In the study of Al3+ resistance, each of the three tested parameters (genotype, Al3+. concentration and exposure time) had a significant influence on the callus growth rate. It is therefore suggested that Al3+ inhibited meristematic cell number in the Al-S genotype by interfering with cell division.
GENERAL INTRODUCTION
AI in solution
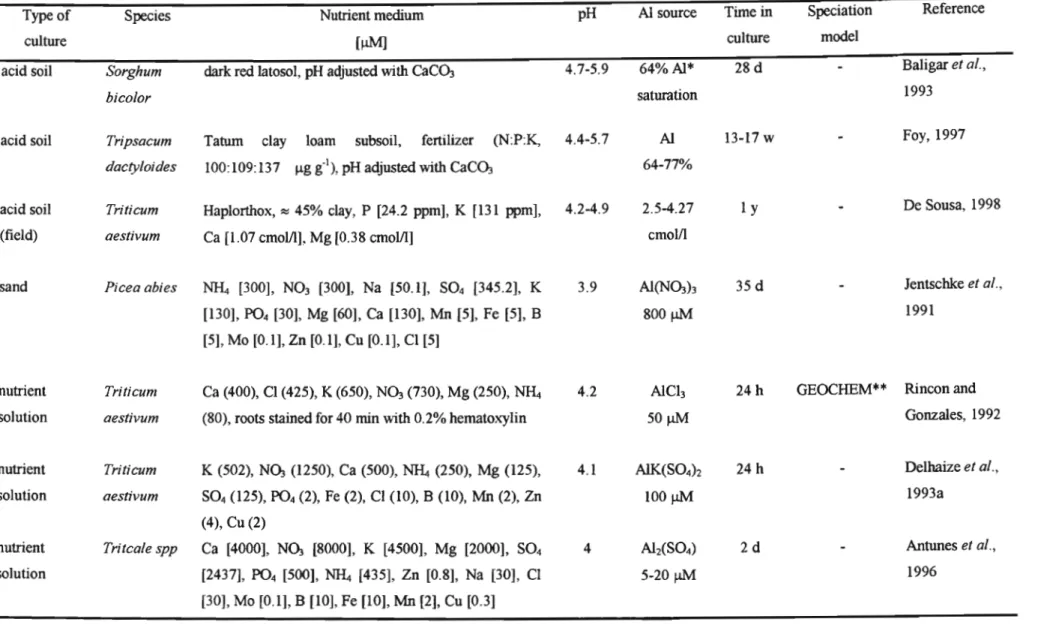
Although Al in solution represents a small fraction of total Al in soil, it is the most important chemically and biologically available form (Driscoll and Schecher, 1990). Hydrolysis of AI3+ involves the progressive loss of hydration shell protons to water molecules in solution (Bertsch and Parker, 1996). The four hydrolysis reactions and the predicted availability of different types of AI at different pH values are shown in Table 1.1. In acidic solutions « pH 4) AI3.

Phytotoxic AI form
Triticum AI-S and AI-T AI exclusion Exudation of AI-chelating ligand Malate efflux by AI-induced anion Zhanget al., 2001. Triticum AI-S and AI-T AI exclusion Exudation of AI-chelating ligands AI3+ activity reduced , reduced RGI Pelletet al., 1997.
Aims and Objectives of this study . 1 Research aims
Further studies were conducted to investigate possible reasons for the greater inhibitory effect of Al3+ on the Al-S than the Al-R genotype (Chapter 4). Finally, the benefits of using an alternative approach to investigate mechanisms of Al3+ toxicity and resistance were considered (Chapter 5).
