KARAKTERISASI Chili veinal mottle virus
STRAIN LEMAH DAN POTENSINYA
SEBAGAI AGENS PROTEKSI SILANG
ASNIWITA
SEKOLAH PASCA SARJANA INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Karakterisasi Chili veinal mottle virus Strain Lemah dan Potensinya sebagai Agens Proteksi Silang
adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2013
Asniwita
ASNIWITA. Karakterisasi Chili veinal mottle virus Strain Lemah dan Potensinya sebagai Agens Proteksi Silang. Dibimbing oleh SRI HENDRASTUTI HIDAYAT, GEDE SUASTIKA, SLAMET SUSANTO, dan SRIANI SUJIPRIHATI.
Chili veinal mottle virus (ChiVMV) merupakan salah satu penyebab penyakit yang penting pada tanaman cabai. Infeksi ChiVMV menyebabkan tanaman mengalami perubahan antara lain daun menjadi belang hijau tua, malformasi, ujung daun meruncing, lamina menyempit, dan tanaman kerdil. Hingga saat ini belum ada teknik pengendalian yang efektif, sehingga perlu dilakukan evaluasi pengendalian penyakit melalui proteksi silang. Proteksi silang merupakan teknik pengendalian secara biologi dengan menggunakan virus strain lemah yang sekerabat dengan virus penyebab penyakit (strain kuat).
Penelitian ini bertujuan untuk (i) mendapatkan ChiVMV strain lemah dari pertanaman cabai, (ii) mempelajari kisaran inang ChiVMV strain lemah, (iii) mempelajari interaksi antara ChiVMV strain lemah dengan strain kuat, (iv) menguji kemampuan ChiVMV strain lemah dalam melindungi tanaman cabai dari infeksi ChiVMV strain kuat melalui teknik proteksi silang, dan (v) mengkarakterisasi secara molekuler ChiVMV strain lemah.
Chili veinal mottle virus isolat lemah diperoleh dari pertanaman cabai pada beberapa daerah di Provinsi Jambi, Sumatera Barat, dan Jawa Barat. Isolat ChiVMV yang diperoleh dari lapangan diseleksi melalui uji biologi secara bertahap. Pertama-tama, isolat-isolat ChiVMV tersebut ditularkan secara mekanis ke tanaman cabai IPB C13. Isolat-isolat yang mampu menginfeksi tetapi tidak menimbulkan gejala digunakan untuk pengujian kisaran inang yang mencakup 8 genotipe cabai dan 11 spesies tanaman Solanaceae. Isolat-isolat yang mampu menginfeksi tetapi tidak menimbulkan gejala digunakan untuk pengujian selanjutnya yaitu evaluasi interaksi ChiVMV isolat lemah dengan isolat kuat. Berdasarkan hasil evaluasi tersebut diseleksi beberapa isolat yang digunakan pada tahap evaluasi proteksi silang. Konfirmasi infeksi ChiVMV pada tahapan-tahapan seleksi isolat ChiVMV dilakukan dengan uji serologi DAS-ELISA, DIBA, atau TBIA. Tahap akhir penelitian adalah melakukan karakterisasi isolat-isolat lemah ChiVMV yang potensial, yaitu menggunakan metode RT-PCR dan analisis keragaman genetik (sikuensing).
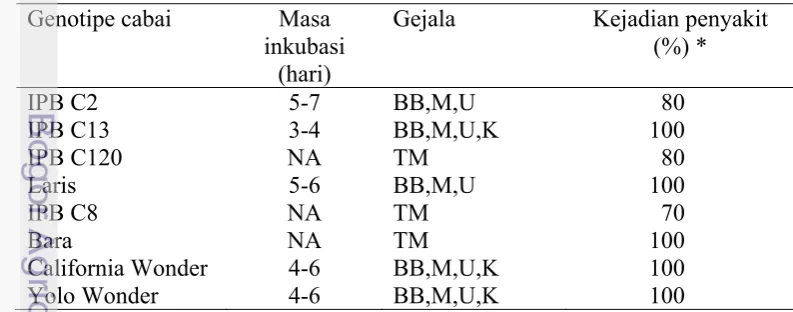
Hasil penelitian menunjukkan bahwa dari 223 sampel daun cabai diperoleh 30 sampel yang terinfeksi tunggal ChiVMV. Setelah inokulasi ke tanaman cabai merah IPB C13 diperoleh 16 isolat lemah, 2 isolat sedang, dan 3 isolat kuat. Berdasarkan hasil inokulasi ChiVMV isolat lemah ke berbagai jenis tanaman cabai dan tanaman Solanaceae diperoleh 8 isolat yang mampu menginfeksi dan
tidak menimbulkan gejala pada tanaman cabai Capsicum annuum (IPB C2, IPB
C13, IPB C120, Laris, IPB C8, Bara, California Wonder, dan Yolo Wonder) dan
tanaman C. frutescens, Datura metel, D. stramonium, Lycopersicon esculentum,
Physalis floridana, Solanum melongena, dan S. nigrum. Pada pengujian kisaran
inang ini semua ChiVMV isolat lemah tidak dapat menginfeksi Nicotiana
seminggu sebelum inokulasi isolat kuat ChiVMV-CKB menunjukkan penekanan keparahan penyakit hingga 97.23%. Dengan demikian dapat dikatakan bahwa ChiVMV isolat lemah dapat menekan ekspresi gejala yang disebabkan oleh isolat kuat ChiVMV-CKB.
Pada pengujian proteksi silang ternyata 5 ChiVMV isolat lemah (KAR, SPR, SKT, CSR, dan PGL) efektif melindungi tanaman cabai dari infeksi isolat kuat ChiVMV-CKB. Tanaman cabai yang diinokulasi isolat lemah sebelum inokulasi isolat kuat tidak menunjukkan gejala atau belang ringan, gejala muncul lebih lambat sampai 110 hari, ekspresi gejala lebih ringan, dan tidak mempengaruhi produksi. Tanaman yang tidak diberi perlakuan isolat lemah menunjukkan gejala yang berat, tidak menghasilkan buah atau buah sangat sedikit (rata-rata jumlah buah pada berbagai waktu inokulasi adalah 0.33 sampai 7.33), ukuran buah kecil, dan bobot buah sangat rendah (rata-rata bobot buah pada berbagai waktu inokulasi adalah 0.53g sampai 13.01g). Tanaman yang tidak diberi perlakuan isolat lemah menunjukkan keparahan penyakit 0% sampai 98.61% pada 28 hari, meningkat 55.55% sampai 100% pada 112 hari; sebaliknya tanaman yang diinokulasi isolat lemah sebelum inokulasi isolat kuat menunjukkan keparahan penyakit 0% pada 28 hari, meningkat 1.38% sampai 8.33% pada 112 hari. Inokulasi ChiVMV isolat lemah 7 hari sebelum inokulasi isolat kuat ChiVMV-CKB sudah dapat melindungi tanaman cabai dari infeksi isolat kuat yang menunjukkan produksi berbeda nyata dengan tanaman yang tidak diproteksi (inokulasi dengan isolat kuat ChiVMV-CKB saja). Inokulasi ChiVMV isolat lemah 14 hari sebelum inokulasi dengan isolat kuat ChiVMV-CKB menunjukkan produksi tidak berbeda nyata dengan tanaman yang diinokulasi hanya dengan isolat lemah saja, kecuali pada isolat lemah ChiVMV-KAR.
Amplifikasi DNA ChiVMV isolat lemah dengan RT-PCR menggunakan pasangan primer ChiVMV F Ind dan ChiVMV R Ind diperoleh fragmen DNA gen CP berukuran 900 bp. Analisis kesamaan sikuen membuktikan bahwa kelima ChiVMV isolat lemah mempunyai kekerabatan yang sangat erat satu sama lain dengan tingkat kesamaan sikuen nukleotida dan asam amino masing-masing berturut-turut 92.8% sampai 99.6% dan 90.5% sampai 99.6%. Isolat-isolat lemah tersebut mempunyai tingkat kesamaan sikuen nukleotida dan asam amino terhadap isolat kuat ChiVMV-CKB masing-masing berturut-turut 92.6% sampai 98.1% dan 90.8% sampai 97.5%. Analisis filogenetik menunjukkan bahwa kelima ChiVMV isolat lemah dan isolat kuat ChiVMV-CKB berada dalam satu kelompok.
Berdasarkan evaluasi yang dilakukan dapat disimpulkan bahwa ChiVMV isolat lemah (KAR, SPR, SKT, CSR, dan PGL) mempunyai potensi sebagai agens proteksi silang karena dapat menekan infeksi isolat kuat ChiVMV-CKB dan mempertahankan produksi tanaman. Selain itu, isolat-isolat lemah ChiVMV tersebut tidak menyebabkan penyakit pada tanaman lain dan memiliki kekerabatan yang dekat dengan isolat kuat ChiVMV-CKB sehingga sangat efisien sebagai agens proteksi silang.
Kata kunci : cabai, Chili veinal mottle virus, isolat kuat, isolat lemah, proteksi
ASNIWITA. Characterization of Weak Isolates of Chili veinal mottle virus and Its Potency as Agents for Cross Protection. Supervised by SRI HENDRASTUTI HIDAYAT, GEDE SUASTIKA, SLAMET SUSANTO, and SRIANI SUJIPRIHATI.
Chili veinal mottle virus (ChiVMV)is considered as an important factors for production of chili pepper in Indonesia. Disease control strategies have been evaluated previously involving resistant cultivars, insecticide application, and several cultural practices. Cross protection approach has been implemented for many plant viruses, therefore it is interesting to evaluate its effectiveness for controlling ChiVMV. Cross protection in plant viral disease is known as the phenomenon where the plants systematically infected with one strain of a virus are protected from infection by a second related strain of the same virus.
The objectives of this research are: (i) to obtain weak isolates of ChiVMV; (ii) to study host range of weak isolates of ChiVMV; (iii) to study interaction between weak isolates and severe isolate of ChiVMV; (iv) to study ability of weak isolates in cross protection; (v) to determine molecular character of potential weak isolates of ChiVMV.
Weak isolates of ChiVMV was obtained from chili pepper fields in West Sumatera, Jambi, and West Java. Field isolates were selected using biological assays : (1) Isolates of ChiVMV were transmitted mechanically to chili pepper genotipe IPB C13 to screen isolates that could infect the plants without showing any symptoms. (2) The selected isolates were used in next experiment, i.e. to evaluate their host range involving 8 chili pepper genotipes and 11 plant species in the family Solanaceae. (3) Based on the second experiment the selected isolates were used in further experiments, i.e. to evaluate the interaction between weak isolates and severe isolate of ChiVMV. The presence of ChiVMV was confirmed through DAS-ELISA, DIBA, or TBIA. Finally, molecular characterization of weak isolates was done by sequencing the coat protein gene .
