Thereafter, an abrupt loss of sea ice is observed, even more so compared to the Arctic (Parkinson, 2019). Winter sea ice algae dynamics (composition, abundance, community distribution and relationship between sea ice interfaces), particularly within the seasonally ice-covered areas of the SO Marginal Ice Zone (MIZ) is poorly documented (Edwards et al., 1998).
Physical description of the Southern Ocean
The area of Antarctica covered by seasonal ice is called the Seasonal Sea Ice Zone (SSIZ), which includes the SBACC and interacts with the ACC, the Antarctic Divergence and the Antarctic Coastal Current (Massom & Stammerjohn, 2010; Robinson & Brink, 2006). East of this region, sea ice propagates along the general wind and circulation in the ACC (Talley et al., 2011).
Sea-ice dynamics
These processes are mostly responsible for the microstructure and porosity of sea ice (Mock & Thomas, 2005). If sea ice permeability is high or the vertical gradient is unstable (caused by increased density and salinity), gravity drainage or convective overturning is also observed (Meiners & Michel, 2016; Vancoppenolle et al., 2010).

Relationship between sea-ice dynamics and sea-ice algae
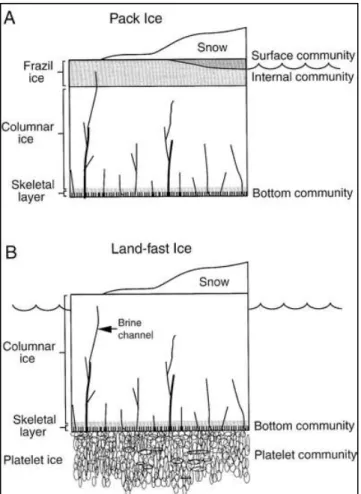
This is markedly different from the continuous ice layer formed as a result of conductive heat loss along the temperature gradient of dense columnar crystalline structure of land fast ice. However, findings by Clarke & Ackley (1984) revealed that congel ice can have two orders of magnitude higher chlorophyll a concentrations than that of frazil-dominated ice.
Relationship between biochemistry of sea-ice and sea-ice algae
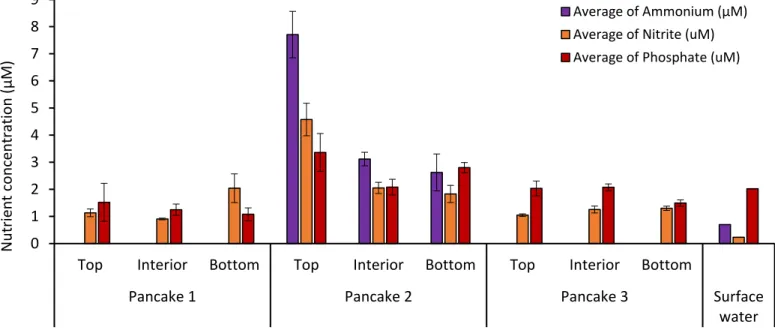
Bottom communities (typically with the highest biomass) showed a preference for newly formed nitrate, while interior communities showed higher ammonium uptake (considered part of the regeneration microbial loop) (Meiners et al., 2016). Additionally, experimental studies have shown increased (20%) growth of sea ice algae on the surface layer of sea ice when exposed to increased CO₂ at constant pH (McMinn et al., 2014).
Relationship between sea-ice and sea-ice algae
These processes are also considered to be essential for the accumulation of sea ice algal cells (Eicken, 2003). Snow loading not only changes the microstructure of sea ice, but also plays a central role in sea ice growth and primary production (Massom & Stammerjohn, 2010).
Physiological adaptations of sea-ice algae against challenging sympagic environment
Narrow brine channels ≤ 200 µm in diameter may also provide a refuge for sea ice microbial communities (Krembs et al., 2000). They contribute to the success of diatom species in sea ice and freezing temperatures (Janech et al., 2006).
Ecological role of sea-ice algae
Therefore, the extracellular secretion of the antifreeze IBP prevents recrystallization of sea ice and subsequent damage to diatom cell membranes (Janech et al., 2006). These hollow appendages are a morphological advantage for planktonic species (increase buoyancy, nutrient uptake and to ward off grazers) (Kooistra et al., 2010), therefore peculiar to find in high abundance and even grow in sea ice environments (Ligowski et al. al. , 2012).
