DIVERSITAS BAKTERI ASAL SPORA FUNGI MIKORIZA
ARBUSKULA GIGASPORA SP. DAN GLOMUS SP. SERTA
POTENSINYA SEBAGAI MYCORRHIZA HELPER BACTERIA
NUNANG LAMAEK MAY
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS
DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis “Diversitas Bakteri asal Spora Fungi Mikoriza Arbuskula Gigaspora sp. dan Glomus sp. serta Potensinya sebagai
Mycorrhiza Helper Bacteria” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2011
Nunang Lamaek May
ABSTRACT
NUNANG LAMAEK MAY. Bacterial Diversity from The Spores of Arbuscular Mycorrhizal Fungi Gigaspora sp. and Glomus sp. and Their Potential as Mycorrhiza Helper Bacteria. Under academic supervision of SRI WILARSO BUDI R and ARUM SEKAR WULANDARI.
Mycorrhiza Helper Bacteria are bacteria that potentially assist the development of mycorrhiza. This study was aimed to explore the Mycorrhizal Helper Bacteria (MHB) from the spore of Arbuscular Mycorrhizal Fungi (AMF) Gigasprora sp. and Glomus sp. The enzymatic activity (cellulase, protease and pectinase), antagonists with pathogen Sclerotium sp., Rhizoctonia sp., Ganoderma sp. and mycorrhizal stimulation were conducted. The results showed that there were 7 species of bacteria isolated from the spores of Gigaspora sp. and 5 species from the spores of Glomus sp. Of the 12 bacteria, 7 bacteria had the potential to produce the enzymes of hydrolytic cellulase and proteases. Based on in vitro test, there were 4 species of bacteria found to have the ability to inhibit the growth of 3 species of pathogens tested. Eight types of bacteria were found to be capable of stimulating the development of mycorrhizal hyphae. There were 3 isolates of bacteria showed the antagonistic toward pathogenic fungi as well as stimulation of AMF. The bacteria obtained were very potential to be developed as biological agents in improving the quality of soil and plants growth.
RINGKASAN
NUNANG LAMAEK MAY. Diversitas Bakteri asal Spora Fungi Mikoriza Arbuskula Gigaspora sp. dan Glomus sp. serta Potensinya sebagai Mycorrhiza Helper Bacteria. Dibimbing oleh Sri Wilarso Budi R. dan Arum Sekar Wulandari. Bakteri dikatakan MHB ketika bakteri itu bersifat endofit dengan kata lain bakteri tersebut harus berada di salah satu bagian tubuh mikoriza, dan berperan terhadap perkembangan mikoriza. Bakteri yang diisolasi dari fungi mikoriza dapat menstimulasi infeksi mikoriza, produksi spora dan juga perlawanan terhadap patogen tanaman. Dengan demikian penting untuk mengeksplorasi bakteri-bakteri tersebut dan menguji potensinya sebagai MHB dan sebagai upaya mengembangkan potensi FMA.
Tujuan dari penelitian ini untuk menemukan jenis-jenis bakteri pada spora FMA Gigaspora sp. dan Glomus sp., mendapatkan bakteri yang mampu menstimulir perkembangan FMA, mendapatkan isolat bakteri yang berpotensi menghasilkan enzim hidrolitik dan menemukan bakteri yang memiliki sifat antagonis terhadap patogen tanaman.
Penelitian ini dilaksanakan di Laboratorium Silvikultur Fakultas Kehutanan Institut Pertanian Bogor selama 11 bulan. Peneltian dibagi ke dalam 4 bagian penelitian yaitu 1) eksplorasi bakteri yang berasal dari spora FMA menggunakan metode Gardeman dan Nicholson (1963) dan Budi et al. (1999); 2) uji stimulasi mikoriza menggunakan metode Budi et al. (1999); 3) uji enzimatik menggunakan metode Teather dan Wood (1982), untuk pektinase dan selulase, serta metode Dunne et al. (1997) untuk protease; dan 4) uji antagonis menggunakan metode Varese et al. (1996); dan. Penelitian dirancang dengan menggunakan Rancangan Acak Lengkap dengan menggunakan bakteri hasil isolasi sebagai perlakuan dan diulang sebanyak 3 kali.
Hasil penelitian menunjukan bahwa ditemukan 12 jenis bakteri yang berasal dari spora FMA. Tujuh bakteri dijumpai pada spora Gigaspora sp. dan 5 jenis bakteri dari spora Glomus sp. Dari ke-12 bakteri tersebut, didapatkan 8 jenis bakteri mampu menstimulir perkembangan hifa mikoriza. Tujuh bakteri ditemukan mampu menghasilkan enzim hidrolitik selulase, dan protease. Dalam pengujian in vitro ditemukan 4 jenis bakteri memiliki kemampuan untuk menghambat pertumbuhan 3 jenis patogen yang diujikan (Sclerotium sp.
Rhizoctonia sp. dan Ganoderma sp.).