ESTABLISHMENT OF IN VITRO REGENERATION PROTOCOLS
Discussion
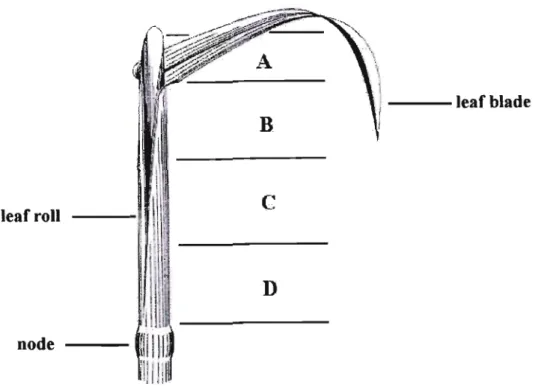
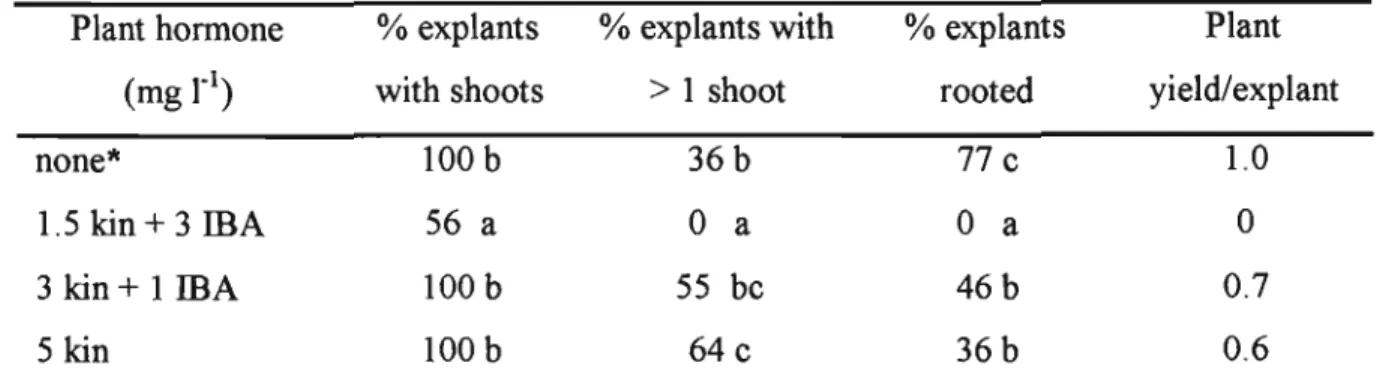
Identifying the different stages of somatic embryo development is therefore important because mutant embryos will not germinate (Dodeman et al., 1997; Fischer et al., 1997). These hormones have been shown to positively influence embryogenic callus formation (Rajasekaran et al., 1987b; Wenck et al., 1988). Murashige and Skoog (1962) based media supplemented with sucrose are commonly used in grass tissue culture (Krishnaraj and Vasil, 1995 ~ Wang et al., 2001).
Although sorbitol (with 2,4-D) was successful for other cultures (Ryschka et aI., 1991; . Okamoto et aI., 1996), this sugar failed to stimulate callus formation in the explants of C. In some other studies an increase in sucrose concentration led to the suppression of non-embryogenic callus and the increased compactness of embryogenic callus (Elkonin et al., 1995, Pedrosa and Vasil, 1996; Saidi et al., 1997), the opposite was found true. in this work. Since dehydration of mature zygotic embryos causes germination and development into normal plants (Gray, 2000; von Arnold et al., 2002), a similar approach for C.
Early studies found that physical drying treatments stimulated germination in Dactylis glomerata (Gray, 1987) and Zea mays (Compton et al., 1992) and this approach has also been used more recently for some gymnosperms (Capuana and Debergh, 1997; . Norgaard, 1997). . Reports in the literature show that in vitro-produced plants are successfully acclimatized to ex vitro growth conditions and develop normally without morphological differences compared to seed-grown plants (Kebebew et al., 1998; Lauzer et al., 2000; Aguado-Santacruz et al. ., 2001; Grando et al., 2002; Wang et al., 2003). In fact, the in vitro regenerants are sometimes found to have better growth patterns than their seed-derived counterpart, as shown by Lolium perenne regenerantes which produced a significantly higher seed yield than the seed-grown plants (Stadelmann et al., 1998).

DEVELOPMENT AND USE OF AN IN VITRO CALLUS CULTURE SYSTEM FOR SCREENING FOR AI3+ RESPONSE
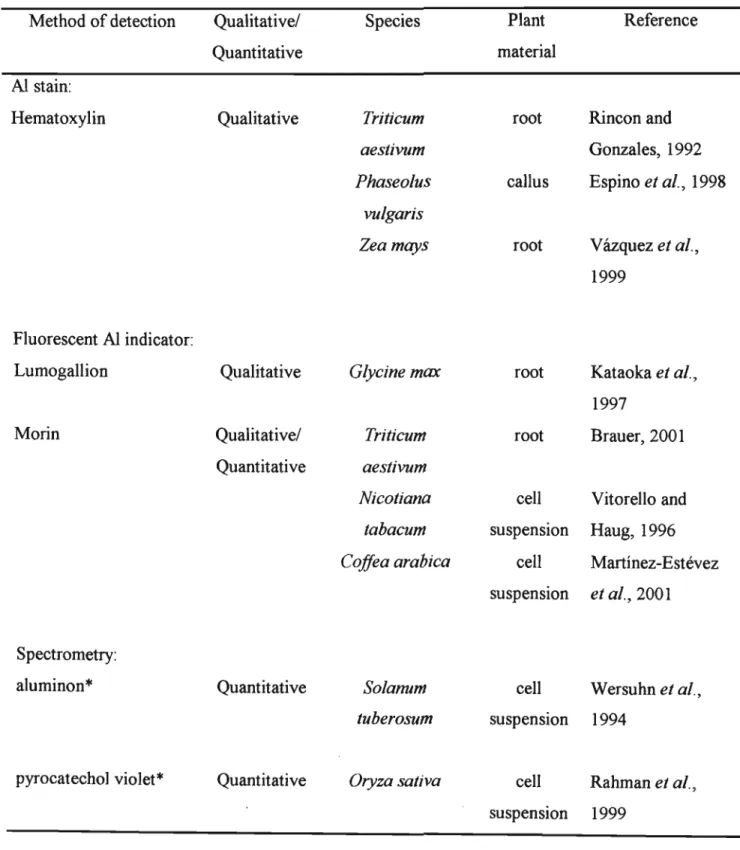
In order to increase the availability of AI in this culture medium, some researchers have modified the composition of the nutrient solution for plant growth. Each of the AI stains forms a complex with AI such that AI-high (AI-S) roots. As in whole plant screening, the use of a simplified salt medium eliminates the interaction of Al with various components of the whole nutrient medium, increases the activity of free Al3+ ions and shortens the cultivation time (Deviet al., 2001; Zhu et al. ., 2003).
