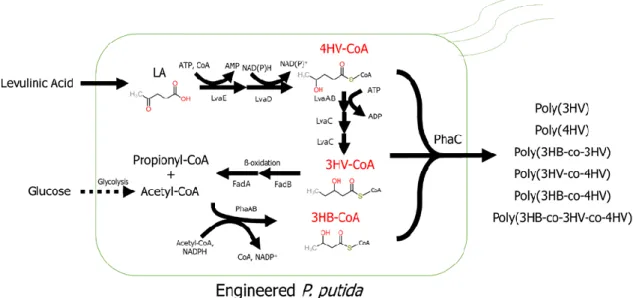
Polyhydroxybutyrate (PHB) is considered a good alternative to petroleum-based polymer due to its biodegradability and renewability. To this end, we produced four different short-chain polyhydroxyalkanoates of levulinic acid in construct Pseudomonas putida and express heterologous PHA synthase in. Furthermore, fadB deletion to block 3HV-CoA degradation affected relatively high 3HV ratios in the production of P( 3HV -co-4HV).
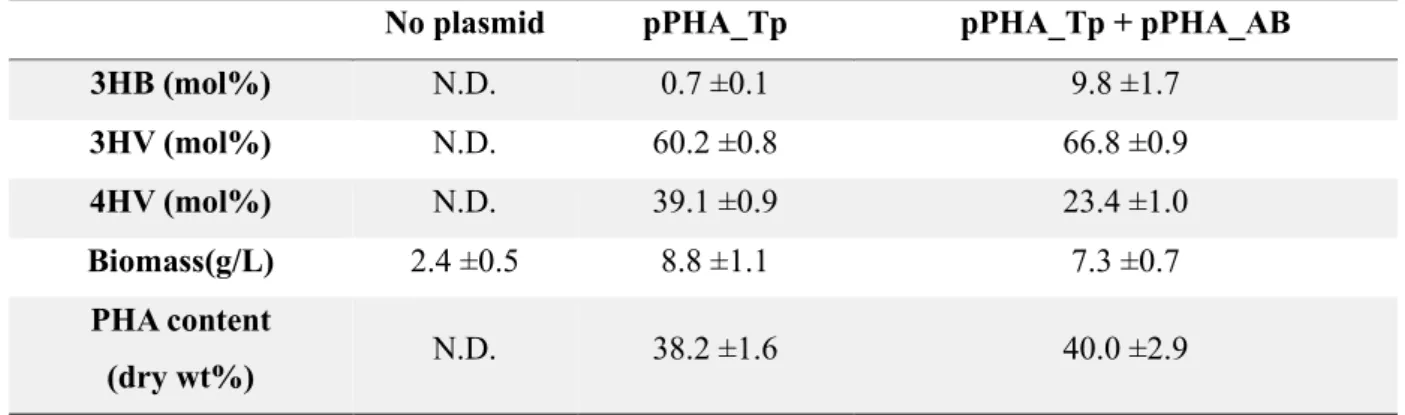
CDW, and PHA (%) content and composition from strain SP01 with plasmid pPHA_Tp and pPHA_AB. CDW, and PHA (%) content and composition from strain SP02 with plasmid pPHA_Tp and pPHA_AB. CDW, and PHA (%) content and composition from strain SP03 with plasmid pPHA_Tp and pPHA_AB.
Introduction
In this study, we investigated the production of different PHAs from monomers produced by the engineered LA to P catabolic pathway. We demonstrated that biomass-derived LA can replace petroleum-based precursors, such as propionate, valeric acid, or γ- valerolactone, for the production of PHAs with 3HV and 4HV monomers.
