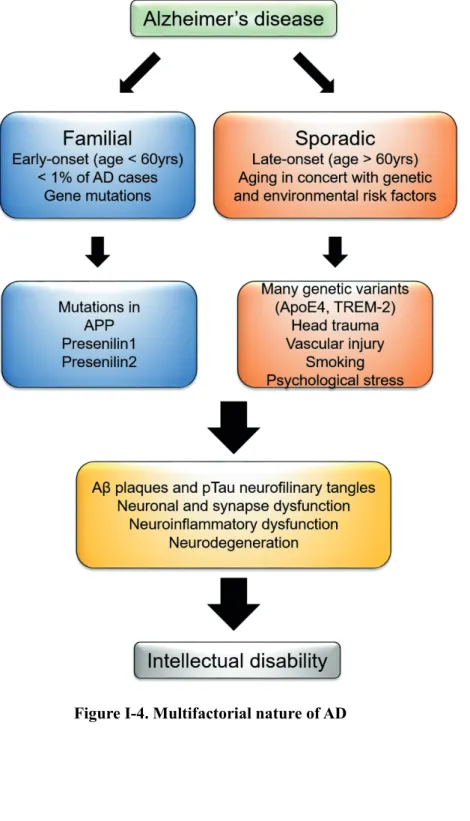
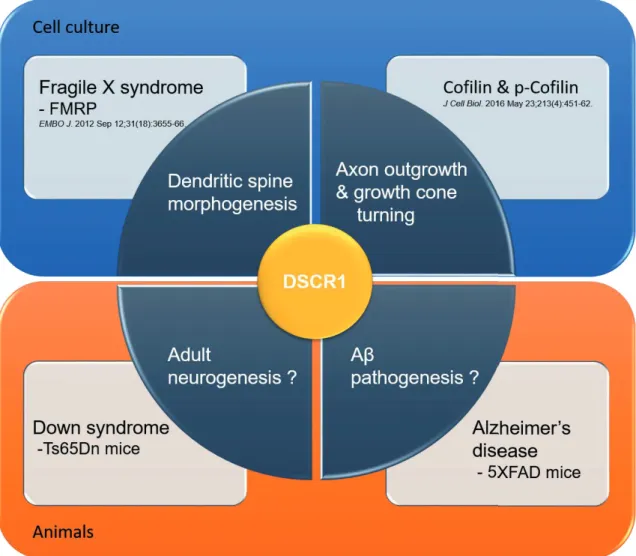
Down syndrome (DS) is one of the most common chromosomal disorders in the world, as it occurs in one out of 600 to 800 live births in the general population. However, it remains unclear which critical region or genes in the extra copy of chromosome 21 are associated with cognitive impairment in DS patients. DSCR1 (Down syndrome critical region 1, also called RCAN1), located on chromosome 21 of Homo sapiens, is highly enriched in the brain, particularly in the hippocampus.
Patients with Down syndrome and Ts65Dn, a mouse model of Down syndrome, showed greatly increased expression of DSCR1 in the brain, suggesting that DSCR1 may play a major role in cognitive impairment in DS. However, it is still unknown how DSCR1 contributes to the intellectual process associated with adult hippocampal neurogenesis (AHN). In this thesis, we demonstrated the novel role of DSCR1 as a regulator of AHN in the rat brain. Although overexpressed amyloid precursor protein (APP), encoded by chromosome 21, is the most likely cause of AD in Down syndrome patients, the effect of other genes in the pathogenesis of AD, including DSCR1, has not been well studied.
Since DSCR1 is overexpressed in AD brain, we hypothesize that DSCR1 may be closely associated with AD neuropathology. Moreover, DSCR1 overexpression in 5XFAD mice shortens pathological defect of amyloid-beta in the brain and restores memory deficit. Blocking drainage via ligation surgery for cervical lymphatic vessels afferent to deep cervical lymph nodes abrogated the positive impact of DSCR1 and cognitive function on Aβ clearance.