Southern Ocean food web
The importance of the role of sea ice algae in synthesizing and exporting carbon to the pelagic environment and higher trophic levels remains to be quantified, as there is great spatial and temporal variability (Kohlbach et al., 2018). It is predicted that if sea ice extent retreats by 50%, there will be an 86% reduction in primary production derived from sea ice algae (Kohlbach et al., 2018).
Seeding of phytoplankton blooms
Influence of sea ice on the composition of spring phytoplankton blooms in the northern Baltic Sea. The influence of sea ice on the structure of phytoplankton communities in the northern Antarctic Peninsula.
36
Methodology
- Study area
- Sample melting and subsequent analysis ..................................................... Error! Bookmark not defined
- Standard spectrophotometric nutrient analysis
- Salinity and Temperature analysis
- Statistical analysis
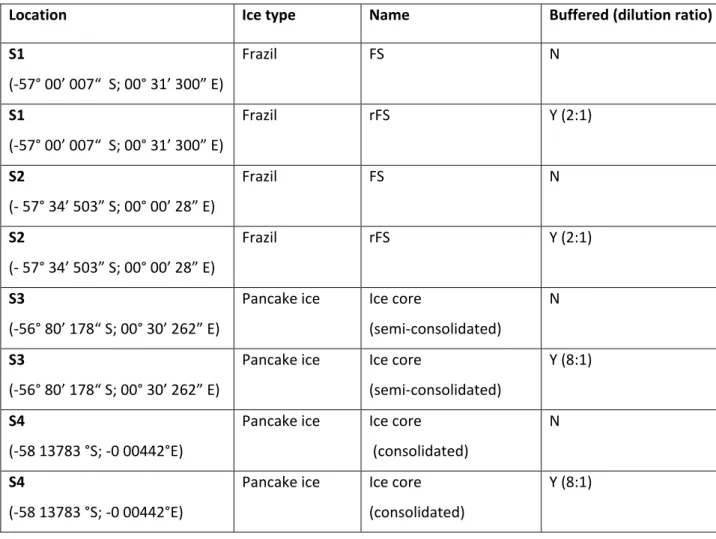
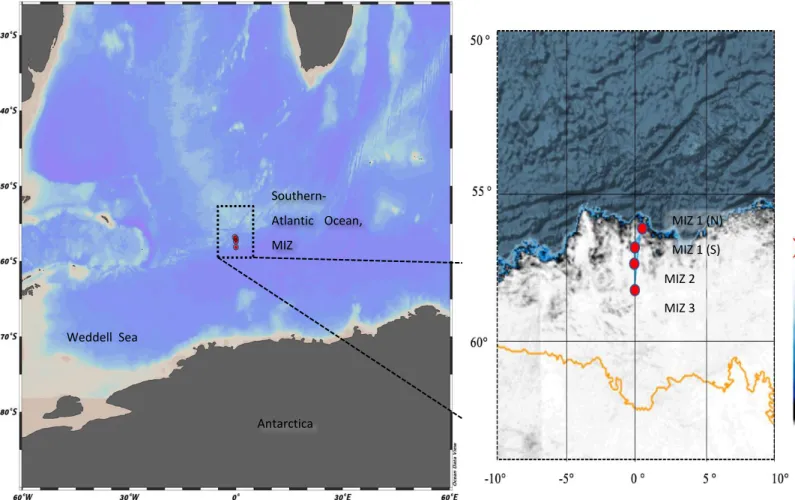
The sea ice samples were collected during the austral winter of July 2017 in the MIZ of the southern Indian Ocean at 65°S, 30°E (Figure 2. 1). The sea ice samples used for algal identification were directly melted at room temperature in the dark to prevent algal growth and osmotic stress. At the Cape Peninsula University of Technology's ecotoxicology laboratory, the sea ice samples were determined using the Utermöhl method (Utermöhl, 1931).
Forty ml of three samples of melted sea ice containing a wide variety of species were analyzed using scanning electron microscopy (SEM). Sea ice and surface seawater samples used for chlorophyll A analysis were analyzed aboard the SA Agulhas II. MDS graphs illustrate "natural algal groups" by sea ice depth and individual samples.