Out of 223 field samples, 30 samples were positively reacted in DAS-ELISA with only ChiVMV antibody. Based on the response on chili pepper genotype IPB C13, ChiVMV isolates can be differentiated into weak isolates (16 samples), mild isolates (2 samples), and severe isolates (3 samples). Based on host range study, 8 isolates out of 16 weak isolates were selected for further evaluation. These isolates were able to cause systemic infection without showing
visible symptom in Capsicum annuum (IPB C2, IPB C13, IPB C120, Laris, IPB
C8, Bara, California Wonder, and Yolo Wonder), C. frutescens, Datura metel, D.
stramonium, Lycopersicon esculentum, Physalis floridana, Solanum melongena, and S. nigrum.
symptoms up to 110 days after inoculation with severe isolate and had no detrimental effects on yield and quality of chili pepper On the other hand, all plants inoculated with only severe isolate ChiVMV-CKB showed severe symptoms, i.e. dark green mottle, malformation, shoestring, stunting, and low yield (average fruit 0.32 to 7.33 per plant with average weight of 0.53 to 13.01 g per fruit). Inoculation only with severe isolate ChiVMV-CKB caused disease severity 0% to 98.61% at 28 days and increased to 55.55% to 100% at 112 days. Inoculation with weak isolate prior severe isolate was able to protect the plants from severe isolate, showed by no infection at 28 days and disease severity reached only 1.38% to 8.33% at 112 days. Inoculation with weak isolates 14 days prior inoculation with severe isolate showed no significant effect on yield, except weak isolate ChiVMV-KAR.
Molecular characterization of ChiVMV isolates was conducted based on analysis of their coat protein gene. Fragment of coat protein gene (900 bp) was successfully amplified using universal primers ChiVMV F Ind and ChiVMV R Ind. Sequence analysis of the coat protein gene showed high homology between 5 weak isolates with nucleotide and amino acid identity varied from 92.8% to 99.6% and 90.5% to 99.6% respectively. Similarly, the weak isolates have high percent similarity of their nucleotide and amino acid with severe isolate ChiVMV-CKB i.e. 92.6% to 98.1% and 90.8% to 97.5%, respectively. Further more phylogenetic tree analysis showed that the weak isolates were in the same group with severe isolate ChiVMV-CKB.
As conclusion, 5 weak isolates of ChiVMV (KAR, SPR, SKT, CSR, and PGL) were identified as potential agents for cross protection due to their ability to protect the plant from infection of severe isolate ChiVMV-CKB. The selected weak isolates were also capable to maintain yield, showed no evidence to cause diseases on other plants and showed a close genetic relationship with severe isolate.
Key words: Chili veinal mottle virus, chili pepper, cross protection, severe
© Hak Cipta milik Institut Pertanian Bogor, tahun 2013
Hak Cipta dilindungi Undang-Undang
1.Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a.Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
b.Pengutipan tidak merugikan kepentingan yang wajar IPB
KARAKTERISASI Chili veinal mottle virus
STRAIN LEMAH DAN POTENSINYA
SEBAGAI AGENS PROTEKSI SILANG
ASNIWITA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Fitopatologi
SEKOLAH PASCA SARJANA INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Dr Ir Suryo Wiyono, MSc. Agr
Dr Ir Sandra Arifin Aziz, MS
Penguji pada Ujian Terbuka: Dr Ir Abdul Muin Adnan, MS
Dr Ir Sri Sulandari, MS
Nama NIM
Program Studi
: Asniwita : A362080011 : Fitopatologi
Disetujui
Komisi Pembimbing
Dr Ir Sri Hendrastuti Hidayat, MSc Ketua
Dr Ir Gede Suastika, MSc Prof Dr Ir Slamet Susanto, MSc Anggota Anggota
Mengetahui
Ketua Program Studi Fitopatologi Dekan Sekolah Pascasarjana
Dr Ir Sri Hendrastuti Hidayat, MSc Dr Ir Dahrul Syah, MScAgr
RIWAYAT HIDUP
Penulis dilahirkan di Bukittinggi pada tanggal 15 November 1966 dari Bapak Basir (Alm) dan Ibu Khasmidar. Tahun 1992, penulis menikah dengan Ir Zainem Efendi dan dikaruniai dua orang putera (Andika Perbawa Wiguna dan Andreas Aulia Rachman), dan satu orang puteri (Andini Vermita Bestari).
Pendidikan Sarjana ditempuh di Jurusan Hama dan Penyakit, Fakultas Pertanian Universitas Andalas lulus pada tahun 1989. Pada tahun 1996, penulis melanjutkan strata 2 di Program Studi Fitopatologi, Sekolah Pascasarjana IPB, lulus pada tahun 1998. Kesempatan melanjutkan ke program doktor pada program studi dan perguruan tinggi yang sama diperoleh tahun 2008. Sejak tahun 1990 sampai sekarang penulis diangkat sebagai staf pengajar tetap pada Fakultas Pertanian, Universitas Jambi.
Syukur Alhamdulillah penulis ucapkan kepada Allah SWT yang telah memberikan segala rahmat dan Karunia-Nya sehingga karya ilmiah ini dapat diselesaikan dengan baik. Tema yang dipilih dalam penelitian ini adalah proteksi
silang, dengan judul Karakterisasi Chili veinal mottle virus Strain Lemah dan
Potensinya sebagai Agens Proteksi Silang.
Pada kesempatan ini, penulis menyampaikan penghargaan dan terima kasih kepada Dr Ir Sri Hendrastuti Hidayat, MSc selaku ketua komisi pembimbing, Dr Ir Gede Suastika, MSc, Prof Dr Ir Slamet Susanto, MSc, dan Prof Dr Ir Sriani Sujiprihati, MS (Alm) selaku anggota komisi pembimbing atas arahan dan bimbingan selama pelaksanaan penelitian dan penyusunan disertasi ini.
Terima kasih disampaikan kepada Rektor Universitas Jambi, Dekan Fakultas Pertanian Universitas Jambi, atas izin yang diberikan kepada penulis untuk mengikuti program doktor (S3) di program studi Fitopatologi, Sekolah Pascasarjana, Institut Pertanian Bogor. Terima kasih disampaikan kepada Rektor Institut Pertanian Bogor, Dekan Sekolah Pascasarjana, seluruh staf pengajar dan administrasi Departemen Proteksi Tanaman, Fakultas Pertanian IPB. Ucapan terima kasih disampaikan kepada tim manajemen Beasiswa Program Pascasarjana (BPPS) Direktorat Jenderal Pendidikan Tinggi Kementerian Pendidikan dan Kebudayaan, Tim Hibah Bersaing Direktorat Penelitian dan Pengabdian Kepada Masyarakat Direktorat Jenderal Pendidikan Tinggi Kementerian Pendidikan dan Kebudayaan, Pemda Provinsi Jambi yang telah memberikan dana dan bantuan dalam mengikuti program doktor.
Terima kasih kepada Mba Tuti Susanti Legiastuti, Pak Edi, seluruh staf rumah kaca, para petani cabai yang bersedia untuk diambil contoh tanaman cabainya untuk keperluan penelitian ini, seluruh teman-teman seperjuangan S3 dan S2, adik-adik mahasiswa S1, dan semua pihak yang telah membantu dalam penelitian dan penyelesaian karya ilmiah ini.
Ucapan terima kasih kepada Ibunda Khasmidar, Ayahanda Basir (Alm), Tante Masdiar, Suami Ir Zainem Efendi dan Anak-anakku tersayang Andika Perbawa Wiguna, Andini Vermita Bestari dan Andreas Aulia Rachman atas dukungan dan kesabaran yang selalu diberikan.
Akhirnya penulis berharap semoga karya ilmiah ini memberikan manfaat bagi kita semua.
Bogor, Januari 2013
DAFTAR ISI DAFTAR TABEL DAFTAR GAMBAR
1 PENDAHULUAN
Latar Belakang Tujuan Penelitian Hipotesis Manfaat Penelitian 1 1 3 4 4
2 TINJAUAN PUSTAKA
Asal Usul dan Arti Penting Tanaman Cabai Virus Penting pada Tanaman Cabai
Kisaran Inang Chili veinal mottle virus
Penularan Chili veinal mottle virus
Karakter Molekuler Chili veinal mottle virus
Interaksi antara Chili veinal mottle virus Isolat Lemah dengan
Isolat Kuat
Pengendalian Chili veinal mottle virus
6 6 7 7 8 9 10 11
3 EKSPLORASI Chili veinal mottle virus ISOLAT LEMAH
DARI PERTANAMAN CABAI Abstrak
Abstract Pendahuluan Bahan dan Metode Hasil dan Pembahasan Kesimpulan 13 13 13 14 15 16 21
4 KISARAN INANG Chili veinal mottle virus ISOLAT LEMAH
Abstrak Abstract Pendahuluan Bahan dan Metode Hasil dan Pembahasan Kesimpulan 22 22 22 23 23 25 30
5 INTERAKSI ANTARA Chili veinal mottle virus ISOLAT
LEMAH DENGAN ISOLAT KUAT Abstrak
7
8
9
Abstrak Abstract Pendahuluan Bahan dan Metode Hasil dan Pembahasan Kesimpulan
KARAKTER MOLEKULER Chili veinal mottle virus
ISOLAT LEMAH Abstrak
Abstract Pendahuluan Bahan dan Metode Hasil dan Pembahasan Kesimpulan
PEMBAHASAN UMUM
KESIMPULAN DAN SARAN Kesimpulan
Saran
DAFTAR PUSTAKA Lampiran
RIWAYAT HIDUP
41 41 42 43 44 54
55 55 55 56 57 59 68
69
72 72 72
73 82
2.1
3.1
3.2
3.3
4.1
4.2
4.3
4.4
5.1
5.2
5.3
5.4
6.1
6.2
Fungsi protein pada organisasi genom Potyvirus
Deteksi Chili veinal mottle virus pada sampel tanaman cabai
tidak bergejala atau bergejala belang ringan yang dikumpulkan dari berbagai daerah pertanaman cabai
Pengelompokan isolat Chili veinal mottle virus berdasarkan
respon tanaman cabai IPB C13
Akumulasi virus direpresentasikan dengan nilai OD 405 nm hasil DAS-ELISA dari daun tanaman cabai IPB C13 pada 28 hari
setelah inokulasi dengan berbagai isolat Chili veinal mottle virus
Karakteristik genotipe cabai yang digunakan dalam pengujian
kisaran inang isolat-isolat Chili veinal mottle virus
Respon genotipe cabai terhadap isolat Chili veinal mottle
virus-CKB (isolat kuat)
Kejadian penyakit pada berbagai genotipe cabai yang terinfeksi
isolat- isolat Chili veinal mottle virus
Kejadian penyakit yang disebabkan oleh isolat-isolat Chili veinal
mottle virus pada tanaman Solanaceae
Skor (indeks) penyakit berdasarkan gejala yang muncul pada
tanaman cabai setelah inokulasi Chili veinal mottle virus
Jenis interaksi Chili veinal mottle virus antara isolat lemah
dengan isolat kuat berdasarkan nilai keparahan penyakit
Respon tanaman cabai IPB C13 terhadap inokulasi Chili veinal
mottle virus isolat lemah dengan isolat kuat
Jenis interaksi antara Chili veinal mottle virus isolat lemah dengan
isolat kuatberdasarkan respon tanaman cabai IPB C13
Masa inkubasi penyakit yang disebabkan Chili veinal mottle
virus pada tanaman cabai IPB C13 pada berbagai interval waktu
inokulasi antara isolat lemah dengan isolat kuat
Tipe gejala penyakit yang disebabkan Chili veinal mottle virus
pada tanaman cabai IPB C13 pada berbagai interval waktu inokulasi antara isolat lemah dengan isolat kuat
9
17
19
20
24
25
27
29
34
35
37
38
45
6.4
6.5
7.1
7.2
7.3
antara isolat lemah dengan isolat kuat
Rata-rata bobot buah pertanaman pada berbagai perlakuan
interval waktu inokulasi antara isolat-isolat lemah Chili veinal
mottle virus dengan isolat kuat ChiVMV-CKB
Rata-rata jumlah buah pertanaman pada berbagai perlakuan
interval waktu inokulasi antara isolat-isolat lemah Chili veinal
mottle virus dengan isolat kuat ChiVMV-CKB
Sikuen nukleotida Chili veinal mottle virus yang berasal dari
GeneBank yang digunakan sebagai pembanding pada analisis keragaman isolat-isolat lemah ChiVMV
Homologi Chili veinal mottle virus isolat lemah dan isolat
ChiVMV dari beberapa negara berdasarkan sikuen nukleotida gen CP
Homologi Chili veinal mottle virus isolat lemah dan isolat
ChiVMV dari beberapa negara berdasarkan sikuen asam amino gen CP
48
52
53
59
63
1.1
2.1
3.1
3.2
4.1
4.2
5.1
6.1
6.2
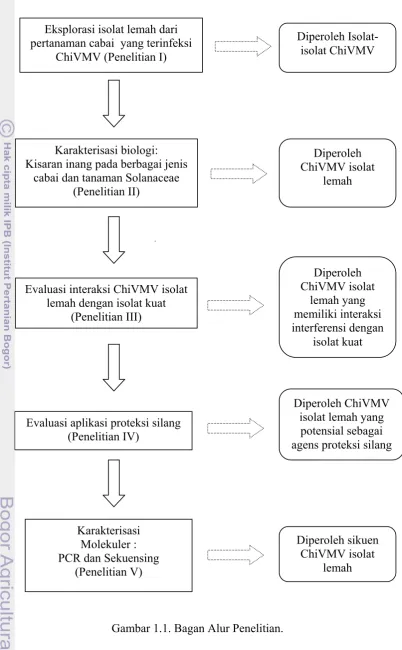
Bagan Alur Penelitian.
