal, motivation and reinforcement, the ultra-Dian alternation between non- REM and REM sleep and, in the canine model of narcolepsy, expression of cataplexy. Moreover, the wake-promoting and REM-sleep-inhibiting effects of orexins delivered ICV [4, 5] may be partially mediated by actions in these regions, as microinjections of orexins into the LC [6] and LDT [7] ].
Orexin has Two Distinct, Direct Actions on LDT, DR, and LC Neurons
In addition, orexin-A also increases intracellular Ca2+ in DR neurons—an effect mediated by the activation of voltage-gated Ca2+ channels by depolarization and by the enhancement of Ca2+. Collectively, these data indicate that orexins, like LDT and DR neurons, both directly excite LC neurons and increase Ca2+ influx via L-type Ca2+.
Which Receptors Are Necessary for the Postsynaptic Actions
Therefore, the absence of orexin signaling in the LDT, DR and LC in DKO mice is consistent with a role for this signaling in suppressing the symptoms of narcolepsy. Our findings are consistent with a role for direct orexin excitation in the LDT, DR, and LC in suppressing cataplexy since OX2R−/− mice rarely express cataplexy.
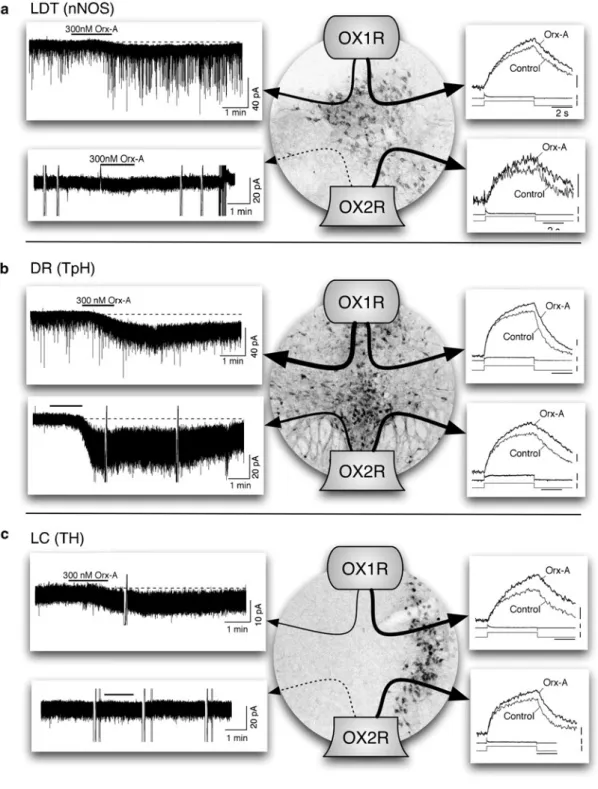
Orexin Receptor Knockout Mice Express Enhanced Cholinergic
In the absence of both orexin receptors, Orexin-A excitation and Ca2+ transient increases are abolished. We found that ChAT, VAChT, and CHT1 message levels were two- to fourfold higher in DKO samples.
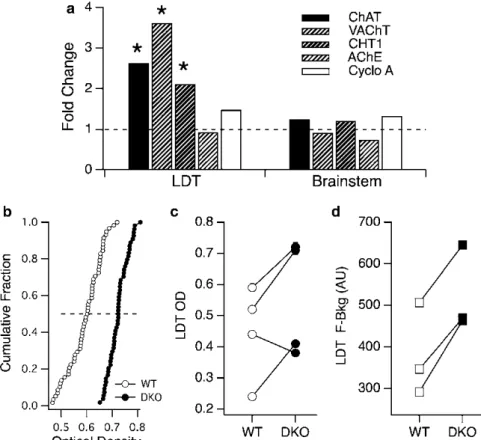
Can These Implied Changes in Brainstem Cholinergic Transmission
Among the various changes noted, a large increase in ACh release in the pons was observed during periods of cataplexy elicited by the food-evoked cataplexy test. Taken together, these data from narcoleptic DKO mice provide a plausible mechanism for key neurochemical events underlying cataplexy in the narcoleptic dogs.
