PENGARUH PEMBERIAN
TEPUNG TAHU DAN TEMPE KEDELAI
DALAM RANSUM TERHADAP
MASSA DAN DENSITAS TULANG
TIKUS BETINA OVARIEKTOMI
Oleh
SAMSU UDAYANA NURDIN
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
SAMSU UDAYANA NURDIN. Pengaruh Pemberian Tepung Tahu dan Tempe Kedelai dalam Ransum terhadap Massa dan Densitas Tulang Tikus Betina Ovariektomi. Dibimbing oleh DEDDY MUCHTADI, ITA DJUWITA, dan SUYANTO PAWIROHARSONO.
Tempe dan tahu dikonsumsi oleh berbagai lapisan masyarakat Indonesia dan telah menjadi sumber zat gizi yang sangat penting. Selain sebagai sumber zat gizi yang baik, tahu dan tempe juga merupakan sumber isoflavon yang penting karena dapat menyediakan 30 hingga 40 mg isoflavon setiap penyajian. Isoflavon diduga mempunyai efek positif terhadap kesehatan tulang. Penelitian ini bertujuan untuk mengetahui pengaruh pemberian tahu dan tempe terhadap massa dan densitas tulang tikus percobaan betina yang diovariektomi. Untuk melihat ada atau tidak adanya efek estrogenik isoflavon dalam tahu dan tempe, maka diamati berat uterus tikus. Evaluasi juga dilakukan terhadap jumlah konsumsi pakan, pertambahan berat badan, dan
efisiensi pakan tikus selama percobaan.
Persiapan pakan meliputi pembuatan tahu dan tempe, tepung tahu dan tepung tempe, dan formulasi pakan. Analisis proksimat dilakukan terhadap kasein, tepung tahu, dan tepung tempe. Khusus untuk tepung tahu dan tepung tempe, dilakukan juga analisis terhadap kandungan isoflavonnya. Data hasil analisis proksimat selanjutnya dijadikan sebagai dasar formulasi ransum berdasarkan persamaan yang
direkomendasikan oleh AOAC. Pakan dibuat isonitrogenus dengan kadar kalsium dan fosfor sama untuk semua jenis pakan. Pakan diberikan secara ad libitum dan diberi rninum air bebas ion selama 32 hari. Tikus yang digunakan adalah tikus betina Sprague-Dawley yang berusia 120 hari, yang terdiri dari 6 ekor tikus normal dan 18 tikus ovariektomi. Dua puluh empat tikus tersebut dibagi ke dalam 3 kelompok berdasarkan berat badan awal. Setiap kelompok terdiri dari 8 ekor tikus, yaitu 2 ekor tikus betina ovariektomi yang diberi pakan dengan sumber protein kasein sebagai kontrol negatif (OvxC), 2 tikus betina ovariektomi yang diberi pakan dengan sumber protein tahu (OvxH), 2 tikus betina ovariektomi yang diberi pakan dengan sumber protein t e m p (OvxT), dan 2 tikus betina normal yang diberi pakan dengan sumber protein kasein (NonOvx) sebagai kontrol positif. Pengamatan dilakukan terhadap massa dan densitas tulang femur sebelah kanan dan tulang lumbar keempat, kadar kalsium serum, dan berat uterus tikus. Selama percobaan juga dievaluasi
perkembangan berat badan dan konsumsi pakan harian.
Tempe memiliki kandungan isoflavon total lebih besar (48,873 mg per 100 g berat kering) jika dibandingkan dengan tahu (39,796 mg per 100 g berat kering). Isoflavon tertinggi pada tempe adalah daidzein dan pada tahu adalah genistein dengan konsentrasi masing-masing 26,237 mg dan 20,862 mg per 100 g berat kering. Faktor I1 pada tahu dan tempe ditemukan sebagai isoflavon dengan kadar terendah, dengan konsentrasi masing-masing 1,483 mg dan 1,439 mg per 100 g berat kering.
dibandingkan dengan berat badan tikus ovariektomi yang mendapat pakan kasein (OvxC), tahu (OvxH) atau t e m p (OvxT) yaitu masing-masing 30,OO g, 41,33 g, 48,33 g, dan 38,73 g. Konsumsi pakan rata-rata tidak berbeda nyata antarperlakuan, dengan rata-rata konsumsi untuk NonOvx, OvxC, OvxH, dan OvxT masing-masing sebesar 9,56 g, 10,91 g, 10,67 g, dan 10,72 g. Efisiensi pakan tempe pada tikus ovariektomi tidak berbeda nyata dengan efisiensi pakan kasein pada tikus normal, yaitu masing-masing sebesar 3,13 dan 3,62, tetapi lebih rendah jika dibandingkan efisiensi pakan kasein dan tahu pada tikus ovariektomi (3,84 dan 4,34). Ovariektomi meningkatkan laju resorpsi tulang yang ditandai dengan meningkatnya kadar kalsium serum (yaitu 2,19 mg/100 ml serum untuk OvxC, 2,5 1 mg/100 ml serum untuk OvxH, dan 2,29 mg/100 ml serum untuk OvxT) dibandingkan tikus normal (1,39 mg/lOO ml serum). Densitas tulang lumbar keempat tikus normal dan tikus ovariektomi yang mendapat pakan tahu tidak berbeda nyata (yaitu masing-masing
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul:
PENGARUH PEMBERIAN
TEPUNG TAHU DAN TEMPE KEDELAI
DALAM RANSUM TERHADAP
MASSA DAN DENSITAS TULANG
TIKUS BETINA OVARIEKTOMI
Adalah benar merupakan hasil karya saya sendiri dan belum pernah dipublikasikan. Semua sumber data dan informasi yang digunakan telah dinyatakan secara jelas dan dapat diperiksa kebenarannya.
Bogor, April 2002
PENGARUH PEMBERIAN
TEPUNG TAHU DAN TEMPE KEDELAI
DALAM RANSUM TERHADAP
MASSA DAN DENSITAS TULANG
TIKUS BETINA OVARIEKTOMI
SAMSU UDAYANA
NURDIN
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Pangan
PROGRAM PASCASARJANA
INSTITUT
PERTANIAN
BOGOR
Judul Tesis : Pengaruh Pemberian Tepung Tahu dan Tempe Kedelai dalam Ransurn terhadap Massa dan Densitas Tulang Tikus Betina Ovariektomi
Nama : Samsu Udayana Nurdin
NRP : 99209
Program Studi : Ilmu Pangan
Menyetujui, 1. Komisi Pembimbing
Prof. DR. Ir. Deddv Muchtadi. MS. Ketua
DR. Drh. l% Diuwita, M.Phil. Anggota I
DR. Suyanto Pawiroharsono, APU Anggota I1
Mengetahui,
2. Ketua Program Studi Ilmu Pangan
yphg-
Prof. DR. Ir. B. Sri Laksmi Jenie. MS.
RIWAYAT HIDUP
Penulis dilahirkan di Pringsewu, Kabupaten Tanggamus Propinsi Lampung
pada tanggal 6 Juni 1967 dari pasangan Bapak Johanis (alm.) dan Ibu Siti Hasanah. Penulis merupakan putra pertarna dari 5 bersaudara dan telah menikah dengan Yeni
Widarsih, SP.
Penulis berhasil menyelesaikan pendidikan sekolah dasar pada tahun 198 1 di
SD Negeri 1 Adiluwih dan pendidikan sekolah menengah pertama pada tahun 1984 di
SMP Negeri 1 Tanjung Karang. Pendidikan sekolah menengah atas dapat penulis
selesaikan pada tahun 1987 di SMA Negeri 1 Tanjung Karang dan pada tahun yang
sama diterima di Fakultas Pertanian Universitas Lampung. Gelar Sarjana Pertanian
(Ir) berhasil diperoleh penulis pada tahun 1992. Sejak tahun 1994 penulis bekerja
sebagai Staf Pengajar di Jurusan Teknologi Hasil Pertanian, Fakultas Pertanian
Universitas Lampung hingga sekarang.
UCAPAN TERIMAKASIH
Puji syukur penulis panjatkan ke hadirat Allah SWT, yang dengan rahmat dan
karunia-Nya sehingga penulis dapat menyelesaikan tesis ini. Tanpa ridho-Nya, tesis
ini tak akan pernah terselesaikan.
Banyak pihak yang berperan sangat besar dalam penyelesaian tesis ini. Berbagai bantuan baik fisik maupun moral telah penulis terima dengan nilai yang tak
terhingga. Karena itu, pada kesempatan ini penulis ingin mengucapkan terima kasih
kepada:
1. Bapak Prof. DR. Ir. Deddy Muchtadi, MS., Ibu DR. Drh. Ita Djuwita, M.Phil., dan
DR. Suyanto Pawiroharsono, APU. yang telah memberikan bimbingan, motivasi,
dan wawasan kepada penulis dalam menyelesaikan tesis ini.
2. Istriku Yeni Widarsih, Mamak dan Mama yang menjadi surnber motivasi dan
semangat. Semoga apa yang penulis lakukan dapat membahagiakan mereka.
3. Bapak (alm.) dan Papa (alm.), semoga Allah S WT melimpahkan pahala kebaikan
yang penulis dapat lakukan pada penelitian ini kepada mereka.
