In the first, I present data from equicaloric choice experiments to determine sugar preferences of both specialist and occasional nectar-feeding birds over a range of concentrations. In the second section of the thesis, I investigate the associations between plant characteristics and nectar-preferential nectar preferences.
Introduction
In addition to these findings in the botanical literature, researchers began to question the relevance of the results of choice test experiments to determine the sugar preferences of nectar-feeding birds (Fleming et al. 2004). 5% higher energy value for sucrose over the equivalent weight of the hexose sugar (Bumstead 1980; Fleming et al. 2004). Methods for investigating the sugar preferences of avian nectarivores are receiving critical reexamination (Fleming et al Brown et al. 2008).

SUGAR PREFERENCES OF A GENERALIST NON- PASSERINE FLOWER VISITOR, THE AFRICAN
SPECKLED MOUSEBIRD
This has led to a growing interest in sugar preferences and apparent digestive efficiency in both passerine and non-passerine occasional avian nectarivores (Lotz and Schondube 2006; Brown et al. 2008; Brown et al. 2010). Common African representatives include starlings, weavers, white-eyes, voles, buzzards, barbets, orioles and drongos (Oatley1964; Symes et al. 2008). Preliminary research on sugar type preferences in occasional passerines has revealed consistent preferences for hexose sugars at low concentrations and either hexose preference or no preference at high concentrations (Brown et al. submitted; in press; Odendaal et al. in press).
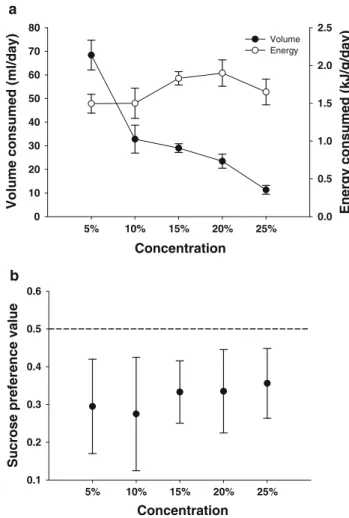
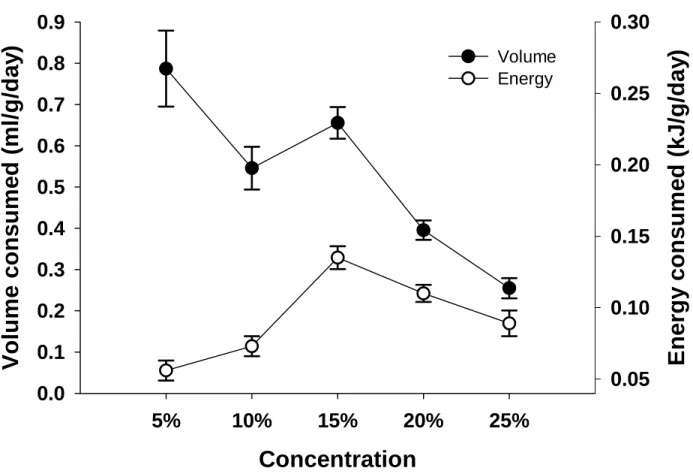
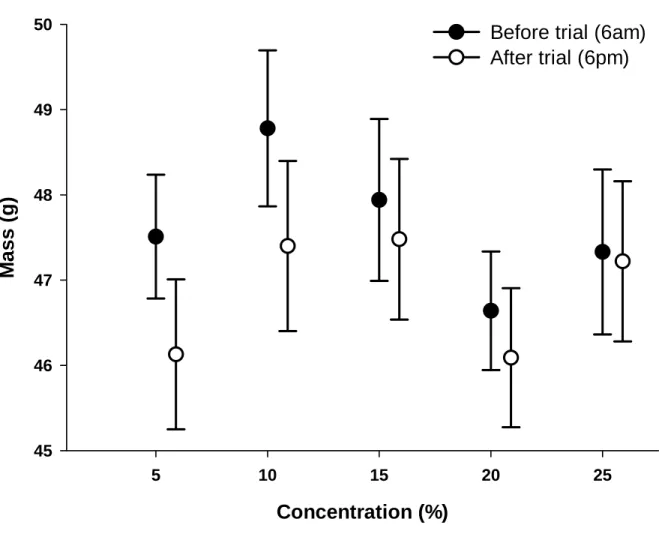
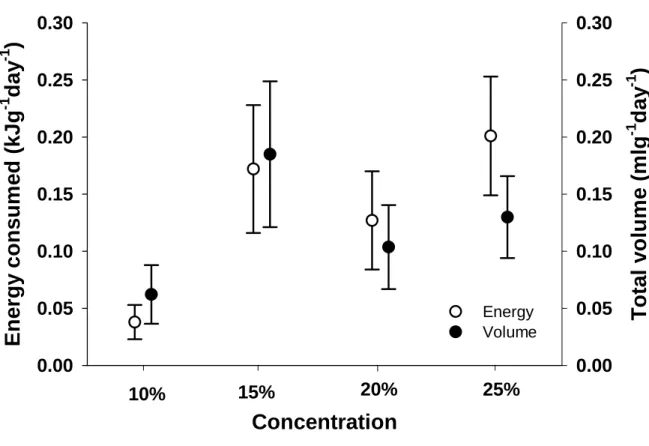
Most of this research was conducted using a single concentration, and equal or equimolar solutions (see Brown et al. 2008). Spotted mousebirds are omnivores and feed on fruit, nectar, insects, and leaves (Downs et al. 2000). We also converted the total volume consumed of each solution to energy consumed, expressed as kJ g-1 day-1 (Brown et al. 2008).
