PENGARUH PENGKAYAAN ZOOXANTHELLAE
DARI BERBAGAI SUMBER INANG TERHADAP
PROSES TRANSLOKASI DAN KALSIFIKASI
BINATANG KARANG
PUJIONO WAHYU PURNOMO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Pengaruh Pengkayaan Zooxanthellae dari Berbagai Sumber Inang Terhadap Proses Translokasi dan Kalsifikasi Binatang Karang adalah karya saya dengan arahan dari Komisi Pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, 19 Juli 2011
ABSTRACT
PUJIONO WAHYU PURNOMO. The Influence of Zooxanthellae Enrichment from Some Host Concerning Translocation Process and Coral Calsification (Advisor by : DEDI SOEDHARMA as Chairman, NEVIATY P. ZAMANI and HARPASIS S. SANUSI as Members).
Translocation process are the biotechnique activity to cross of clade on coral bleaching by enrichment of zooxanthellae. Succesfully of translocation process hope can support idea of coral recovery after bleaching. The research do manner sequensial in the three steps. The first step is exploring of zooxanthellae growth and examine genetic diversity be based on the artificial environment and purposed for : (a) Evaluating factor needed irradiance, temperature and nutrient that supporting of zooxanthellae growth, (b) explore genetic diversity of zooxanthellae based on profill it’s DNA from sea anemone, Tridacna, Acropora, Favites and Goniastrea, (c) Explore the effort optimalization growing of zooxanthellae. The second step is examine degradation polyp histologis of
Goniastrea aspera concerning application of temperature levels and endurance after bleaching. This research aims to : (a) investigate effect temperature concerning change content zooxanthellae density in the polyp histologies, (b) investigate effect temperature on process change of polyp histologies and (c) the evaluation ability of coral after bleaching. The third step is explore translocation several clade of zooxanthellae on the survival and growth rate of Goniastrea aspera. Research aims to : (a) Evaluating effect of zooxanthellae enrichment on the growth and resettlement of zooxanthellae on the polyp histology and (b) Explore the influence zooxanthellae translocation on the calcification of Goniastrea aspera.
Research did in the Main Center of Brackishwater Aquaculture Development Jepara, Ecodevelopment Coastal FPIK UNDIP and Southern of Pulau Panjang Waters at Jepara. Result of the first research are : (a) The irradiance, temperature and nutrient for supporting growth of zooxanthellae are : green radiance (with comparison 84,56 - 102,78 µmol quanta m-2 dt-1), 20 – 23oC and initial nutrient 200 µM of NaNO3, (b)
Diversity variation clade from host (organism test) are clade A, clade B and clade C, (c) In controlling environment, the potention of peak growth of third clade are can be maintenanced with adding NO3-N amount 0,0445 mg/l and repeat again for 16 days.
Result of the second research are : (a) Optimum temperatur for resistent of zooxanthellae in the polyp histologist of Goniastrea aspera is 36oC until 6 hours, (b) More than optimal temperature and incubation time efected on the histologist degradation of the polyp of Goniastrea aspera, zooxanthellae release and coral death, (c) Recovery organism (Goniastrea aspera) examine on the condition partial bleaching and (d) Coral recovery marked with zooxanthellae regulation process in the polyp hystologies.and to achieve perfect systematic after week 12th. Result of the third research are : a) In the artificial waters, translocation zooxanthellae to polyp tissue of
Goniastrea aspera occured at day 17 and more fast in the natural waters; b) In the controlling of temperature environment on translocation provided positive response of
Goniastrea aspera’s normal life, relocation and growth rate of zooxanthellae as in nature and c) recognition, resettlement, and growth process of zooxanthellae made it possible for Goniastrea aspera to grow normally in natural waters.
RINGKASAN
PUJIONO WAHYU PURNOMO. Pengaruh Pengkayaan Zooxanthellae dari Berbagai Sumber Inang Terhadap Proses Translokasi dan Kalsifikasi Binatang Karang. (Di bawah bimbingan : DEDI SOEDHARMA sebagai Ketua, NEVIATY P. ZAMANI dan HARPASIS S. SANUSI sebagai anggota).
Karang merupakan biota laut yang mampu membentuk ekosistem. Ancaman degradasinya semakin meningkat tidak saja oleh pengaruh alami akan tetapi juga pengaruh antropogenik. Kejadian ini sangat ironis dengan keberadaannya yang sangat penting baik terhadap fungsinya sebagai kawasan penyangga kehidupan internal dan eksternal ekosistem bentukannya maupun fungsinya sebagai penyangga sistem pulau. Pengaruh alami yang menonjol terhadap penurunan kualitas terumbu karang dalam dekade terakhir adalah adanya pengaruh pemanasan. Efek pemanasan perairan terhadap karang adalah timbulnya pemutihan atau bleaching. Pemanasan perairan laut dalam kurun waktu lama dikhawatiran menyebabkan kerusakan terumbu karang secara permanen. Kekhawatiran ini perlu dijembatani dengan penelusuran terhadap proses bleaching dan pemulihannya. Dukungan sifat fluktuasi air laut serta potensi strategi pengaturan faali karang melalui proses bleaching diperkirakan dapat mengurangi kekhawatiran tersebut.
Penelitian ini dilakukan secara sekuensial dalam tiga tahap. Tahap pertama adalah Kajian pertumbuhan zooxanthellae secara massal dan uji keragaman genetik berdasarkan penumbuhan di lingkungan binaan. Kajian tahap pertama ini bertujuan untuk : (a) Mengevaluasi faktor kebutuhan cahaya, suhu dan nutrien optimum yang mendukung pertumbuhan zooxanthellae, (b) Mengkaji diversitas zooxanthellae berdasarkan profil DNAnya dari sumber inang sea anemon, tridacna, Acropora, Favites dan Goniastrea aspera dan (c) Mengkaji upaya optimalisasi pertumbuhan zooxanthellae secara massal. Tahap kedua adalah uji degradasi jaringan polip Goniastrea aspera terhadap terapan berbagai taraf suhu dan ketahannya pasca bleaching. Kajian tahap dua bertujuan untuk : (a) Mengkaji efek suhu terhadap perubahan kadar zooxanthellae pada jaringan polip karang; (b) Mengkaji efek suhu terhadap proses perubahan jaringan polip karang dan (c) Mengevaluasi kemampuan pulih karang pasca bleaching. Tahap ketiga adalah kajian translokasi beberapa clade zooxanthellae terhadap sintasan dan pertumbuhan karang Goniastrea aspera. Kajian ini bertujuan untuk (a)
Mengevaluasi efek pengkayaan zooxanthellae terhadap perkembangan
zooxanthellae pada jaringan polip Goniastrea aspera; (b) Mengevaluasi efek pengkayaan zooxanthellae terhadap translokasi pada jaringan polip binatang karang dan (c) Mengkaji pengaruh translokasi terhadap pertumbuhan karang.
102,78 µmol quanta m-2 dt-1, kisaran suhu yang mendukung adalah antara 20oC
hingga 23oC dengan penambahan nutrien awal 200 µM NaNO3. (b) Variasi
keragaman clade dari sumber inang Sea anemon, Tridacna, Acropora, Favites dan
Goniastrea adalah clade A, clade B dan clade C. (c) Pada kondisi lingkungan yang terkendali, maka potensi pertumbuhan puncak zooxanthellae clade A, B dan C dapat
dipertahankan dengan penambahan ulang NO3-N sebesar 0,0445 mg/l dalam selang
waktu 16 hari.
Kajian tahap kedua dilakukan baik dalam media binaan di lingkungan BBPBAP Jepara dan Laboratorium Pengembangan Wilayah Pantai FPIK UNDIP Jepara dan lingkungan perairan terumbu karang selatan Pulau Panjang Jepara. Perlakuan yang diterapkan adalah dengan memberikan shock suhu kepada biota uji Goniastrea aspera dalam 4 taraf yaitu 40oC, 36oC, 32oC dan 28oC. Hasil yang menonjol dari kajian tahan ini adalah : (a) Suhu optimum bagi zooxanthellae yang
masih dapat bertahan pada jaringan polip karang Goniastrea aspera adalah 36oC
selama 6 jam, (b) Di atas suhu optimum terjadi proses degradasi jaringan polyp, pelepasan zooxanthellae dan kematian karang, (c) Pemulihan biota uji hanya dapat terjadi pada kondisi pemutihan parsial (d) Pemulihan karang ditandai dengan terjadinya proses regulasi zooxanthellae di dalam jaringan polip karang dan mencapai penataan sempurna setelah minggu ke 12.
Kajian tahap ketiga dilakukan baik dalam media binaan di lingkungan BBPBAP Jepara dan Laboratorium Pengembangan Wilayah Pantai FPIK UNDIP Jepara dan lingkungan perairan terumbu karang selatan Pulau Panjang Jepara. Hasil penelitian menunjukkan bahwa : a) Translokasi zooxanthellae pada
Goniastrea aspera pasca bleaching pada lingkungan binaan terjadi pada hari ke 17, sedangkan di perairan alami dapat terjadi lebih dini; b) Translokasi yang terkendali oleh kondisi lingkungan khususnya suhu memberikan respon positif terhadap proses
relokasi dan pertumbuhan zooxanthellae pada jaringan polyp Goniastrea aspera; c)
Proses recognisi, resettlement dan pertumbuhan zooxanthellae memungkinkan karang
Goniastrea aspera dapat tumbuh secara normal di media alami.
©
Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-UndangDilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
PENGARUH PENGKAYAAN ZOOXANTHELLAE
DARI BERBAGAI SUMBER INANG TERHADAP
PROSES TRANSLOKASI DAN KALSIFIKASI
BINATANG KARANG
PUJIONO WAHYU PURNOMO
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Kelautan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi Ujian Tertutup :
1. Dr. Ir. Etty Riani, MS : Staf pengajar pada Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor.
2. Dr. Ir. Fajar Basuki, MS : Staf pengajar pada Fakultas Perikanan dan Ilmu Kelautan, Universitas Diponegoro.
Penguji Luar Komisi Ujian Terbuka :
1. Prof. Dr. R. Suharsono : Kepala Pusat P2O LIPI Jakarta
Judul Disertasi : Pengaruh Pengkayaan Zooxanthellae dari Berbagai Sumber Inang Terhadap Proses Translokasi dan Kalsifikasi Binatang Karang
Nama : Pujiono Wahyu Purnomo
NIM : C661020071
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Dedi Soedharma, DEA Ketua
Dr. Ir. Neviaty P. Zamani, M.Sc. Prof. Dr. Ir. Harpasis S. Sanusi, M.Sc.
Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Ilmu Kelautan
Dr. Ir. Neviaty P. Zamani, M.Sc. Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karuaniaNya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Mei 2004 ini adalah translokasi zooxanthellae, dengan judul Pengaruh Pengkayaan Zooxanthellae dari Berbagai Sumber Inang Terhadap Proses Translokasi dan Kalsifikasi Binatang Karang.
Disertasi ini memuat 3 bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Artikel yang diajukan untuk diterbitkan pada jurnal ilmiah tersebut adalah pada Bab 3 berjudul Uji Keragaman Genetik Zooxanthellae dari beberapa Sumber Inang dan Kajian Pertumbuhannya di Lingkungan Binaan dan Bab 5 berjudul Kajian Translokasi Beberapa Clade Zooxanthellae terhadap Sintasan dan Pertumbuhan Karang Goniastrea aspera. Satu dari tiga jurnal yang dapat dikembangkan dari penelitian ini telah diterbitkan di Majalah Saintek FPIK UNDIP, sementara lainnya masih menunggu penerbitan.
Pada kesempatan ini penulis menyampaikan ucapan terima kasih dan penghargaan setinggi-tingginya kepada :
1. Prof. Dr. Dedi Soedharma, DEA, selaku ketua komisi pembimbing yang telah memberikan curahan waktu, nasehat, arahan dan motivasi secara terus menerus dengan penuh dedikasi dari awal perencanaan penelitian sampai selesainya disertasi ini;
2. Dr. Ir. Neviaty P. Zamani, M.Sc dan Prof. Dr. Ir. Harpasis S. Sanusi, M.Sc, selaku anggota komisi pembimbing yang telah memberikan bimbingan, arahan, saran, semangat dan koreksian-kooreksian yang kritis dan tajam sehingga menambah kualitas disertasi ini;
3. Dr. Ir. Neviaty P. Zamani, M.Sc, selaku Ketua Program Studi Ilmu Kelautan beserta staff administrasinya atas segala bantuan administratif dan akademik selama penyelesaian studi ini;
4. Rektor Institut Pertanian Bogor dan Dekan Sekolah Pascasarjana IPB yang telah memberikan kesempatan kepada penulis untuk mengikuti program Doktor di Institut Pertanian Bogor.
5. Rektor Universitas Diponegoro dan Dekan Fakultas Perikanan dan Ilmu Kelautan UNDIP, atas ijin pendidikan yang diberikan kepada penulis untuk mengikuti program doktor di Institut Pertanian Bogor.
6. Dr.Ir. I Wayan Nurjaya, M.Sc, mewakili Wakil Dekan FPIK-IPB selaku pimpinan sidang ujian tertutup; Dr. Ir. Etty Riani, MS dari Fakultas Perikanan dan Ilmu Kelautan IPB dan Dr. Ir. Fajar Basuki dari Fakultas Perikanan dan Ilmu Kelautan UNDIP selaku penguji dalam ujian tertutup yang telah menambah wawasan pengetahuan keilmuan penulis,
7. Prof. Dr. Ir. Indra Jaya, M.Sc, selaku pimpinan sidang ujian terbuka; Prof. Dr. Soeharsono dari Pusat Penelitian Oseanografi (P2O) LIPI dan Prof. Dr. Ir. Muhammad Zainuri, DEA dari Fakultas Perikanan dan Ilmu Kelautan UNDIP, selaku penguji dalam ujian terbuka yang telah menambah bobot pengetahuan keilmuan penulis,
memberikan ijin penelitian. Ir. Bambang Widyo Prastowo, MSi dan staf di Lingkungan Laboratorium Penyakit Ikan dan Genetika; Ir. Fairus M. Soni, M.Sc dan staf di Lingkungan Laboratorium pakan alami BBPBAP yang berkenan membantu pelaksanaan studi ini,
9. Ir. Imam Triarso, MS, selaku Kepala Laboratorium Pengembangan Wilayah Pantai FPIK UNDIP beserta staf yang berkenan memberikan bantuan selama penelitian ini berlangsung;
10. Rekan-rekan mahasiswa Pascasarjana Program Studi Ilmu Kelautan IPB khususnya angkatan 2002 atas kebersamaan dan kerjasamanya selama menempuh pendidikan.
11. Orang Tua dan Mertua saya, adik serta seluruh keluarga yang telah memberikan doa, semangat dan kasih sayangnya selama penulis menempuh pendidikan Doktor di Institut Pertanian Bogor.
12. Istriku Sulistiyaningsih dan putra putriku Tantri Bagus Satrio, S,Ked; Azafilmi Hakiim, S.T.; Arsyzilma Hakiim, S.Ked.; Willy Noorhasheed dan Athasalma Monica atas segala pengorbanan dan dorongan doa, semangat serta kasih sayangnya yang diberikan selama penulis menempuh pendidikan.
Penulis menyadari bahwa disertasi ini masih jauh dari kesempurnaan, namun demikian penulis berharap semoga disertasi ini bermanfaat.
Bogor, 19 Juli 2011
RIWAYAT HIDUP
Penulis dilahirkan di Demak pada tanggal 11 Mei 1962 dari ayah Zoechdi Soewoto (Alm) dan ibu Soemirah (Alm). Penulis merupakan putra pertama dari empat bersaudara. Pendidikan sarjana ditempuh di Program Studi Manajemen Sumberdaya Hayati Jurusan Perikanan Fakultas Peternakan dan Perikanan Universitas Diponegoro lulus pada tahun 1986. Pada tahun 1989 penulis diterima di Program Studi Ilmu Perairan pada program Pascasarjana IPB dan menamatkannya pada tahun 1992. Kesempatan untuk melanjutkan ke program doktor pada program studi Ilmu Kelautan pada perguruan tinggi yang sama diperoleh pada tahun 2002. Bea siswa pendidikan pascasarjana diperoleh dari Due Like.
Penulis bekerja sebagai staf pengajar pada program studi Manajemen Sumberdaya Perairan Jurusan Perikanan Universitas Diponegoro. Mata kuliah yang diampu oleh peneliti adalah Avertebrata, Biologi Laut dan Pengelolaan Terumbu Karang.
Karya-karya ilmiah yang diterbitkan sebagai bagian dari penelitian program doktor adalah Model Kehidupan Zooxanthellae dan Penumbuhan Massalnya pada Media Binaan di Jurnal Saintek Perikanan Universitas Diponegoro Semarang. Sementara itu karya ilmiah lain dengan judul Kajian Pertumbuhannya di Lingkungan Binaan dan Kajian Translokasi Beberapa Clade
Zooxanthellae terhadap Sintasan dan Pertumbuhan Karang Goniastrea
xix
DAFTAR ISI
DAFTAR TABEL ... xxi
DAFTAR GAMBAR ... xxiii
DAFTAR LAMPIRAN ... xv
PENDAHULUAN Latar Belakang ... 1
Ruang Lingkup Penelitian ... 4
Tujuan Penelitian ... 11
Manfaat Penelitian ... 11
Hipotesis Penelitian ... 21
Lokasi Penelitian ... 12
TINJAUAN PUSTAKA Fakta dan Faktor-faktor Destruktif bagi Karang ... 15
Bioekologi Zooxanthellae dan Karang ... 17
Peranan Faktor Fisika Kimia Perairan terhadap Hubungan Karang dan Zooxanthellae ... 22
Proses Simbiosis Zooxanthellae dan Peran Fungsionalnya ... 35
UJI KERAGAMAN GENETIK ZOOXANTHELLAE DARI BEBERAPA SUMBER INANG DAN KAJIAN PERTUMBUHANYA DI LINGKUNGAN BINAAN Abstrak ... 41
Pendahuluan ... 42
Metodologi... 46
Hasil Percobaan ... 55
Pembahasan ... 66
Simpulan ... 79
Daftar Pustaka ... 81
UJI DEGRADASI JARINGAN POLIP Goniastrea aspera TERHADAP TERAPAN BERBAGAI TARAF SUHU dan KETAHANANNYA PASCA PEMUTIHAN Abstrak ... 89
Pendahuluan ... 90
Metodologi... 93
Hasil Percobaan ... 102
Pembahasan ... 115
Simpulan ... 126
xx
KAJIAN TRANSLOKASI BEBERAPA STRAIN ZOOXANTHELLAE TERHADAP SINTASAN DAN PERTUMBUHAN KARANG
Goniastrea aspera
Abstrak ... 133
Pendahuluan ... 134
Metodologi ... 136
Hasil Percobaan ... 144
Pembahasan ... 158
Simpulan ... 166
Daftar Pustaka ... 167
PEMBAHASAN UMUM SIMPULAN DAN SARAN Simpulan ... 195
Saran... 196
DAFTAR PUSTAKA ... 199
xxi
DAFTAR TABEL
Halaman
1 Rangkuman tentang Identifikasi Coral Bleaching Akibat Beberapa
Stressor ... 16