Background
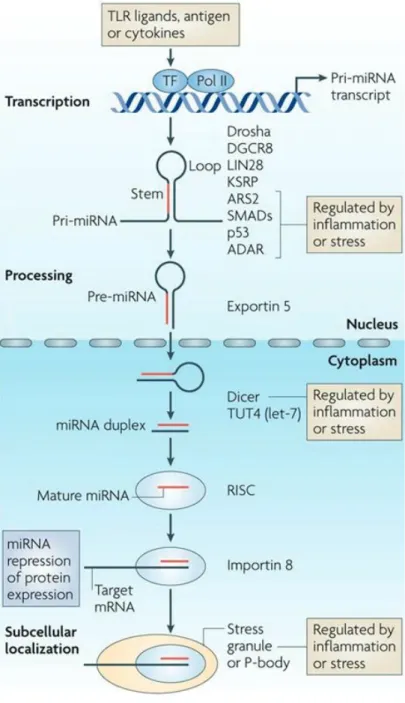
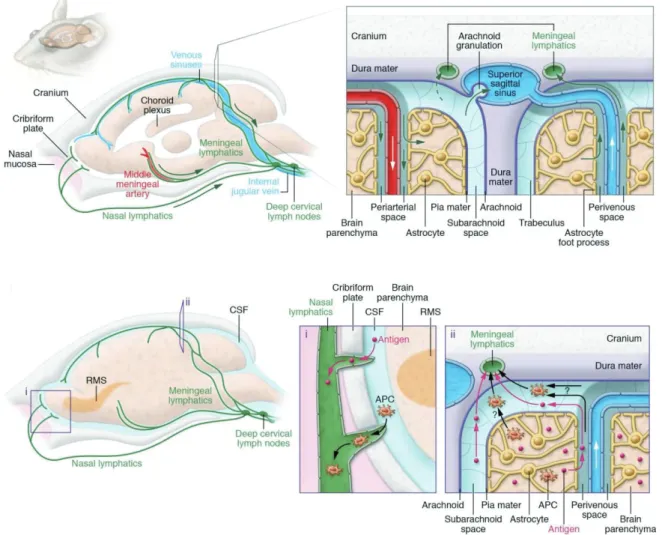
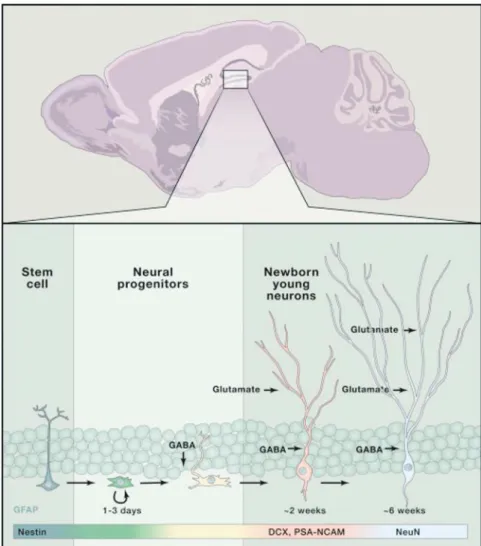
A recent study suggested that miR-124 promotes neurogenesis in the subventricular zone (SVZ) of the adult mammalian brain 9. Conversely, Tet 1 enzyme plays a crucial role in the active demethylation of methylated DNA via oxidation of 5mC. Recent studies in the field of CNS have revealed the crucial role of the meningeal lymphatic system in various neurological disorders with age 17 , 18 .
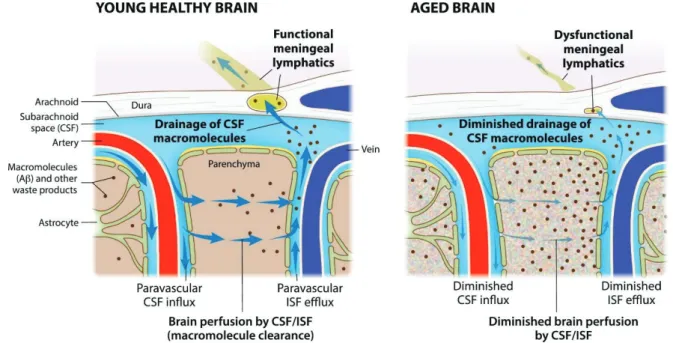
Notably, meningeal lymphatic dysfunction has been shown to accelerate amyloid plaque burden in the brain and meninges and develop memory loss in aged tg mouse models of AD 15 , 16 (Fig. I-6). The Down syndrome critical region1 (DSCR1, also called RCAN1) is located on human chromosome 21 and shows an enriched expression pattern in the brain, particularly in the hippocampus. Meanwhile, adult neurogenesis has been linked to hippocampus-dependent intellectual disabilities associated with disorders located in the neurodevelopmental group such as DS.