Cataplexy in Mouse Models?
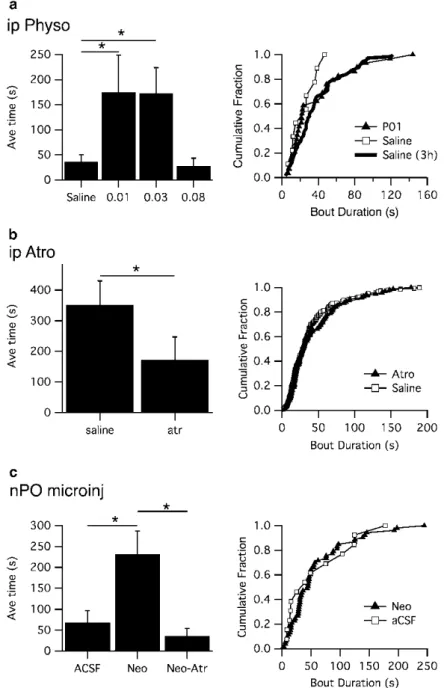
3 Cholinergic modulation of behavioral arrests in DKO mice. a) Low doses of physostigmine increased the time spent on behavioral arrests in the first hour of monitoring (left). Aminergic and cholinergic afferents to REM sleep induction areas of the pontine reticular formation in the rat.
Afferent Control of the Hypocretin/
Given the importance of the Hcrt system in such functions, it is of considerable interest to understand the control of the Hcrt neurons. While hypothalamic regions preferentially innervate the medial and perifornical parts of the Hcrt neuron field, projections from the brainstem are generally directed toward the lateral part of the field [11].
Orexin Neurons
For example, mice expressing the enhanced green fluorescent protein under the control of the hcrt/orexin promoter (orexin/EGFP mice) have facilitated in vitro studies of putative neurochemical inputs to the Hcrt neurons [ 14 , 15 ]. The following sections provide some examples that illustrate how such mouse strains have been used to advance our understanding of the Hcrt system.
Neuromodulatory Control of the Hcrt Neurons
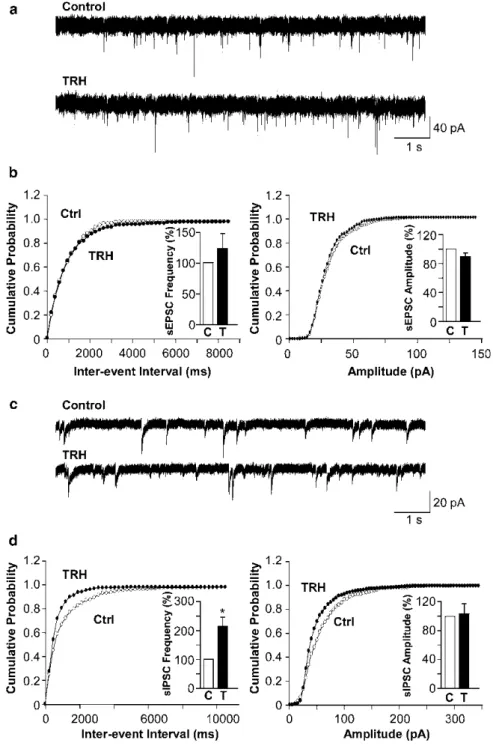
Finally, orexin/ataxin-3 mice, a model of human narcolepsy in which the Hcrt neurons degenerate, have facilitated in vivo studies of Hcrt neuron function [ 17 ]. In GABAB1flox/flox; orexin-Cre brain slices, the absence of GABAB receptors reduced the sensitivity of Hcrt neurons to both excitatory and inhibitory inputs [ 38 ].
Nociceptin/Orphanin FQ
This hypothesis was recently tested in conditional knockout mice in which the GABAB1 gene was specifically eliminated from Hcrt neurons (GABAB1flox/. flox; orexin-Cre mice). Thus, GABAB receptors on Hcrt neurons appear to be crucial for proper control of Hcrt tone during sleep and wakefulness and appear to play a role in consolidating sleep-wake periods by stabilizing sleep-wake transitions.