Multivariate analysis with combined dependent variables was used to test for homogeneity of variances among sea ice samples. The Shapiro Wilkinson normality test was used to test for equality of variance in sea ice algal abundance across sea ice samples.
Results
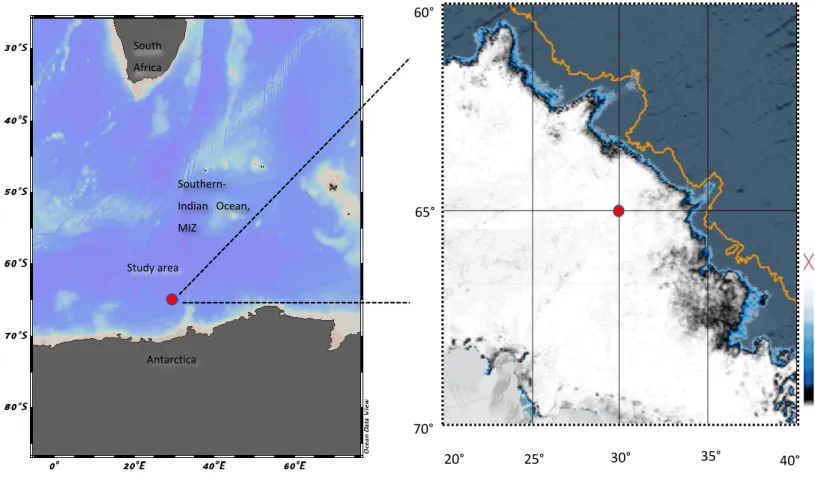
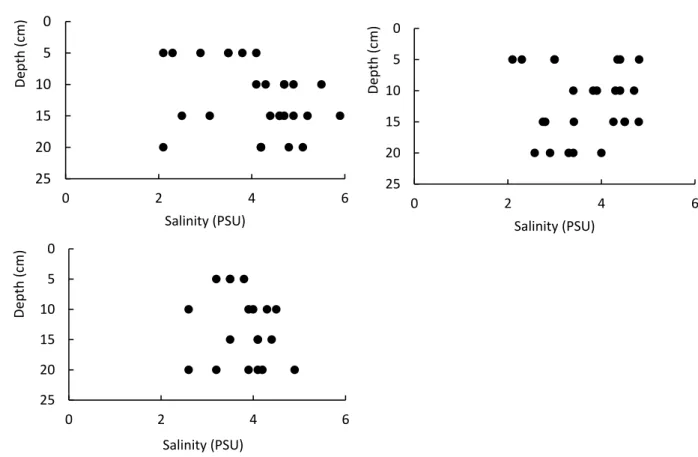
- Physicochemical environment
- Sea-ice algae biomass
- Biological community structure
- Relationship between biochemical sympagic environment and algae community structure
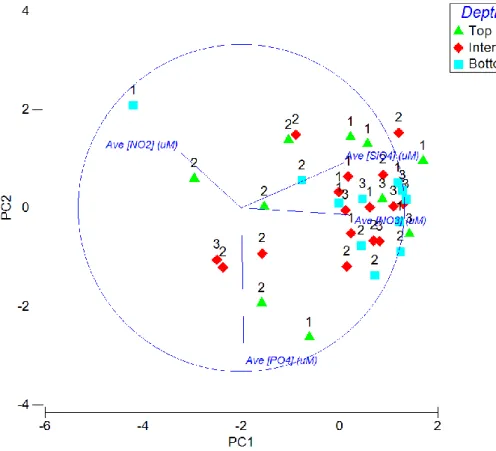
Conversely, sea ice pennate diatom cell concentration was markedly higher compared to surrounding surface water. No clear pattern was observed for diversity indices (species richness and diversity) in sea ice samples (Table 2. 3). The MDS analysis showed clear groupings between community structure by depth and pancake sea ice sample (Figure 2. 13).