Volume of each solution drunk was recorded to determine a daily intake and converted to energy consumption per g per day (Brown et al. 2008). We compared energy intake (kJg-1day-1) and volume consumption (mlday-1) at each of the four hexose concentrations offered simultaneously to birds in the concentration trial using RMANOVA to determine whether a concentration preference was evident ( Brown et al. in press).
Concentration
Sugar concentration
ConcentrationEnergy consumed (kJg-1day-1)
Total volume (mlg-1day-1)
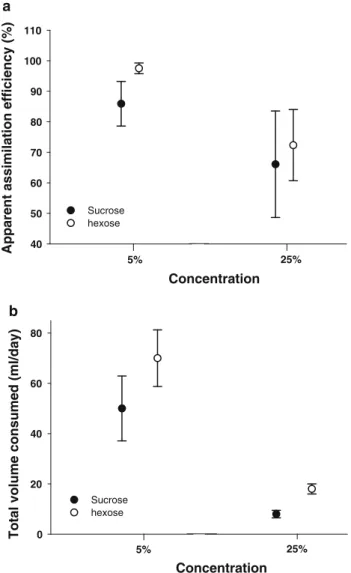
This contrasts with results obtained for passerines with occasional nectarers, which generally show either hexose preferences at all concentrations (Brown et al. in press) or hexose preference at low concentrations with no preference at high concentrations (Brown et al. submitted; Odendaal et al. in press). Speckled buzzards mirror that of both passerines and non-passerine specialist nectar birds (Fleming et al. Brown et al. 2010). Speckled Mousebirds efficiently assimilated both types of sugar when fed low- and high-concentration solutions, which is similar to specialized nectar-feeding birds (Markman et al. 2006; McWhorter and Lopez-Calleja 2000).
This is similar to results found in other occasional bird-nectar species (Brown et al. in press; submitted; Odendaal et al. in press). Rather surprisingly, spotted voles showed a pattern of sugar preference across a range of concentrations that was similar to all specialist nectarivores studied to date, both passerines and non-passerines (Fleming et al. Brown et al. 2010). In contrast, occasional nectar-feeding passerines that have been studied so far show varying patterns of sugar preference (at a range of concentrations) that are undoubtedly associated with varying differences in apparent assimilation efficiencies of sucrose solutions and the ability to increase volumetric intake to accommodate the daily energy intake at low. concentrations (Brown et al. under pressure;.
Previous research on hummingbirds suggests that adaptation, not phylogeny, is responsible for differences in sugar preferences, concentration preferences, and apparent assimilation efficiency between specialist and occasional nectarivores (Fleming et al. Brown et al. 2010, in press). Interpretation of nectar properties of flowers pollinated by generalist avian nectarivores: sugar preferences and digestive efficiency of the African weaver bird.