The callus growth rate data were transformed (square root) to achieve a normal distribution and the one-sample Kolmogorov-Smirnov normality test (p > 0.05) was applied to the residuals of the transformed data. At neutral pH, most of the aluminum supplied is available as Al(OH)3 and (AI(OHk). As with AICh, approximately 60% of the aluminum supplied forms an aluminate ion (Al(OH)n at pH 7.
The chemical interactions of aluminum were modeled with the components of the nutrient media and the resulting Al3+ activity was predicted with increasing aluminum supply (Figure 3.3). Simulations of the MINTEQA2 model showed that when using the tested media, the maximum Al3+ activity achieved is 7. In addition, the confidence limits for the mean callus growth rate (in the absence of Al3+) were calculated for each of the.
Genotype
Other interactions showed that the change in callus growth rate over time differed among the three genotypes and that the negative effects of increasing Al3+ concentration on callus growth rate appeared more pronounced with longer culture time than after a short culture interval. When exposed to Al3+, the response of each of the three genotypes was different from each other. It was clear that the growth rate of callus exposed to 0.8, 1.9, and 2.3 mM Al3+ were all significantly lower than that of the control, but not significantly different from each other.
Also, at each culture interval, there was no significant difference in the growth rate of callus exposed to 0.08 mM Al3+. In genotype 2, at all time intervals tested, except for 4 weeks, exposure to 0.08 mM Al3+ resulted in a significantly lower growth rate than the control. Furthermore, similar to genotype 1, there were no significant differences in the growth rate of callus exposed to 0.8, 1.9 and 2.3 mM Al3+ at a time mterva .h · · I.
In genotype 3, after 2 weeks of culture, only the highest AI3+ concentration tested (2.3 mM) led to a significant reduction in callus growth rate compared to the control. As in genotype 1, the lowest Al3+ concentration had no effect on callus growth rate at all tested time intervals. Only after 6 and 8 weeks of cultivation was the growth rate of callus exposed to 0.8, 1.9 and 2.3 mMAl3+ significantly lower than the control.
Genotype 2
Callus was initiated in the first culture phase of the somatic embryogenesis protocol (section 2.3.2). In the present study, a speciation model (MINTEQA2) was used to evaluate the merits of published Al screening media, including that of Conner and Meredith (1985a) (Figure 3.3). This component of the MS nutrient formulation was reduced as a decrease in Mg2+ did not have a negative effect on callus growth in the control medium (Figure 3.5).
In these studies, the long culture time could be attributed to a low availability of free AI3+ ions due to interactions of AI with components of the culture medium. Thus, in the development of AI screening media for callus cultures, it is important to ensure both an available supply of free AI3+ as well as sufficient nutrient levels for active callus growth. In this study, it was also possible to differentiate between the three genotypes using the frequency of non-embryogenic callus production (Figure 3.9).
It is possible that low pH in the absence of AI would adversely affect the in vitro response of some species (ie, callus growth). This does not seem to be the case in this study, as the control callus (-AI, pH 3.6) maintained a high growth rate throughout the culture time investigated (Figure 3.6). In this work, AI-S and AI-R C could be distinguished using differences in callus growth.