Notably, DSCR1 is abundantly enriched in the hippocampus; however, the function of DSCR1 in adult neurogenesis was previously unknown. Interestingly, according to early studies on lymphangiogenesis, DSCR1 was one of the most strongly induced genes by VEGF-C and VEGF-A, which were previously reported to be involved in the promotion of lymphangiogenesis 28 - 32 .
Experimental Method and Material
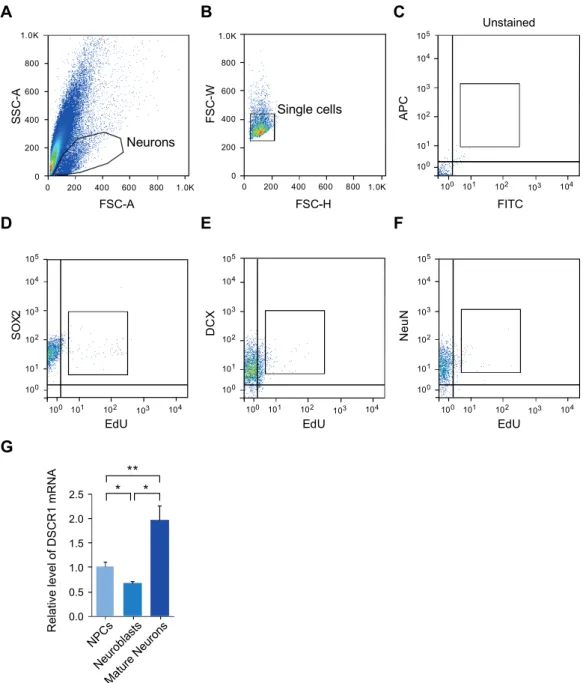
Samples were collected from fine dissections of the adult hippocampus and were incubated in a dissociation medium with the enzyme mixture (Dispase 1U/ml, Sigma Aldrich and Papain 2.5 U/ml, Sigma Aldrich). After anesthesia with 2% isoflurane, 8-week-old mice were stereotaxically injected with the virus into the hippocampal DG using a stereotaxic apparatus. Next, washing 3 times with ice-cold lysis buffer and elution of the resin-bound proteins was performed, and then immunoblotting was performed to detect the binding of DSCR1.
All mice were adequately anesthetized by intraperitoneal injection of tribromoethanol ('Avertin') after transcardial perfusion with chilled 0.1 M PBS. The scalp was excised by cutting the rostral tip of the nasal bone, and then the facial muscles attached to the skull were removed. Mice were anesthetized with an intraperitoneal injection of Avertin and then the head was fixed in a stereotaxic apparatus.
Injection of 1 μl of Ovalbumin Alexa Fluor 488 conjugate was performed stereotaxically 2.5 mm below the surface of the skull with a microsyringe (Hamilton) at an injection rate (0.2 μl per minute over 5 min) 45 , and the needle was held to avoid a reflux of the injected solution for at least 5 minutes. After injection of fluorescent tracers, the brain and lymph nodes were harvested and images were acquired on a ZEISS Axio Zoom. V16 Stereo Zoom fluorescence microscope equipped with an Andor's iXon EMCCD camera. A vertical skin incision was made in the midline from the neck to the manubrium of the sternum and then the opening was made exposing the trachea, posterior belly of the digastric muscle (PBD), sternocleidomastoid muscle (SCM) to reveal the lymphatic vessels of the neck. uterus. (cLVs) 15.
For the intracerebroventricular (i.c.v.) injection, titrated viruses were delivered stereotaxically into the left side of the lateral ventricular CSF in 4-month-old AD mice. After injecting the purified AAV1 particles (1 µl, 3 x 1011 genome copies (GC)/ml) at an injection rate (0.2 µl per minute for 5 minutes), the needle was kept in place for another 5 minutes to prevent backflow. prevent. . Branch point analysis was implemented via the Skeleton plugin of ImageJ software by counting the number of branch crossings in the branchial network of meningeal lymphatic vessels.
Light-sheet fluorescence microscopy (Lightsheet Z.1) was used to image whole-mount meninges, and a 3D image was subsequently reconstructed using Imaris software (Bitplane) for quantitative analysis of the lymph volume. The Lyve1 positive surface ROI was additionally created to calculate meningeal lymph volume. Then the centrifuged cells were passed through a column (LS MACS) placed in the strong magnet of the MACS separator.