A Peptidergic Inhibitor of Hcrt Neuron Activity
N/OFQ also indirectly modulated the activity of Hcrt cells by inhibiting presynaptic glutamate release, the reverse of CRF effects on Hcrt neurons [37]. Moreover, while CRF facilitates SIA, N/OFQ reduces SIA, and the presence of Hcrt neurons is essential for SIA.
Thyrotropin-Releasing Hormone
A Peptidergic Activator of Hcrt Neuron Activity
Note that the location of the injection site is dorsal to the distribution of hypocretin neurons. Neuropeptide Y inhibits hypocretin/orexin neurons by multiple presynaptic and postsynaptic mechanisms: tonic depression of the hypothalamic arousal system.
Overview of Mouse Models of Narcolepsy
This chronic sleepiness is often severe and can significantly impair interpersonal relationships, the ability to drive safely, and performance at school and work. All of these hypotheses have their merits, but have been difficult to fully test in clinical studies.
The Neural Basis of Sleepiness in Narcoleptic Mice
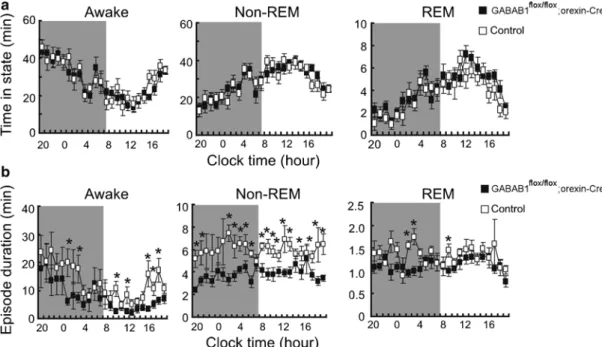
Previous reports have shown that double KO mice lacking OX1 and OX2 receptors have severe sleepiness similar to that seen in orexin KO mice [10]. Mice lacking OX1 and OX2 receptors (OX1/2 TD) show severe fragmentation between sleep and wakefulness, almost as severe as orexin KO mice, but have less cataplexy (indicated by triangles).

Behavioral State Instability in Narcoleptic People
Furthermore, in the midst of what should be stable, full wakefulness, orexin KO mice showed rapid transitions to NREM sleep, suggesting that even high levels of arousal can be unstable. The latter phenomenon may be similar to the occasional "sleep attacks" seen in people with narcolepsy and warrants further investigation, as it suggests that in the absence of orexins, activity in wakefulness-promoting systems can rapidly fail.
How Mouse Models Provide New Insights into the Sleepiness of
Yet wakefulness-promoting circadian signals can reach key arousal regions even in the absence of the orexin neurons. In rats with lesions of the lateral hypothalamus that kill most of the orexin neurons (and other nearby neurons), adenosine levels in the basal forebrain do not rise over 6 h of sleep deprivation [35] .
Appendix
Instead, there is much evidence that sleepiness may be due to reduced activity in a number of wake-promoting regions or insufficient inhibition of sleep-promoting regions. For example, narcoleptic mice have reduced or less consistent activity in monoaminergic neurons during wakefulness.
Advantages and Limitations of Using Mice to Understand Sleep
One can raise the same concern that some of the narcolepsy phenotype in orexin KO mice is caused by abnor-. Contrasting effects of ibotenate lesions of the paraventricular nucleus and subparaventricular zone on sleep-wake cycle and temperature regulation.