Tiga jenis bakteri yaitu Bacillus subtilis, Pseudomonas diminuta dan
Enterobacter hormaechei berpotensi menjadi MHB plus. Bakteri-bakteri tersebut ditetapkan sebagai MHB plus karena merupakan bakteri hasil isolasi dari spora FMA (endofit), mampu menstimulir pertumbuhan hifa, mempunyai kemampuan menghasilkan enzim hidrolitik dan mempunyai sifat antagonis terhadap patogen.
© Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebut sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
DIVERSITAS BAKTERI ASAL SPORA FUNGI MIKORIZA
ARBUSKULA
GIGASPORA
SP. DAN
GLOMUS
SP. SERTA
POTENSINYA SEBAGAI
MYCORRHIZA HELPER BACTERIA
NUNANG LAMAEK MAY
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Silvikultur Tropika
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Diversitas Bakteri asal Spora Fungi Mikoriza Arbuskula
Gigaspora sp. dan Glomus sp. serta Potensinya sebagai
Mycorrhiza Helper Bacteria Nama : Nunang Lamaek May
NRP : E451090011
Disetujui, Komisi Pembimbing
Dr. Ir. Sri Wilarso Budi R., MS Dr. Ir. Arum Sekar Wulandari, MS
Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Silvikultur Tropika
Dr. Ir. Sri Wilarso Budi R., MS Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Dengan penuh kerendahan hati penulis panjatkan ucapan syukur ke hadirat ALLAH YEHUWA, sumber kehidupan dan penyedia segala perkara yang baik atas perlindungan, kasih dan kesempatan menikmati kehidupan ini beserta segala aspeknya termasuk penyertaanNya selama penelitian dan penulisan tesis berjudul Diversitas Bakteri asal Spora Fungi Mikoriza Arbuskula Gigaspora sp. dan Glomus sp. serta Potensinya sebagai Mycorrhiza Helper Bacteria.
Dari sanubari yang terdalam penulis manyampaikan terima kasih, hormat dan penghargaan yang setulus-tulusnya kepada :
1. Dr. Ir. Sri Wilarso Budi R., MS selaku ketua komisi pembimbing dan Dr. Ir. Arum Sekar Wulandari, MS selaku anggota komisi pembimbing yang senantiasa mencurahkan tenaga, pikiran dan waktu dalam membimbing penulis sejak penyusunan rencana penelitian hingga penyelesaian tesis ini. 2. Dr. Ir. Prijanto Pamoengkas, M.Sc.F.Trop selaku penguji luar komisi yang
telah memberikan kritikan, masukan dan saran guna penyempurnaan tesis. 3. Ibunda, Oma tercinta dan Marlin May yang senantiasa memberikan doa restu
serta selalu mengingatkan penulis akan makna hidup ini.
4. Rektor Universitas Negeri Papua dan Dekan Fakultas Kehutanan UNIPA atas izin dan motivasi kepada penulis.
5. Kementerian Pendidikan Nasional RI yang telah memberikan Beasiswa Program Pascasarjana (BPPS 2009).
6. PEMDA Provinsi Papua atas pemberian Hibah Pendidikan Pascasarjana. 7. Seameo Biotrop yang telah memberikan DIPA penelitian 2010.
8. Civitas Akademika Program Pascasarjana IPB dan Departemen serta Mayor Silvikultur Tropika.
9. Dr. Irdika Mansur, Dr. Abimanya D. Nusantara, dan Dr. Supriyanto atas informasi mengenai mikoriza yang diberikan.
10. Evelin A Tanur, S.Hut dan Descarlo Worabai, S.Hut atas bantuan dan motivasinya.
11. Keluarga Yohanes Kwando dan Rozalina Mandabayan.
12. Emy Sadjati, S.Hut, Victor J.G. May, S.Hut, Kusriner F. Mabubedari, S.Hut, M.Si., dan Matheus Beljai, S.Hut atas kebersamaannya dalam berdiskusi. 13. Teman-teman SVK 2009 Agung, Rida, Desti, Andi, Anna, Fitri, Arief dan
Benyamin serta adik-adik SVK 2010 Mahardika, Jin won dan Paranita atas hari-hari indah bersama selama kuliah dan bantuan serta semangatnya. 14. Adik-adikku Oni, Novi, Adel, Elda, Laurensia, Osval, Sandra, Sartika, Willy,
Monalisa, Ina, Biman, Agil, Noni, Dio dan Valeria
15. Semua pihak yang pada kesempatan ini tak dapat penulis sebutkan satu persatu namanya namun budi baiknya akan selalu dikenang.
Semoga karya ilmiah ini bermanfaat bagi perkembangan Ilmu pengetahuan terutama bidang Kehutanan.