Results
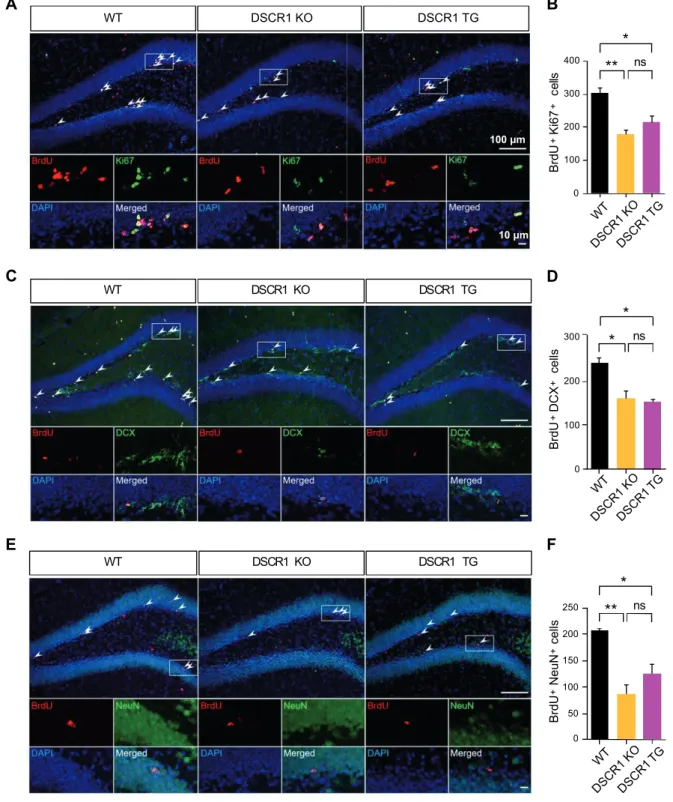
Adult hippocampal neurogenesis was modulated by DSCR1. A) Representative image with BrdU and Ki67 to detect dividing cells in the subgranular zone of the DG. Furthermore, upregulation of the expression of miR-124 induced the AN defects in the SVZ, located along the lateral ventricle 55, 56. We first validated the expression of different forms of miR-124 in the hippocampi, including primiRNA and pre-miRNA/mature using qRT-PCR (Fig Ⅲ-1-5A) 57.
We induced a specific mutation at CpG sites in the normal miR-124 promoter to block methylation. We also induced mutations at all methylation sites residing in the hippocampus of DSCR1 TG mice. We tested whether DSCR1 affects the interaction with intron 8 of TET1 in the presence of U1/U2 snRNA 59 .
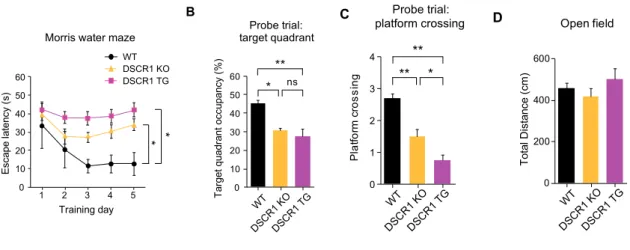
Subsequently, DSCR1/5XFAD mice showed improvement in the spatial learning and memory task in the MWM test, suggesting that DSCR1 plays a protective role in 5XFAD mice. -E) Control littermates, 5xFAD mice, and DSCR1/5XFAD mice were tested in the MWM at 5 months of age. Interestingly, we also noted no obvious structural differences in the dural lymphatics between the DSCR1/5XFAD mice and wild-type mice (Fig. Ⅲ-2-5B).
We found that dcLN leakage of dye in sham-operated DSCR1/5XFAD mice, whereas tracer uptake was abolished in ligated mice. Interestingly, removal of meningeal lymphatic vessels from ligation worsened Aβ deposition in meningeal lymphatics and brain parenchyma (Fig Ⅲ-2- 6D-G) and also worsened learning and memory ability (Fig Ⅲ-2-6H-K). These findings strengthened that DSCR1 is necessary for recovery of the lymphatic system in the AD mouse model, which alleviates amyloid pathology and behavioral abnormalities.
Quantification of the OVA-AF647 drainage in the dcLN with a sham operation compared with ligation surgery. NFAT-calcineurin signaling is one of the major pathways induced by DSCR1 in the mature and developing CNS 80 . The first of these conclusions supports a novel role of DSCR1 in the processes of AHN.