2 Kondisi Kesehatan Terumbu Karang di Indonesia ... 18
3 Beberapa Zooxanthellae yang berasosiasi dengan Cnidaria ... 20
4 Komposisi Kimia Media Agar Allen untuk Kultur Biakan Murni Zooxanthellae... 48
5 Komposisi Kimia Media Agar Bacto untuk Kultur Biakan Murni Zooxanthellae... 48
6 Peubah yang diukur, metoda dan periode pengukuran ... 54
7 Respon Penumbuhan Beberapa Zooxanthellae pada Media Agar ... 55
8 Ragam DNA Zooxanthellae dari Beberapa Inang ... 62
9 Densitas Zooxanthellae tertinggi hasil kultur dari Beberapa Inang ... 63
10 Komposisi Komunitas Endolitik Dan Aktivitas Metaboliknya dalam Lingkungan Konstituen Rangka Karang Hidup ... 75
11 Manajemen Koloni Contoh untuk Setiap Ulangan perlakuan ... 96
12 Peubah yang diukur, metoda dan periode pengukuran ... 101
13 Hasil Analisis Kualitas Air Perairan Terumbu Karang (Lokasi Inkubasi Alamiah) ... 115
14 Pengelolaan Sample Tiap Perlakuan ... 139
15 Peubah yang diukur, metoda dan periode pengukuran ... 143
16 Diversitas DNA pada Polip Goniastrea aspera pasca bleaching dalam masa pengeraman 10 hari media Pengkayaan Zooxanthellae ... 148
17 Diversitas DNA pada Polip Goniastrea aspera pasca bleaching dalam masa pengeraman 17 hari media Pengkayaan Zooxanthellae ... 152
18 Hasil analisis Beberapa Peubah Lingkungan Pengeraman Biota Uji Goniastreas aspera ... 163
19 Variasi Clade Berdasarkan Jenis, Sumber dan Sebaranny ... 174
20 Tipe Kelarutan Konsentrasi Nitrogen Inorganik (dalam mol/l) di Beberapa Lokasi Terumbu Karang di Dunia ... 187
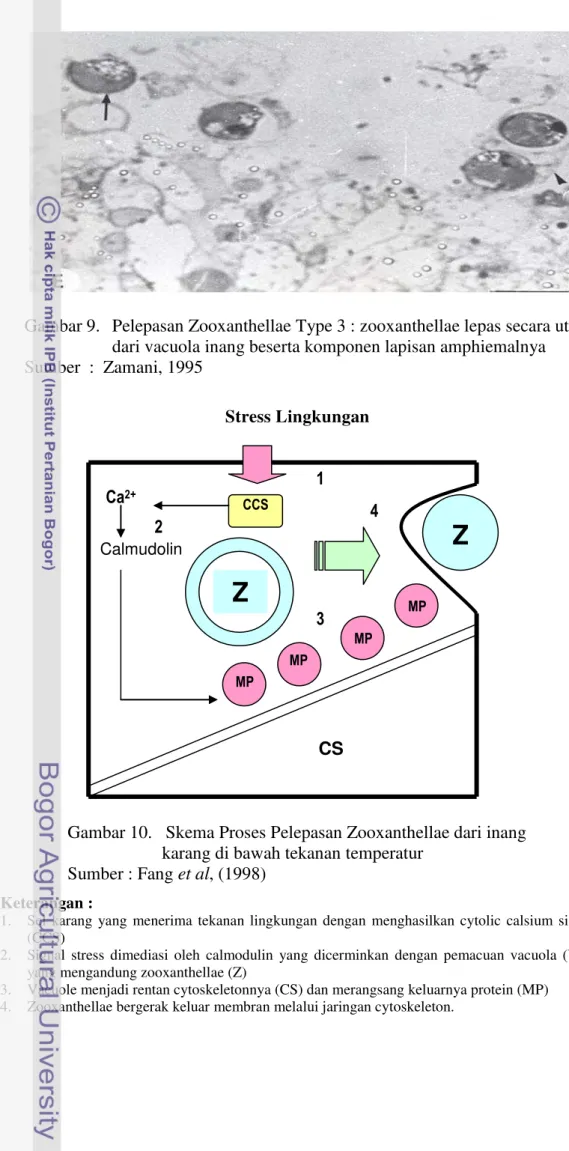
xxiii 7 Pelepasan Zooxanthellae Type 1 : zooxanthellae menyelimuti
cytoplasma Inang ... 33 8 Pelepasan Zooxanthellae Type 2 : zooxanthellae secara parsial
atau menyeluruh dikelilingi oleh sisa-sisa jaringan ... 33 9 Pelepasan Zooxanthellae Type 3 : zooxanthellae lepas secara utuh
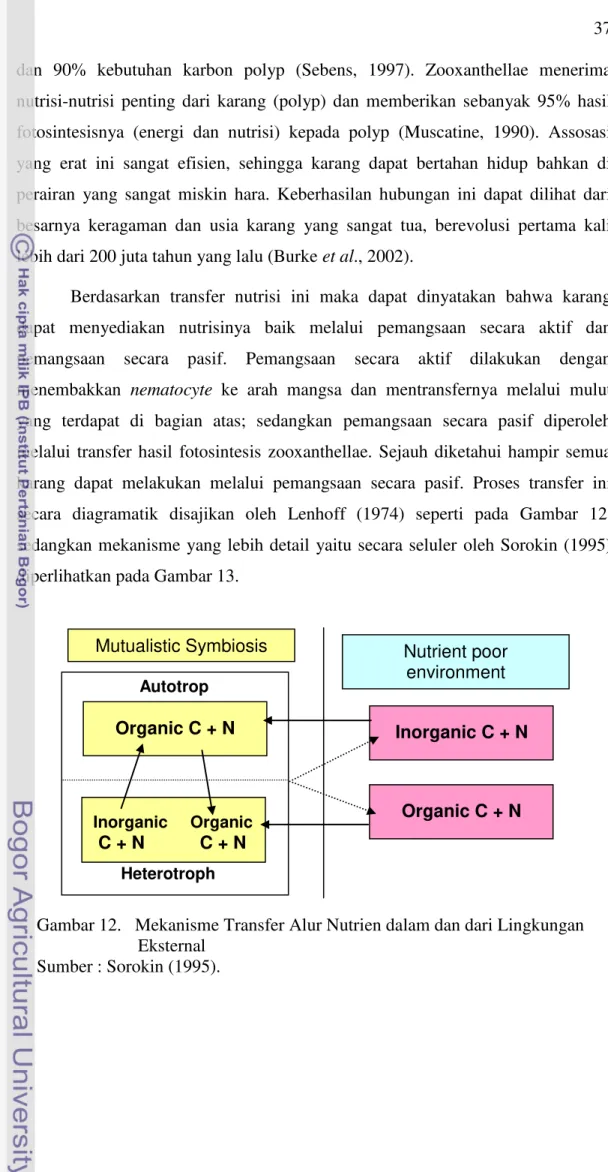
dari vacuola inang beserta komponen lapisan amphiemalnya ... 34 10 Skema Proses Pelepasan Zooxanthellae dari inang karang
di bawah tekanan suhu ... 34 11 Penampang Lintang Jaringan karang dan Konsentrasi
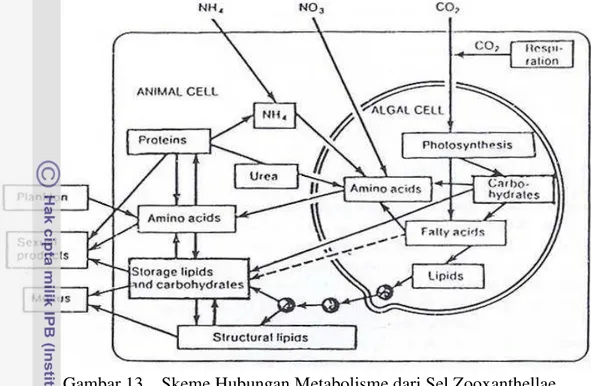
Zooxanthellae di dalamnya ... 36 12 Mekanisme Transfer Alur Nutrien dalam dan dari Lingkungan
Eksternal ... 37 13 Skema Hubungan Metabolisme dari Sel Zooxanthellae
dalam Inang Polip ... 38 14 Densitas Zooxanthellae hasil Adaptasi dari Media Inokulan ... .... 56 15 Pertumbuhan Zooxanthellae pada Berbagai Pengubahan
Kondisi Media ... 57 16 Genotip RFLP dari contoh zooxanthellae yang dimurnikan dari
beberapa sumber inang ... 60 17 Hubungan kekerabatan DNA dari beberapa jenis zooxanthellae
berdasarkan Analisis Cluster ... 61 18 Pertumbuhan Zooxanthellae antar Clade ... .... 64 19 Perubahan Kadar NO3-N selama Budidaya Zooxanthellae ... .... 65
20 Pertumbuhan Zooxanthellae pada Bak Massal dengan
Penambahan Nutrien ... 66 21 Pemiskinan NO3-N dan Perkembangan Zooxanthellae ... .... 76
22 Respon Penambahan NO3-N terhadap upaya
Mempertahankan Perkembangan Zooxanthellae ... 77 23 Perubahan Kandungan Zooxanthellae Karang Goniastrea aspera
padaBerbagai Perlakuan Suhu ... 103 24 Profil Jaringan Polip Karang Selama Proses Penerapan Berbagai Tingkat
xxiv
25 Sintasan Goniastrea aspera pasca Bleaching ... ... 114 26 Perkembangan Zooxanthellae dari Adaptasi Karang Goniastrea
aspera pada Berbagai Tingkat Media Suhu ... 117 27 Proses Addisi Sel Zooxanthellae pada Jaringan Polip Karang ... ... 123 28 Sketsa Penataan Peralatan Uji Translokasi Zooxanthellae
Pada Goniastrea aspera yang telah dibleachingkan ... 138 29 Pola Pita DNA Zooxanthellae pada Jaringan Polip Goniastrea aspera
yang diperkaya dengan Zooxanthellae Clade A, B dan C pada masa
inkubasi 10 hari ... 147 30 Hubungan kekerabatan DNA dari beberapa jenis zooxanthellae
berdasarkan Analisis Cluster pada tahap uji diversitas DNA
zooxanthellae dari jaringan polip Goniastrea aspera pasca Pemutihan
dalam 10 hari waktu inkubasi pada media pengkayaan zooxanthellae ... 149 31 Pola Pita DNA Zooxanthellae pada Jaringan Polip Goniastrea aspera
yang diperkaya dengan Zooxanthellae Clade A, B dan C pada masa
inkubasi 17 hari ... 152 32 Hubungan kekerabatan DNA dari beberapa jenis zooxanthellae
berdasarkan Analisis Cluster pada tahap uji diversitas DNA
zooxanthellae dari jaringan polip Goniastrea aspera pasca Pemutihan
dalam 17 hari waktu inkubasi pada media pengkayaan zooxanthellae ... 153 33 Perkembangan Zooxanthellae pada Goniastrea aspera
dalam Masa Inkubasi di Perairan Alami ... 155 34 Perubahan Tampilan Jaringan Polip Karang selama masa
Pengeraman di Perairan Laut selama 6 minggu ... 156 35 Pertumbuhan Specimen Goniastrea aspera Berdasarkan
Sumber Pengkayaan Clade ... 158 36 Nilai Beberapa Peubah Fisika Kimia Perairan Binaan
tempat Pengeraman Biota uji pasca bleaching ... 164 37 Profil histologis jaringan polyp karang pada pengaruh pemanasan
Media pada suhu 40oC selama 6 jam dan suhu 36oC selama 12 jam ... 181 38 Bentukan Struktur Diameter Rongga pada Rangka Karang
xxv
DAFTAR LAMPIRAN
Halaman 1 Pencacahan Densitas Zooxanthellae pada Tahap Adaptasi ... 215 2 Hasil Uji antar perlakuan pada tiap tahap pertumbuhan zooxanthellae ... .... 217 3 Densitas Zooxanthellae pada percobaan pertumbuhan antar clade
dan tingkat nutrien ... 218 4 Analisis Perubahan NO3-N (mg/l) pada Media Budidaya Zooxanthellae... .... 221
5 Densitas Zooxanthellae dan analisis ragam model perubahan zooxanthellae pada penerapan level shock suhu ... 224 6 Analisis ragam densitas zooxanthellae pada polip karang Goniastrea
aspera Pada pengkayaan ragam clade (A, B dan C) pada minggu ke 15
PENDAHULUAN
Latar Belakang
Menurut Veron (1995), terumbu karang merupakan ekosistem khas daerah tropis dengan pusat penyebaran di wilayah Indo Pasifik. Terbatasnya penyebaran terumbu karang di perairan tropis dan bentang latitudinal dari wilayah selatan Jepang sampai dengan Australia dikendalikan oleh faktor temperatur dan sirkulasi permukaan (surface circulation) air. Penyebaran terumbu karang secara longitudinal dipengaruhi oleh adanya konektivitas berupa stepping stone (area penyebaran). Kombinasi antara faktor lingkungan fisik (temperatur dan sirkulasi air permukaan) dengan banyaknya jumlah area penyebaran yang terdapat di wilayah Indo Pasifik diperkirakan menjadi faktor pendukung luasnya terumbu karang di kawasan tersebut. Kini hampir 800 jenis karang yang tergolong kelompok Schleractinia telah dideskripsikan. Dari sejumlah karang yang ditemukan ini, 600 jenis berada di Asia Tenggara khususnya Indonesia dan Philipina (Burke, Selig dan Spalding, 2002), dan dengan pertimbangan luas kawasannya sebesar 34% (51% konstribusi kawasan terumbu karang Indonesia) dari total kawasan terumbu karang di dunia maka secara biogeografi kawasan ini dinyatakan sebagai pusat sebaran karang di dunia (Veron, 1995).