Skema organisasi genom Potyvirus (Revers et al. 1999).
Gejala infeksi Chili veinal mottle virus pada tanaman cabai
genotipe IPB C13. A) tidak menunjukkan gejala, B) belang
sedang, malformasi, C) belang berat, malformasi, D) ujung daun
meruncing dan lamina menyempit.
Kejadian penyakit pada tanaman cabai genotipe IPB C13 yang
diinokulasi dengan berbagai isolat Chili veinal mottle virus.
Genotipe cabai yang diinokulasi dengan Chili veinal mottle virus.
A) IPB C13, B) IPB C2, C) IPB C120, D) Laris, E) IPB C8, F) Bara, G) Yolo Wonder, dan H) California Wonder
Tanaman Solanaceae yang diinokulasi dengan isolat Chili veinal
mottle virus: A) Capsicum frutescens, B) Datura metel, C)
Datura stramonium, D) Lycopersicon esculentum, E) Nicotiana benthamiana, F) Nicotiana tabacum cv. Samsun, G) Nicotiana tabacum cv. White Burley, H) Nicotiana tabacum cv. Xanthi, I)
Physalis floridana, J) Solanum melongena, dan K) Solanum nigrum. Semua tanaman yang diinokulasi dengan ChiVMV tidak menunjukkan gejala sampai selesai pengamatan.
Gejala infeksi Chili veinal mottle virus pada enam jenis
inokulasi. A) inokulasi dengan bufer, tidak menunjukkan gejala, B) inokulasi dengan isolat lemah, tidak menunjukkan gejala, C) inokulasi dengan isolat kuat, gejala kerdil (tanaman sebelah kanan), D) inokulasi isolat lemah seminggu kemudian inokulasi isolat kuat, gejala belang ringan, E) inokulasi isolat kuat seminggu kemudian inokulasi isolat lemah, gejala ujung daun meruncing, dan F) inokulasi campuran isolat lemah dengan isolat kuat pada waktu bersamaan, belang berat dan malformasi.
Tanaman cabai yang diinokulasi dengan Chili veinal mottle
virus. A) isolat lemah saja, B) isolat kuat saja, C) isolat kuat setelah isolat lemah.
Perkembangan jumlah tanaman tidak bergejala pada perlakuan berbagai isolat lemah (KAR, SPR, SKT, CSR, dan PGL), isolat kuat (CKB) dan interval waktu inokulasi antara isolat lemah
dengan isolat kuat ChiVMV-CKB. Pada CSR garis dan ,
PGL garis dan , CKB garis dan masing-masing
berada pada posisi yang sama.
5
9
18
21
26
29
36
47
7.1
7.2
7.3
7.4
7.5
28 hari, B) interval 21 hari, C) interval 14 hari, D) interval 7
hari, E) tanpa lemah saja, F) tanaman sehat, dan G) isolat kuat
ChiVMV-CKB saja.
Visualisasi pita DNA hasil amplifikasi dengan primer ChiVMV F Ind dan ChiVMV R Ind pada gel agarosa TBE; M) marker (1 kb ladder), 1) tanaman sehat, 2) KAR, 3) SKT, 4) SPR, 5) CKB, 6) CSR, dan 7) PGL.
Perbandingan hasil sikuen nukleotida 5 isolat lemah ChiVMV
dan isolat kuat CKB-2 (GeneBank aksesi DQ854960). Huruf
dengan latar belakang hitam menunjukkan kesamaan sikuen, sedangkan huruf dengan latar belakang putih menunjukkan
ketidaksamaan sikuen dengan isolat lainnya. Allignment
menggunakan software Bioedit versi 7.0.0 (Isis
Pharmaceuticals, Inc).
Pohon filogenetika berdasarkan sikuen nukleotida Chili veinal
mottle virus, dibuat dengan software Geenbee yang tersedia
pada http://www.genebee.msu.su.
Perbandingan hasil sikuen asam amino 5 isolat lemah ChiVMV
dan isolat kuat CKB-2 (GeneBank aksesi DQ854960). Huruf
dengan latar belakang hitam menunjukkan kesamaan sikuen, sedangkan huruf dengan latar belakang putih menunjukkan
ketidaksamaan sikuen dengan isolat lainnya. Allignment
menggunakan software Bioedit versi 7.0.0 (Isis
Pharmaceuticals, Inc).
Pohon filogenetika berdasarkan sikuen asam amino Chili veinal
mottle virus, dibuat dengan software Geenbee yang tersedia
pada http://www.genebee.msu.su.
52
59
60
63
65
1 PENDAHULUAN
Latar Belakang
Cabai (Capsicum annuum L) merupakan sayuran yang penting. Di Indonesia cabai merah merupakan salah satu komoditas unggulan hortikultura. Tanaman cabai ditanam di seluruh provinsi di Indonesia dan memiliki nilai yang baik sehingga mendapat prioritas untuk dikembangkan (Direktorat Jenderal Hortikultura 2008). Cabai merupakan komponen penting dalam resep masakan karena membentuk rasa dan aroma serta mengandung kalori. Kandungan gizi buah cabai merah (100 g) antara lain protein (12 g), asam lemak (2.78 g), karbohidrat (56.11 g), kalsium (148 mg), fosfor (293 mg), besi (7.78 mg), vitamin A (41610 IU), vitamin B (2.05 mg), dan vitamin C (76.39 mg). Cabai mengandung alkaloid seperti capsaicin (C18H27O3N) yang mencirikan cabai manis, pedas, atau sedang (Indian Institute of Spices Research 2004).
Luas pertanaman cabai di Indonesia pada tahun 2011 mencapai 121 063 ha, dengan produksi sebesar 888 852 ribu ton, dan produktivitas 7.34 ton/ha (Badan Pusat Statistik 2012), sedangkan potensi produksi cabai dapat mencapai sekitar 12 ton/ha (Purwati et al. 2000). Salah satu penyebab rendahnya produktivitas cabai adalah infeksi virus. Virus mengakibatkan kerusakan paling tinggi pada tanaman cabai melalui penurunan kuantitas dan kualitas buah. Sekitar 65 jenis virus dapat menginfeksi cabai di dunia (Asian Vegetable Research and Development Center
(AVRDC) 2001). Virus yang banyak menginfeksi cabai antara lain Chili veinal mottle virus (ChiVMV), Cucumber mosaic virus (CMV), Pepper mild mottle virus
(PMMV), dan Pepper yellow leaf curl virus (PYLCV) (AVRDC 2001; Rashid et al. 2007). Hasil survei South Asian Vegetable Research Network (SAVERNET) diketahui bahwa CMV dan ChiVMV paling banyak terdapat pada pertanaman cabai diikuti oleh PYLCV (AVRDC 1996; Weerraratne dan Yapa 2002).
Chili veinal mottle virus termasuk anggota genus Potyvirus yang merupakan salah satu virus penting yang menginfeksi tanaman cabai di Asia (Lee et al. 2005; Moury et al. 2005; Rashid et al. 2007), di daerah tropik dan subtropik (AVRDC 2000). Survei di 16 negara Asia menunjukkan 30% tanaman cabai terinfeksi oleh ChiVMV, menyebabkan kehilangan hasil sekitar 50% di Pakistan (Shah et al. 2011), dan di Malaysia (Ong et al. 1980). Anggota Potyvirus lain yang menginfeksi tanaman cabai antara lain: Potato virus Y (PVY), Tobacco etch virus (TEV), Pepper mottle virus (PepMoV), dan Pepper veinal mottle virus (PVMV) (Moury et al. 2005).
occidentalis, Solanum nigrum. Datura metel, Physalis floridana (Shah et al. 2008), akan tetapi kisaran inang ChiVMV strain lemah belum pernah dilaporkan.
Penularan ChiVMV di lapangan terjadi melalui serangga vektor yaitu Myzus persicae dan Aphis gossypii (Shah et al. 2008). Serangga vektor yang paling efektif pada pertanaman cabai di dataran tinggi adalah M. persicae, sedangkan di dataran rendah atau di wilayah iklim panas adalah A. gossypii. Pencegahan infeksi virus ini melalui pengendalian serangga vektornya sulit dilakukan karena ChiVMV tergolong non persisten (keberadaan virus hanya terbatas pada stilet) sehingga virus dapat diakuisisi dan ditularkan dalam waktu singkat yaitu hanya beberapa detik (Matthews 2002).