African Red-winged Starlings feed on nectar, but don’t like it too sweet
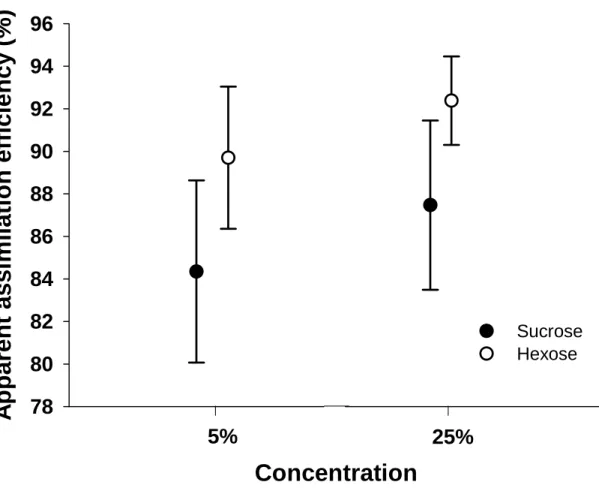
Plants pollinated by these birds possess a set of floral traits (floral morphology, nectar volume and concentration and sugar type) that are distinct from plants pollinated by specialist avian nectarivores (Botes et al. 2008; Johnson & Nicolson 2008; Symes & Nicolson 2008; Symes & Nicolson 2008) . Nicolson 2008; Brown et al. 2009). However, if we exclude specialized passerine nectarivores (which generally have an assimilation efficiency for sucrose of >95%) and passerines of the Sturnidae-Muscicapoidea genus (which lack sucrase), we find that the assimilation efficiency of sucrose is generally lower than that of hexose sugars in passerines (Lane 1997; Brown et al. in press; Odendaal et al. in press; Wellmann & Downs 2009). In this study, we investigated the sugar preference, concentration preference and assimilation efficiency of red-winged starlings over a range of concentrations (5–25%) that reflect nectar concentrations in bird-pollinated plants (Johnson & Nicolson 2008).
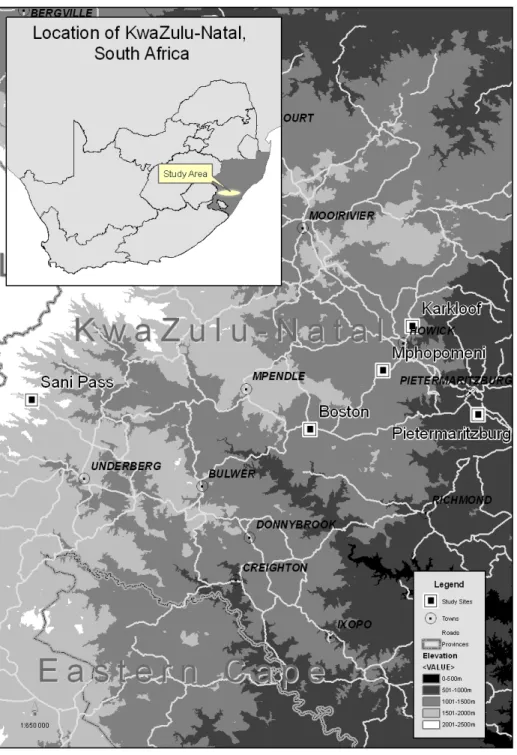
We captured nine Red-winged Starlings using the Ecotone mist net at Waterfall, KwaZulu-Natal (2944S 3049E) in February 2008. See Fleming et al. Solutions were delivered in modified 50 ml burette tubes calibrated at 0.1 ml intervals, alternating position at midday to avoid lateral displacement ( Jackson, Nicolson & Lotz 1998b ; Fleming et al. 2004 ).
We also converted the total volume of each solution consumed to the energy consumed per gram of body weight per day, expressed as kJg-1day-1 (Brown et al. 2008). The volume of each solution consumed was converted into energy expended per gram of body weight per day (Brown et al. 2010).