3. Adik-adikku, Iyal, Adi, Hendra, Heni, yang bersabar karena kakaknya tidak dapat berbuat banyak untuk kehidupan mereka.
4. Staf Lab. Embriologi FKH IPB, terutarna Ibu DR. Drh. Ita Djuwita, M.Phil. dan Bapak Drh. M. Kusdiantoro, yang membantu penulis dalam mempersiapkan
hewan percobaan dan pembedahannya.
5. Kawan-kawan terbaikku di IPB, Budi dan Mbak Lince, atas kebersamaannya. Semoga kita tetap menjaga persahabatan kita selamanya.
6. Pak Gatot dan Ibu, Mbak Ida, Pak Toni, Pak Pai, Pak Ahyar, Pak Wahid, dan Adi
yang banyak membantu selama di laboratorium dan di kandang.
7. Fitri dan Ju', semoga data-data yang ada dapat membalas bantuan yang penulis terima.
9. Bu' Elsi, Neti dan kawan-kawan IPN serta semua pihak yang tidak dapat saya sebutkan satu persatu atas bantuannya.
Akhirnya penulis berdoa, semoga semua kebaikan mereka mendapat pahala dari Allah SWT dan karya ini dapat bermanfaat dan memberikan keberkahan kepada semua pihak.
DAFTAR IS1
halaman DAFTAR IS1
...
DAFTAR TABEL
...
DAFTAR GAMBAR...
DAFTAR LAMPIRAN...
I.
PENDAHULUAN...
1.1 Latar Belakang...
...
1.2 Tujuan Penelitian
I1
.
TINJAUAN PUSTAKA...
2.1 Kedelai, Tempe Kedelai, dan Tahu...
2.2 Isoflavon...
...
2.3 Biologi Tulang
...
2.4 Pengaruh Hormon terhadap Kesehatan Tulang
...
2.5 Isoflavon dan Kesehatan Tulang
...
2.6 Tikus Putih sebagai Model untuk Osteoporosis
...
I11
.
BAHAN DAN METODE PENELITIAN...
3.1 Tempat dan Waktu Penelitian
...
3.2 Pakan. Hewan Percobaan. dan Pelaksanaan Percobaan
...
3.3 Pengamatan
...
3.4 Rancangan Penelitian dan Analisis Data
...
IV
.
HASIL DAN PEMBAHASAN4.1 Kandungan Senyawa Isoflavon Tepung Tahu dan Tepung Tempe
...
...
4.2 Formulasi Ransum
4.3 Perkembangan Berat Badan. Jumlah Konsumsi Pakan. dan
...
...
4.4 Berat Uterus dan Berat Uterus per Berat Badan...
4.5 Kadar Kalsium Serum
...
4.6 Massa dan Densitas Tulang Femur...
4.7 Massa dan Densitas Tulang Lumbar Keempat...
4.8 Rekapitulasi Hasil Penelitian.
...
V KESIMPULAN DAN SARAN
5.1 Kesimpulan
...
5.2 Saran
...
DAFTAR PUSTAKA
...
DAFTAR TABEL
Tabel Halaman
[image:107.588.68.507.78.664.2]1
.
Kandungan isoflavon kedelai dan pada berbagai produk olahan kedelai..
2...
2
.
Komposisi zat gizi tempe kedelai per 100 gram berat kering 6...
3
.
Kandungan zat gizi tahu segar per 100 gram berat kering 7 4.
Perkiraan kandungan isoflavon kedelai dan produk olahan kedelai perpenyajian
...
8...
5
.
Hasil analisis kuantitatif isoflavon tepung tahu dan tepung tempe 27 6.
Hasil malisis proksimat. kadar kalsium dan fosfor kasein. tepung tahu....
dan tepung tempe 29
...
7
.
Komposisi bahan untuk pembuatan 1 kg pakan 308
.
Pengaruh perlakuan terhadap pertambahan berat badan dan terhadap per-bandingan pertarnbahan berat badan dengan jumlah konsumsi pakan rata-
rata
...
32 9.
Pengaruh perlakuan terhadap berat uterus dan terhadap perbandingan...
berat uterus dengan berat badan 36
...
10
.
Pengaruh perlakuan terhadap kadar kalsiwn serum 38...
1 1
.
peng& perlakuan terhadap massa dan densitas tulang femur 4012
.
Pengaruh perlakuan terhadap massa dan densitas tulang lumbar...
keempat 41
...
DAFTAR GAMBAR
Gambar Halaman
DAFTAR LAMPIRAN
Lampiran Halaman
...
1
.
Hasil uji sidik ragam rata-rata konsumsi pakan tikus 52...
2
.
Hasil uji sidik ragam pertambahan berat badan tikus 52...
3
.
Hasil uji Beda Nyata Terkecil pertambahan berat badan tikus 52...
4
.
Hasil uji sidik ragam efisiensi pakan tikus 53...
5
.
Hasil uji Beda Nyata Terkecil efisiensi pakan tikus 53...
6
.
Hasil uji sidik ragam berat uterus tikus 537
.
Hasil uji Beda Nyata Terkecil berat uterus tikus...
54...
8
.
Hasil uji sidik ragam berat uterudberat badan tikus 54...
9
.
Hasil uji Beda Nyata Terkecil berat uteruslberat badan tikus 54 10.
Hasil uji sidik ragam kadar kalsium serum tikus...
55...
1 1
.
Hasil uji Beda Nyata Terkecil kadar kalsiurn serum tikus 55 12.
Hasil uji sidik ragam massa tulang femur tikus...
55 13.
Hasil uji sidik ragam densitas tulang femur tikus...
5614
.
Hasil uji sidik ragarn massa tulang lumbar keempat...
5615
.
Hasil uji sidik ragam densitas tulang lumbar keempat tikus...
5616
.
Hasil uji Beda Nyata Terkecil densitas tulang lumbar keempat tikus...
5717
.
Prosedur analisis kandungan isoflavon...
5818
.
Kromatogram standar isoflavon...
601 9
.
Kromatogram isoflavon tepung tahu...
61 20.
Kromatogram isoflavon tepung tempe...
622 1
.
Prosedur analisis kadar air...
63 22.
Prosedur analisis kadar abu...
63...
23
.
Prosedur analisis kadar protein 63...
24
.
Prosedur analisis kadar lemak kasar 65...
25
.
Prosedur analisis kadar serat kasar 65...
27. Prosedur penentuan densitas dan massa tulang femur dan tulang lumbar
I. PENDAHULUAN
1.1 Latar Belakang
Kedelai merupakan bahan pangan penting yang digunakan harnpir di seluruh dunia. Di negara-negara timur kedelai telah digunakan sejak zarnan dulu sebagai sumber gizi, bahan baku berbagai jenis makanan, dan bahan obat. Sementara itu, di negara-negara barat hingga saat ini kedelai sebagian besar hanya dikonsumsi para vegetarian dan mereka yang memahami manfaat kedelai bagi kesehatan.
Bertambahnya laporan-laporan penelitian tentang manfaat kedelai bagi kesehatan, telah meningkatkan permintaan kedelai dari negara-negara barat (Messina, 1999a).
Berbagai produk dapat dihasilkan dari kedelai baik sehagai bahan makanan maupun pakan ternak. Produk kedelai hasil industri tradisional yang terdapat dan berpotensi di pasaran Indonesia antara lain adalah temp, tahu, tauco, kecap, dan kembang tahu.
Tempe dikonsurnsi oleh jutaan masyarakat di Indonesia clan telah menjadi sumber gizi yang sangat penting. Tempe mempunyai kandungan protein yang tinggi dengan nilai
PER
yang hampir setara dengan kasein dan susu skim. Tempemerupakan sumber mineral seperti kalsium, fosfor, besi, serta berbagai vitamin (S yarief et. al., 1999).
sebagai daging talc bertulang, karena tahu merupakan produk nabati yang mutu proteinnya setingkat dengan produk hewani, yaitu daging dan ayam, dengan nilai NPU 65% (Shurtleff dan Aoyagi, 1979).
Selain sebagai sumber gizi yang baik, tempe dan tahu juga merupakan sumber isoflavon yang penting karena &pat menyediakan 30 hingga 40 mg setiap penyajian (Indiana Soybean Board, 1998). Kandungan isoflavon tempe, tahu dan beberapa produk olahan kedelai lain dapat dilihat pada Tabel 1.
Tabel 1. Kandungan isoflavon kedelai dan pada berbagai produk olahan kedelai*
Produk Isoflavon (mgfg)
Tahu Miso Tempe Susu kedelai
Tepung kedelai (fiull fat)
Tepung kedelai (defatted) - -
Konsentrat kedelai (aqueous washed) Konsentrat kedelai (alcohol washed) Isolat kedelai
Kedelai (raw) 1,4
Sumber: Archer Daniels Midland Company, 1999
Keterangan:
*
= Total isoflavon dinyatakan sebagai unit aglikonIsoflavon adalah salah satu senyawa yang termasuk golongan flavonoid dan merupakan bagian terbesar dalam golongan tersebut. Isoflavon yang ditemukan pada Leguminoceae be rjumlah cukup besar yaitu sekitar 0,25%. Sebanyak 99% isoflavon pada kedelai berupa glikosida d m terdiri dari 64% genistin, 23% daidzin, dan 13% glisitin 7-0-P-glikosida (Naim et.al., 1974). Pada umumnya isoflavon terdapat dalam tanaman kacang-kacangan atau Leguminoceae dan tidak terdapat pada
[image:112.588.94.502.281.475.2]3 Beberapa penelitian menunjukkan bahwa isoflavon kedelai dapat mengurangi resiko osteoporosis. Isoflavon memiliki struktur kimia yang sangat mirip dengan hormon estrogen dan obat osteoporosis ipriflavon yang merupakan isoflavon sintetis. Estrogen dan ipriflavon telah diketahui dapat melindungi densitas mineral tulang wanita postmenopouse. Beberapa data menunjukkan bahwa isoflavon tidak hanya menghambat kerusakan tulang tetapi juga menstimulasi pembentukannya (Messina,
1999a).