2
terumbu karang Samudera India, pemutihan karang terjadi pada tahun 1991 dan 1995. Di kawasan terumbu karang Laut Karibia khususnya di Jamaika juga dilaporkan terjadi pemutihan pada Tahun 1987, 1988, 1990 dan 1995 serta beberapa tempat lain.
Peningkatan kegiatan ekonomi di kawasan pesisir yang mempunyai potensi terumbu karang dan atau yang berdaya guna secara langsung di lingkungan terumbu karang merupakan bentuk pengaruh antropogenik terhadap keberadaan terumbu karang. Meskipun gangguan antropogenik terjadi pada variasi spasial maupun temporal, tetapi seringkali overlap (tumpang tindih) diantaranya memberikan pengaruh yang sangat besar terhadap terumbu karang. Di beberapa lokasi di pesisir Indonesia, adanya pengaruh antropogenik dapat mematikan atau memusnahkan ekosistem ini. Pengaruh antropogenik ini akan semakin besar dampaknya terhadap karang akibat tumpang tindih dengan pengaruh peningkatan temperatur air laut secara global. Kejadian ini mempunyai implikasi kronik dalam jangka panjang sebagaimana diperlihatkan oleh efek eutrofikasi. Fenemona ini akan menurun kepada gangguan seperti penyakit bakteri yang dapat mengakibatkan pemutihan karang sebagaimana terjadi akibat peningkatan temperatur (Rosenberg dan Ben-Haim, 2002). Efek gangguan campuran secara umum dapat mengganggu proses pertumbuhan karang, mempengaruhi rekruitmen serta proses regeneratif (Hughes dan Connell, 1999). Penurunan kualitas dan kuantitas ekosistem terumbu karang tersebut selanjutnya memberikan implikasi efek terhadap berbagai hal seperti menurunnya kelimpahan ikan (Wilkinson, 1999); eutrifikasi (Berner dan Izhaki, 1994; Stambler dan Vago, 1994; Wilkinson, 1999). Kejadian ini sangat ironis dengan keberadaannya yang sangat penting baik terhadap fungsinya sebagai kawasan penyangga kehidupan internal dan eksternal ekosistem bentukannya maupun fungsinya sebagai penyangga sistem pulau.
3
gejala alam yang kian akan terus mempengaruhi kehidupan terumbu karang. Berkenaan dengan fenomena di atas, maka pemutihan karang akan terus menjadi fenemena yang harus diterima oleh biota karang. Dalam kondisi fluktuasi ini karang akan terus melakukan proses adaptasi untuk mempertahankan diri terhadap kematian (Fitt et al. 2001). Salah satu faktor penting terkait dengan hal tersebut adalah kemungkinan terjadinya translokasi zooxanthellae pasca proses pemutihan,
sebagai bagian dari mekanisme adaptasi yang dilakukan oleh karang dalam mengantisipasi perubahan lingkungan eksternalnya. Ini menjadi pertanyaan utama dari kekhawatiran akan masa depan terumbu karang akibat tekanan yang luas. Tekanan yang luas ini tidak saja mengantisipasi efek pemanasan global yang bersifat alamiah juga akibat langsung dan tidak langsung dari pengaruh anthropogenik (Westmacott, et al., 2000). Selanjutnya dijelaskan bahwa pendekatan yang perlu dilakukan adalah dengan penerapan beberapa model pengelolaan secara terpadu. Namun dibalik itu semua, pada prinsipnya gangguan yang terjadi pada karang dalam suatu runtun waktu tertentu memerlukan proses pemulihan. Pernyataan Peters, (1997); Nystrom, Folke dan Moberg (2000) dan Fitt et al. (2001) perlu dicermati terhadap fenomena ini. Dikemukakan bahwa hasil dari perubahan ini mungkin mempunyai peran penting dalam rangka meningkatkan daya dukung yang akan berkompromi terhadap kemampuan terumbu karang mengatasi gangguan pada masa mendatang.
4
bebas dari pengaruh temperatur pada kisaran kedalaman yang lebar (Jokiel dan Coles, 1990) dan lebih lanjut dikemukakan bahwa pemulihan karang mungkin saja terjadi karena dimungkinkannya lebih dari satu takson zooxanthellae dapat menempati inang polip karang (Rowan dan Power; 1991). Pendapat tersebut semuanya mendukung teori translokasi. Veron (1995) menyatakan bahwa temperatur hanya sebagai suatu syarat yang menghantarkan ke proses pemutihan karang, sebagaimana peranannya dalam pengaturan siklus reproduksi.
Ruang Lingkup Penelitian
Dewasa ini kondisi terumbu karang baik yang ditemukan di perairan Indonesia maupun di berbagai lokasi sebarannya telah mengalami degradasi. Berkaitan dengan proses degradasi tersebut, Westmacott, et al (2000) menyatakan bahwa gangguan terbesar bagi terumbu karang baik secara lokal maupun global menimbulkan pertanyaan mengenai masa depan terumbu karang. Berkenaan dengan proses degradasi karang, maka pada dasarnya tekanan yang diterima oleh biota ini pada awalnya merupakan suatu aktivitas pemutusan hubungan fungsional antara zooxanthellae dengan karang. Proses pemutusan hubungan fungsional ini merupakan mekanisme pelepasan zooxanthellae dari jaringan-jaringan polip karang. Mekanisme awalnya dicirikan oleh proses pemutihan karang. Dengan pelepasan zooxanthellae ini maka proses transfer energi beserta dampak fungsional akan terganggu. Dalam tekanan yang bersifat kontinyu dan dalam jangka waktu yang lama maka akan dapat menyebabkan kematian bagi karang.
5
karang yang resisten (Fitt et al., 2001). Kembalinya ekosistem terumbu karang ke fungsi semula setelah degradasi bergantung kepada kesuksesan reproduksi dan rekolonisasi karang-karang yang tersisa serta dari karang-karang yang berada di luar populasi sumber terumbu dalam kondisi kelabilan lingkungan.
Dalam kaitan tersebut, terlaksananya reproduksi dan rekolonisasi merupakan suatu kejadian dari suatu proses kehidupan karang untuk membantu melakukan pemulihan. Reproduksi dan rekolonisasi tidak akan dapat terlaksana dan mempunyai peluang yang sangat kecil dalam kondisi terganggunya faali biota karang (Fitt et al., 2000). Selanjutnya dikemukakan bahwa gangguan faali ini harus terselesaikan terlebih dahulu untuk mendapatkan kondisi karang yang stabil. Stabilitas faali merupakan salah satu faktor penting dalam pemulihan individu karang. Menurut Lenhoff (1974) stabilitas faali tersebut adalah kelengkapan stuktur biokimiawi polip. Selanjutnya dikemukakan bahwa struktur biokimiawi cnidaria mencakup sistem syaraf dan sistem transport nutrien. Keduanya memegang peranan penting dalam proses reproduksi serta adaptasi karang.
Regulasi zooxanthellae berkaitan dengan proses pemulihan organ dalam polip karang. Menurut Stokes (1972) dinyatakan bahwa dalam kondisi eleminasi sebagian organ jaringan mesoglea dan endoderm maka terbentuknya rajutan baru ditentukan oleh sambungan fungsional dari sistem syaraf dan sistem nutrisi karang. Masuknya zooxanthellae ke dalam jaringan ini merupakan pemula untuk dapat terwujudnya jaringan ikat baik syaraf maupun nutrisi dari polip karang. Demikian seterusnya hingga akan mengisi kerusakan-kerusakan organel dalam jaringan mesoglea dan endoderm untuk membentuk struktur organ yang sempurna dari polip karang. Dalam kondisi inilah proses reproduksi dan rekolonisasi akan dapat berlangsung.
6
(2003) bahwa pada saat musim penghujan terumbu karang di paparan terumbu karang Selatan Pulau Panjang hampir memperlihatkan kepunahan, namun demikian pasca musim hujan berlalu terindikasi mempunyai pemulihan dengan struktur komunitas yang sama. Dengan demikian memunculkan pertanyaan apakah coral bleaching hanya semata-mata merupakan suatu efek ataukah suatu strategi biota untuk mempertahankan diri masih menyisakan pertanyaan. Buddemier dan Futin (1983) berpendapat bahwa hal tersebut merupakan kemungkinan yang terjadi di alam, dengan konsepnya bahwa pemutihan karang merupakan mekanisme adaptasi; meskipun pendapat ini ditentang oleh beberapa peneliti lainnya.