Gejala pada tanaman cabai yang terinfeksi ChiVMV strain kuat berupa belang, perubahan bentuk (malformasi), dan pengurangan ukuran daun (daun mengecil) (Ong 1995; Shah et al. 2009). Berbagai isolat ChiVMV yang diinokulasikan ke C. annuum var. grossum dapat menimbulkan gejala yang berbeda-beda mulai dari gejala ringan (belang ringan, tidak terjadi malformasi daun atau malformasi daun ringan) sampai gejala berat (belang berat, malformasi daun, tanaman kerdil) (Opriana 2009).
Sampai saat ini belum ada strategi pengendalian ChiVMV yang efektif. Usaha yang telah dilakukan adalah mengurangi sumber inokulum dengan mencabut tanaman yang menunjukkan gejala ChiVMV, namun cara ini tentu tidak efektif mengingat proses penularan virus terjadi dengan cepat. Penggunaan kultivar resisten merupakan cara yang paling ekonomis, tetapi pada cabai sedikit sumber ketahanan terhadap ChiVMV (AVRDC 2000). Mengingat belum berhasilnya usaha pengendalian ChiVMV maka perlu dicari alternatif pengendalian yang lain. Salah satu upaya yang mungkin dilakukan adalah melalui proteksi silang dengan menggunakan ChiVMV strain lemah.
Virus strain lemah dapat dimanfaatkan untuk mencegah infeksi virus kedua (strain kuat) pada virus yang sekerabat. Proteksi silang ditemukan oleh McKinney pada tahun 1929 ketika meneliti tanaman tembakau yang terinfeksi oleh “green” strain Tobacco mosaic virus (TMV) yang dapat melindungi dari infeksi oleh strain kuat TMV (Thresh 2006). Proteksi silang merupakan metode biologi (Komar et al. 2008), tidak menyebabkan polusi, tidak beresiko terhadap petani dan konsumen, tidak mengganggu teknik pengendalian yang lain dalam pengelolaan penyakit, aplikasi simpel dan tidak mahal (Rezende dan Pacheco 1998), tidak berbahaya terhadap ekosistem. Proteksi silang dapat digunakan sebagai salah satu komponen pengendalian penyakit terpadu dan dikombinasikan dengan teknik pengendalian yang lain seperti kultur teknis dan sanitasi (Tsuda 2005).
Kemampuan strain lemah untuk melindungi tanaman dari kerusakan yang ditimbulkan oleh strain kuat pada virus yang sekerabat telah diteliti dengan mempelajari hubungan antara virus dan antara strain virus (Thresh 2006). Proteksi silang sama dengan vaksinasi atau immunisasi yang telah digunakan untuk mencegah penyakit pada manusia dan hewan (Tsuda 2005).
interferensi atau kompetisi, virus yang satu akan mendominasi (Roossinck 2005). Menurut Kosaka et al. (2006) interaksi interferensi sesama virus dapat menguntungkan tanaman, karena gejala yang ditimbulkan lebih ringan daripada infeksi tunggal yang disebabkan virus strain kuat, dengan kata lain virus pertama dapat melindungi tanaman dari infeksi kedua (strain kuat) pada virus yang sekerabat. Lecog (1998) menyatakan umumnya virus pertama adalah virus strain lemah yang tidak menimbulkan gejala atau gejala lemah pada tanaman, hal ini mengindikasikan terjadinya proteksi silang pada virus yang menginfeksi tanaman.
Beberapa contoh proteksi silang telah dilaporkan diantaranya Citrus tristeza virus pada jeruk (Lin et al. 2002), TMV pada tomat (Lu et al. 1998), PRSV pada pepaya dan Cucurbitaceae (Rezende dan Pacheco 1998; Wang et al. 1991; You et al. 2005), Cacao swollen shoot virus di Ghana (Ollennu dan Owusu 2003), Pepino mosaic virus (PepMV) pada tomat (Hanssen et al. 2010), dan Grapevine fanleaf virus (GFLV) pada anggur (Komar et al. 2008), ZYMV pada Cucurbitaceae (Kosaka et al. 2006; Lecog et al. 1991; Rezende dan Pacheco, 1998; Walkey et al. 1992). ZYMV strain lemah efektif untuk mengendalikan penyakit yang disebabkan oleh ZYMV pada Cucurbitaceae dan telah dilakukan lebih dari 10 tahun di Brazil (Bonilha et al. 2009) dan menjadi salah satu alternatif pengendalian ZYMV (Gal-on 2007; Wang et al. 1991). Akan tetapi proteksi silang belum pernah dilaporkan pada ChiVMV.
Sebelum menerapkan proteksi silang untuk mengendalikan ChiVMV perlu dilakukan penelitian dengan tahap-tahap sebagai berikut: eksplorasi ChiVMV isolat lemah dari pertanaman cabai, karakterisasi ChiVMV baik karakterisasi biologi maupun molekuler, mempelajari interaksi ChiVMV strain lemah dengan strain kuat, dan evaluasi efisiensi proteksi silang untuk menekan kehilangan hasil akibat infeksi strain kuat (Gambar 1).
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Mendapatkan isolat- isolat lemah ChiVMV dari pertanaman cabai.
2. Mempelajari kisaran inang isolat-isolat ChiVMV pada berbagai genotipe cabai dan tanaman Solanaceae.
3. Mempelajari interaksi antara ChiVMV isolat lemah dengan isolat kuat. 4. Mengevaluasi proteksi silang menggunakan ChiVMV isolat lemah yang
terpilih.
Hipotesis
1. ChiVMV isolat lemah telah ada dan menyebar di beberapa pertanaman cabai dan tidak menimbulkan gejala atau menimbulkan gejala lemah pada tanaman cabai.
2. ChiVMV mempunyai kisaran inang pada beberapa tanaman Solanaceae. 3. ChiVMV isolat lemah memiliki interaksi interferensi dengan ChiVMV isolat
kuat.
4. ChiVMV isolat lemah dapat melindungi tanaman cabai dari infeksi ChiVMV isolat kuat.
5. Terdapat perbedaan sikuen nukleotida dan asam amino antara sesama isolat ChiVMV.
Manfaat Penelitian
Gambar 1.1. Bagan Alur Penelitian. Evaluasi aplikasi proteksi silang
(Penelitian IV)
Evaluasi interaksi ChiVMV isolat lemah dengan isolat kuat
(Penelitian III) Karakterisasi biologi: Kisaran inang pada berbagai jenis
cabai dan tanaman Solanaceae (Penelitian II)
Eksplorasi isolat lemah dari pertanaman cabai yang terinfeksi
ChiVMV (Penelitian I)
Karakterisasi Molekuler : PCR dan Sekuensing
(Penelitian V)
Diperoleh Isolat-isolat ChiVMV
Diperoleh ChiVMV isolat lemah yang potensial sebagai agens proteksi silang
Diperoleh ChiVMV isolat
lemah
Diperoleh ChiVMV isolat
lemah yang memiliki interaksi interferensi dengan
isolat kuat
Diperoleh sikuen ChiVMV isolat
2 TINJAUAN PUSTAKA
Asal Usul dan Arti Penting Tanaman Cabai
Cabai termasuk dalam genus Capsicum, famili Solanaceae, ordo Solanales, klas Magnolipsida dan divisi Magnoliophyta (Bosland dan Votaya 2000). Tanaman cabai berasal dari Meksiko Amerika Tengah dan Pegunungan Andes di Amerika Selatan. Pada tahun 1493 Colombus membawa dan memperkenalkan benih cabai ke Spanyol dan menyebar dengan cepat di Eropa. Pada abad ke 16 tanaman cabai dibawa ke Asia termasuk Indonesia oleh bangsa Portugis dan Spanyol dari Amerika Selatan, kemudian tersebar luas di daerah tropik dan sub tropik (Rubatsky dan Yamaguchi 1997).
Beberapa spesies cabai yang penting adalah C. annuum, C. frutescens, C. pubescens, C. baccatum, C. chinensis. Spesies cabai liar antara lain C. praetermisum, C. chacoense, C. galapogoense, C. cardenassi, C. eximium, C. tovarii (Bosland dan Votaya 2000). Di Indonesia terdapat dua jenis cabai yang banyak ditanam yaitu cabai besar (C. annuum) dan cabai rawit (C. frutescens). Cabai besar yang sering ditanam adalah cabai merah besar dan cabai merah keriting.
Cabai merupakan salah satu komoditas hortikultura yang penting. Cabai sudah menjadi kebutuhan pokok masyarakat sehari-hari, disamping itu cabai memiliki nilai ekonomis yang cukup tinggi. Cabai mengandung vitamin A dan C, serta antioksidan yang berfungsi untuk menjaga tubuh dari serangan radikal bebas, lasparaginase dan capsaicin yang berperan sebagai zat anti kanker, mencegah flu dan demam, meredakan batuk, meringankan penyakit asma dan bronkitis. Cabai berperan penting dalam masakan dan tidak bisa disubstitusi dengan komoditas lain, serta dimanfaatkan sebagai bahan campuran berbagai industri pengolahan makanan dan pembuatan obat-obatan (Macrae et al. 1993).
Virus Penting pada Tanaman Cabai
Virus yang menginfeksi tanaman cabai adalah ChiVMV, CMV, PVMV, TEV, TSWV, TLCV, TMV, ToMV (Shah et al. 2008). ChiVMV pertama kali dilaporkan di Malaysia oleh Barneet 1947. ChiVMV tersebar luas dan mengurangi hasil pada pertanaman cabai di Peninsula Malaysia (Ong et al. 1979). ChiVMV terdapat di Indonesia (Taufik et al 2005), Papua New Guinea, Australia, Korea, Filipina, Taiwan, Thailand. Distribusi geografi ChiVMV tidak absolut seperti terdapat secara sporadik di negara lain di dunia seperti Afrika Timur (Nono-Wondin et al. 2001).
Sama halnya seperti kelompok Potyvirus lain, ChiVMV memiliki struktur khusus (badan inklusi) berbentuk cakra yang banyak ditemukan dalam sitoplasma dan inti. Lipid tidak dilaporkan pada ChiVMV. Titik pengenceran (dilution end point DEP) adalah 10 -5, thermal inactivation point (TIP) adalah 60 oC, longevity in vitro (LIV) adalah 7 hari pada suhu kamar. Pengamatan dengan mikroskop elektron menunjukkan ChiVMV mempunyai panjang 750 nm, lebar 12 nm (Ong et al. 1979). Berat molekul coat protein (CP) ChiVMV adalah 28 kDa berdasarkan analisis polyacrylamide gel electrophoresis (PAGE) (Shah 2006).
Tanaman cabai yang terinfeksi ChiVMV menunjukkan gejala belang hijau tua, pada gejala lanjut menyebabkan berkurangnya ukuran daun, distorsi daun dan buah kecil (Weerraratne dan Yapa 2002), belang, distorsi, dan mosaik pada buah, bercak hijau tua tidak beraturan, penebalan tulang daun, malformasi daun, daun menjadi kecil (Siriwong et al. 1995), hijau tua, kemudian distorsi dengan mosaik, tanaman terinfeksi menyebabkan berkurang jumlah bunga dan buah, sehingga menyebabkan kehilangan hasil (Wang et al. 2006).