Ada beberapa perbedaan pada hasil penelitian tentang pengaruh isoflavon dan produk olahan kedelai terhadap kesehatan tulang. Wangen et. al. (2000) mendapatkan bahwa isoflavonper se tidak terbukti mempunyai pengaruh yang menguntungkan pada remodeling tulang wanita pada saat pre- dan postmenopouse. Sementara Alekel (2000) menemukan bahwa isolat protein yang kaya akan isoflavon menghambat kehilangan tulang lumbar spine wanita premenopouse. Penelitian pada hewan percobaan menunjukkan bahwa produk olahan kedelai memberikan pengaruh positif pada kesehatan tulang (Arjmandi, et. al., 1996; Arjmandi et. al., 1998)
Penelitian pengaruh tahu dan t e m p terhadap kesehatan tulang menjadi topik yang menarik, karena penelitian-penelitian yang dilakukan pada hewan percobaan pada umumnya menggunakan isoflavon murni. Tahu dan t e m p selain rnengandung isoflavon, kandungan proteinnya juga mempunyai pengaruh positif terhadap
dari berbagai dokter, merekomendasikan bahwa lebih baik mengkonsumsi produk olahan kedelai yang mengandung isoflavon dibandingkan dengan suplemen isoflavon.
1.2 Tujuan Penelitian
11. TINJAUAN PUSTAKA
2.1 Kedelai, Tempe Kedelai, dan Tahu
Di antara jenis kacang-kacangan, kedelai merupakan sumber protein yang baik. Di sarnping itu, kedelai dapat juga digunakan sebagai sumber lemak,
karbohidrat, vitamin, mineral dan serat (Koswara, 1992). Kedelai merupakan sumber yang baik bagi asam linoleat dan asam a-linolenat (Messina, 1999a).
Tempe kedelai adalah makanan tradisional Indonesia yang berasal dari kedelai yang difermentasi. Fennentasi t e m p terjadi karena aktivitas kapang Rhizopus sp.
pada kedelai rebus sehingga membentuk massa yang padat dan kompak. Menurut Shurtleff dan Aoyagi (1979), tempe yang baik adalah tempe yang berwarna putih, kompak, dan beraroma khas tempe.
Kedelai mengalami berbagai perubahan komposisi selama proses
pembuatannya. Keterlibatan mikroorganisme pada perubahan komposisi selama proses pembuatan tempe terjadi pada saat perendarnan akibat aktivitas bakteri pembentuk asam dan saat fermentasi oleh aktivitas kapang. Sebagai akibat dari perubahan-perubahan tersebut tempe menjadi lebih enak, lebih bergizi, dan lebih mudah dicerna.
Aktivitas enzim protease kapang menyebabkan protein teruai menjadi asam amino bebas yang bersifat lebih mudah dimanfaatkan oleh tubuh. Muata et. al.
6 demikian, jumlah total asam amino tempe relatif sama dengan jumlah total asam
amino kedelai.
Tempe merupakan sumber vitamin yang baik t e r u m a vitamin B. Selain itu, tempe juga merupakan sumber mineral khususnya kalsium, fosfor dan besi. Beberapa
mineral lain juga terdapat dalam tempe walau dalam jumlah kecil, seperti Mg (236
mg1100 g bk), Zn (3,s mg1100 g bk) dan Mn (1,; mg1100 g bk) (Shurtleff dan Aoyagi,
Metode dasar pembuatan tempe di Indonesia meliputi tahap pembersihan biji
kedelai kering, pencucian dan perendaman, perebusan pertama, pengupasan kulit,
perendaman (hidrasi), perebusan kedua, penirisan dan pendinginan, inokulasi,
pembungkusan dan inkubasi (Shurtleff dan Aoyagi, 1979).
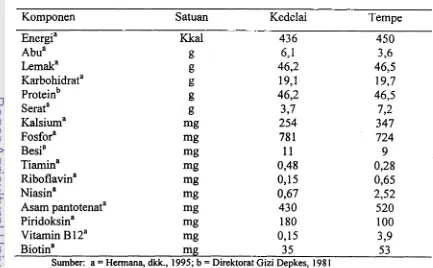
Tabel 2. Komposisi zat gizi tempe kedelai per 100 gram berat kering
Komponen Satuan Kedelai Tempe
Energia Kkal 436 450
Abua g 6,1 3,6
Lemaka g 46,2 46,5
Karbohidrata g 19,l 19,7
proteinb !I 46,2 46,5
Serata g 3,7 7 2
Kalsiuma mg 254 347
Fos for a mg 78 1 724
Besia mg 11 9
Tiamina mg 0,48 0,28
Riboflavina mg 0,15 0,65
Niasina mg 0,67 2,52
Asam pantotenata mi? 430 520
Piridoksina mg 180 100
Vitamin B 1 2a mg 0,15 3,9
Biotina mg 35 5 3
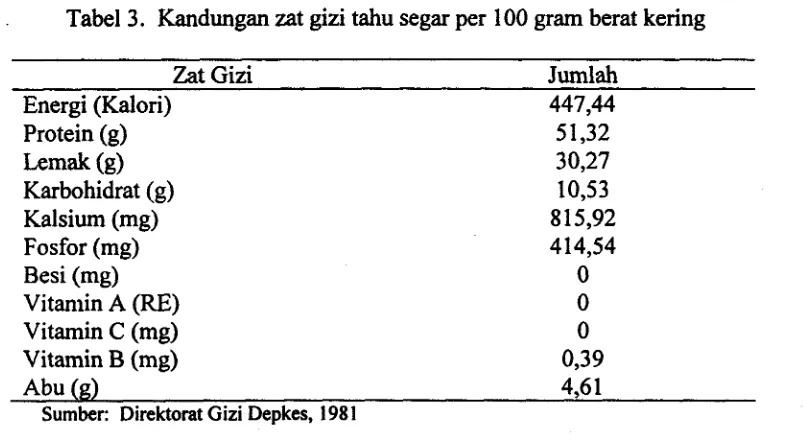
[image:116.588.71.504.387.655.2]Tahu merupakan konsentrat protein kedelai (Shurtleff dan Aoyagi, 1979). Menurut Standar Industri Indonesia (SII, 1990), tahu adalah suatu produk makanan berbentuk padat lunak yang dibuat melalui proses pengolahan kedelai (Glycine sp.) dengan cara pengendapan proteinnya, dengan atau tanpa penarnbahan bahan lain yang diizinkan. Tahu sering disebut sebagai daging tak bertulang, karena tahu merupakan produk nabati yang mutu proteinnya setingkat dengan produk hewani, yaitu daging dan ayam. Tahu merupakan surnber gizi yang lebih baik dibandingkan dengan kedelai biasa, sebab pada tahu enzim inhibitor tripsin telah diinaktivasi, proteinnya terdenaturasi, rasa dan flavornya telah diperbaiki (Sulaeman, Marliyati, dan Anwar,
1992). Komposisi zat gizi setiap 100 gram tahu dapat dilihat pada Tabel 3. Tabel 3. Kandungan zat gizi tahu segar per 100 gram berat kering
Zat Gizi Jumlah
Energi (Kalori) 447,44
Protein (g) IAmak (g) Karbohidrat (g) Kalsium (mg) Fosfor (mg) Besi (mg) Vitamin A (RE)
Vitamin C (mg) Vitamin B (mg)
Abu (g) 4,6 1
Sumber: Direktorat Gizi Depkes, 198 1
[image:117.589.104.507.341.561.2]8
mengandung komponen bukan kedelai. Kecap (soy sauce) dan minyak kedelai (soy oil) tidak mengandung isoflavon (Indiana Soybean Board, 1998). Perkiraan
kandungan isoflavon kedelai dan produk olahan kedelai dapat dilihat pada Tabel 4.