RESPONSES OF AI-SENSITIVE AND AI-RESISTANT MERISTEMATIC CALLUS CELLS TO AI3+
- Discussion
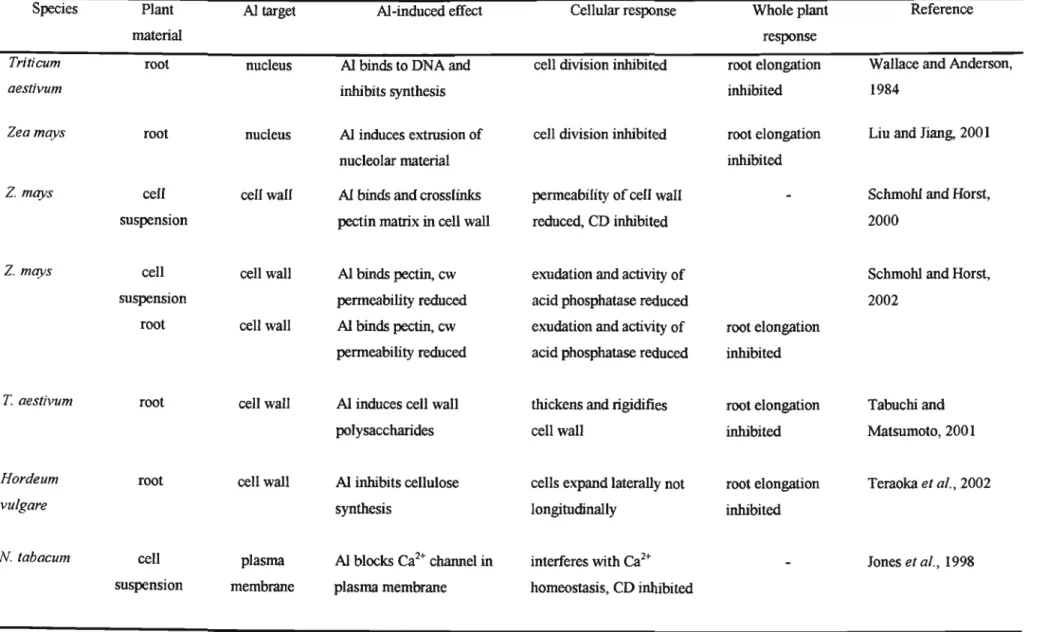
In the Al-R genotype, only the highest concentration of colchicine (2.5 mM) resulted in any significant reduction (36%) in embryogenic cells. In both genotypes, none of the treatments resulted in a significant increase in the number of non-embryogenic cells, except for exposure to 2.5 mMcolchicine in the Al-R genotype (Table 4.4). The results show that a significantly higher concentration of Al was found in Al-S than Al-R genotype (Table 4.6).
Furthermore, the medium supporting the Al-R genotype had a 5.9-fold lower Al3+ concentration at the center of the Petri dish than the Al-S medium. Thus, it is possible that a similar AI resistance strategy is evident in the AI-R genotype. Furthermore, increased AI content in the nuclei of AI-S cells disrupted cell division and consequently inhibited root growth (Crawford et al., 1998).
Some researchers have shown that AI is preferentially located in actively growing cells: 1) DTZ cells in the maize root tip (Sivaguru et al., 1999a). In this study, it is possible that the inhibited cell division in the AI-S genotype occurred as a result of microtubule disruption of the meristematic callus cells. Furthermore, although there were no differences in cytometric AI content between the two genotypes tested in this study (Table 4.7), the AI-S genotype accumulated three times more AI in the nucleus compared to the AI-R genotype.
FINAL DISCUSSION
Ahn BJ, Huang F.H and King lW 1985 Plant regeneration through somatic embryogenesis in conventional Bermuda grass tissue culture. Cardona CA and Duncan RR 1997 Callus induction and highly efficient plant regeneration through somatic embryogenesis in Paspalum. Chandler S.F. and Vasil i.K. 1984 Optimization of plant regeneration from long-term embryogenic callus cultures of Pennisetum purpureum Scum.
De la Fuente-Martinez J.M and Herrera-Estrella L 1999 Advances in the understanding of aluminum toxicity and the development of aluminum-resistant transgenic plants. Jackson LA, Dalton S.J and Dale P.J 1986 Regeneration of plants from root callus in the forage grass Lolium multiflorum. Miyasaka S.C. and Hawes M.C. 2001 A possible role for root border cells in the detection and avoidance of aluminum toxicity.
Pedrosa LF and Vasil 1.K 1996 Optimization of somatic embryogenesis and long-term regeneration in callus cultures of Diploperennial teosinte (Zea diploperennis litis, Doebley and Guzman). Pellet D.M, Grunes D.L and Kochian L.V 1995 Organic acid secretion as an aluminum tolerance mechanism in maize (Zea maysL.). Pifieros M.A and Kochian L.V 2001 A patch-clamp study on the physiology of aluminum toxicity and aluminum tolerance in maize.
Polle E, Konzak C.F and Kittrick LA 1978 Visual detection of aluminum tolerance levels in wheat by hematoxylin staining of seedling roots. Sivaguru M, Baluska F, Volkmann D, Felle H.H and Horst WJ 1999a Effects of aluminum on the cytoskeleton of the maize root tip.