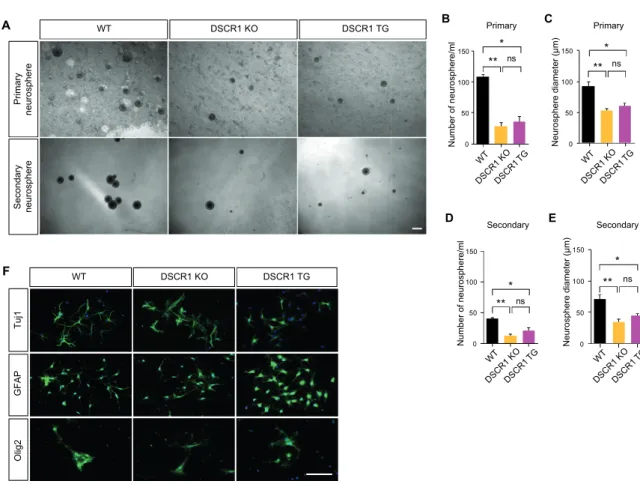
Next, DSCR1 knockdown enhanced TET1 splicing and expression, which was confirmed in the hippocampus of DSCR1 KO mice. Taken together, these results suggest that proper expression of DSCR1 in the hippocampus is required for the normal AHN, which subsequently regulates miR-124 and TET1 expression.
DSCR1 regulates AHN by regulating crosstalk between TET1 and miR-124
Tables
- Discussion
DSCR1 forward TCATCGACTGCGAGATGGAG DSCR1 reverse TGGTGTCCTTGTCATATGTTCTG TET1 pre-mRNA forward CGCCATCACACCATGCAAA TET1 pre-mRNA reverse GCCATCTGCTGCCCTCTTCT TET1 mature mRNA forward GAAGGAGAGCAGCCGATGETag forward GGT pri-miR-124 forward CATCCTCCCTCTCTTCCATC pri-miR-124 reverse TTAAATAAGGTCCGCTGTG pre-miR-12 4 before AGGCCTCTCTCTCTCCGTGTTCA before -miR -124 reverse CAGCCCCATTCTTGGCATTC mature miR-124 forward GCGAATGCATTAAGGCACGCGG. Meanwhile, dysfunction or altered levels of RBPs have been associated with various disorders, such as neurological disorders and cancers 76, 77. However, the exact role of DSCR1 in the study of RNA-protein interactions is not fully understood. Surprisingly, correcting DSCR1 levels is sufficient to recover the defects of enhanced hippocampal neurogenesis and Aβ pathogenesis in DS and AD mice, resulting in improved cognitive function.
However, overexpressed DSCR1 induces impaired adult neurogenesis in the hippocampus, while DSCR1 overexpression alleviates Aβ pathology and improves learning and memory by potentiating the meningeal lymphatic system. Thus, additional functional study of DSCR1-enhanced neurogenesis remains to be investigated in AD pathogenesis. Although DSCR1 is the representative calcineurin regulator, we identified that DSCR1 also controls the miR-124-TET1 pathway.
Therefore, it would be interesting to determine whether the DSCR1-mediated calcineurin/NFAT signaling pathway controls AHN together or independently with miR-124-TET1. We identify that TET1 mediates the methylation status in the promoter region of miR-124, affecting its levels. These findings demonstrated the relationship between DNA methylation and microRNA (miRNA) expression in the AHN, which is supported by our several results.
Furthermore, correcting a single copy of DSCR1 in Ts65Dn mice fully rehabilitated AHN and learning and memory abnormalities. Blockade of cLVs by ligation surgery in hybrid mice aggravates Aβ pathogenesis in the dural lymphatics and parenchyma of the brain, which shows the reduced cognitive ability. RQVHW$O]KHLPHUVGLVHDVHUHYLVLVHG$O]KHLPHUV 'HPHQWLD . .XU] $ 3HUQHF]N\ 5 $P\ORLG FOHDUDQFH DV D WUHDWPHQW WDUJHW DJDLQVW .. amp; & 2OLYHU * .LSQLV - &16 O\PSKDWLF GUDLQDJH DQG QHXURLQIODPPDWLRQ DUH UHJXODWHGE\PHQLQJHDOO\PSKDWLFYDVFXODWXUH1DWXUH1HXURVFLHQFH.
0DNH\HY(9 =KDQJ- &DUUDVFR0$0DQLDWLV77KH0LFUR51$PL5 SURPRWHVQHXURQDOGLIIHUHQWLDWLRQE\WULJJHULQJEUDLQVSHFLILFDOWHUQDWLYHSUH P51$VSOLFLQJ0ROHFXODUFHOO . .. amp;KDU DFWHUL]DWLRQIURP