Tabel 4. Perkiraan kandungan isoflavon kedelai dan produk olahan kedelai per penyajian
Ukuran Genistein Total mg isoflavon Jenis makanan penya- (pg/g prot) Isoflavon per penyajian
jian ( ~ g / g pro0
(€9
Mature soybeans, uncooked 93 1106 1891 175,6
Roasted soybeans 86 1214 1942 167,O
Tepung kedelai 21 1185 2084 43,8
Textured soy protein, dry 3 0 472 928 27,8
Green soebeans, uncooked 128 301 548 70,l
Susu kedelai 228 3 0 56 20,O
Tempe mentah 114 277 53 1 60,5
Tahu mentah 144 209 336 38,3
Isolat kedelai kering 28 1100 2174 56,5
Konsentrat kedelai kering 28 1 1 1 195 12,4
Sumber:HCF, 1998
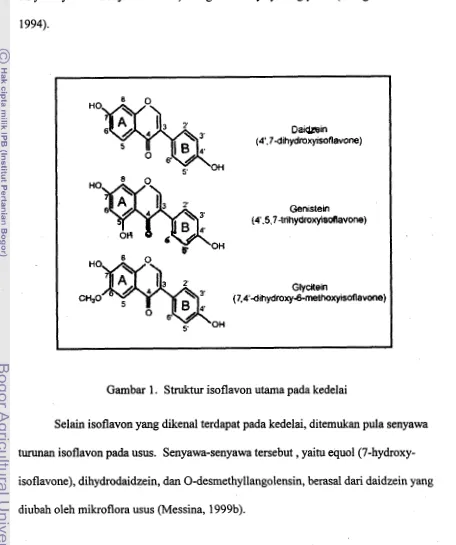
2.2 Isoflavon
Isoflavon merupakan merupakan senyawa terbesar dalam golongan flavonoid. Struktur dasar senyawa flavonoid adalah inti flavon yang terdiri dari 2 cincin benzen (A dan B) yang berikatan melalui cincin karbon pran heterosiklik (Gambar 1). Posisi cincin B benzen merupakan dasar untuk menggolongkan flavonoid menjadi
flavonoid (posisi 2) dan isoflavon (posisi 3).
Isoflavon utarna pada kedelai dan produk olahan kedelai adalah genistein
9 menempel pada posisi C-7 cincin A (Messina, 1999b). Selain itu ditemukan pula
senyawa isoflavon lain dalarn jumlah yang jauh lebih kecil yaitu glycitein (7,4'-
dihydroxy-6-methoxy-isoflavone) dan glikosidanya yaitu glycitin (Wang et. al.,
[image:119.588.66.522.121.666.2]1994).
Gambar 1. Struktur isoflavon utama pada kedelai
Selain isoflavon yang dikenal terdapat pada kedelai, ditemukan pula senyawa turunan isoflavon pada usus. Senyawa-senyawa tersebut
,
yaitu equol(7-hydroxy-isoflavone), dihydrodaidzein, dan 0-desmethyllangolensin, berasal dari daidzein yang
10 Turunan senyawa isoflavon yang ditemukan pada tempe dan tidak ditemukan
pada kedelai adalah Faktor-II(6,7,4' tri-hidroksi isoflavon). Senyawa ini terbentuk
selama proses fermentasi oleh aktivitas mikroorganisme. Faktor-I1 ini terikat dengan
senyawa karbohidrat melalui ikatan glikosidik dan akan dibebaskan pada proses
fermentasi (Pawiroharsono, 2000)
Isoflavon dalarn kedelai secara alarni sebagian besar ada dalam bentuk glikon.
Bentuk ini dipertahankan oleh tanaman sebagai bentuk inaktif hingga dibutuhkan
sebagai antioksidan (Anderson, 1998).
2.3. Biologi Tulang
Tulang tersusun dari sel-sel, matriks organik, dan mineral. Matriks organik
terdiri dari kolagen dan bahan dasar yang mengandung mukopolisakarida. Pada
komponen matriks ini mengendap kristaloid yang terdiri dari kalsium-fosfat
(Djojosoebagio, 1 996).
Sel-sel tulang yang terdiri dari osteosit, osteoblas, dan osteoklas, mempunyai
fungsi khusus dan terletak pada posisi yang berbeda-beda (Djojosoebagio, 1996). Osteoblas dan osteoklas merupakan sel utarna yang berperan dalam pembentukan dan
resorpsi tulang. Osteoblas adalah sel-sel pembentuk tulang yang berasal dari
prekursor sel stroma di sumsum tulang. Sel-sel ini mensekresikan sejumlah besar
kolagen tipe I, protein matriks tulang yang lain, dan alkalin fosfatase. Sel-sel ini berdiferensiasi menjadi osteosit, yaitu sel-sel bundar yang dikelilingi oleh matriks
Garnbar 2. Organisasi sel tulang (Ganong, 1983); Horrnon paratiroid (PTH); Kalsitonin (CT); 1,25-dihidrokolekalsiferol (DHC).
Osteoklas adalah sel multinukleus yang menimbulkan erosi dan menyerap tulang yang sebelumnya telah terbentuk. Sel-sel ini berasal dari stem sel
hematopoietik melalui monosit. Osteoklas melekat ke tulang melalui perluasan membran yang mengelilingi suatu daerah terpisah antara tulang dan bagian osteoklas (Ganong, 1 995).
Mineral tulang merupakan kristal hidroksiapatit (3Ca3(P04)2.Ca(OH)2).
Selain kalsium dan fosfor, tulang juga mengandung sitrat, natrium, barium, strontium, timah, karbonat, fluor, klor, magnesium, kalium, dan air. Karbonat, sitrat, strontium dan natrium terletak di permukaan kristal (Djojosoebagio, 1996).
lapisan-lapisan tulang yang tersusun rapat dan merupakan bagian terbesar dari tulang
panjang, sedangkan tulang trabekular terdiri atas lapisan-lapisan tipis yang
membentuk bagian dalam tulang (Ganong, 1995;
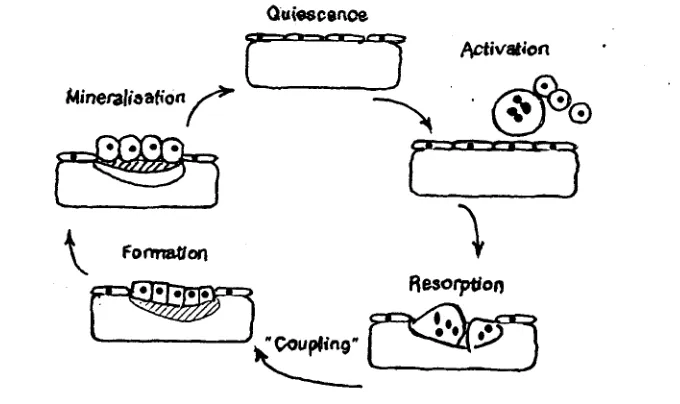
Selama hidup, tulang secara terus menerus diresopsi dan tulang baru dibentuk
kembali. Kalsium dalam tulang mengalami pertukaran dengan kecepatan 100% per tahun pada bayi dan 18% per tahun pada orang dewasa. Remodeling tulang sebagian besar adalah proses lokal yang berlangsung pada daerah-daerah kecil oleh populasi sel
yang disebut unit-unit remodeling tulang (Ganong, 1995).
Ouiescenae
Activation
Gambar 3. Siklus remodeling tulang (Gowen, 199 1 ).
Remodeling tulang merupakan proses fisiologis normal atau sebagai respon
[image:122.584.130.470.309.511.2]13
Tulang d i m (quiescence bone) dilapisi oleh satu lapis sel yang terlihat inaktif yang berasal dari osteoblas. Tahap pertama dari siklus remodeling tulang adalah penempatan prekursor pada tempat tertentu, proliferasi, diferensiasi dan fusi prekursor tersebut ke dalam osteoklas (tahap aktivasi). Tahap selanjutnya adalah resorpsi, yaitu tahap dimana osteoid dan matriks yang telah mengalami mineralisasi dirusak oleh aktivitas osteoklas. Sebelum memasuki tahap pembentukan (formation), terdapat tahapan inaktif, dimana osteoklas hilang dan permukaan tulang yang rusak tersebut tidak dilapisi oleh sel selain kadang-kadang terdapat mononuclear
phagocytes. Mononuclear phagocytes ini mungkin terlibat di dalam penempatan prekursor osteoblas, yang selanjutnya akan mengalami proliferasi dan diferensiasi pada lokasi yang sedang mengalami remodeling. Sel matang kemudian membentuk matriks tulang baru hingga kerusakan tergantikan (formation). Osteoid selanjutnya mengalami mineralisasi dan lapisan sel terlihat kembali (quiescence) (Gowen, 1991)
2.4 Pengaruh Hormon terhadap Kesehatan Tulang
Remodeling tulang dipengaruhi oleh berbagai hormbn dan faktor
pertumbuhan. Terdapat tiga hormon utama yang paling berpengaruh, dan ketiganya berkaitan dengan metabolisme kalsium. Hormon paratiroid (PTH) meningkatkan ~ a ~ + ~ l a s m a dengan memobilisasinya dari tulang. Hormon ini meningkatkan reabsorpsi ca2+di dalam ginjal, yang diimbangi dengan peningkatan ca2+ yang difiltrasi. Paratiroid juga meningkatkan pembentukan 1,25-dihidrokolekalsiferol
usus, memobilisasi ion dari tulang dan meningkatkan reabsorpsi ca2+ dalam ginjal. Sementara itu, hormon kalsitonin (CT) menghambat resorpsi tulang dan
meningkatkan jumlah ca2+ dalam plasma (Ganong, 1995).