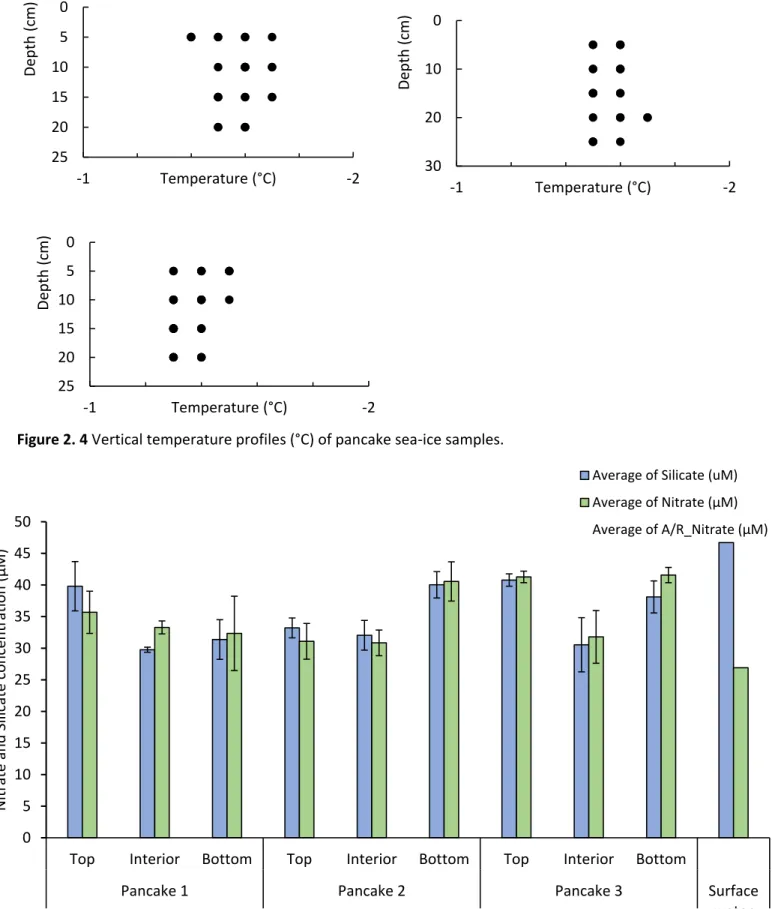
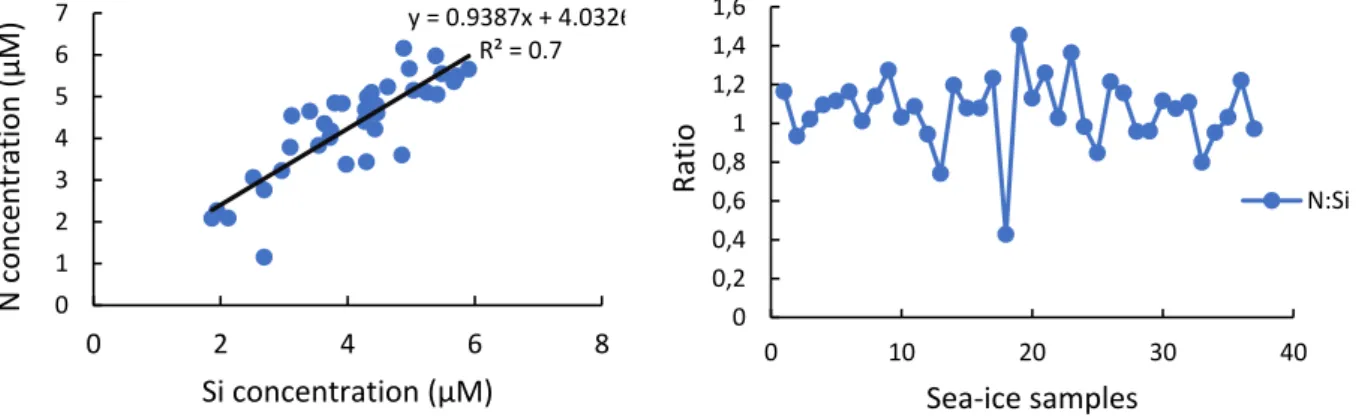
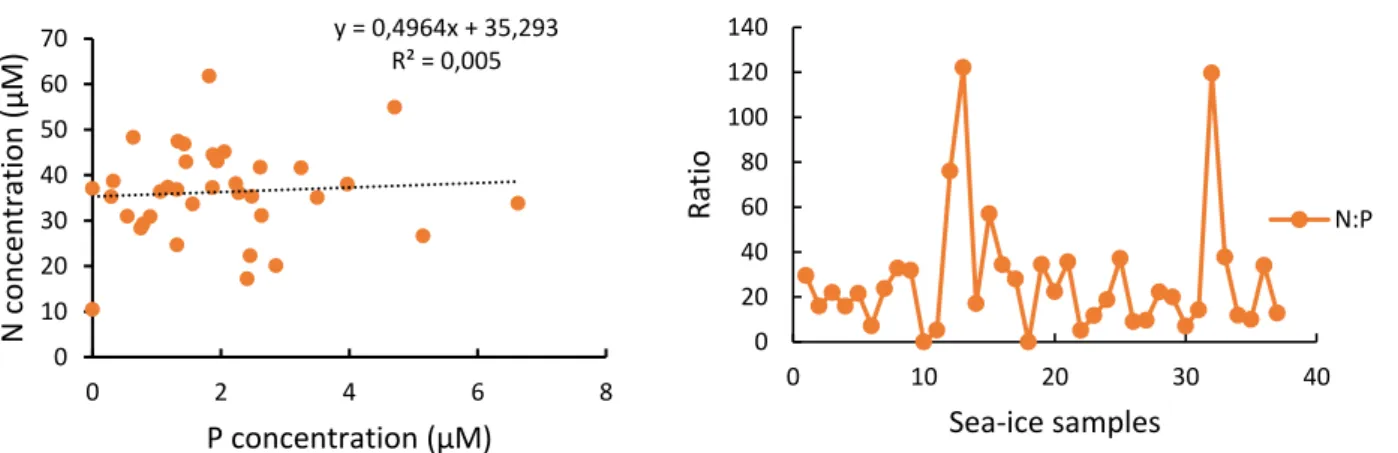
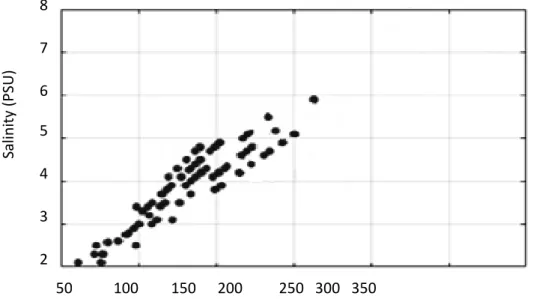
Several significant relationships (p<0.05) were identified between nutrients and sea ice algal biomass (chlorophyll a) when pancake ice flakes were tested, regardless of sea ice depth layers. A significant relationship within the top layer of sea ice samples was found between silicate and large sea ice algal cells. The inner layer showed a significant (p<0.05) negative correlation between nutrients (nitrate and silicate concentrations) and pico-sea-ice algal biomass, as well as nitrite and total chlorophyll a concentration.
The bottom layer also showed a positive correlation between pico-sea-ice algae (p<0.05) and nitrite concentrations. In addition, ciliates had a significant (p<0.05) strong positive association with bottom layer silicate concentration in all sea ice samples.
Discussion
- Biochemical environment
- Size fractionation of sea-ice algae ................................................................ Error! Bookmark not defined
This generated extreme regional variability with respect to MIZ sea ice behavior and morphology (Vichi et al., 2019). The SO sea-ice environment has been depicted as a biogeochemically active reservoir for iron (Lannuzel et al., 2016). This is contrary to what was predicted for the early-winter sea-ice microbial community (Fripiat et al., 2017).
The dominance of larger cells may also be characteristic of a more settled sea ice community (Tison et al., 2020). Larger contributions from nano phytoplankton cells (2–20 µm) to bottom seawater and sea ice in the MIZ and polar frontal zone have been postulated by Detmer & Bathmann (1997), suggesting a lower importance and dominance ( cellular abundance) of autotrophic pico. -seaweed-ice. In the PIPERS study within the central and western Ross Sea MIZ (CRS and WRS) during early winter (Tison et al., 2020).
Species of the genus Fragilaria are often found dominating the sea ice environment (Krell et al., 2008), especially pack ice (Lizotte, 2001) and open waters of Antarctic ice-marginal areas (Kang & Frexyll, 1992). . Contrary to the findings of Fripiat et al. 2017), who hypothesized that sea ice algae are not active during winter on Antarctic sea ice.