Teknik translokasi zooxanthellae antar inang pada ekosistem terumbu karang yang diteliti ini merupakan salah satu upaya untuk memberikan pembuktian terhadap teori adaptasi sebagaimana yang diusulkan oleh Buddemeier dan Futin (1983). Selanjutnya bahwa tantangan ke depan khususnya terhadap meningkatnya perdagangan karang akhir-akhir ini perlu diantisipasi massalisasi produk karang melalui teknik-tektik buatan dengan tanpa menggantungkan sepenuhnya pada sediaan alamiah. Bioteknik translokasi zooxanthellae pada karang diperkirakan dapat memberikan pemecahan masalah tersebut; sehingga nilai keberadaan (existence value) dari karang akan tetap terpelihara secara alamiah; sementara nilai ekonomi (economic value) dapat ditarik berdasarkan pendekatan etik seperti yang dipersyaratkan sebagai produk ekonomik dan lestari dari sumberdaya alam (Primack, 1995). Dengan demikian, kajian translokasi tidak hanya dapat memberikan jawaban mendasar tentang pemulihan terumbu karang, akan tetapi juga merupakan suatu langkah alternatif terhadap bagi upaya-upaya pemanfaatan terumbu karang secara lebih bijaksana.
Pemikiran translokasi zooxanthellae antar inang sebagaimana yang akan dituju dalam penelitian ini, secara konseptual didasarkan kepada tiga aspek, yaitu :
1. Peranan simbiosis antara zooxantellae dan binatang karang.
7
a. Merombak sisa metabolik (Goreau, 1961 dalam Veron, 1995) b. Meningkatkan proses kalsifikasi (Goreau, 1961 dalam Veron, 1995), c. Memberikan konstribusi nutrien secara langsung dan
d. Mendaur ulang (recycling) nutrien pembatas (nitrogen dan fosfor).
Dengan adanya simbiosis, semua kegiatan tersebut atau semua manfaat tersebut akan terlaksana; dan dengan adanya simbiosis ini maka secara fototropis dapat memperpanjang kehidupan karang dalam suatu periode tertentu dan jika tidak untuk jangka waktu tak terbatas memberikan tambahan nutrien minor.
2. Proses Relokasi zooxantellae.
Proses kalsifikasi pada binatang karang mempunyai ketergantungan yang sangat besar terhadap kelimpahan zooxanthellae dalam jaringan tubuhnya. Di lain pihak bahwa proses relokasi zooxanthellae sangat bergantung baik kepada mekanisme transduksi maupun proses respon interselnya. Menurut Lenhoff (1974), bahwa terdapat beberapa tahap dalam mekanisme relokasi zooxanthellae dalam inang binatang karang, yaitu :
a. Kontak dan Pengenalan (Recognition). Meskipun terdapat argumentasi bahwa transduksi zooxanthellae pada jaringan seluler inangnya terjadi pada saat pelepasan planula, namun tahap ini juga dapat terjadi pada setiap perkembangan dari binatang karang. Berkaitan dengan mekanisme kontak ini oleh Borneman (1998) diinformasikan adanya beberapa mekanisme
recognition yang terjadi yaitu melalui 5 cara :
1) Planula Larva. Dalam hal ini keberadaan zooxanthellae pada karang diperkirakan terjadi sejak masih larva. Pada saat planula karang dilepaskan dari induknya, maka pada saat itu telah ditemukan zooxanthellae pada jaringan planula. Kemudian sejalan dengan pertumbuhan karang, maka zooxanthellae melakukan proses regulasi pertumbuhannya dalam jaringan polip karang;
8
mesoglea, selanjutnya ketersediaan nutrien tersebut secara nyata dapat merangsang pertumbuhan zooxanthellae dalam jaringan karang;
3) Inang penyela (intermediet host). Cara ini dijelaskan bahwa masuknya zooxanthellae dapat terjadi melalui pemangsaan eksternal yang dilakukan oleh polip karang. Suatu biota penyela (zooplankton) seperi udang kecil, hasil sisa cernaan zooplankton dan alga yang dimangsa oleh polip karang merupakan komponen penunjang dimungkinkannya transfer zooplankton ke dalam jaringan karang. Zooplankton atau biota penyela ini selanjutnya akan diangkut ke dalam mesentri polip kemudian sisaannya akan tersimpan disana.
4) Feces predator. Ini prinsipnya adalah keluarnya zooxanthallae dari pemangsaan yang tidak sempurna, atau ketidak mampuan beberapa biota untuk mencernanya. Keluar bersama feces yang akhirnya dapat terlarut ke dalam lingkungan perairan.
5) Kontak Acak; yang terjadi melalui terjadinya proses pertemuan secara acak akibat sifat planktonik dari zooxanthellae. Setelah terjadi kontak tersebut, maka terjadinya proses endositosis apabila keduanya mengalami persesuaian.
b. Endocytosis. Merupakan proses pemasukan suatu sel alga ke dalam jaringan inang. Prosesnya dilakukan setelah mengalami tahap pengenalan dengan kecepatan dan jumlah yang bergantung kepada jenis dan kapasitas dari binatang karang.
c. Relokasi intraselluler dari simbion, ini berkaitan dengan sistem endoskeleton dari binatang karang. Proses enzymatik yang membantu pelaksanaannya ditentukan oleh fluktuasi pH seluler.
9
3. Proses Kalsifikasi Binatang Karang.
Menurut Goreau dalam Muscatine (1974) proses kalsifikasi merupakan kombinasi pengaruh eksternal dan internal. Pengaruh faktor eksternal diperlihatkan dalam suplai Ca2+ dan kekurangan CO2 dan perubahan pH ke dalam
jaringan seluler. Adapun pengaruh eksternal adalah adanya kerja enzym hydroksida dan proses transfer nutrien diantara zooxanthellae dan binatang karang. Pengaruh eksternal juga dimungkinkan apabila terdapat fluktuasi tekanan
insitu dimana binatang karang tersebut ditemukan.
Sementara itu dukungan hasil penelitian yang mempunyai keeratan dengan dimungkinkannya proses translokasi zooxanthellae antar inang pada karang didasarkan kepada keterangan yang menyebutkan bahwa zooxanthellae bukan merupakan species endosimbion tunggal. Selanjutnya berkenaan dengan uji DNA terhadap ragam zooxanthellae pada ragam inang sebagaimana dilakukan oleh Rowan dan Powers (1991) diperoleh keterangan bahwa clade zooxanthellae ternyata beragam. Di samping itu diinformasikan pula bahwa Pocillopora damicornis dan Pocillopora meandrina mempunyai dua clade zooxanthellae yang sama. Ini berarti bahwa genotip alga yang sama ditemukan pada inang yang berbeda. Keterangan ini memberikan dukungan bahwa dimungkinkan terjadinya proses translokasi zooxanthellae antar inang.
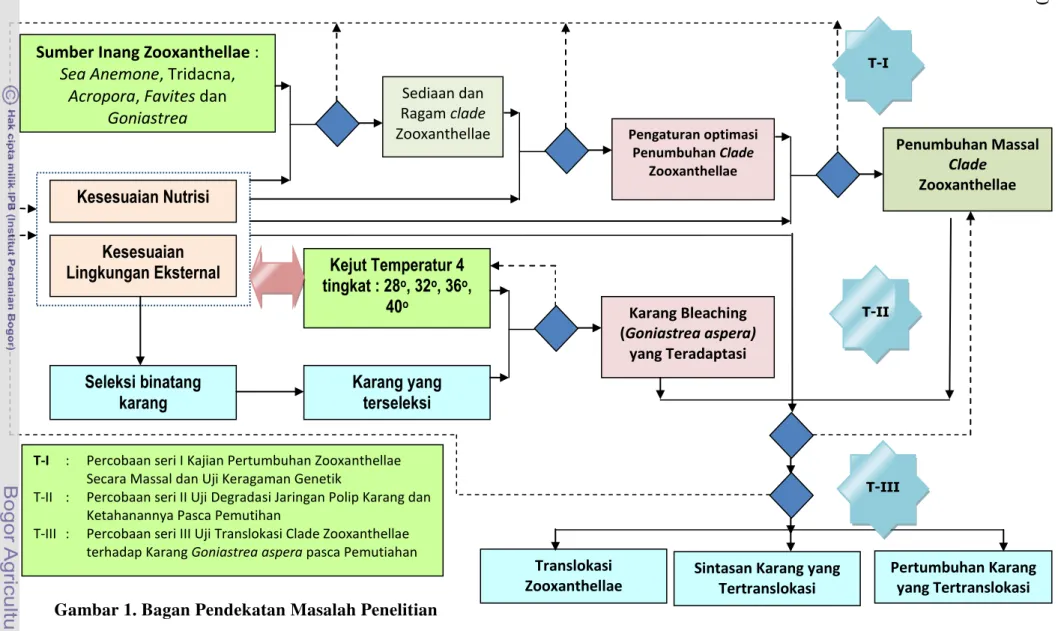
Berkaitan dengan berbagai proses dalam fenomena relokasi zooxanthellae dan peranannya terhadap proses kalsifikasi pada biota karang maka dalam telaahannya diperlukan tahapan kajian sebagai berikut (Gambar 1):
1. Tahap pemurnian zooxanthellae dari beberapa sumber inang, penumbuhan zooxanthellae secara massal dan uji keragaman genetik merupakan tahap penelitian awal yang bertujuan untuk mengetahui seberapa jauh binatang karang dapat bertahan dalam lingkungan binaan dan seberapa jauh berbagai jenis zooxanthellae dapat dipertahankan dalam penumbuhan massal;
10
T-I : Percobaan seri I Kajian Pertumbuhan Zooxanthellae Secara Massal dan Uji Keragaman Genetik
T-II : Percobaan seri II Uji Degradasi Jaringan Polip Karang dan Ketahanannya Pasca Pemutihan
T-III : Percobaan seri III Uji Translokasi Clade Zooxanthellae terhadap Karang Goniastrea aspera pasca Pemutiahan
Gambar 1. Bagan Pendekatan Masalah Penelitian
11
3. Tahap translokasi adalah upaya silang clade dilakukan dengan dua langkah yaitu : Tingkat penjenuhan zooxanthellae, dalam hal ini pengukuran diarahkan kepada dua aspek yaitu kelimpahan zooxanthellae dan profil penempatannya ke dalam jaringan karang hasil adaptasi. Kajian pada tahap penjenuhan karang dilakukan setelah obyek penelitian ini dibleachingkan dengan mempergunakan kejut temperatur. Kedua tahap kalsifikasi merupakan tahap penelitian yang diarahkan untuk menerangkan seberapa jauh proses transfer nutrien antara simbion dan inang dapat berlangsung. Untuk keperluan ini maka akan dilakukan pengukuran pertumbuhan karang pasca penjenuhan zooxanthellae.