Selain ketiga hormon utarna di atas, beberapa hormon dan faktor perturnbuhan juga dapat mempengaruhi pembentukan tulang dan metabolisme kalsium. Glukokor-
tikoid merendahkan kadar ca2+ plasma dengan menghambat pembentukan dan aktivitas osteoklas, tetapi pada jangka panjang, hormon ini dapat menyebabkan osteoporosis dengan m e n d a n pembentukan tulang dan meningkatkan resorpsi tulang. Glukokortikoid m e n d pembentukan tulang dengan menghambat replikasi sel dan sintesis protein di dalam tulang serta menghambat fungsi osteoblas. Glukokortikoid juga m e n d a n absorpsi ca2+ dan PO?- dari usus oleh kerja anti vitamin D clan meningkatkan ekskresi ion-ion tersebut dari ginjal. Penurunan
konsentrasl ca2+ plasma meningkatkan sekresi hormon paratiroid dan resorpsi tulang pun difasilitasi. Hormon pertumbuhan meningkatkan ekskresi kalsium di dalam urin, tetapi juga meningkatkan absorpsi kalsium usus yang dapat lebih besar dari efek ekskresi sehingga menghasilkan keseimbangan positif. Di samping itu IGF-I (insulin like growth factor I) yang dibentuk oleh kerja hormon pertumbuhan merangsang sintesis protein di dalam tulang. Estrogen mencegah osteoporosis dengan efek langsung atau tidak langsung terhadap osteoblas, sedangkan insulin dapat meningkatkan pembentukan tulang (Ganong, 1995).
terhadap berbagai hormon yang secara langsung mempengaruhi kesehatan tulang. Anggapan ini berubah setelah ditemukannya reseptor estrogen beta pada tulang.
Estrogen adalah keluarga hormon yang disintesis dalam jaringan ovarium dan ekstra ovarium. 17P-Estradiol adalah estrogen primer berasal dari ovarium. Pada beberapa spesies, estron lebih banyak tetapi berasal dari jaringan ekstra ovarium. Estrogen dibentuk dengan aromatisasi androgen dalam proses kompleks yang memerlukan 3 langkah hidroksilasi, yang masing-masing membutuhkan Oz dan NADPH. Kompleks enzim aromatase diduga meliputi P-450 oksidase yang mempunyai h g s i ganda. Estradiol dibentuk jika subtrat kompleks enzim ini testoteron, sedang estron berasal dari aromatisasi androstenedion (Granner, 1990).
Fungsi utama hormon ovarium adalah mempersiapkan determinan (penentu) struktural sistem reproduksi wanita untuk reproduksi dengan mematangkan sel benih primordial, mengembangkan jaringan yang menerima implantasi blastosit,
melengkapi hormonal timing untuk ovulasi, menetapkan, dengan hormon plasenta, lingkungan yang diperlukan untuk mempertahankan kehamilan, dan melengkapi pengaruh hormonal untuk persalinan dan laktasi (Granner, 1990).
Estrogen merangsang perkembangan jaringan yang terlibat dalam reproduksi. Umwnnya hormon ini merangsang ukuran dan jumlah sel dengan meningkatkan kecepatan sintesis protein, mRNA, rRNA, tRNA, dan DNA. Di bawah perangsangan estrogen, epitel vagina berproliferasi dan berdiferensiasi; endometrium uterus
16 dengan mempengaruhi pembuluh darah tepi, estrogen khas menyebabkan vasodilatasi dan pembuangan panas (Granner, 1990).
Estradiol juga mempunyai efek anabolik pada tulang dan tulang rawan, sehingga menambah pertumbuhan tulang (Granner, 1 990). Estrogen dapat menstimulasi sel tulang untuk menghasilkan IGF-1. Selanjutnya IGF-1 akan
menstimulasi proliferasi dan produksi kolagen tipe 1 oleh osteoblas (Gowen, 1991). Percobaan menggunakan tikus menunjukkan bahwa estrogen dapat
meningkatkan absorbsi kalsium dari pakan, meningkatkan deposisi kalsium di dalam tulang, clan menurunkan ekskresi kalsium dari tubuh. Sedangkan pada burung dara, estrogen mampu meningkatkan kadar fosfat dalam serum (Djojosoebagio, 1996). Estrogen dapat berpengaruh langsung terhadap kesehatan tulang melalui reseptor estrogen beta. Estrogen dapat menekan produksi IL-6 oleh osteoblas, sehingga menekan produksi osteoklas (Girasole et. al., 1 992).
2.5 Isoflavon dan Kesehatan Tulang
Kemiripan struktur antara isoflavon dan estrogen dan penemuan bahwa estrogen menunjukkan sifat estrogenik yang lemah merupakan awal timbulnya spekulasi bahwa isoflavon dapat meningkatkan kesehatan tulang. Kemunculan spekulasi ini juga didorong oleh kemiripan isoflavon kedelai dengan isoflavon sintetik, 7-isoprop-oxyisoflavone (ipriflavone), yang telah terbukti dapat meningkatkan masa tulang wanita pascamenopouse (Messina, 1999a).
alfa, isoflavon lebih bersifat agonis karena senyawa ini dapat memelihara jaringan tulang atau minimal dapat menjaga keseimbangan antara aktivitas osteoblastik dan osteoklastik. Pada jaringan yang banyak mengandung reseptor estrogen alfa, seperti pada jaringan reproduksi, isoflavon bersifat sebagai antagonist. Pada jaringan ini isoflavon menempati reseptor estrogen dan memblok stimulasi DNA (genome) dan sintesis protein oleh estradiol (Anderson, 1998).
Anderson et a1.(1995) melaporkan bahwa genistein memberikan efek berbeda terhadap tulang pada dua model tikus percobaan steril yang berbeda, yaitu tikus muda yang sedang tumbuh dan tikus menyusui, yang diberi diet rendah kalsium. Setelah 2 minggu percobaan pada tikus muda dan 5 minggu percobaan pada tikus menyusui, genistein pada dosis 1,O mg/hari memiliki pengaruh yang sama dengan conjugated equine estrogen pada dosis Spglhari.
A rjmandi et. al. (1 996) mempelajari pengaruh protein kedelai terhadap kehilangan tulang akibat strerilisasi (ovariectomy). Penelitian yang menggunakan tikus Sprague-Dawley tersebut menghasilkan kesimpulan bahwa protein kedelai lebih bersifat protektif terhadap tulang trabekular bila dibandingkan dengan tulang kortikal. Pada penelitian lanjutan Arjmandi et. al. (1 998) yang menggunakan model yang sama dengan penelitian Arjmandi et. al. (1996) diperoleh kesimpulan lebih jelas bahwa isoflavon yang bertanggung jawab terhadap pengaruh positif terhadap tulang.
18 meningkat secara nyata bila dibandingkan dengan nilai baseline pada wanita
postrnenopouse yang mengkonsumsi 40 gram protein kedelai yang mengandung 2,25
mg isoflavonlg protein, sementara itu pada wanita yang mengkonsumsi 40 g protein
yang mengandung isoflavon lebih rendah (1,39 mglg protein) densitas tulangnya
tetap. Wangen et.a. l(2000) mendapatkan bahwa isoflavon per se tidak terbukti
mempunyai pengaruh yang menguntungkan pada pergantian tulang wanita pada saat
pre dan postmenopouse. Alekel et. al. (2000) melaporkan bahwa isolat protein yang kaya akan isoflavon dapat menghambat kehilangan tulang lumbar spine wanita
premenopouse. Studi epidemilogis yang dilakukan di Jepang menunjukkan bahwa
komsumsi yang tinggi terhadap produk olahan kedelai ternyata dapat meningkatkan
masa tulang wanita pada saat postmenopouse (Somekawa et. al,. 200 1).
2.6 Tikus Putih sebagai Hewan Model untuk Osteoporosis
Osteoporosis pada manusia merupakan penyakit kronis yang datang secara
perlahan-lahan yang belum sepenuhnya dipahami. Dengan alasan ini maka tidak ada
satupun hewan model yang sempurna untuk penyakit ini. Data-data dari percobaan
yang telah menghasilkan obat clan terapi baru untuk penyakit-penyakit tulang
menunjukkan bahwa model yang sempurna untuk osteoporosis pada manusia tidaklah
diperlukan. Dapat diprediksi, responnya cepat, sensitif, dan murah merupakan model
yang bagus dan praktis yang dibutuhkan (Tuukkanen, 2001).
Tikus yang diovariektomi biasanya merupakan model pilihan, tetapi hams
pada jangka waktu penelitian yang lama. Walaupun hewan model tidak mengalami patah tulang, kemungkinan untuk melakukan uji biokimiawi dengan tulang hewan dapat menjadi cara yang akurat untuk menganalisa kerusakan yang dapat
mengakibatkan patah tulang (Tuukkanen, 200 1).
Terdapat beberapa galur atau varietas tikus yang memiliki kekhususan tertentu. Galur Sprague-Dawley memiliki kekhususan berwarna albino putih, berkepala kecil dan ekornya lebih panjang dari badannya. Galur Wistar ditandai dengan kepala besar dan ekor yang lebih pendek, serta galur Long-Evans yang lebih kecil dari tikus putih dan memiliki warna hitam pada kepala dan tubuh bagian depan (Malole dan Pramono, 1 989).
Tikus dapat dikandang bersarna dalam satu kelompok besar yang terdiri dari jantan dan betina dari berbagai tingkat tanpa terjadinya perkelahian yang berarti.