Satu tanaman cabai dapat terinfeksi oleh beberapa jenis virus. Tanaman cabai sering ditemui terinfeksi ChiVMV bersama-sama dengan CMV (Taufik et al. 2005). Sampel tanaman cabai yang dikumpulkan dari lapangan menunjukkan terinfeksi ganda ChiVMV dan CMV sekitar 10% (Shah et al 2001), 37% (Weeraratne dan Yapa 2002). Infeksi ganda ini menyebabkan penghambatan pertumbuhan tanaman dan penurunan produksi lebih berat dari pada infeksi tunggal ChiVMV atau CMV (Subekti et al 2006).
Pada beberapa kombinasi inang-virus, penyakit yang disebabkan oleh virus strain kuat dapat dihindari jika tumbuhan tersebut pertama di inokulasi dengan strain lemah dari virus yang sama (sekerabat) melalui teknik proteksi silang yang dapat melindungi tumbuhan terhadap infeksi virus strain kuat (Agrios 2005), namun proteksi silang pada ChiVMV belum pernah dilaporkan.
Kisaran Inang Chili veinal mottle virus
glutinosa, N. occidentalis, C. quinosa, S. nigrum, D. metel, P. floridana pada 7 sampai 14 hari setelah inokulasi (Shah et al. 2008).
Sejumlah gulma banyak terdapat di sekitar pertanaman cabai, kemungkinan gulma ini dapat menjadi sumber infeksi ChiVMV (Shah et al. 2009). D. metel dilaporkan dapat terinfeksi ChiVMV baik di lapangan dan di rumah kaca. Gulma ini merupakan inang alternatif atau sumber inokulum untuk infeksi sekunder pada musim tanam berikutnya (Shah et al. 2008).
Penularan Chili veinal mottle virus
Penularan virus yang paling umum dan sangat penting di lapangan adalah melalui serangga vektor. Kutudaun merupakan golongan serangga yang paling banyak berperan sebagai vektor virus tumbuhan. Kutudaun umumnya mendapatkan virus setelah makan pada tumbuhan sakit hanya selama beberapa detik (30 detik atau kurang) dan dapat menularkan virus tersebut setelah pindah dan makan pada tumbuhan sehat dalam waktu yang singkat. Panjang waktu kutudaun tetap bersifat viruliferous setelah mendapat virus bervariasi dari beberapa menit sampai beberapa jam (Agrios 2005; Matthews 2002).
Hasil survei hama dan penyakit oleh AVRDC pada pertanaman cabai di beberapa negara di Asia menyimpulkan bahwa hama yang banyak pada pertanaman cabai adalah kutudaun (A. gossypii, M. persicae), dan penyakit yang sering ditemui adalah penyakit yang disebabkan oleh ChiVMV dan CMV (Berke 2002). Oleh karena itu peran kutudaun sangatlah penting yaitu sebagai hama dan sebagai vektor virus.
Chili veinal mottle virus ditularkan oleh kutudaun (Wang et al. 2006), dan secara mekanis (Nono-Womdim et al. 2001). ChiVMV ditularkan secara non persisten oleh A. gossipii, A. craccivora, A. spiraecola, M. persicae, Toxoptera citricidus, Hysteroneura setarieae, Rhopalosiphum maidis, secara non persisten (Ong 1979; Shah et al. 2009). Virus dapat diperoleh dan kemudian ditularkan oleh kutudaun dalam waktu 2 detik sampai beberapa menit melalui makan pada tanaman sehat (Matthews 2002).
Chili veinal mottle virus dapat ditularkan secara mekanis ke tanaman C. annuum, C. frutescens, D. metel, D. stramonium, L. esculentum, N. glutinosa, N. tabacum, P. floridana, dan P. hybrida, (Siriwong et al. 1995). ChiVMV dapat ditularkan secara mekanis ke N. tabacum, N. glutinosa, N. occidentalis, D. metel, P. floridana, S. nigrum (Shah et al. 2008). Di Afrika, ChiVMV dapat ditularkan secara mekanis dari terung ke tembakau (N. tabacco cv. Xanthi), cabai (C. annuum cv Yolo Wonder), tomat (L. esculentum cv Tengeru 97), dan S. aethiopicum cv. Tengeru White, penularan secara mekanis ini tidak terjadi pada melon (Cucumis melo), ketimun (C. sativus) dan Vigna unguiculata yang bukan inang ChiVMV (Nono-Wondim et al. 2001).
tersebut (Agrios 2005). Penularan secara mekanis melalui inokulasi cairan tanaman sakit biasanya dilakukan untuk menguji kisaran inang dan sifat ketahanan tanaman terhadap virus. Dari hasil penelitian Wang et al. (1991) terhadap infeksi ZYMV strain kuat pada Cucurbitaceae, inokulasi pada fase bibit memberikan hasil yang lebih baik dibanding fase pertumbuhan lanjut. Pada tanaman yang diperbanyak dengan biji, metode yang efisien untuk inokulasi adalah pada fase bibit (Rezende dan Pacheco 1998). ChiVMV tidak terbawa benih, tidak terdeteksi pada organ reproduksi tanaman (androecium dan gynaecium). ChiVMV terdeteksi hanya pada sepal. Jadi ChiVMV tidak ditularkan melalui benih dan tidak berada pada bagian reproduksi (Shah et al 2008).
Karakter Molekuler Chili veinal mottle virus
Chili veinal mottle virus termasuk genus Potyvirus, berbentuk benang dengan panjang 680 sampai 900 nm dan lebar 11 sampai 13 nm, memiliki genom berupa untai tunggal RNA, positive sense (Nono-Wondim et al. 2001), dengan ukuran 9.7 kb (Shukla et al 1994). Viral protein genome-linked (VPg) terdapat pada ujung 5’ dan poly (A) pada ujung 3’ (Gambar 2.1). Protein di kode oleh 10 genom yang memiliki berbagai fungsi (Tabel 2.1). Coat protein (CP) mengkode 858 sampai 864 nukleotida dan 3” untranlated regions (3’ UTR) mengkode 275-289 nukleotida. (Tsai et al. 2008).
5´NTR 3´NTR
Gambar 2.1 Skema organisasi genom Potyvirus (Revers et al. 1999)
Tabel 2.1 Fungsi protein pada organisasi genom Potyvirus
Protein Fungsi P1
HC-Pro
P3 6 K1 CI
6 K2
NIaVPg NIaPro NIb CP
Proteinase, pergerakan dari sel ke sel
Penularan oleh vektor, proteinase, patogenisitas, penekan RNA silencing, pergerakan dari sel ke sel
Belum diketahui, kemungkinan replikasi Belum diketahui, kemungkinan replikasi
Replikasi genom (RNA helicase), stimulasi asam nukleat aktivitas ATP ase, pergerakan dari sel ke sel
Belum diketahui, kemungkinan replikasi, mengatur penghambatan translokasi NIa nuclear
Replikasi genom (primer untuk memulai sintesa RNA) Proteinase
Replikasi genom (RNA-dependent RNA-polimerase [RdRp]) Penularan oleh vektor, patogenisitas, encapsidase RNA, pergerakan dari sel ke sel
Genom Potyvirus memiliki satu open reading frame (ORF) yang mengkode polyprotein 340 kDa. Translasi RNA dimulai pada start codon AUG pada 145 sampai 205 dari ujung 5’ dan stop codon terdapat pada 186 sampai 250 dari ujung 3’ yang ditandai dengan poly (A) seperti AAUAAA. Protein virus dihasilkan dengan mengkode poliprotein yang diproses oleh 3 proteinase virus yaitu nuclear inclusion protein a (NIa), helper component-Proteinase (HC-Pro), dan P1 proteinase (P1) (Verchot et al. 1991).
Sikuen nukleotida dan asam amino ChiVMV gen CP sesama 24 isolat dari China, India, Indonesia, Taiwan, dan Thailand menunjukkan kesamaan masing-masing lebih dari 89.5% dan 94.4%, sedangkan kesamaan Potyvirus lain adalah kurang dari 83.6% untuk sikuen nukleotida dan kurang dari 79.4% untuk sikuen asam amino (Tsai et al. 2008). Kesamaan sikuen nukleotida sesama 3 isolat ChiVMV (Beijing, Taiwan, dan Thailand) sekitar 90.3 sampai 92.8%, dengan isolat PVMV dari 77.7 sampai 82.2%, sedangkan kesamaan sikuen nukleotida dengan Potyvirus lain adalah kurang dari 70%. Group yang sama dideterminasi berdasarkan gen CP dan 51 nukleotida pada 5’-proximal region. Berdasarkan kesamaan sikuen nukleotida paling rendah 85% dalam genus Potyvirus berarti dengan kesamaan sikuen nukleotida kurang dari 85% maka dikelompokkan pada spesies yang berbeda. Hal ini valid menempatkan PVMV dan ChiVMV sebagai 2 spesies yang berbeda. Tingginya kesamaan asam amino antara PVMV dengan ChiVMV dapat dijelaskan melalui uji serologi (Moury et al. 2005). Kesamaan sikuen asam amino gen CP CVbMV-CM1 (ChiVMV-CM1) dengan 18 Potyvirus lain berkisar antara 54% sampai 65% (Ikegami et al. 1999).
Pemilihan primer untuk ChiVMV dapat dilakukan pada daerah coat protein (CP), 3 untranslated region (3 UTR), nuclear inclusion protein (NIb). Primer D [GGIAA(A/G)GC(G/A/T/C)CC(G/A/T/C)TA(C/T)AT] merupakan primer yang berhubungan dengan ‘GKAPYL’ motif asam amino pada protein NIb dari 8421 sampai 8438 nukleotida ChiVMV. Primer E (CGCGCTAATGACATATCGGT) berhubungan dengan ‘TDMSLAR’ asam amino motif konversi dalam CP dari 9138 sampai 9157 nukleotida ChiVMV. RT-PCR dengan primer D dan E dapat mendeteksi ChiVMV pada fragmen DNA berukuran 788 bp, PVMV pada 737 bp. Penambahan 51 nukleotida yang berhubungan dengan 17 kodon terdapat pada region ini dari 3 macam isolat ChiVMV bila dibandingkan dengan sikuen PVMV. 708 nukleotida pada 3’ dari gen CP digunakan untuk mengkalkulasikan adanya kesamaan ChiVMV, PVMV dan Potyvirus lain. Untuk mengetahui keragaman suatu virus atau untuk mengetahui adanya infeksi campuran pada inang yang sama dilakukan deteksi secara RT-PCR (Moury et al. 2005).