Tujuan Penelitian
Penelitian ini bertujuan untuk :
1. Mengevaluasi penumbuhan massal zooxanthellae berdasarkan tahapan pemurnian
clade dari berbagai inang (Sea anemon, Tridacna, Acopora, Favites dan
Goniastrea);
2. Mengevaluasi dan menginventarisir profil zooxanthellae berdasarkan karakter DNAnya;
3. Mengkaji proses bleaching dan mengembangkan batasan tolok ukurnya; 4. Mengevaluasi kemampuan pulih karang pasca pemutihan;
5. Mengevaluasi tahapan translokasi zooxanthellae pada jaringan polip binatang karang;
6. Mengkaji pengaruh translokasi terhadap sintasan dan pertumbuhan karang.
Manfaat Penelitian
Penelitian ini diharapkan memberikan manfaat berupa :
1. Konstribusi pemikiran terhadap upaya penyediaan stock zooxanthellae berbagai
clade dengan sifat bawaan sebagai upaya translokasinya pada beberapa jenis karang baik bagi kepentingan ekonomis maupun konservasi;
12
3. Konstribusi pemikiran terhadap penambahan tolok ukur bagi evaluasi status kesehatan terumbu berdasarkan profil endosimbiosis.
Hipotesis Penelitian
Hipotesis yang diajukan untuk dianalisis dan dibuktikan adalah :
1. Kesesuaian nutrisi dan kondisi lingkungan dapat memelihara pertumbuhan optimal zooxanthellae;
2. Pemutihan parsial dapat mempertahankan pemulihan karang;
3. Translokasi zooxanthellae meningkatkan ketahanan dalam mempertahankan kelangsungan hidup karang;
4. Zooxanthellae yang tertranslokasikan dapat mengalami pengaturan pertumbuhan di jaringan polyp;
5. Tingkat penjenuhan zooxanthellae yang optimal dapat menunjang proses kalsifikasi.
Lokasi Penelitian
TINJAUAN PUSTAKA
Fakta dan Faktor-faktor Destruktif bagi Karang
Tingkat tumpang tindih dampak yang terjadi pada ekosistem terumbu karang semakin meningkat dengan adanya fenomena alam yang menyebabkan pemutihan karang. Istilah pemutihan karang itu sendiri didefinisikan oleh Brown (1997) sebagai pemutihan karang yang diikuti dengan lepasnya symbiotik alga (zooxanthellae) dan atau peluluhan pigmennya. Selanjutnya secara sistemik dipetakan oleh Westmacott, et al., 2000; seperti terlihat pada Gambar 3.
Gambar 3. Penyebaran Global Peristiwa Pemutihan Karang (1998-2000) Sumber : Westmacott et al (2000)
16
Tabel 1. Rangkuman Tentang Identifikasi Pemutihan Karang akibat beberapa Stressor
No Stressor Observasi Penelitian
Lapangan
Observasi Laboratorium
1 Peningkatan temperatur air laut
Glynn, 1993 Hoegh-Guldberg dan Smith, 1989; Glynn dan DCroz, 3 Peningkatan Irradiance Fisk dan Done, 1985; Glesson dan
Wellington, 1993; Brown et al., 1994; Glesson dan Wellington, 1995
Hoegh-Guldberg dan Smith, 1989; Lesser, 1989, Lesser et at., 1990; Lesser dan Shik,
5 Penurunan salinitas Goreau, 1964; Van Woesik et al., 1995; De Vantier et al., in press
Berdasarkan keterangan di atas, nampak bahwa penyebab pemutihan karang dapat bermacam-macam. Namun demikian indikasi terjadinya perubahan pemutihan karang secara global lebih banyak disebabkan oleh pengaruh peningkatan temperatur; meskipun dalam skala lokal indikasi pengaruh irradian yang dalam hal ini terutama pengaruh sinar ultraviolet diuraikan beberapa peneliti juga memberikan andil. Pengaruh pencahayaan terutama direspon oleh fitobiotik yang dalam hal ini zooxanthella dengan cara memacu proses eksitasi elektronik. Mekanisme eksitasi elektronik ini secara rinci dikemukakan oleh Page (1990) bahwa apabila cahaya ini kuat mengenai fitobiotik maka dapat menyebabkan proses luluhnya pigmen, sehingga selanjutnya pigmen tidak mampu melakukan reduksi dalam proses fotosintesis. Ini berarti bahwa secara luas pengaruh temperatur lebih dominan dengan dampak penyebaran yang luas. Meskipun demikian, secara teoritis memang dampak peningkatan irradiance dapat mempengaruhi photosistem I dan II terhadap zooxanthellae. Namun apabila mengacu kepada proses pelepasan zooxanthellae sebagaimana diinformasikan oleh Fang et al., (1998), maka pemutihan karang lebih ditentukan oleh karang dalam merespon stress lingkungan. Dalam hal ini dipicunya katalisis temperatur
dengan meningkatnya produk calmodulin yang selanjutnya memacu
17
dapat terjadi pemecahan sel yang diikuti oleh keluarnya zooxanthellae dari jaringan endoderm karang. Dengan demikian secara substansial efek temperatur lebih dominan dibandingkan efek radiasi matahari. Meskipun hal terakhir dapat menyebabkan peluluhan pigmentasi (chlorophyll c) (Rivers et al., 2002; Bynagle
et al., 2003). Ini dipertegas oleh hasil penelitian Brown et al., (1994) bahwa retraksi jaringan secara nyata dipengaruhi oleh adanya pengaruh temperatur. Akibat antropogenik tidak diinformasikan memberikan pengaruh yang berarti terhadap pemutihan karang sebagaimana dilakukan pengakajian di beberapa wilayah Indonesia (Edinger et al., 1998).
Pada masa yang akan datang dengan semakin meningkatnya berbagai aktivitas hulu terhadap ekosistem karang, diperkirakan akan lebih meningkatkan tekanan terhadap ekosistem ini. Indikasi ini semakin menguat dengan telaah representatif terhadap pola pemanfaatan sumberdaya pesisir yang selama ini berlangsung khususnya di Indonesia dan di Asia Tenggara umumnya (Westmacott
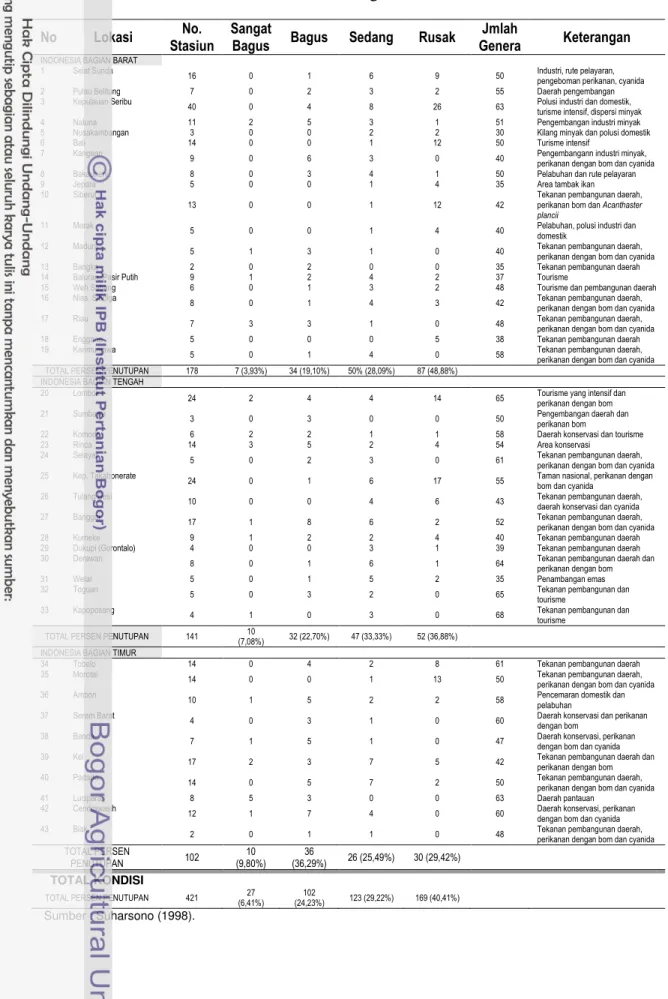
et al., 2000). Kondisi kualitas terumbu karang saat ini di perairan Indonesia juga menambah ke arah indikasi tersebut (Tabel 2; Suharsono, 1998). Kepunahan ekosistem ini sangat merugikan dipandang dari berbagai sudut. Secara ekologis kehilangan ekosistem ini dapat menyumbangkan kerugian yang tidak sedikit mengingat bahwa ekosistem ini adalah salah satu penyumbang terbesar terhadap produksi ikan bersih di kawasan pantai; di samping keindahannya yang juga tidak ditemukan lagi dan yang jauh lebih penting potensi genetik serta berbagai potensi obat-obat penting akan ikut musnah.
Bioekologi Zooxanthellae dan Karang
18
Tabel 2. Kondisi Kesehatan Terumbu Karang di Indonesia
No Lokasi No. TOTAL PERSEN PENUTUPAN 141 10
19
Karang yang ada sekarang mempunyai produktivitas kotor yang sebanding dengan produksi ekosistem terestrial.
Kata simbiosis didefinisikan sebagai hidup bersama dengan organisme yang berbeda. Endosimbiont (organisme yang hidup dalam binatang) secara umum diistilahkan dengan nama zooxanthellae, zoochlorellae dan cyanobacteria, yakni sebagai nama origin dalam dasar taksonomi, tetapi sekarang dideskripsikan sebagai suatu kompleks taksa yang secara luas saling melengkapi dalam taksonomi tanaman atau binatang. Zooxanthellae sendiri merupakan dinoflagellata laut yang ditemukan dalam beberapa invertebrata tropis termasuk karang (Trench, 1979). Dalam perkembangannya, penamaan zooxanthellae berkembang sebagai kumpulan dari beberapa familia dan mungkin klas dari dinoflagellata laut (Blank dan Trench, 1985; Roman dan Power, 1991). Secara formal maupun informal penamaan dari kelompok tersebut sesuai dengan taksonomi alga yang tetap diaplikasikan dalam identifikasinya pada endosimbiont dalam jaringan polip karang. Zooxanthellae secara efektif sekarang diartikan dengan endosymbiotik dari produser primer.