Methods and materials
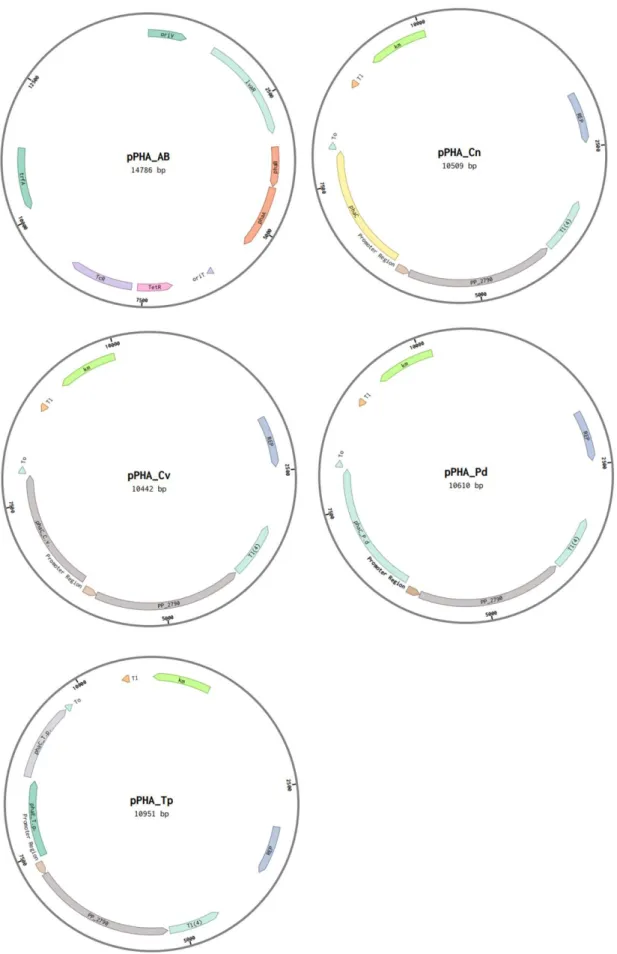
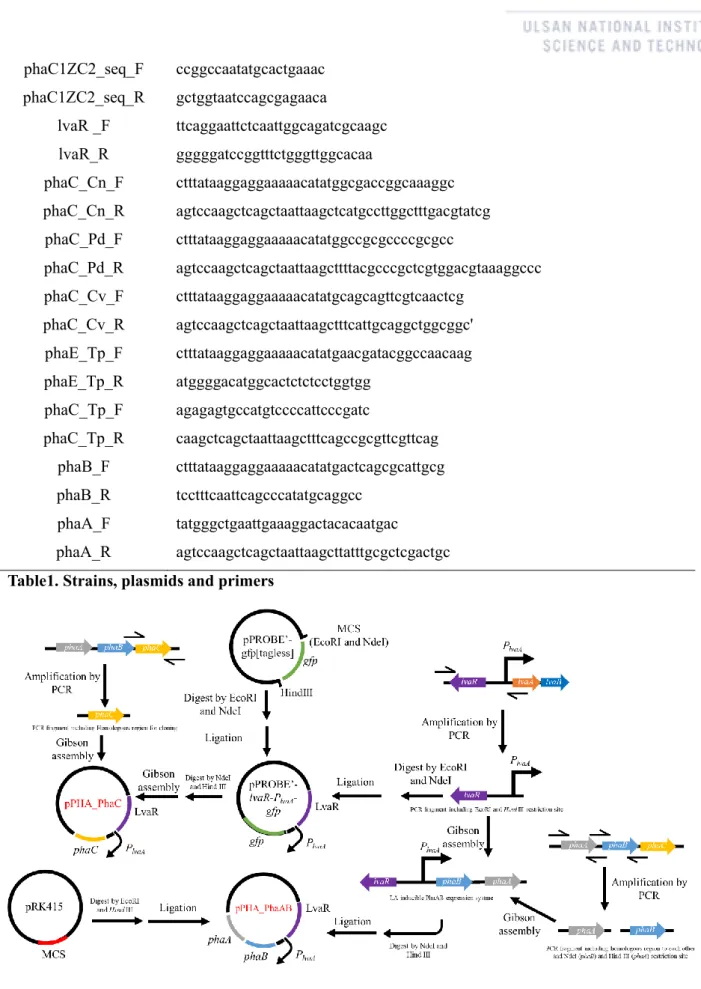
- Strains, plasmids and primers
- Enzymes, chemicals and media
- Cultivation of P. putida
- PHA analysis
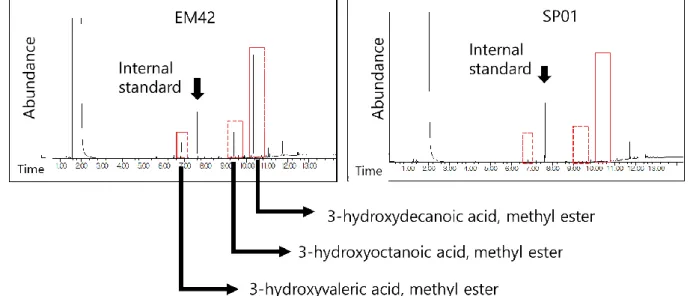
Restriction enzymes (Fermentas), DNA ligase (NEB) and polymerase (Ex Taq, Takara) were used for plasmid construction. Gamma-valerolactone, methyl-3-hydroxyvalerate (Santa Cruz Biotechnology), methyl-3-hydroxybutylate (Sigma-Aldrich), methyl-benzoate (Acros Organic), Poly(3-hydroxybutyric acid-co-3-hydroxyvaleric acid) (Sigma) - Aldrich) and benzoic acid were used for GC-MS standard. Three colonies were picked from the plates and incubated for 15 h in 5 ml LB.
Then, 1 ml of the sample was inoculated into a minimal culture medium to which 7.5 g/L glucose and 7.5 g/L LA were added. The frozen pellets were lyophilized overnight to measure the dry cell weight of the dried cell. After measuring the weight, the cells were mixed with 2 ml of methanol-sulfuric acid (15% v/v 95% sulfuric acid) and 2 ml of chloroform solution and reacted at 100 °C for 4 hours.