Energy consumed (kJg-1day-1)
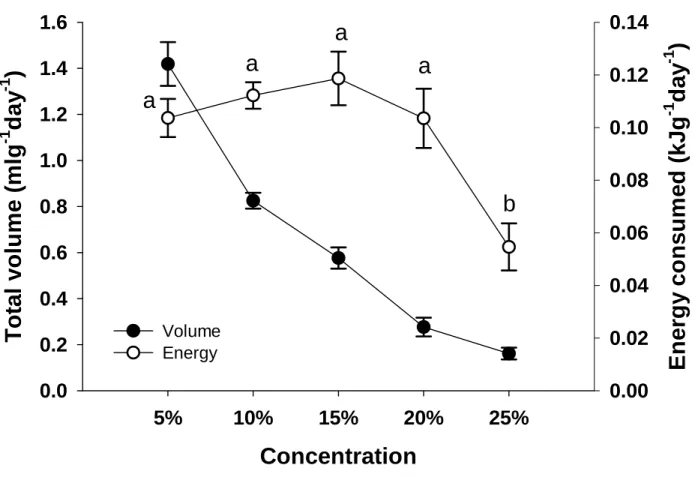
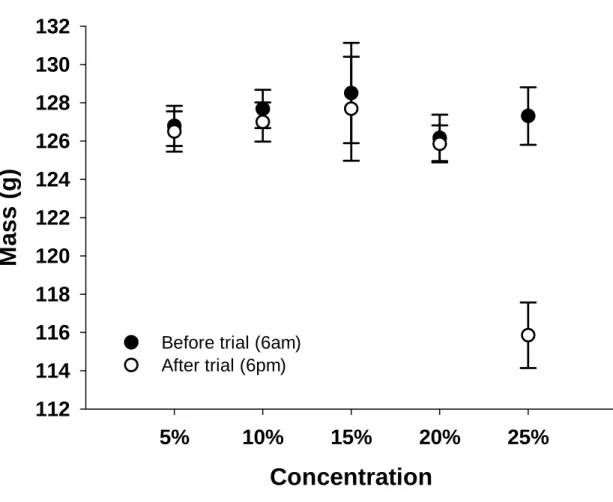
Both black-headed starlings (Brown et al. in press) and red-winged starlings drank less of the 25% solution (by volume) when given a choice of four different concentrations, and most strikingly, red-winged starlings were unable to maintain energy balance ( and actually significant body weight lost) at this high concentration. The results of this study and those of other similar studies on sparrows, occasional avian nectarivores (Brown et al. in press; Odendaal et al. in press) are beginning to help evolutionary biologists explain the phenomenon of nectar-dominant, low-concentration hexose pollinators in plants. From data obtained in bird elimination experiments, Symes et al. this volume) suggest that bees may be more important than sunbirds as pollinators of A.
A recent study of Kniphofia caulescens Baker, a species with relatively shallow flowers (about 20 mm depth), identified short-billed generalist birds as pollinators (Brown et al., 2009). Johnson and Brown (2004) made anecdotal observations of visits by Malachite Sugarbills (Nectarinia famosa) to flowers of Kniphofia laxifloraKunth in one population, while Johnson et al. To assess the efficiency of birds as pollinators, we collected pollen loads from birds that were miscaptured in the Curry's Post population.
Because our study sites were more representative of the typical habitat of K. laxiflorathan, the mountainous sites were selected by Johnson et al. Although few Kniphofia species have been studied to date (Brown et al., 2009; Johnson et al., 2009; current study), it appears that sugar composition is phylogenetically restricted in the genus.
Covariation of flower traits and bird pollinator assemblages among populations of Kniphofia
Despite recent molecular and morphological studies, the phylogenetics of this group is still problematic (Ramdhani et al. 2006). We conducted controlled pollination experiments at the Sani Pass and Pietermaritzburg populations to determine the compatibility system and dependence of K. We conducted exclusion experiments at the Sani Pass and Pietermaritzburg populations to determine the effectiveness of different pollinator groups.
This was then fused to microscope slides in the field (Brown et al. 2009) and compared to K. Corolla reference slides, there was a significantly longer length at Sani Pass and Boston than at Mpophomeni, Karkloof and Pietermaritzburg (Fig. 3a). Plants at Sani Pass were visited only by specialist avian nectarivores (3 species), and those in Boston by specialists (2 species) and one species of generalist avian nectarivore.
Pollen loads obtained by netted birds at Sani Pass and Pietermaritzburg were very high (Table 2). Fruit set in both ecotypes were similar to those recorded in other Kniphofia species pollinated by generalist (Brown et al. 2009) and specialist avian nectarivores (Brown et al. in press).
Can bird sugar preferences explain plant nectar properties?
Indeed, specialists show a strong preference for solutions with the highest sugar concentrations when given the choice (chapter 3). However, occasional avian nectarivores show a preference for low-concentration solutions, or lack a concentration preference, and are able to maintain energy balance on low-concentration solutions. This, coupled with their preference for hexose solutions, suggests that they may be responsible for the evolution of high amounts of hexose sugars in the low concentration of nectar in plants they pollinate (Johnson & Nicolson 2008).