Interaksi Chili veinal mottle virus Isolat Lemah dengan Isolat Kuat
Dua atau beberapa virus dapat menginfeksi satu tanaman secara bersama-sama. Virus-virus tersebut dapat menginfeksi pada sel atau jaringan yang sama atau berbeda dalam satu tanaman. Infeksi dua atau lebih jenis virus dalam tanaman berpotensi untuk berinteraksi. Virus yang menginfeksi bersama-sama bisa saja tidak sekerabat atau sekerabat (Roossinck 2005).
replikasi atau distribusi virus lain (Atabekov dan Dorokhov 1984), sebaliknya interaksi interferensi adalah keberadaan satu virus dapat mencegah infeksi virus yang lain (virus kedua). Fenomena virus pertama dapat melindungi dari infeksi kedua yang menyebabkan gejala yang parah pada inang yang sama dikenal dengan proteksi silang. Virus pertama ini digunakan untuk mengendalikan virus kedua dari virus yang sekerabat (Aguilar et al. 2000), kompetisi biasanya terjadi pada virus yang sekerabat (Roossinck 2005).
Interaksi dapat menguntungkan pada satu atau kedua virus dengan menyebabkan gejala lebih parah, biasanya pada interaksi sinergis antara PVY dengan PVX. Sinergis ini dicirikan oleh peningkatan 3 sampai 10 kali titer PVX dibanding infeksi tunggal. PVY dan PVX jelas sinergis dalam infeksi campuran, akan tetapi hubungan yang saling menguntungkan antara kedua virus ini berlangsung dalam waktu yang pendek, karena pada infeksi campuran ini penyakit lebih parah dan sering mengakibatkan kematian tanaman (Roossinck 2005).
Dua genotipe Tobacco mild green mosaic virus (TMGMV) yaitu large (TMGMV-L) dan small (TMGMV-S) merupakan virus sekerabat dapat berada bersama-sama dalam tanaman N. glauca dan terjadi interaksi interferensi atau virus berkompetisi satu sama lain secara alami di lapangan (Bodaghi et al. 2004). Kemudian Lin et al. (2002) menyatakan bahwa isolat lemah dan isolat kuat CTV dapat berada bersama-sama dalam tanaman jeruk besar secara alami, adanya kedua isolat lemah dan kuat CTV pada jeruk menyebabkan tanaman tidak bergejala yang mengindikasikan terjadinya proteksi silang. Dilaporkan isolat lemah CTV dapat melindungi tanaman jeruk besar dalam waktu yang lama terhadap infeksi isolat kuat CTV. Proteksi silang dengan isolat lemah kemungkinan terjadi interferensi terhadap kemampuan strain kuat untuk mencapai konsentrasi yang tinggi. Interaksi dua virus yang saling berkompetisi dapat menguntungkan terhadap tanaman.
Pengendalian Chili veinal mottle virus
Pengendalian penyakit yang disebabkan oleh virus pada umumnya sulit dilakukan (Hull 2002). Upaya pengendalian virus pada tanaman cabai yang dilakukan oleh petani adalah dengan mengurangi sumber inokulum dengan cara mencabut tanaman yang menunjukkan gejala terinfeksi virus, tetapi cara ini tidak efektif karena penularan virus dapat terjadi dengan cepat. Pengendalian ChiVMV cukup sulit mengingat (1) tanaman yang terinfeksi ChiVMV tidak dapat disembuhkan, (2) ChiVMV mempunyai kisaran inang yang luas, dan (3) ChiVMV ditularkan secara non persisten oleh kutu daun.
Usaha pengendalian ChiVMV yang baik adalah dengan penanaman varietas tahan (resisten) (Agrios 2005), akan tetapi hingga saat ini belum diperoleh kultivar cabai yang tahan terhadap ChiVMV (Rashid et al. 2007). Semua genotipe cabai yang ditanam di Pakistan rentan terhadap infeksi ChiVMV (Shah et al.
tahan terhadap ChiVMV (Subekti et al. 2006). Untuk itu perlu dicari alternatif pengendalian yang lain, salah satunya adalah melalui teknik proteksi silang dengan cara menginfeksi tanaman dengan virus strain lemah yang menimbulkan gejala lemah untuk melindungi tanaman dari infeksi strain kuat. Proteksi silang merupakan teknik untuk melindungi tanaman dengan memanfaatkan virus strain lemah yang dapat mencegah exspresi virus kedua pada virus yang berhubungan (virus dari spesies yang sama) (Bonilha et al. 2009).
Pada konsep proteksi silang, digunakan strain virus yang menginduksi proteksi silang sebagai “protecting strain”, dan strain yang digunakan untuk mengevaluasi efisiensi proteksi silang sebagai “challenging strain”. Protecting strain umumnya adalah strain lemah yang tidak menimbulkan gejala atau gejala lemah dan tidak mempengaruhi produktivitas tanaman. Challenging strain (virus kedua) dapat diinokulasikan secara mekanis dengan waktu yang berbeda setelah inokulasi dengan protecting strain (Lecog 1998; Matthews 2002). Proteksi silang tidak terjadi jika kedua virus diinokulasikan secara bersama-sama, jadi diperlukan beberapa hari setelah virus pertama menginfeksi tanaman untuk keseimbangan proteksi silang terhadap virus kedua (Roossinck 2005). Replikasi mungkin dapat terjadi pada virus kedua, tetapi mungkin gagal menimbulkan gejala (Lecoq 1991).
Mekanisme proteksi silang adalah kompetisi dalam hal tempat replikasi, pada pengamatan dengan mikroskop elektron, pada proteksi silang menyebabkan hampir semua tempat (site) replikasi virus pada tanaman ditempati oleh strain lemah, konsekuensinya bila di challenge dengan strain kuat menyebabkan sedikit atau tidak ada tempat yang tersedia pada tanaman yang dilindungi untuk replikasi virus strain kuat (Matthews 2002; Walkey et al. 1992).
Proteksi silang pada ZYMV berdasarkan pada mekanisme RNA silencing, yang dapat mencegah virus yang sekerabat dari multiplikasi dalam sel yang sama. Pada tanaman yang diinokulasi dengan ZYMV strain lemah maka akan terjadi penumpukan viral small interfering RNA (vsi RNA) dalam sel terinfeksi dan sel didekatnya, dan kemudian apabila ZYMV strain challenge masuk ke dalam sel, maka ZYMV strain lemah akan mengaktifkan RNA-induced silencing complex (RISC) (mengandung vsi RNA) yang akan mendegradasi strain challenge sebelum strain challenge menjadi seimbang, ZYMV strain lemah tetap ada dengan aktifnya RISC. Interval antara inokulasi ZYMV strain lemah dengan ZYMV strain challenge diperlukan untuk perlindungan karena diperlukan konsentrasi tertentu dari vsiRNA dan atau aktivitas RISC, perlu sampai terakumulasi dalam sel terinfeksi dan atau sel didekatnya (Gal-on 2007).
Aplikasi yang tepat dalam hal interval, yaitu selisih waktu yang diperlukan antara inokulasi strain proteksi dengan strain challenge untuk mendapatkan perlindungan yang efektif. Interval 14 hari antara inokulasi strain proteksi dengan strain challenge efektif untuk melindungi tanaman. Interval 7 atau 8 hari menghasilkan perlindungan sedang, dan tidak ada perlindungan yang diperoleh pada interval 2 hari (Walkey et al. 1992).
3 EKSPLORASI
Chili veinal mottle virus
ISOLAT LEMAH
DARI PERTANAMAN CABAI
(Exploration of Weak Isolates of
Chili veinal mottle virus
from Chili Pepper)
Abstrak
Salah satu virus utama yang menginfeksi tanaman cabai adalah Chili veinal mottle virus (ChiVMV) yang potensial menyebabkan kehilangan hasil. Strategi pengendalian virus melalui proteksi silang mengandalkan kemampuan virus strain lemah untuk melindungi tanaman dari infeksi virus strain kuat. Penelitian dilakukan untuk mengeksplorasi ChiVMV isolat lemah dari pertanaman cabai di provinsi Jambi, Sumatera Barat, dan Jawa Barat. Isolat-isolat ChiVMV yang diperoleh berhasil ditularkan dan diperbanyak pada tanaman cabai (Capsicum annuum L.) di rumah kaca. Berdasarkan gejala penyakit isolat ChiVMV dapat dibedakan menjadi tiga kelompok yaitu isolat lemah (KAR, SPR, EAK, SKT, LGM, SKR, CSR, PGL, KRS, KMT, KRD, STG, KLT, SLO, BTL, dan WTE), isolat sedang (CGN dan NSI) , dan isolat kuat (CKW, CKB, dan BLG). Isolat-isolat lemah ChiVMV selanjutnya dapat digunakan sebagai kandidat agens proteksi silang dalam pengendalian penyakit belang yang disebabkan oleh ChiVMV.
Kata kunci : Chili veinal mottle virus, isolat lemah, tanaman cabai
Abstract
Chili veinal mottle virus (ChiVMV) is known as an important virus infecting chili pepper and may cause significant yield loss. Management strategy of virus diseases using cross protection relies on the ability of mild strain of virus to protect plant from infection by severe strain of the same virus. Initial exploration was conducted in several chili pepper growing areas in West Sumatera, Jambi, and West Java to obtain mild isolates of ChiVMV. Field isolates of ChiVMV were propagated in chili pepper (Capsicum annuum L.) in the screenhouse. Based on symptom development ChiVMV isolates can be differentiated into 3 groups, i.e. weak isolates (KAR, SPR, EAK, SKT, LGM, SKR, CSR, PGL, KRS, KMT, KRD, STG, KLT, SLO, BTL, and WTE), mild isolates (CGN and NSI), severe isolates (CKW, CKB, and BLG). Further characterization of promising ChiVMV isolates is required to confirm the potency of virus isolates to be used in cross protection strategy to suppress disease incidence.
Pendahuluan
Penyakit yang disebabkan oleh Chili veinal mottle virus (ChiVMV) diketahui hanya menyebar di wilayah tertentu saja. Distribusi geografi ChiVMV mencakup Indonesia, Malaysia, Papua New Guinea, Korea, Filipina, Taiwan, Thailand, Australia (Nono-Wondim et al. 2001), Afrika Timur (Moury et al.
2005), China (Wang et al. 2006), Pakistan (Shah et al. 2009). Ledakan penyakit yang disebabkan oleh ChiVMV pada pertanaman cabai terjadi di China selama tahun 2003 sampai 2004 (Wang et al. 2006). Infeksi ChiVMV dapat menurunkan kualitas buah, dan menyebabkan kehilangan hasil mencapai 50% di Pakistan (Shah et al. 2011), dan Malaysia (Ong et al. 1980).
Tanaman cabai yang terinfeksi ChiVMV menunjukkan gejala tunggal atau kombinasi (Ong et al. 1979) yaitu belang hijau tua, penebalan tulang daun, malformasi daun, dan daun menjadi kecil (Ong et al. 1979; Siriwong et al. 1995; Weeraratne dan Yapa 2002). Berbagai isolat ChiVMV yang diinokulasikan ke
C.annuum dapat menimbulkan gejala yang berbeda-beda, mulai dari gejala ringan (belang ringan, tidak terjadi malformasi daun atau malformasi daun ringan) sampai gejala berat (belang berat, malformasi daun) (Opriana 2009).