After the reaction was complete, 1 mL of TDW was added and vortexed to mix the solution. Finally, 100 µl sample was taken from the lower organic solvent layer and diluted in 900 µl chloroform to prepare a sample for GC-MS analysis.
Results and Discussion
- Elimination of the ability of P. putida KT2440 to produce mcl-PHAs
- Scl-PHA production in the phaC1ZC2-deleted P. putida strain
- Effect of the lvaAB deletion on PHA production and monomer composition
- Effect of the fadB deletion on PHA production and monomer composition
- Comparison of different PHA polymerase on scl-PHA production
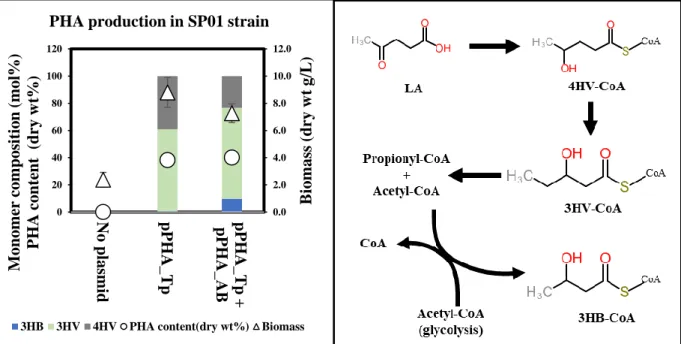
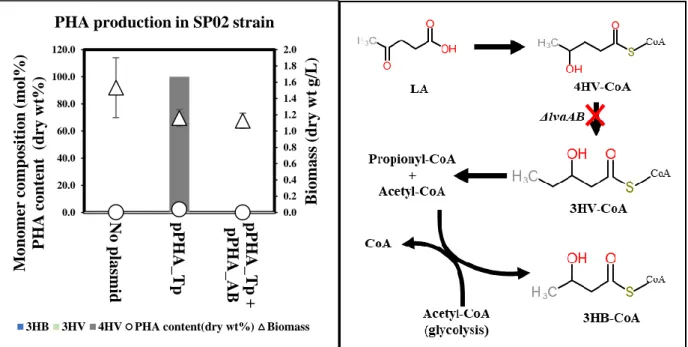
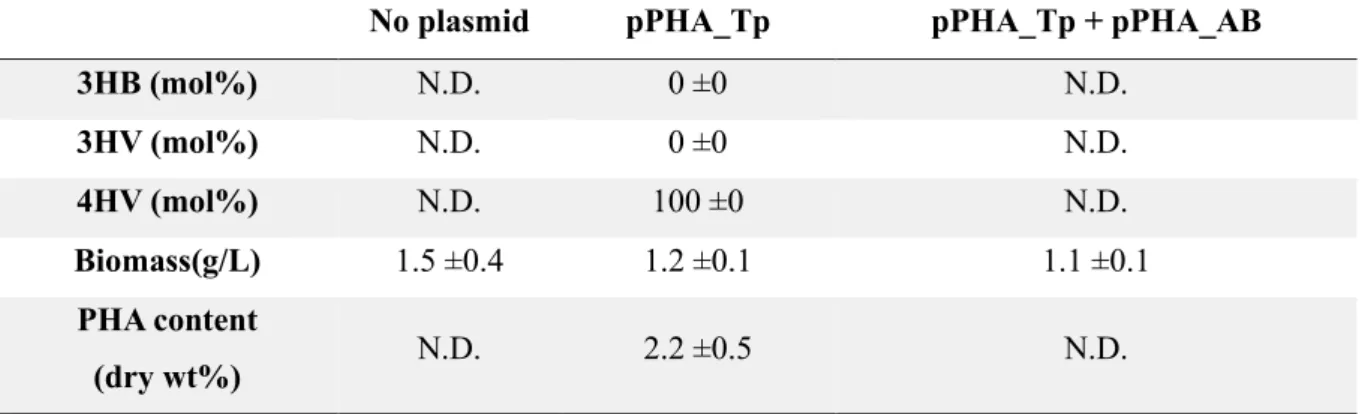
To test whether the two intermediates, 4HV-CoA and 3HV-CoA, are catabolic pathways of LA in P. These results include the possibility of increased flux from 4HV-CoA to 3HV-CoA upon PhaAB expression and possible increased metabolism of LA due to consumption of acetyl-CoA , because two acetyl-CoAs were consumed to produce one 3HB-CoA monomer. In order to produce different types of scl-PHA using the LA catabolic pathway, a lvaAB deletion 4HV-CoA accumulation strain was constructed.