Tikus betina mengalami kematangan seksual pada usia 50 hingga 72 hari (Bennett dan Vickery, 1970). Tikus dapat hidup lebih dari 3 tahun dan produktif untuk berbiak selama lebih dari sembilan bulan atau sampai usia satu tahun. Tikus sudah tidak produktif lagi pada usia 450-500 hari.
Tikus betina sebagai model telah banyak digunakan untuk penelitian
osteoporosis. Pada penelitian ini tikus dibuat kekurangan hormon estrogen dengan cara membuang ovariumnya (ovariectomy). Varietas yang umum digunakan adalah Sprague-Dawley (Anderson et. al., 1 995; Arjmandi et. al., 1 996; Arjmandi et. al,.
111. BAHAN DAN METODE PENELITIAN
3.1 Tempat dan Waktu Penelitian
Penelitian ini dilakukan di Laboratorium Biokimia Pangan dan Gizi Jurusan Teknologi Pangan dan Gizi, Fakultas Teknologi Pertanian; kandang hewan percobaan Jurusan Teknologi Pangan dan Gizi, Fakultas Teknologi Pertanian; Pusat
Pengembangan Teknologi Pangan IPB; Laboratorium Teknologi Bioindustri, Pusat Pengkajian dan Penerapan Teknologi Bioindustri (P3TB) Badan Pengkajian dan Penerapan Teknologi (BPPT) di kawasan Pusat Pengembangan Ilmu dan Teknologi Serpong. Penelitian dilaksanakan mulai bulan Juli 2001 sampai Januari 2002. 3.2. Pakan, Hewan Percobaan, dan Pelaksanaan Percobaan
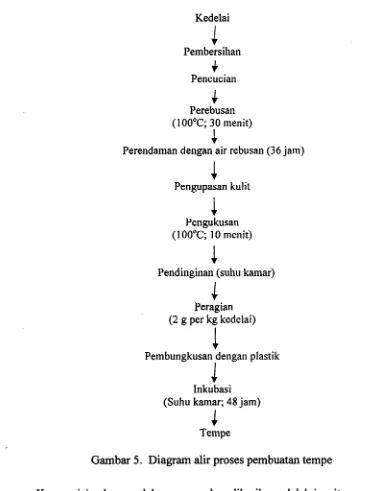
Persiapan pakan meliputi pembuatan tahu dan tempe, tepung tahu dan tepung tempe, dan pencampuran pakan. Diagram proses pembuatan tempe dan tahu masing- masing dapat dilihat pada Gambar 4 dan 5. Kedelai yang digunakan adalah impor dari Amerika yang diperoleh dari agen kedelai di daerah Cilendek Bogor. Ragi yang digunakan adalah ragi Hanoman (BPPT)
.
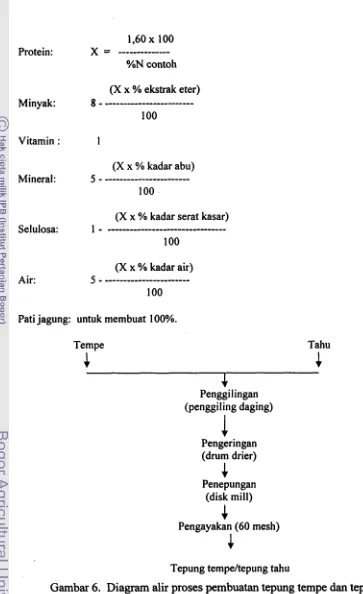
Bahan penggumpal yang digunakan dalam pembutan tahu adalah CaS04. Tahu dan tempe yang diperoleh selanjutnyadikeringkan dengan drum drier (R Simon Dryers Ltd., England) dan digiling menjadi tepung. Diagram proses pembuatan tepung tahu dan tempe dapat dilihat pada
Gambar 6. Tepung yang dihasilkan digunakan sebagai bahan pakan tikus pada pelaksanaan penelitian
2 1 tepung tahu dan tepung tempe sebagai dasar perhitungan dalam formulasi pakan
tikus. Selain itu dianalisis pula kandungan kalsium dan fosfor (Lampiran 26) tepung tahu dan tepung tempe.
Kedelai bersih
Perendaman (1 2 jam)
I
+
PenggilinganI
+
Penambahan air (5 bagian air)Perebusan
(1 00'-1 10°C; 0,7 kg/cm)
Penyaringan
A
Susu kedelai Ampas
Curd Whey
Pengepresan din Pencetakan
Pendinginan dalam air (20 menit)
1
Tahu
[image:131.584.205.491.131.600.2]22
Selama menunggu waktu analisis dan pemberian pakan, tepung tahu dan tepung tempe dikemas dalam kantung plastik polietilen, kemudian disimpan dalam lemari es. Kedelai Pembersihan
+
Pencucian4
Perebusan (1 00°C; 30 menit)4
Perendaman dengan air rebusan (36 jam)
Pengupasan kulit
Pengukusan (1 00°C; 10 menit)
Pendinginan (suhu kamar)
Peragian (2 g per kg kedelai)
Pembungkusan dengall plastik
(Suhu kamar; 48 jam)
4
Tempe
Gambar 5. Diagram alir proses pembuatan tempe
[image:132.588.81.460.126.617.2]1,60 x 100 Protein:
x
=---
%N contoh
(X x % ekstrak eter) Minyak: g
- -- --
---
---
100
Vitamin : 1
(X x % kadar abu) Mineral: 5
-
---
---
---
100
(X x % kadar serat kasar) Selulosa: 1
-
...
100
(X x % kadar air) Air: 5
- --- --- -- ---- --- -- --
100
Pati jagung: untuk membuat 100%.
Tempe
4
Tahu4
4
Penggi lingan (penggiling daging)
Pengeringan
(drum drier)
4
Penepungan (disk mill)
4
Pengayakan (60 mesh)
I
[image:133.586.67.430.53.647.2]Tepung tempeltepung tahu
Semua bahan pakan diaduk hingga homogen dan diukur kadar airnya untuk
mengetahui berat kering pakan yang diberikan. Kadar air sisa pakan juga diukur
untuk mengetahui berat kering pakan yang tersisa. Berat kering pakan yang dimakan
tikus adalah selisih antara berat kering pakan yang diberikan dengan berat kering
pakan sisa. Kalsium dan fosfor pada pakan yang menggunakan tepung tahu dan
tepung tempe diatur sama dengan pakan kasein menggunakan kalsium karbonat atau
potasium fosfat monobasik.
Pemberian pakan perlakuan dilakukan selama 32 hari. Tikus diberi pakan secara ad libitum dan diberi minum air bebas ion. Kandang yang digunakan adalah
kandang plastik yang diberi lubang pada bagian bawahnya.
Ovariektomi tikus dilakukan berdasarkan prosedur Hogan et. al. (1986)
(Lampiran 28). Tikus yang akan diovariektomi dibius dengan ketamin (Ketavet)
dengan dosis lmY200 gram BB. Ovariektomi tikus dilakukan pada saat tikus berusia 90 hari. Kemudian dilakukan pemulihan hingga jahitan siap dilepas (20 hari).
Sebelum percobaan dimulai, tikus normal dan yang diovariektomi diadaptasikan di
lingkungan laboratorium percobaan selama 10 hari.
Percobaan menggunakan 24 (dua puluh empat) tikus betina Sprague-Dawley
yang berusia 17 minggu. Tikus-tikus tersebut dibagi menjadi 3 kelompok berdasarkan berat badan awal. Setiap kelompok terdiri dari 8 ekor tikus, yaitu 2 ekor tikus betina steril yang akan diberi ransum dengan sumber protein kasein sebagai kontrol negatif
tepung tempe (OvxT), dan 2 tikus betina normal yang akan diberi ransum dengan
sumber protein kasein sebagai kontrol positif (NonOvx).
Setelah 32 hari percobaan, tikus dibius dan dimatikan
,
kemudian darah dari leher ditampung untuk diambil serumnya. Serum dari setiap tikus ditempatkan kedalam botol kecil dan disimpan pada suhu lemari es hingga waktu analisis. Tulang yang dianalisis adalah tulang paha sebelah kanan (right femur) dan tulang lumbar
keempat Wurth lumbar bones) yang dikeluarkan dari tubuh tikus melalui operasi.
3.3. Pengamatan
Untuk mengamati pengaruh perlakuan terhadap tulang tikus percobaan maka
diamati massa dan densitas tulang (Lampiran 27) femur sebelah kanan dan tulang
lumbar keempat, serta kadar kalsium (Lampiran 26) serum. Data lain yang juga diukur
adalah berat tikus awal dan setelah percobaan, serta jumlah ransum yang dikonsumsi. Untuk melihat efek estrogenik dari isoflavon, maka diamati juga berat uterus tikus
setelah selesai percobaan.
3.4. Rancangan Penelitian dan Analisis Data
Rancangan percobaan yang digunakan pada penelitian ini adaiah Rancangan Acak Kelompok Lengkap dengan perlakuan tikus betina ovariektomi yang diberi
ransum dengan surnber protein kasein (OvxC), tikus betina ovariektomi yang diberi
ransum dengan sumber protein tepung tahu (OvxH), tikus betina ovariektomi yang
diberi ransum dengan sumber protein tepung tempe (OvxT), dan tikus betina normal
26
Masing-masing perlakuan diulang sebanyak 6 kali. Data yang diperoleh kemudian diolah dengan sidik ragam untuk mengetahui pengaruh perlakuan terhadap parameter yang diuji. Untuk mengetahui perbedaan antarperlakuan, data yang
IV. HASIL DAN PEMBAHASAN
4.1 Kandungan Senyawa Isoflavon Tepung Tahu dan Tepung Tempe
Hasil analisis tepung tahu dan tepung tempe menunjukkan adanya empat jenis senyawa isoflavon yaitu, faktor-2, daidzein, glisitein, dan genistein (Tabel 5).