Pengendalian ChiVMV melalui pengendalian serangga vektor dinilai tidak efektif. Hal tersebut berkaitan dengan sifat retensi virus (lamanya vektor dapat menularkan virus) dan waktu untuk akuisisi dan inokulasi virus. ChiVMV tergolong bersifat non persisten (keberadaan virus hanya terbatas pada stilet serangga) sehingga virus dapat diakuisisi dan ditularkan dalam waktu singkat yaitu beberapa detik, dan kemampuan vektor menularkan virus berlangsung selama beberapa menit (Shah et al. 2008). Pengendalian melalui perakitan varietas tahan mengalami kendala karena terbatasnya sumber ketahanan terhadap ChiVMV (Rashid et al. 2007). Salah satu strategi pengendalian yang belum banyak dievaluasi adalah melalui pendekatan proteksi silang (cross protection).
Proteksi silang adalah strategi untuk melindungi tanaman dengan memanfaatkan virus strain lemah untuk mencegah infeksi virus strain kuat pada virus yang sekerabat (Bonilha et al. 2009). Virus strain lemah tidak menimbulkan gejala atau menimbulkan gejala sangat lemah pada tanaman sepanjang musim tanam, virus strain lemah tidak berbahaya terhadap ekosistem seperti tanaman, serangga, dan mamalia, serta tidak menyebabkan polusi terhadap lingkungan dan efektif untuk lingkungan pertanian berkelanjutan. Jika virus strain lemah aman pada segala kondisi maka virus strain lemah dapat digunakan untuk skala luas (Tsuda 2005). Proteksi silang merupakan salah satu komponen pengendalian penyakit terpadu dan dapat dikombinasikan dengan teknik pengendalian yang lain.
Upaya menerapkan proteksi silang untuk mengendalikan ChiVMV diawali dengan kegiatan eksplorasi ChiVMV isolat lemah dari pertanaman cabai. Adanya informasi tentang ChiVMV isolat lemah dapat bermanfaat dalam pemilihan ChiVMV isolat lemah yang akan digunakan untuk proteksi silang. Oleh karena itu tujuan penelitian ini adalah mendapatkan ChiVMV isolat lemah melalui eksplorasi pada pertanaman cabai.
Bahan dan Metode
Penelitian dilakukan di beberapa daerah pertanaman cabai, Laboratorium Virologi Tumbuhan dan Rumah Kaca, Departemen Proteksi Tanaman, Institut Pertanian Bogor, Bogor dari bulan Februari sampai Juli 2011.
Pengumpulan Sampel dari Pertanaman Cabai
Daun sampel tanaman cabai diambil dari beberapa pertanaman di daerah Jambi, Sumatera Barat, dan Jawa Barat selama musim tanam. Daun cabai yang diambil ialah daun yang menunjukkan gejala belang ringan atau daun yang tidak bergejala dari tanaman yang berada di sekitar tanaman bergejala belang berat. Pada penelitian ini juga digunakan isolat koleksi dari Laboratorium Virologi Departemen Proteksi Tanaman Fakultas Pertanian IPB untuk optimalisasi seleksi isolat virus.
Deteksi dengan Double Antibody Sandwich Enzyme-linked Immunosorbent Assay (DAS-ELISA)
Daun sampel yang diperoleh dari pertanaman cabai diberi kapas yang dibasahi akuades pada bagian tangkai daun, dimasukkan kedalam plastik polythene lalu disimpan dalam kotak pendingin sebelum dibawa ke laboratorium. Sampel tersebut selanjutnya diuji dengan metode DAS-ELISA menggunakan tiga jenis antiserum secara terpisah, yaitu antiserum ChiVMV, CMV, dan TMV.
pada suhu ruang, sambil diamati terjadinya perubahan warna menjadi kuning pada sumuran platmikrotiter. Nilai absorbansi secara kuantitatif dibaca dengan spektrofotometer Microplate reader BIO-RAD Model 550 pada panjang gelombang 405 nm. Reaksi dihentikan dengan penambahan 3 M NaOH sebanyak 50 μl tiap sumuran.
Penularan ChiVMV ke Tanaman Cabai
Berdasarkan hasil DAS-ELISA dilakukan seleksi untuk tahapan penelitian selanjutnya. Sampel yang dipilih adalah sampel yang positif terinfeksi hanya oleh ChiVMV dengan titer DAS-ELISA tertinggi atau terendah dari setiap daerah pengambilan sampel. Sampel daun tersebut digunakan untuk inokulasi ke tanaman cabai (C. annuum) genotipe IPB C13 dengan metode inokulasi mekanis menggunakan cairan perasan tanaman sakit (sap). Berdasarkan hasil penelitian Opriana (2009) genotipe IPB C13 sangat rentan terhadap ChiVMV.
Sap dibuat dengan cara daun tanaman cabai digerus yang sebelumnya ditambah larutan penyangga fosfat (0.01 M, pH 7) dengan perbandingan 1:5 (b/v). Tanaman cabai yang akan diinokulasi ditaburi carborundum 600 mesh pada permukaan atas daun yang berguna untuk membuat pelukaan kecil tanpa mematikan sel daun yang akan diinokulasi ChiVMV. Sap dioleskan pada permukaan daun tanaman cabai menggunakan kapas steril. Inokulasi dilakukan pada dua helai daun bagian atas (daun termuda yang telah membuka penuh), kemudian daun tanaman disiram dengan air mengalir untuk membersihkan sisa carborundum yang masih melekat (Green 1991). Tanaman dipelihara dan ditempatkan dalam rumah kaca kedap serangga.
Seleksi Isolat ChiVMV
Pengamatan terhadap tanaman cabai yang sudah diinokulasi ChiVMV dilakukan setiap hari selama empat minggu setelah inokulasi mencakup pengamatan gejala, masa inkubasi, dan kejadian penyakit. Infeksi ChiVMV dideteksi dengan metode DAS-ELISA (seperti diuraikan sebelumnya).
Kejadian penyakit dihitung dengan rumus :
Jumlah tanaman terinfeksi
Kejadian penyakit = --- x 100 % Jumlah tanaman di inokulasi
Pengelompokan isolat ChiVMV didasarkan pada gejala yaitu isolat lemah (tidak ada gejala, belang ringan), isolat sedang (gejala belang sedang, malformasi), dan isolat kuat (gejala belang berat, malformasi, ujung daun meruncing, tanaman kerdil).
Hasil dan Pembahasan
Eksplorasi dan Deteksi Virus dengan DAS-ELISA
dan Yapa (2002) di Sri Lanka dan Shah et al.(2009) di Pakistan yaitu belang, malformasi, daun menjadi kecil. Hasil pengamatan di lapangan di sekitar tanaman yang menunjukkan gejala belang berat ditemukan juga tanaman yang tidak bergejala dan kelihatan sehat serta tidak mengalami penghambatan pertumbuhan. Pada beberapa areal pertanaman cabai dapat dijumpai tanaman yang menunjukkan gejala belang ringan tetapi tidak mengalami penghambatan pertumbuhan.
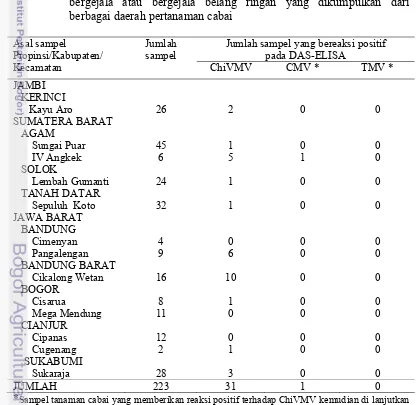
[image:35.595.98.516.338.743.2]Sebanyak 223 sampel diperoleh dari pertanaman cabai dari 13 lokasi pengamatan. Sampel daun cabai terdiri atas daun yang menunjukkan gejala belang ringan dan daun yang tidak menunjukkan gejala. Semua sampel dari lapangan dideteksi dengan DAS-ELISA menggunakan antiserum ChiVMV dan menunjukkan infeksi ChiVMV pada 31 sampel (31/223). Selanjutnya 31 sampel tersebut dideteksi dengan DAS-ELISA menggunakan antiserum CMV dan infeksi CMV hanya terdeteksi pada satu sampel (1/31). Selanjutnya 30 sampel yang terinfeksi ChiVMV tanpa CMV dideteksi dengan DAS-ELISA menggunakan antiserum TMV dan ternyata tidak ada sampel yang terinfeksi TMV (0/30) (Tabel 3.1).
Tabel 3.1 Deteksi Chili veinal mottle virus pada sampel tanaman cabai tidak bergejala atau bergejala belang ringan yang dikumpulkan dari berbagai daerah pertanaman cabai
Asal sampel
Propinsi/Kabupaten/ Kecamatan
Jumlah sampel
Jumlah sampel yang bereaksi positif pada DAS-ELISA
ChiVMV CMV * TMV *
JAMBI KERINCI
Kayu Aro 26 2 0 0
SUMATERA BARAT AGAM Sungai Puar IV Angkek SOLOK Lembah Gumanti TANAH DATAR Sepuluh Koto
45 6 24 32 1 5 1 1 0 1 0 0 0 0 0 0 JAWA BARAT BANDUNG Cimenyan Pangalengan BANDUNG BARAT Cikalong Wetan BOGOR Cisarua Mega Mendung CIANJUR Cipanas Cugenang SUKABUMI Sukaraja 4 9 16 8 11 12 2 28 0 6 10 1 0 0 1 3 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
JUMLAH 223 31 1 0
Metode DAS-ELISA terbukti dapat digunakan untuk kegiatan eksplorasi isolat lemah karena mampu mendeteksi infeksi virus walaupun gejala penyakit sangat lemah atau bahkan tidak tampak. Metode ELISA telah banyak digunakan untuk mendeteksi berbagai virus tanaman karena sangat sensitif (Clark dan Adam 1977), dan dapat mendeteksi infeksi virus yang tidak menimbulkan gejala (Kositratana et al. 1991). Dilaporkan oleh Kosaka et al. (2006) bahwa ZYMV strain lemah (ZYMV-2002) menimbulkan gejala sangat lemah atau tidak bergejala pada tanaman, tetapi setelah dikonfirmasi dengan DAS-ELISA menunjukkan hasil yang positif. Metode ELISA menjadi salah satu teknik serologi yang populer karena selain sensitif juga mudah dilakukan,akurat, simpel dan tidak memerlukan biaya tinggi(Clark dan Adam 1977).