The PHA synthase of Thiocapsa pfennigii (PhaC_T.p.) was expressed with and without PhaAB in the resulting strain. These results indicate that there may be steric effects between the PhaC enzyme and the P(4HV) homopolymer, resulting in lower enzyme activity [ 40 ]. When PhaC was co-expressed with PhaAB in the lvaAB-deleted strain, the strain did not produce polymer in flask culture.
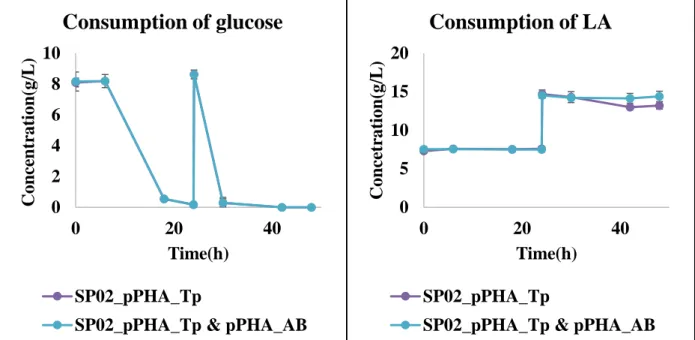
These results indicate that, in addition to the low enzyme activity described above, there is an additional effect due to PhaAB expression. Considering that expression of PhaAB leads to excessive consumption of acetyl-CoA and that production of 4HV-CoA consumes two ATPs and one NAD(P)H, this result is due to a lack of energy from additional expression of PhaAB. LA is converted to acetyl-CoA and propionyl-CoA via the Lva operon containing lvaABCDEFG and beta-oxidation fadBA genes and is then completely oxidized via the TCA cycle.