76
Methodology
- Study area
- Sample collection and processing ................................................................ Error! Bookmark not defined
All sea ice cores were transported in black plastic bags to avoid light exposure and prevent further photosynthesis. Sea ice samples were then examined for sea ice algae using light microscopy and the chlorophyll a concentration was measured. Biomass of large algal cells (shown as >20 µm) was below 0.2 µg/L for all sea ice and surface water samples.
In addition, the maximum mean biomass (>2.7 µg/L) was recorded for frazil ice samples, which showed similar high concentrations as the consolidated bottom sea ice layers (Figure 3). Most notably, the biomass of the surrounding surface water was significantly (p<0.05) lower compared to sea ice samples (frazil and sea ice) (Figure 3.2). The sea ice algae community was dominated by the class Bacillariophyceae and the order Pennales, i.e.
The number of individual species and the diversity of the surface seawater samples were similar to those of the sea ice samples. In contrast, no significant differences were found between deep layers of sea ice and between surface water and frasile ice (MIZ S) samples.
Discussion
- Sea-ice algae biomass
- Species community structure
However, the accumulation of sea ice algae did not increase from the initial frazil ice cell concentration compared to other sea ice types. Average sea ice algal biomass was significantly higher compared to the surrounding surface water. In addition, species of Fragilariopsis were the most abundant throughout all sea ice and surface water samples.
Similar dominant sea ice algal taxa were observed in this study compared to the 2017 Southern-Indian Ocean MIZ sea ice (Chapter 2). Like the 2017 sea ice algal communities (Chapter 2), centric diatoms were abundant in all sea ice samples, highlighting their dominance in marginal ice edge zones (Horner, 1985). Ice increases in complexity with age, and as a result a more diverse fauna and flora community composition is expected in the sea ice (Hop et al., 2020).
Lack of microhabitats observed in this study may indicate that the sea ice algal community was recently established due to the relatively young ice age. However, dense benthic communities exist when sea ice is relatively young and thick (between 0.4 and 1 m, respectively) (Meiners et al., 2012).
Conclusion
Exploring the spatial heterogeneity of Antarctic sea ice algae using an autonomous vehicle-mounted irradiation sensor. Physiological responses of a small Antarctic diatomaceous earth (Chaetoceros sp.) to simulated environmental constraints associated with sea ice formation. Changes in sea ice protist diversity with declining sea ice in the Arctic Ocean from the 1980s to 2010s.
Exopolymer alteration of sea ice physical properties and implications for ice habitability and biogeochemistry in a warmer Arctic. Reviews and syntheses: Ice acidification, the effects of ocean acidification on sea ice microbial communities. The importance of marine biota to the ecosystem of the northwestern Weddell Sea.
Interannual variability of phytoplankton biomass and species composition in northern Marguerite Bay (western Antarctic Peninsula) is governed by both winter ice cover and summer stratification. 2015: A study of the impact of lead ice formation on sea ice mass balance using a new parameterization of frazil and greasy ice.
99
- Osmoregulation
- Osmolytes
- Sea-ice processing
- Methodology
- Sample collection
- Statistical analysis
- Results
- Discussion
- Species-specific response to melting process
- Temporal effect of melting procedures
- Conclusion
- References
Sea ice algae experience disturbances in cellular homeostasis or osmotic stress under different natural conditions in the SO. This is problematic because the sea ice lives in the brine network (Garrison & Buck, 1986). The sea ice samples were stirred every hour to homogenize the sample and prevent rapid heating.
The sea ice was then examined for the identification of chlorophyll a and sea ice using light microscopy (see Chapter 2). However, species richness, number of species and species diversity were lower in all sea ice samples melted with FSW. The effect of melt treatments on assessment of biomass and nutrients in sea ice (Saroma-ko Lagoon, Hokkaido, Japan).
Comparison between the two winter campaigns revealed clear differences in the biomass and distribution of sea ice algae. Overwintering sea ice algal communities (in both consolidated and semi-consolidated pancake ice) were generally uncorrelated with underlying pelagic phytoplankton communities.