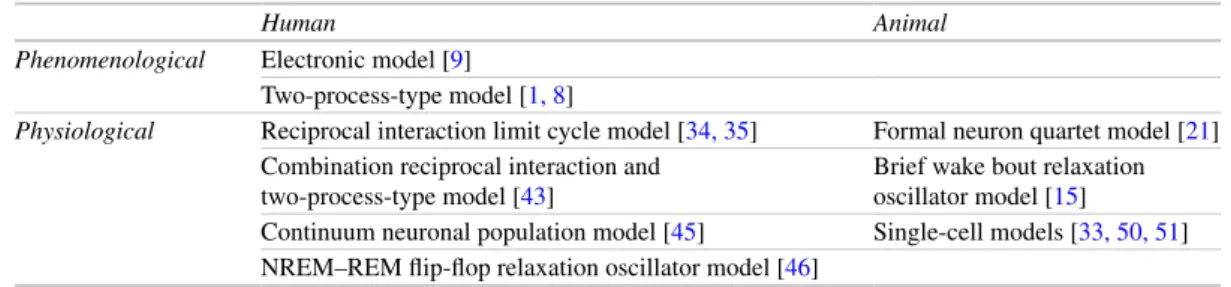
Phenomenological Models of Narcolepsy in Humans
Although mathematical modeling approaches have generally only recently been applied to narcolepsy, developments in modeling normal sleep/wake behavior have laid an excellent foundation for linking the experimental insights about the orexin (also known as hypocretin) system with the observed features of the narcolepsy phenotype. Physiologically based models generate predictions about mechanisms underlying specific elements of the narcolepsy phenotype.
Mathematical Models of Narcolepsy
Therefore, the two-process model alone may not be able to capture certain aspects of the narcolepsy phenotype, but the interactions between Process S and Process C will be important elements of more detailed models. Lawder attempted to model many aspects of the narcolepsy phenotype, including sleep attacks, cataplexy, sleep paralysis, and hypnagogic hallucinations, while maintaining normal nighttime sleep duration.
Phenomenological Models of Narcolepsy in Animals
Although phenomenological models are limited in their ability to predict mechanisms for the properties they describe, they provide an important formalization of conceptual hypotheses. Furthermore, by linking conceptual elements such as the S process to clinical measures such as SWA, phenomenological models provide general insights that can ultimately be explored in a relevant biological substrate.
Physiologically Based Models of Narcolepsy in Animals
Therefore, fragmented sleep/wake behavior was the main focus of the modeling approach of Diniz Behn and colleagues [15]. When simulated orexin effects were omitted, the model reproduced the fragmented sleep-wake behavior of OXKO mice.
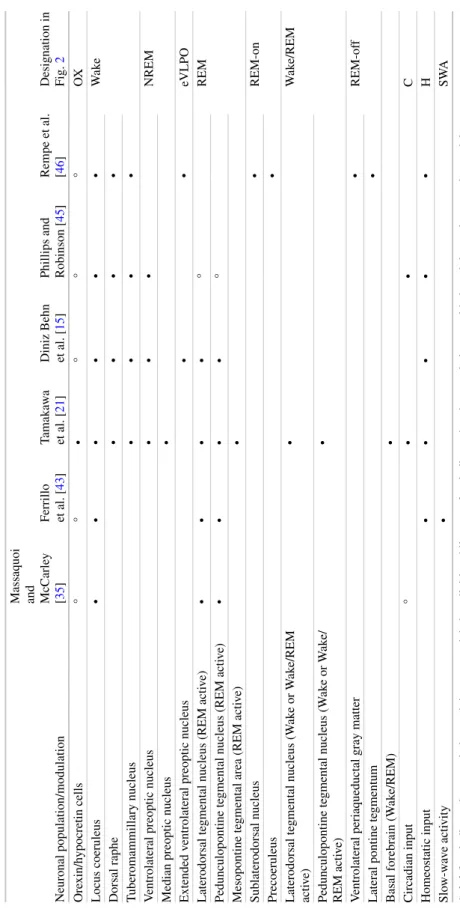
Physiologically Based Models of Narcolepsy in Humans
Interestingly, desensitization to dynorphin lasts approximately 90 seconds, consistent with the delay in functional orexin effects identified in the survival analysis. Therefore, the colocalization of peptides combined with the dynamics of desensitization may result in complex modulation of postsynaptic targets manifesting as a delay in the onset of orexin effects.