The strain that can accumulate more 4HV-CoA and 3HV-CoA was constructed by deleting fadB in order to make more LA incorporated into scl-PHA by blocking its complete oxidation. There was more than a two-fold increase in the molar content of 3HV in the copolymer compared to 4HV. Considering the low 3HB production and low growth, these results indicate that LA metabolism was delayed by fadB deletion and Acetyl-CoA depletion due to PhaAB expression significantly delayed cell growth.
It is known that PHA synthases determine the compositional variation of the resulting polymer and efficient production of PHAs with desired properties is of industrial importance. Four different PHA synthetases (Cupriavidus necator H16, Chromobacterium violaceum, Thiocapsa pfennigii and Paracoccus denitrificans) were expressed in the SP01 strain. In this case, 36.7–40% of the total PHA content was found, which is comparable to before and after PhaAB expression.

Conclusions
Amirul, "Studies on Microbial Synthesis and Characterization of Polyhydroxyalkanoates Containing 4-Hydroxyvalerate Using γ-Valerolactone," Applied Biochemistry and Biotechnology, vol. Wang et al., "Biosynthesis and Thermal Properties of PHBV Produced from Levulinic Acid by Ralstonia eutropha, " PLoS ONE, vol. Steinbüchel, "Development of a process for biotechnological large-scale production of 4-hydroxyvalerate-containing polyesters and characterization of their physical and mechanical properties," Biomacromolecules, vol.
Pradella, "Production of poly(3-hydroxybutyrate-co-3-hydroxyvalerate) P(3HB-co-3HV) with a wide spectrum of 3HV content at high yields by Burkholderia sacchari IPT 189," World Journal of Microbiology and Biotechnology, vol. . Chen, "Production and characterization of homopolymer poly(3-hydroxyvalerate) (PHV) accumulated by wild-type and recombinant Aeromonas hydrophila strain 4AK4," Bioresource Technology, vol. Kachrimanidou et al., "Sunflower-based biorefinery: Poly(3-hydroxybutyrate) and poly (3-hydroxybutyrate-co-3-hydroxyvalerate) production from crude glycerol, sunflower meal and levulinic acid,” Bioresource Technology , vol.
Park, "Development of a deletion mutant of Pseudomonas denitrificans that does not degrade 3-hydroxypropionic acid," Applied Microbiology and Biotechnology, vol. Shang, "Pseudomonas putida KT2440 markerless gene deletion using a combination of λ Red recombination and Cre/loxP site-specific recombination, " FEMS Microbiology Letters, vol. Durfee et al., "The complete genome sequence of Escherichia coli DH10B: insights into the biology of a laboratory workhorse," J Bacteriol, vol.
Lee, “Construction of the lva operon and optimization of culture conditions for improved production of 4-hydroxyvalerate from levulinic acid in Pseudomonas putida KT2440,” Journal of Agricultural and Food Chemistry , vol. Davis et al., "Conversion of grass biomass into fermentable sugars and its use for medium chain length polyhydroxyalkanoate (mcl-PHA) production by Pseudomonas strains." Olivera et al., "Molecular characterization of the phenylacetic acid catabolic pathway in Pseudomonas putida U: the phenylacetyl-CoA catabolon," (in eng), Proc Natl Acad Sci U S A, vol.
De Eugenio et al., “Biochemical evidence that the phaZ gene encodes a specific intracellular medium-chain polyhydroxyalkanoate depolymerase in Pseudomonas putida KT2442 characterization of a paradigmatic enzyme,” Journal of Biological Chemistry, vol. Wittmann, “In silico-driven metabolic engineering of Pseudomonas putida for improved production of poly-hydroxyalkanoates,” Metabolic Engineering, vol. Ueda, "Growth-associated production of poly(3-hydroxyvalerate) from n-pentanol by a methylotrophic bacterium, Paracoccus denitrificans". in eng), Applied and Environmental Microbiology, vol.