Berdasarkan klasifikasinya maka zooxanthellae dikelompokkan sebagai mikroalga dengan klas dinoflagellata (Borneman, 1998). Dikemukakan lebih lanjut bahwa eksistensi biota ini mulai diketahui sebagai sel yang berada di dalam jaringan karang pada 1881. Awal mula nomenklaturnya dinamakan
Zooxanthellae nutricula. Kemudian dengan berkembangnya teknik identifikasi, jenis ini ditemukan sebagai jenis yang masuk dalam klas dinoflagellata pada tahun 1922. Dalam perkembangannya maka penamaan zooxanthellae berubah yaitu dengan nama Symbiodinium sp; Gymnodinium sp; Symbiodinium microadriaticum
dan Zooxanthellae microadriaticum.
20
aspek tersebut, yaitu : morfologi, struktur tubuhnya, infectivitas, photoadaptif, isoenzym, steropol dan bentuk isoelektrik dari protein peridinium dalam chlorophyll c-nya. Selanjutnya, dengan ditemukannya Symbiodinium pilosum dan
Symbiodinium kawaguti yang ditemukan pada Montipora verrucosa, maka sejak itu ditemukan banyak jenis dari zooxanthellae. Berkenaan dengan hal tersebut, Trench (1979) menginventaris beberapa clade zooxanthellae yang bervariasi, yaitu sebagaimana diinfomasikan pada Tabel 3.
Tabel 3. Beberapa Zooxanthellae yang berasosiasi dengan Cnidaria
No Jenis Zooxanthellae Karang Inang
1 Symbiodinium Sp Oculina divisa
2 Symbiodinium Sp. Millepora dichotoma
3 S. microadriaticum Cassiopeia xamachana
4 S. goreauii Ragactis lucida
5 S. kawaguti Montipora verrucosa
6 S. pilosum Zoanthus sociatus
7 S. meandrinae Meandrina meandrites
8 S. pulchorum Aiptasia pulchellae
9 S. cariborum Condylactus gigantean
10 S. bermudense Aiptasia pallida
11 S. californicum Anthopleua elegantissima
12 Symbiodinium Sp Acropora cervicornis (high ligh)
13 Symbiodinium Sp Acropora cervicornis (low ligh)
14 Gledinium visucum Millepora dichotoma (low ligh) Sumber : Trench (1979)
21
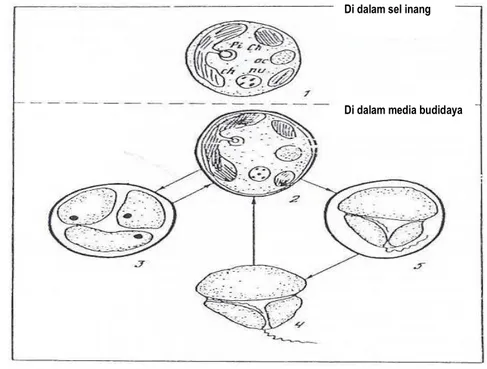
Gambar 4. Skema Siklus Hidup Zooxanthellae
Sumber : Sorokin (1993)
Keterangan :
A. Zooxanthellae dalam jaringan inang 1. Cyst zooxanthellae dalam jaringan inang Pi = Pyrenoid
Ch = Chloroplast Ac = Accumulatif body Nu = Nucleus
B. Zooxanthellae dalam media budidaya 2. Cyst dengan dinding yang keras 3. Zoosporangia
4. Zoospora motil 5. Zoospora immotil
Sejak 1970, semua dinoflagellata simbiotik dianggap sebagai species endemik tunggal Symbiodinium microadriaticum Freudenthal (Taylor, 1974). Dengan berkembangnya ragam penelitian lanjutan yang meliputi perilaku (behavior), fisiologi dan ultrastruktural sebagaimana diinformasikan oleh Baker (2003); La Jeunesse (2001); Santos et al. (2001), secara genetik dijumpai
bermacam jenis Symbiodinium. Perkembangan mengenai ragam clade
zooxanthellae dikemukakan beberapa peneliti sebagai berikut : 1. Clade A, B dan C diperoleh oleh Rowan dan Powers (1991). 2. Clade D diperoleh oleh Carlos et al. (1999).
Di dalam sel inang
22
G
3. Clade E (Symbiodinium californium ditemukan oleh La Jeunesse dan Trench, 2000).
4. Clade F ditemukan oleh La Jeunesse, 2001.
5. Clade G ditemukan oleh Pochon et al. (2001) dan yang terbaru adalah 6. Clade H ditemukan oleh Pochon et al. (2004).
Sementara itu Cofforth dan Santos (2005) menganalisis kekerabatannya berdasarkan rangkuman penelitian yang telah berlalu adalah sebagaimana diinformasikan pada Gambar 5.
Gambar 5. Hubungan phylogenic antar clade utama Symbiodinium.
Sumber : Cofforth dan Santos (2005)
Peranan Faktor Fisika Kimia Perairan terhadap Hubungan Karang dan Zooxanthellae
Faktor-faktor fisika lingkungan mempunyai peranan terhadap zooxanthellae dan inangnya yaitu binatang karang dalam hal pembatasan distribusi dan diversitasnya. Pembahasan mengenai faktor lingkungan ini dipersamakan karena keduanya bersifat sinergis yang artinya pembatasan dan triger keduanya baik untuk pertumbuhan, makan dan sintasannya adalah sama. Ini berarti bahwa distribusi karang yang saat ini terinformasikan berdasarkan telaah dari berbagai aspek yang dikemukakan oleh Veron (1995) semakin menguatkan teori akan sifat sinergisme antar kedua biota tersebut. Atas dasar hal tersebut, saat ini para ilmuwan mempercayai bahwa eksistensi zooxanthellae dapat menerangkan diversitas genetik dari distribusi karang. Smith dan Buddemeier (1992) melaporkan ulasan terbarunya tentang efek perubahan lingkungan terhadap ekosistem terumbu karang. Berikut adalah beberapa telaah variable fisik kimia
23
perairan yang mempunyai peranan penting dalam memperngaruhi distribusi karang beserta endosimbionnya.
Temperatur
Temperatur adalah peubah yang berperan dalam mengendalikan distribusi horizontal dari terumbu karang. Hanya yang memenuhi syarat ini oleh beberapa rujukan yang mungkin adalah efek dari arus dalam pendistribusian dari planula karang (Veron, 1995). Meskipun pernyataan ini pada akhir-akhir ini ditentang, namun hal tersebut tetap menjadi satu dari beberapa pernyataan yang berlangsung terus dalam literatur biologi karang.
Pandangan yang telah bertahan lama dimana saat ini tidak banyak didukung adalah bahwa distribusi terumbu karang dibatasi oleh distribusi karang; dan bahwa hal ini secara umum menyangkut proses fisiologi khususnya dalam hal penangkapan pakan dan reproduksi. Logika alternatifnya adalah bahwa temperatur membatasi keduanya baik terumbu karang dan karang melalui proses interaksi ekologi, dimana kebutuhan energi (cahaya dan hubungan simbiosis) dari terumbu karang secara progresif menjadi kurang kompetitif melawan dominasi makroalga dalam ekosistem. Selang waktu efektif dari temperatur minimum bagi karang dan terumbu karang adalah berbeda. Bagi biota karang, temperatur minimum efektif adalah temperatur yang menyebabkan kematiannya di lingkungan alami. Dalam hal ini terjadi pada habitat karang yang terus menerus atau berulang-ulang selama periode tertentu (dalam jam sampai dengan mingguan) mendapat aliran massa air dengan suhu rendah. Adapun temperatur minimum efektif untuk ekosistem terumbu karang adalah pada suatu kondisi mana konstruksi terumbu tidak dapat dipertahankan. Ini juga berkaitan dengan siklus lingkungan alam, tetapi terjadi dalam rangka waktu yang sangat lama, yakni puluhan bahkan ratusan tahun.
24
Korelasi ini baru-baru ini diuji ulang dalam rentang temperatur dan dicatat di Jepang oleh Veron dan Minchin (1992) dan ditemukan benar. Temperatur rendah sering tercatat di lingkungan terumbu karang, tapi dalam sebagian kasus hanya ditemukan adanya kematian parsial (dimana bagian koloni karang mati) atau meliputi terumbu karang yang secara geologis merupakan hasil peninggalan lampau atau secara primer terdiri dari runtuhan yang tidak terkonsulidasi. Fluktuasi temperatur dalam jangka pendek di terumbu karang Teluk Teluk (Arab dan Parsia) (Coles dan Fadlallah, 1991) diketahui sebagai temperatur minimum.
Semua karang akan mati jika terbuka sampai dengan tidak normal oleh pengaruh temperatur rendah yang abnormal, juga organisme lainnya. Sangat sedikit zooxanthellae karang diketahui dapat mentolerir temperatur di bawah 11oC pada kondisi alamiah. Oulastrea crispata di Semenanjung Noto Laut Jepang dapat mentolerir temperatur pada kira-kira 0oC (Tidak diketahui dalam periode waktunya) dan nampaknya zooxanthellae tetap tinggal (Veron dan Minchin, 1992). Siderastrea radians diketahui tercatat hidup dan toleran pada temperatur 4,5 oC (Veron, 1995).
Sebagian besar informasi dari studi tentang toleransi pada temperatur rendah dilakukan di Selat Tanabe dan dekat Kushimoto Jepang Selatan; dimana species Acropora, Porites, Echinophyllia, Hydnophora dan Leptastrea mengalami tekanan secara fisiologis pada temperatur 9,4oC (Fukuda, 1974 dalam Veron dan Minchin, 1992). Studi ini ketika dikombinasikan dengan peneliti lain (Veron, 1992) tentang distribusi regional, memperlihatkan bahwa terdapat kawasan-kawasan tertentu yang mempunyai perbedaan toleransi species karang terhadap temperatur minimum dan bahwa kesuburan atau kekayaan species di kawasan marginal setidak-tidaknya mendapat setengah tekanan toleransi fungsionalnya dari temperatur yang menerpanya.