Kromatogram HPLC isoflavon tepung tempe dan tepung tahu dapat dilihat pada Lampiran 23 dan 24.
Tabel 5. Hasil analisis kuantitatif isoflavon tepung tahu dan tepung tempe Komponen Tepung tahu Tepung t e m p Tepung tahu Tepung tempe
(mg/100g berat basah) (mg/100g berat kering)
Faktor I1 1,327 1,275 1,483 1,439
Daidzein 15,315 23,259 17,121 26,23 7
Glycitein 0,295 0,132 0,330 0,148
Genistein 1 8,66 1 1 8,66 1 20,862 2 1,050
Total 35,598 43,237 39,796 48,873
Berdasarkan Tabel 5, terlihat bahwa senyawa isoflavon yang dominan pada tepung tahu dan tepung tempe adalah daidzein dan genistein. Tepung tempe
mengandung daidzein dan genistein lebih tinggi bila dibandingkan dengan tahu. Hal ini diduga karena pada proses pembuatan tempe terdapat tahapan yang lebih
memungkinkan te rjadinya perubahan senyawa isoflavon dari bentuk glikosida menjadi aglikon.
Isoflavon utama pada kedelai adalah genistein (4',5',7-tryhydroxy-isoflavone)
[image:137.584.81.503.267.420.2]28 daidzein dan 15% glicitein. Secara alami, isoflavon pada kedelai hampir seluruhnya ada dalam bentuk P-glikosida (glikon) (Anderson, 1998). Proses perendaman kedelai pada pembuatan tahu dan tempe, serta proses pencucian pada pembuatan t e m p diduga mampu mengaktifkan enzim P-glukosidase pada kedelai. Enzim ini dapat menghidrolisis glikosida isoflavon menjadi aglikon isoflavon (Ewan, et.al., 1992). Tahap perendaman pada proses pembuatan tempe kurang berpengaruh pada
perubahan bentuk isoflavon bila dibandingkan dengan tahap fermentasi ini. Tahap fermentasi pada proses pembuatan kedelai merupakan tahapan penting yang dapat mengubah glikosida isoflavon menjadi aglikon glikosida. Fungi Rhizopus oligosporus
(Siregar dan Pawiroharsono, 1 997; Pawiroharsono, 1997) dan Rhizopus oryzae
(Siregar dan Pawiroharsono, 1997) serta beberapa jenis kamir dan bakteri kontaminan (Pawiroharsono, 1997) yang terlibat dalam proses fermentasi mampu menghasilkan enzim P-glukosidase yang juga dapat menghidrolisis glikosida isoflavon.
Peningkatan konsentrasi aglikon isoflavon pada proses fermentasi lebih besar jika dibandingkan dengan pada proses perendaman (Pawiroharsono, 1 997).
Isoflavon dalam bentuk glikosida (glikon) bersifat tidak aktif (Pawiroharsono, 1997). Proses hidrolisis yang menyebabkan terputusnya ikatan P-glukosida
mengaktifkan senyawa tersebut. Selain itu, bentuk aglikon mempunyai
bioavailabilitas yang lebih baik jika dibandingkan dengan bentuk glikon (King et.al.,
29 4.2 Formulasi Ransum
Pada Tabel 6 disajikan hasil analisis proksimat, kadar kalsium, dan kadar fosfor kasein, tepung tahu dan tepung tempe. Data hasil analisis ini selanjutnya dijadikan dasar perhitungan formulasi ransum tikus yang digunakan dalam
percobaan. Persamaan yang digunakan dalam perhitungan berdasarkan rumus yang disarankan AOAC (1990). Komposisi bahan untuk pembuatan 1 kg ransum dapat dilihat pada Tabel 7.
Tabel 6. Hasil analisis proksimat, kadar kalsium dan fosfor kasein, tepung tahu dan tepung tempe
Kasein Tepung tahu Tepung t e m p
Komposisi (% berat basah)
Air Protein Lemak Serat kasar Abu Kalsium Fosfor
[image:139.588.64.504.303.499.2]akibat penelitian ini diharapkan benar-benar karena perbedaan surnber protein dan kadar isoflavon pakan.
Tabel 7. Komposisi bahan untuk pembuatan 1 kg pakan
Komposisi Perlakuan
NonOvx OvxC OvxH OvxT
Kasein (g) 109,53 109,53
-
-
Tepung tabu (g)
-
-
230,OOTepung tempe (g)
-
-
269,59Minyak jagung (g) 79,80 79,80 32,50 33,70
Selulosa teknis (g) 20 20 8,35 2,12
Mineral mix* (g) 11,31 11,31 9,40 8,71
c a c o 3 (12) 37,56 37,56 21,99 28,69
w 2 p o 4 (g) 19,15 19,15 16,66 18,42
Vitamin (Bekamin) (g) 10 10 10 10
Air (g) 41,30 41,30 25,73 19,42
Pati jagung (Maizena) (g) 671,35 671,35 645,37 609,35 Ketcrangan:
*
= tanpa CaC03 dan KH2P04; OvxC = Tikus yang diovariektomi yang mendapat pakan dengan sumber protein Casein; NonOvx = Tikus yang tidak diovariektomi yang mendapat pakan dengan sumber protein Casein; OvxH = Tikus yang diovariektomi yang mendapat pakan dengan sumber protein tahu; OvxT = Tikus yang diovariektomi yang mendapat pakan dengan sumber protein tempe.4.3 Perkembangan Berat Badan, Jumlah Konsumsi Pakan, dan Efmiensi Pakan Tikus selama Percobaan
[image:140.588.79.509.150.426.2]3 1 diduga terjadi karena pada minggu kedua ini mulai masuk tikus dari peneliti lain yang menggunakan ruang kandang yang sama, sehingga tikus hams beradaptasi kembali dengan suasana lingkungan yang baru. Hal yang sama juga terjadi pada minggu keempat
.
Pada minggu keempat ini suasana lingkungan ruang kandang kembali berubah karena masuknya pengguna baru.+
NonOvx0 5 10 15 20 24 27 28 32
[image:141.588.77.507.199.636.2]Waktu Percobaan (hari)
Gambar 7. Perkembangan berat badan tikus selama percobaan; Tikus Ovariektomi yang mendapat pakan dengan surnber protein kasein (OvxC); Tikus yang tidak diovariektomi yang menda- pat pakan dengan sumber protein kasein (NonOvx); Tikus ovariektomi yang mendapat pakan dengan sumber protein tahu (OvxH); Tikus ovariektomi yang mendapat pakan dengan sumber protein t e m p (OvxT).
32 disebabkan oleh uap amonia yang berasal dari urin dan feses yang menimbulkan penyakit pa.-paru.
Uji sidik ragam menunjukkan bahwa perlakuan berpengaruh sangat nyata (a
= 0,Ol) terhadap kenailcan berat badan tikus selama percobaan (Lampiran 3). Pengaruh perlakuan terhadap pertambahan berat badan tikus dapat dilihat pada Tabel 8. Tikus yang diovariektomi (OvxC, OvxH, OvxT) mengalami pertambahan berat badan yang lebih besar jika dibandingkan dengan tikus yang tidak
diovariektomi (NonOvx). Arjmandi et. a1. (1 996) mendapatkan data pertambahan berat badan yang sejalan dengan penelitian ini, tetapi peningkatan berat badan akibat ovariektomi tersebut dapat dicegah dengan pemberian 1793 estradiol atau konsentrat protein kedelai.