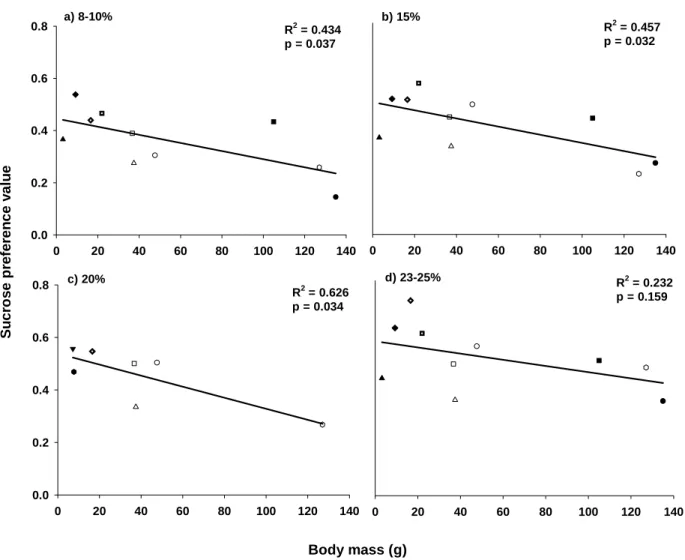
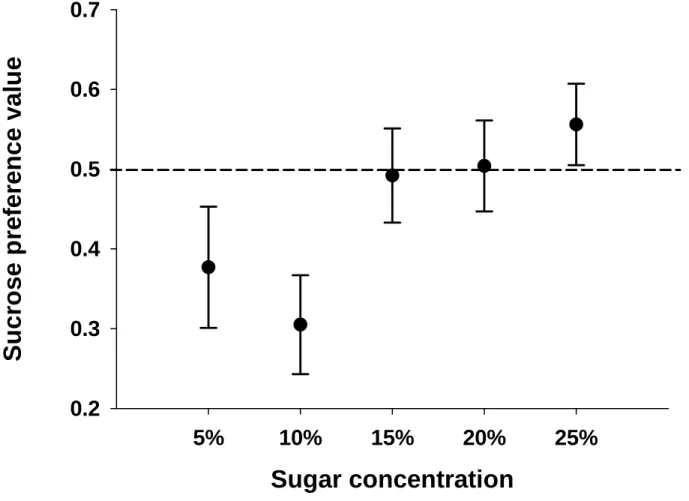
Since bird sugar preferences are known, in many cases, to depend on solution sugar concentrations, it is necessary to statistically separate the effects of bird feeding area and sugar concentration when trying to explain sugar preference. I then used Analysis of Covariance (ANCOVA), using type III sum of squares, with 12 species (8 specialists and 4 occasional nectarivores nested within these two food niches) and sugar solution concentration as covariates. The response variable was the arcsine square root transformed sucrose preference data for solutions ranging from 3–25% sugar concentration ( Figure 1 ).
This result thus raised questions about the validity of the dietary niche hypothesis developed by Johnson and Nicolson (2008). I then developed a new hypothesis (the 'body size hypothesis') considering the relationship between body size and sucrose preference for these 12 species using regression analysis for four different concentrations (Figure 2).
Concentration (%)
These studies indicate that specialized passerines such as blackbirds have sugar preferences and digestive abilities similar to hummingbirds (Fleming et al., 2004; Lotz and Schondube, 2006). Brown et al., 2010b), comparisons have only been reported at single concentrations, and these concentrations have not been representative of the highly diluted nectar found in flowers pollinated by these birds. Also, test solutions in previous studies were not equicaloric; thus conflating tests of sugar preference with an energy difference between test solutions (Brown et al., 2008).
The apparent adsorption efficiency of the two sugars was tested indirectly following a method similar to that used by Jackson et al. Importantly, birds were able to maintain body mass at low concentrations and meet daily energy requirements, similar to dark-capped bulbuls (Brown et al., 2010b). However, unlike bird species that cannot assimilate sucrose at all (Muscicapoidea superfamily), and thus avoid sucrose solutions altogether (Gatica et al., 2006;.
It seems likely that differences in nectar characteristics between plants pollinated by specialist avian nectarivores and those pollinated by generalists are due to differences in selection imposed by these two groups of birds, but this remains poorly understood (Fleming et al. , 2004). Brown et al., 2010b), reveals that the sugar preferences and digestive abilities of occasional nectarivores such as village weavers and dark-capped bulbuls, when offered solutions of biologically realistic concentrations, can explain the evolution of the hexose-dominated nectar in plants pollinated by these birds.