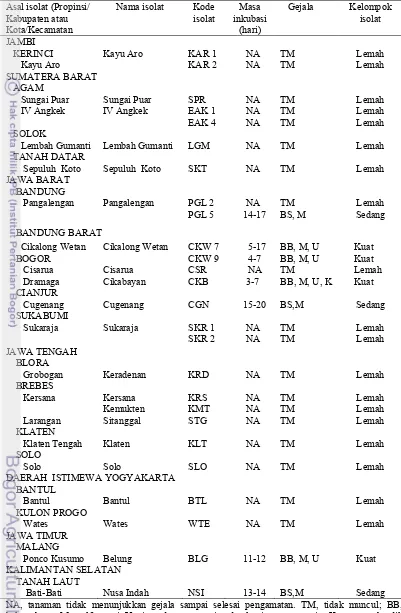
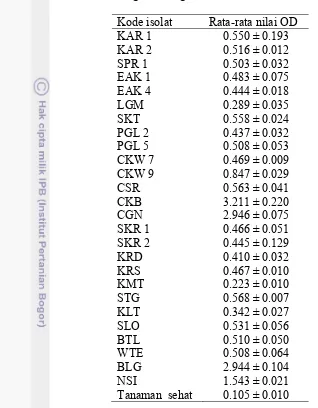
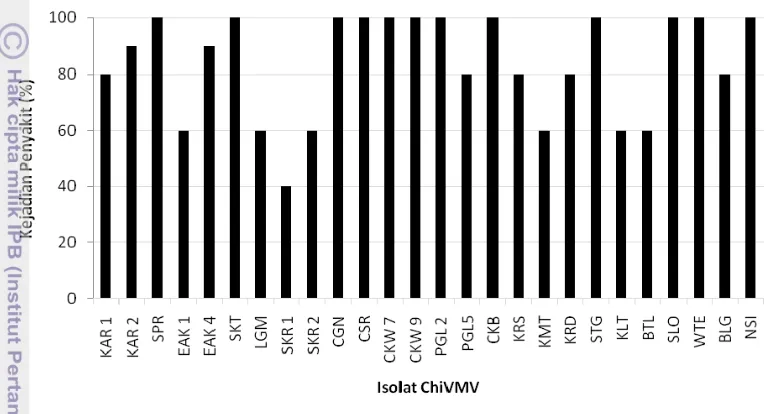
Kadar virus pada tanaman yang terinfeksi ditunjukkan oleh nilai optical density (OD) pada 405 nm. Semakin tinggi nilai OD semakin tinggi pula kadar virus dalam jaringan tanaman (Pacheco et al. 2003). Sebanyak 15 isolat ChiVMV dengan nilai OD tertinggi atau terendah yaitu isolat KAR 1, KAR 2, SPR, EAK 1, EAK 4, SKT, LGM, SKR 1, SKR 2, CGN, CSR, CKW 7, CKW 9, PGL 2, PGL 5 (Lampiran 1) dipilih untuk disertakan dalam pengujian pengelompokan isolat ChiVMV.
Pengelompokan Isolat ChiVMV
Sebanyak 26 isolat ChiVMV yang terdiri dari 15 isolat lapangan (KAR 1, KAR 2, SPR, EAK 1, EAK 4, SKT, LGM, SKR 1, SKR 2, CGN, CSR, CKW 7, CKW 9, PGL 2, dan PGL 5) dan 11 isolat koleksi Laboratorium Virologi (CKB, KRS, KMT, KRD, STG, KLT, SLO, BTL, WTE, BLG, dan NSI) digunakan pada tahapan pengelompokan isolat ChiVMV. Inokulasi isolat-isolat ChiVMV pada cabai genotipe IPB C13 dilakukan secara mekanis dan gejala diamati selama empat minggu. Menurut Aguilar et al. (2000) waktu satu sampai empat minggu sudah cukup bagi virus untuk menginfeksi tanaman secara sistemik. Sebanyak 19 isolat tidak menunjukkan gejala sampai akhir pengamatan, sementara tujuh isolat menunjukkan gejala pada cabai IPB C13 (Tabel 3.2).
Isolat ChiVMV menimbulkan infeksi sistemik dengan berbagai variasi gejala. Isolat PGL 5, CGN, dan NSI menimbulkan gejala belang sedang, dan malformasi daun. Isolat CKW 7, CKW 9, dan BLG menimbulkan gejala belang berat, malformasi, ujung daun meruncing. Isolat CKB menunjukkan gejala belang berat, malformasi, ujung daun meruncing, dan tanaman kerdil (Gambar 3.1). Gejala yang sama pada tanaman cabai yang terinfeksi ChiVMV dilaporkan oleh Weeraratne dan Yapa (2002), Siriwong et al. (1995), dan Wang et al. (2006) yaitu belang hijau tua, malformasi daun, dan daun menjadi kecil.
Gambar 3.1 Gejala infeksi Chili veinal mottle virus pada tanaman cabai genotipe IPB C13. A) tidak menunjukkan gejala, B) belang sedang, malformasi, C) belang berat dan malformasi, D) ujung daun meruncing dan lamina menyempit.
Tabel 3.2 Pengelompokan isolat Chili veinal mottle virus berdasarkan respon tanaman cabai IPB C13
Asal isolat (Propinsi/ Kabupaten atau Kota/Kecamatan
Nama isolat Kode isolat Masa inkubasi (hari) Gejala Kelompok isolat JAMBI KERINCI Kayu Aro
Kayu Aro KAR 1
KAR 2 NA NA TM TM Lemah Lemah SUMATERA BARAT AGAM Sungai Puar IV Angkek SOLOK Lembah Gumanti TANAH DATAR Sepuluh Koto
Sungai Puar IV Angkek
Lembah Gumanti
Sepuluh Koto
SPR EAK 1 EAK 4 LGM SKT NA NA NA NA NA TM TM TM TM TM Lemah Lemah Lemah Lemah Lemah JAWA BARAT BANDUNG Pangalengan Pangalengan PGL 2 PGL 5 NA 14-17 TM BS, M Lemah Sedang BANDUNG BARAT Cikalong Wetan BOGOR Cisarua Dramaga CIANJUR Cugenang SUKABUMI Sukaraja Cikalong Wetan Cisarua Cikabayan Cugenang Sukaraja CKW 7 CKW 9 CSR CKB CGN SKR 1 SKR 2 5-17 4-7 NA 3-7 15-20 NA NA
BB, M, U BB, M, U TM
BB, M, U, K
BS,M TM TM Kuat Kuat Lemah Kuat Sedang Lemah Lemah JAWA TENGAH BLORA Grobogan BREBES Kersana Larangan KLATEN Klaten Tengah SOLO Solo Keradenan Kersana Kemukten Sitanggal Klaten Solo KRD KRS KMT STG KLT SLO NA NA NA NA NA NA TM TM TM TM TM TM Lemah Lemah Lemah Lemah Lemah Lemah DAERAH ISTIMEWA YOGYAKARTA
BANTUL Bantul KULON PROGO Wates JAWA TIMUR MALANG Ponco Kusumo Bantul Wates Belung BTL WTE BLG NA NA 11-12 TM TM
BB, M, U
Lemah Lemah Kuat KALIMANTAN SELATAN TANAH LAUT
Bati-Bati Nusa Indah
NSI
13-14
BS,M Sedang
Tabel 3.3 Akumulasi virus direpresentasikan dengan nilai OD 405 nm hasil DAS-ELISA dari daun tanaman cabai IPB C13 pada 28 hari setelah inokulasi dengan berbagai isolat Chili veinal mottle virus
Kode isolat Rata-rata nilai OD KAR 1
KAR 2
0.550 ± 0.193 0.516 ± 0.012 SPR 1
EAK 1 EAK 4 LGM SKT PGL 2 PGL 5 CKW 7 CKW 9 CSR CKB
0.503 ± 0.032 0.483 ± 0.075 0.444 ± 0.018 0.289 ± 0.035 0.558 ± 0.024 0.437 ± 0.032 0.508 ± 0.053 0.469 ± 0.009 0.847 ± 0.029 0.563 ± 0.041 3.211 ± 0.220 CGN
SKR 1 SKR 2 KRD
2.946 ± 0.075 0.466 ± 0.051 0.445 ± 0.129 0.410 ± 0.032 KRS
KMT STG KLT SLO
0.467 ± 0.010 0.223 ± 0.010 0.568 ± 0.007 0.342 ± 0.027 0.531 ± 0.056 BTL
WTE BLG
0.510 ± 0.050 0.508 ± 0.064 2.944 ± 0.104 NSI
Tanaman sehat
1.543 ± 0.021 0.105 ± 0.010
ketimun menunjukkan replikasi strain lemah ZYMV-2002 sangat rendah dibanding strain kuat ZYMV Z5-1 (Kosaka et al. 2006). Strain lemah ZYMV-WK tetap bertahan walaupun titernya rendah pada tanaman labu (Lin et al. 2007), dan tanaman labu yang diinokulasi dengan isolat lemah ZYMV tidak menimbulkan gejala dan tidak mempengaruhi pertumbuhan vegetatif tanaman (Lecog et al.
[image:39.595.111.497.192.399.2]1991).
Gambar 3.2 Kejadian penyakit pada tanaman cabai genotipe IPB C13 yang diinokulasi dengan berbagai isolat Chili veinal mottle virus.
Berdasarkan gejala pada tanaman cabai genotipe IPB C13, maka isolat-isolat ChiVMV dapat dibedakan menjadi isolat-isolat lemah (KAR, SPR, EAK, SKT, LGM, SKR, CSR, PGL, KRS, KMT, KRD, STG, KLT, BTL, SLO, dan WTE), isolat sedang (CGN dan NSI), dan isolat kuat (CKW, CKB, dan BLG). Isolat-isolat lemah ChiVMV yang diperoleh pada penelitian ini selanjutnya dapat digunakan sebagai kandidat agens proteksi silang untuk melindungi tanaman dari infeksi virus strain kuat. Menurut Lin et al.(2007) kemampuan strain lemah untuk melindungi tanaman dari kerusakan yang ditimbulkan oleh strain kuat dari virus yang sekerabat telah diketahui. Beberapa tahapan penelitian perlu dilakukan untuk memilih isolat-isolat lemah ChiVMV yang potensial, diantaranya adalah melalui pengujian kisaran inang isolat ChiVMV (Shah et al. 2008), interaksi antara isolat lemah dan isolat kuat (Roossinck 2005), dan evaluasi proteksi silang (Kosaka et al. 2006).
Kesimpulan
4 KISARAN INANG
Chili veinal mottle virus
ISOLAT LEMAH
(Host Range Study of Weak Isolates of
Chili veinal mottle virus
)
Abstrak
Chili veinal mottle virus (ChiVMV) merupakan salah satu penyakit penting pada tanaman cabai dan Solanaceae lainnya. Penelitian dilakukan di rumah kaca untuk mempelajari kisaran inang delapan ChiVMV isolat lemah (KAR, SKT, SPR, CSR, PGL, SLO, STG, dan WTE). Delapan genotipe cabai (IPB C2, IPB C13, IPB C120, Laris, IPB C8, Bara, California Wonder, dan Yolo Wonder) dan sebelas tanaman Solanaceae (Capsicum frutescens, Datura metel, D. stramonium, Lycopersicon esculentum, Nicotiana benthamiana, N. tabacum cv. Samsun, N. tabacum cv. White Burley, dan N. tabacum cv. Xanthi, Physalis floridana, Solanum melongena,