Tabel 8. Pengaruh perlakuan terhadap pertambahan berat badan dan terhadap perbandingan pertambahan berat badan dengan jumlah konsumsi pakan rata-rata
Parameter
Perlakuan Pertambahan Konsumsi pakan rata- Pertambahan berat berat badan (g) rata (g bk) badan/konsumsi
pakan rata rata
NonOvx 30,OOa 9,56a 3,13a
OvxC 41,33b 10,91a 3,84bc
OvxH 48,33b 10,67a 4,34c
OvxT 3 8,73 b 10,72a 3,62ab
Keterangan: H w f berbeda ke arah kolom menunjukkan perbedaan yang nyata (a=S%);
OvxC = Tik~ls ovariektomi yang mendapat pakan dengan sumber protein kasein; NonOvx = Tikus
[image:142.586.64.511.428.605.2]3 3 Peningkatan berat badan tikus yang mengalami ovariektomi diduga akibat terjadinya peningkatan simpanan lemak pada jaringan adiposa. Ovariektomi telah menyebabkan berkurangnya produksi estrogen, sehingga terjadi penurunan katabolisme lemak. Akibatnya terjadi peningkatan deposit lemak pada jaringan adiposa. Percobaan menggunakan mencit yang dibuat tidak mampu menghasilkan estrogen endogen (Jones et. al., 2000) dan mencit yang tidak memiliki reseptor estrogen alpa
(ERa)
mendukung dugaan tersebut, yaitu bahwa peningkatan berat badan tikus disebabkan oleh adanya peningkatan berat jaringan lemak putih (White Adipose Tissue). Jones et.al. (2000) mengajukan dua mekanisme biokimiawi yang mungkin berkenaan dengan peran estrogen pada homeostasis lemak. Pertama, adanya gangguan penggunaan dan penyimpanan glukosa otot pada tikus yang kekurangan estrogen (misalnya pada tikus yang mengalami ovariektomi) akan menyebabkan pen- lean body mass. Penurunan penggunaan glukosa oleh otot ini akan menyebabkan meningkatnya jumlah glukosa yang tersedia untuk proses lipogenesis sehingga mendorong terjadinya penimbunan lemak. Otot juga bertanggung jawab atas sebagian besar penggunaan glukosa setelah makan34 Peningkatan berat badan tikus yang mengalami ovariektomi pada penelitian ini ternyata tanpa disertai dengan peningkatan jumlah konsumsi pakan (Lampiran 1 dan Tabel 8). Wade dan Gray (1985) yang dikutif Heine et.al. (2000) mendapatkan hal yang sama pada tikus dan hamster, dan menduga ha1 tersebut disebabkan oleh adanya penurunan pengeluaran energi (energy expenditure). Heine et.01. (2000) menganggap bahwa p e n m a n pengeluaran energi merupakan falctor penting yang menyebabkan kegemukan pada tikus yang tidak memiliki reseptor estrogen alfa (ERa). Tetapi pada mencit yang tidak mampu memproduksi estrogen endogen peningkatan berat badan justru disertai dengan p e n m a n jumlah konsumsi pangan. Pada mencit jenis ini peningkatan berat badan diduga disebabkan oleh adanya
penurunan tingkat aktivitas fisik spontan, penurunan oksidasi glukosa, dan p e n m a n lean bod' mass (Jones et. al., 2000).
Uji sidik ragam menunjukkan bahwa perlakuan berpengaruh nyata ( a = 0,05) terhadap efisiensi pakan (pertambahan berat badan Ijumlah konsumsi pakan )
(Lampiran 5). Dari Tabel 8 terlihat bahwa efisiensi pakan pada tikus yang
mengalami ovariektomi lebih besar dibandingkan dengan tikus normal, kecuali pada tikus yang mengalami ovariektomi yang mendapat pakan tempe. Heine et.al. (2000) menganggap bahwa peningkatan efisiensi pakan dan tidak adanya peningkatan jumlah konsumsi pakan pada tikus jantan dewasa yang tidak memiliki reseptor
3 5
Peningkatan berat badan, peningkatan efisiensi pakan dan jumlah konsumsi pakan yang tetap pada penelitian ini mendukung bukti adanya proses penghematan energi yang dapat berupa penurunan pengeluaran energi, penurunan oksidasi glukosa, penurunan tingkat aktivitas, atau penurunan lean body mass sebagai akibat pengaruh defisiensi estrogen. Pada wanita, fase penghematan ditemukan terjadi pada masa awal kehamilan dan premenopouse sebagai akibat terjadinya penurunan oksidasi lemak yang disebabkan oleh estrogen (O'Sullivan et.al., 2001)
Efisiensi pakan tempe ternyata tidak berbeda nyata dengan tikus normal (Tabel 2). Data ini mendukung dugaan Arjmandi et.al. (1996) yang menganggap bahwa penghambatan peningkatan berat badan (yang juga menunjukkan peningkatan efisiensi pakan) tikus yang mengalami ovariektomi akibat pemberian pakan tempe disebabkan oleh senyawa isoflavon yang bersifat agonis terhadap estrogen. Temuan adanya penurunan lemak abdominal pada tikus yang mengalami ovariektomi yang diberi pakan konsentrat protein kedelai bila dibandingkan dengan tikus normal (Arjmandi et.al., 1996) menunjukkan adanya pengaruh senyawa isoflavon terhadap proses metabolisme lemak.
4.4 Berat Uterus dan Berat Uterus per Berat Badan
3 6 Ovariurn adalah sumber utama estrogen yang berperan dalarn proliferasi sel- sel uterus (Granner, 1990). Estrogen menyebabkan hiperemia dan meningkatkan penggabungan asam amino, sintesis asam nukleat, dan retensi nitrogen di dalam endometrium. Estrogen juga menstimulasi fosforilasi, glikolisis aerobik dan anaerobik, dan deposisi glikogen (Hafez, 1 970).
Tabel 9. Pengaruh perlakuan terhadap berat uterus dan terhadap perbandingan berat uterus dengan berat badan
Parameter
Perlakuan Berat uterus (g) Berat uteruskrat badan
NonOvx 0,392a 0,00 15a
OvxC 0,0667b 0,00023 b
OvxH 0,0709b 0,00027b
OvxT 0,076 1 b 0,00029b
Keterangan: Hurup berbeda ke arah kolom menunjukkan perbedaan yang nyata (a=5%); OvxC = Tikus ovariektomi yang mendapat pakan dengan sumber protein kasein; NonOvx
-
Tikus yang tidak diovariektomi yang mendapat pakan dengan sumber protein kasein; OvxH = Tikus ovariektomi yang mendapat pakan dengan sumber protein tahu; OvxT = Tikus ovariektomi yang mendapat pakan dengan sumber protein tempe.atropi uterus pada tikus menyusui, tikus muda (Anderson et.al, 1995) atau mencit (Ishimi et.al., 1999) yang diovariektomi.
Uterus mengandung lebih banyak reseptor estrogen alfa (ERa) bila dibandingkan dengan reseptor estrogen beta (ERP). Isoflavon bersifat antagonis terhadap estrogen pada ERa, sehingga pada jaringan yang banyak mengandung ERa senyawa ini menempati reseptor estrogen dan memblok stimulasi DNA dan sintesis protein yang diinduksi oleh estradiol (Anderson, 1998). Kuiper et.al. (1997)
sebelumnya juga telah menunjukkan bahwa genistein, sebagai senyawa isoflavon dengan efek estrogenik terkuat, mempunyai afinitas yang lebih tinggi terhadap ERP bila dibandingkan dengan ERa.
4.5 Kadar Kalsium Serum
Uji sidik ragam menunjukkan bahwa perlakuan berpengaruh sangat nyata
(a= 0,Ol) terhadap kadar kalsium serum (Lampiran 11). Dari Tabel 10 dapat diduga bahwa ovariektomi telah menyebabkan terjadinya peningkatan resorpsi tulang pada tikus percobaan. Ovariektomi menyebabkan peningkatan kadar kalsiurn senun, yang merupakan salah satu indikator terjadinya peningkatan proses resorpsi tulang
38 Estrogen menghambat pembentukan interleukin- 1 (IL- I), tumour necrosis factor (TNF) (Gowen, 1991) dan interleukin-6 (IL-6) (Girasole et.al., 1992). Ketiga
jenis sitokin ini dapat menstimulasi aktivitas osteoklas sehingga terjadi peningkatan laju resorpsi tulang. Kekurangan estrogen akibat ovariektomi menyebabkan
peningkatan produksi ketiga jenis sitokin tersebut.
Tabel 1 0. Pengaruh perlakuan terhadap kadar kalsium serum
Perlakuan Kadar kalsium serum (mg/lOOml)
NonOvx 1,39a
OvxC 2,19b
OvxH 2,5 1 b
OvxT 2,29b
Keterangan: Hurup berbeda ke arah kolom menunjukkan perbedaan yang nyata (a=5%);
OvxC = Tikus ovariektomi yang mendapat pakan dengan sumber protein kasein; NonOvx = Tikus yang tidak diovariektomi yang mendapat pakan dengan sumber protein kasein; OvxH = Tikus ovariektomi yang mendapat pakan dengan sumber protein tahu; OvxT = Tikus ovariektomi yang mendapat pakan dengan sumber protein tempe.
Walaupun menemukan beberapa indikator terjadinya peningkatan laju remodeling tulang (seperti peningkatan kadar alkalin fosfatase dan tartrate-resistant acidphosphatase) akibat ovariektomi, tetapi Arjmandi et.al. (1 996) tidak menemukan perbedaan kadar kalsium pada serum tikusnya. Peningkatan indikator tersebut tidak dapat dicegah oleh pemberian isoflavon, walaupun isoflavon terbukti menghambat kehilangan tulang. Berdasarkan hal ini maka Arj mandi et. a1. (1 996) berpendapat bahwa isoflavon meningkatkan laju pembentukan tulang sehingga melampaui laju resorpsinya.
dibandingkan dengan konsumsi rata-rata isoflavon pada penelitian Arjmandi et.al (1 996), yaitu 8,03 1 mg/kg berat badan/hari. Diduga dosis isoflavon pada penelitian ini tidak memberikan efek yang berarti pada laju pembentukan tulang, sehingga laju resorpsinya lebih tinggi jika dibandingkan dengan laju pembentukannya. Kadar kalsium serum meningkat akibat pelepasan kalsium akibat resorpsi dan penyerapan dari makanan tidak diimbangi dengan penggunaanya untuk pembentukan tulang.
Peningkatan kalsium serum pada pene