Catatan lain dari kelangsungan hidup akibat temperatur rendah ditemukan hampir sama, yang dapat dirangkum sebagaimana uraian berikut :
a. Solenastrea hyades di utara Carolina (Macintyre dan Pilkey, 1969 dalam
25
b. Acropora Sp, Porites Spp dan Platygyra daedalea di Teluk Persia dan Teluk Arab pada temperatur 11oC oleh Coles dan Fadlallah (1991).
c. Montipora Spp di P. Yaeyama pada temperatur kurang dari 13oC (Nomura,
dalam Veron dan Minchin, 1992).
Pada kisaran temperatur serupa juga ditemukan di Florida dan selatan Victoria Australia bahwa karang dengan jenis Plesiastrea versipora dan dua jenis
Coscinarea dapat hidup dengan baik (Davis, 1982; dan Burn, 1985).
Catatan tentang toleransi pada temperatur rendah, pada akhirnya bahwa karang dapat menyebar lebar baik dari segi taksonomi maupun secara geografis. Karang selalu mengalami kematian partial dan secara sekuensial akan mengalami pemulihan. Observasi yang relevan diperoleh bahwa sebagian besar penulis mengemukakan bahwa kematian terbesar terjadi di perairan dangkal, dimana udara dingin terjadi. Hubungan yang sama tepat antara rata-rata temperatur minimum air laut permukaan dan mortalitas dapat dilihat di Kushimoto, dimana temperatur minimum rata-rata bulanan sekitar 15,3oC. Acropora hyacinthus
sebagai species karang dominan di kawasan tersebut menderita meskipun tidak mengalami kematian saat temperatur menurun mencapai 13,7oC pada tahun 1980. Beberapa diantaranya mengalami pemutihan dan sebagian mati pada temperatur 13,4oC kemudian kematian secara luas terjadi pada tahun 1984 saat temperatur mencapai 13,2oC (Misaki, 1980 dalam Veron dan Minchin, 1992). Dalam studi tersebut, temperatur kritis untuk bertahan hidup adalah 2oC di bawah rataan temperatur minimum bulanan. Kematian serupa akan dijumpai oleh species lain, dan studi juga memperlihatkan bahwa variasi kematian yang luas akan terjadi saat lamanya perubahan itu berlangsung.
26
juga terjadi pada lokasi lainnya. Kesimpulan yang dapat diutarakan dari catatan data di Jepang bahwa :
a. 22,5% dari total diversitas species karang dapat mentolerir temperatur minimum 10,4oC.
b. 27% dari total diversitas species karang dapat mentolerir temperatur minimum 13,2oC.
c. 40% dari total diversitas species karang dapat mentolerir temperatur minimum 14,1oC.
Data statistik ini mengabaikan penjelasan tentang frequensi dan lama waktu ditemuinya karang pada variasi temperatur, sehingga mereka secara konservative menunjukkan bahwa setengah dari semua karang mentolerir temperatur 14oC, juga bahwa setengah dari species sisanya mentolerir temperatur 4oC kurang dari temperatur minimum yang tercatat di lingkungan terumbu karang di Jepang. Ini adalah fakta yang kuat yang menunjukkan bahwa pembangunan terumbu tidak dibatasi oleh toleransi karang pada temperatur rendah.
Pada saat hampir mati, sebagian besar pengaruh temperatur rendah pada karang adalah terhadap morfologinya. Beberapa species Acropora pada lintang ekstrim di Jepang berbentuk deretan bertingkat yang melebur tidak teratur dengan plates-nya (tempat lekatan yang mati). Pembatasan pertumbuhan ini nampaknya merupakan bagian dari kematian partial dan pemulihannya. Veron dan Minchin (1992) mencatat bahwa pemulihan jenis tersebut dapat cepat melebihi pemulihan akibat bahaya Acanthaster. Di Tateyama (lokasi karang paling utara dunia) karang juga ditemukan; semua jenis kecuali Alveopora japonica yang tumbuh luar biasa, pertumbuhannya merata dalam bentuk encrusting dan beberapa (seperti
27
subspecies secara geografik, walau satu jenis adalah secara primer ditentukan oleh lingkungan dari pada perbedaan genetik.
Sedikit data yang tersedia tentang laju pertumbuhan karang pada temperatur rendah yang ekstrim. Rata-rata pertumbuhan dari jenis Acropora,
diukur di Kushimoto dan subtropis Kuroshima masing-masing adalah 19 dan 99 mm/th (Veron dan Minchin, 1992). Crossland (1984) menyimpulkan bahwa temperatur rendah merupakan determinan (penentu) primer, sedangkan cahaya merupakan determinan sekunder dari pertumbuhan dan kelangsungan hidup
Acropora formosa di Pulau-pulau Houtman Abrolhos barat. Secara nyata karang-karang di pulau-pulau ini nampaknya tumbuh dengan melimpah dan merupakan tingkat produksi karbonat yang relatif tinggi seperti di terumbu karang tropis (Smith, 1981).
Tekanan temperatur panas tidak seperti tekanan yang dialami oleh karang pada temperatur dingin, yakni bukan merupakan fenomena pembatasan dispersi karang dan juga tidak merupakan pengaruh batas-batas lintang. Secara prinsipil, pengaruh temperatur panas menyebabkan breakdown (kerusakan) simbiosis karang dengan zooxanthellae yang diekspresikan dalam bentuk keluarnya zooxanthellae dari jaringan sel karang atau yang lebih dikenal dengan istilah coral bleaching (pemutihan karang). Meskipun tekanan temperatur tinggi dapat terjadi di daerah terumbu karang lintang tinggi (seperti di Hawaii; Jokiel dan Coles, 1990; di Bermuda, Cook et al., 1990), namun hal itu cenderung terjadi dalam areal yang sempit di kawasan ekuator dan secara umum ini berkaitan dengan saat surut atau surut yang tidak normal. Pada skala biogeografi, tekanan temperatur panas selalu berkorelasi dengan fluktuasi cuaca harian yang mana El Nino Southern Oscillation (ENSO) telah diketahui pengaruhnya yang sangat penting (Guzman dan Cortes, 1992).
1989-28
1990 (Williams dan Bunkley Williams, 1990). Bagaimanapun ENSO berhubungan dengan elevasi temperatur yang mungkin mempunyai dampak nyata terhadap biogeografi hanya di sepanjang timur Pasifik, dimana mereka menerima pengaruh tersebut. Secara langsung ataupun tidak langsung dapat mengakibatkan bleaching karena massa airnya melewati kawasan ini (Lessios et al., 1983; Glynn, 1984) dan mungkin juga menyebabkan kepunahan/kematian massal dari Acropora
di sini. Karena efek dari fluktuasi ENSO sinergis dengan surut yang ekstrim pergerakan massa air menuju sepanjang timur Pasifik, menyebabkan fluktuasinya mungkin berpengaruh terhadap kawasan yang dilaluinya karena adanya halangan oleh Tanah Genting Amerika Tengah, pada akhirnya mereka ini menjadi penting seperti Barrier Pasifik timur dalam memelihara keterisolasian kawasan.
Nilai pembatas temperatur tinggi yang secara ekologis berpengaruh terhadap karang maksimum adalah 30oC - 34oC (Jokiel dan Coles, 1990; Glynn, 1984; Hoegh-Guldberg dan Smith, 1989). Nilai maksimum ini secara geografis dan dengan waktu yang terbuka keterpengaruhannya, dengan toleransi lebih tinggi dari 2 oC dari lokasi tropis atau daerah sedang (temperate)(Coles et al, 1976). Temperatur yang optimum untuk pertumbuhan mempunyai variasi yang lebar dengan ketersediaan nutrien dan mempunyai sifat sinergis dengan pengaruh salinitas dan cahaya (Coles dan Jokiel, 1978) dan optimum pada temperatur 26 oC seperti yang laporkan di karang yang ada di Hawaii (Jokiel dan Coles, 1990).
Musim
29
secara musim di daerah tropis (Chalker et al., 1984). Tetapi kompetisi dengan makroalga merupakan faktor pembatas pertumbuhan pada terumbu karang dekat pantai. Hal ini disebabkan oleh kawasan khususnya pada saat musim hujan akan mengalami masukan massa air dengan beragam polutan pembatas kehidupan karang.
Cahaya
Kemampuan karang untuk membangun terumbu adalah dengan cara memanfaatkan energi dari cahaya matahari. Hal ini menjadi kunci bagi eksistensi pandangan teori terumbu karang yang modern dan juga bisa jadi untuk semua terumbu karang dalam skala geologi. Cahaya, bukan temperatur jelas sekali secara ekologis merupakan pembatas dari pada semua parameter fisika lingkungan, oleh sebab itu nampaknya menunjukkan bahwa cahaya dapat menyebabkan pembatasan secara fisik terhadap biogeografi secara horizon. Kepentingan cahaya, dari kajian biogeografi dan evolusi adalah terkait dengan evolusi dari proses simbiosis karang dengan alga simbionnya yang berperan dalam pembangunan terumbu karang yang melampaui waktu evolusi itu sendiri. Terkait dengan hal tersebut dan dalam peran cahaya, hal ini sinergis dengan adanya sedimentasi lingkungan, dimana pengaruhnya akan dapat menyebabkan hilang/tenggelamnya diversitas secara ekologis.
30
dinophyceae, rhaphidophyceae, chrysophyceae, bacillariophyceae, xanthophyceae dan phaeophyceae (Meek, 1974). Komponen chlorophyl-c mengandung 3 komponen yang secara spectruk berbeda nyata, yaitu chlorophyl-c1, c2 dan c3. Dalam hal ini chlorophyl-c2 selalu berada dalam dinophyceae (zooxanthellae termasuk dalam kelompok ini; baik yang hidup bebas maupun alga simbiotik). Apabila ditelusur lebih lanjut bahwa variabilitas cahaya yang ada sebagaimana dikemukakan oleh Glynn (1993); Gleason (1993) dicirikan oleh kepekatan, dimana kesesuaian lokasi dengan kesesuaian adaptasinya dicirikan oleh semakin pekat, tidak diinformasikan tentang perbedaan warna. Tingkat kepekatan itu dimungkinkan oleh sifat fotoadaptif dari zooxanthellae. Peridinin dan pigmen caretenoid lain dalam sel zooxanthellae merupakan komposisi spesifik dari kelompok dinoflagellata sebagai kompleks protein pigmen. Komposisi ini secara kuat merespon cahaya hijau pada kisaran panjang gelombang 490 - 540 nm.