Bogor, Agustus 2011
RIWAYAT HIDUP
Penulis dilahirkan di Manado pada tanggal 26 Oktober 1983 sebagai anak sulung dari pasangan ayahanda Lamek May (Alm) dan ibunda Laurensina Taribaba. Jenjang pendidikan formal mulai SD hingga SMU diselesaikan di Manokwari. Pendidikan sarjana ditempuh di Jurusan Budidaya Hutan, Fakultas Kehutanan, Universitas Negeri Papua melalui jalur undangan seleksi siswa andalan masuk UNIPA (SESAMA UNIPA), lulus pada tahun 2007. Kesempatan untuk melanjutkan ke program magister pada Mayor Silvikultur Tropika, Sekolah Pascasarjana IPB tahun 2009. Beasiswa diperoleh dari Kementerian Pendidikan Nasional ditjen dikti melalui beasiswa pendidikan pascasarjana (BBPS). Penulis bekerja sebagai staf pengajar Fakultas Kehutanan, Universitas Negeri Papua sejak tahun 2008.
DAFTAR ISI
Halaman
DAFTAR TABEL ... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN ... xv
I PENDAHULUAN ... 1
1.1 Latar Belakang ... 1
1.2 Tujuan ... 2
1.3 Hipotesis ... 2
1.4 Manfaat ... 3
1.5 Kerangka Pemikiran ... 3
II TINJAUAN PUSTAKA ... 5
2.1 Diversitas Bakteri Asal Mikoriza ... 5
2.2 Fungi Mikoriza Arbuskula (FMA) ... 8
2.3 Mycorrhiza Helper Bacteria (MHB) ... 9
III METODE PENELITIAN ... 12
3.1 Tempat dan Waktu ... 12
3.2 Bahan dan Alat ... 12
3.3 Prosedur Penelitian ... 12
3.3.1 Eksplorasi Bakteri yang Berasal dari Spora FMA ... 12
3.3.1.1 Isolasi Spora FMA ... 12
3.3.1.2 Isolasi Bakteri dari Spora FMA ... 12
3.3.1.3 Karakterisasi dan Identifikasi Bakteri ... 13
3.3.2 Uji Stimulasi Mikoriza ... 13
3.3.3 Uji Enzimatik ... 13
3.3.4 Uji Antagonis ... 14
IV HASIL ... 15
4.1 Keragaman Jenis Bakteri Asal Spora FMA ... 15
4.2 Bakteri yang Menstimulasi Perkembangan Mikoriza ... 17
4.3 Bakteri yang Berpotensi Menghasilkan Enzim Hidrolitik ... 19
4.4 Bakteri yang Berpotensi Sebagai Antagonis Terhadap Fungi Patogen ... 23
VI KESIMPULAN DAN SARAN ... 37
6.1 Kesimpulan ... 37
6.2 Saran ... 37
DAFTAR TABEL
Halaman
1 Jumlah dan morfologi jenis bakteri yang diisolasi dari sporokarp jamur ektomikoriza ... 6
2 Jenis bakteri yang ditemukan dari sporokarp Glomus mosseae ... 6 3 Jenis-jenis bakteri yang ditemukan dari sporokarp Suillus grevillei
dan ektomikoriza ... 7
4 Jenis-jenis bakteri asal spora FMA ... 15
5 Perkembangan hifa mikoriza ... 17
6 Aktivitas enzimatik (diameter halo (cm)) bakteri yang diisolasi
dari spora Gigaspora sp. dan Glomus sp. ... 20 7 Perkembangan radial miselium dan persen penghambatan patogen
Sclerotium sp. ... 22 8 Perkembangan radial miselium dan persen penghambatan patogen
Rhizoctonia sp. ... 26 9 Perkembangan radial miselium dan persen penghambatan patogen
DAFTAR GAMBAR
Halaman
1 Bagan alir kerangka pemikiran ... 4
2 Jenis-jenis bakteri yang ditemukan dari spora FMA ... 16
3 Grafik perpanjangan hifa Gigaspora sp. ... 18
4 Penampilan perkembangan hifa mikoriza ... 19
5 Grafik diameter zona bening (cm) akibat aktivitas bakteri penghasil selulase ... 21
6 Penampakan zona bening enzim selulase ... 21
7 Grafik diameter zona bening (cm) akibat aktivitas bakteri penghasil protease ... 22
8 Penampakan zona bening enzim protease ... 22
9 Grafik panjang radial miselia patogen fungi Sclerotium sp. ... 24
10 Penampilan penghambatan patogen Sclerotium sp. oleh bakteri ... 25
11 Grafik panjang radial miselia fungi patogen Rhizoctonia sp. ... 26
12 Penampilan penghambatan patogen Rhizoctonia sp. oleh bakteri ... 27
13 Grafik panjang radial miselia fungi patogen Ganoderma sp. ... 28
14 Penampilan penghambatan patogen Ganoderma sp. oleh bakteri ... 29
DAFTAR LAMPIRAN
Halaman
1 Identifikasi bakteri dari spora Gigaspora sp. ... 42 2 Identifikasi bakteri dari spora Glomus sp. ... 44 3 Sidik ragam aktivitas bakteri yang berpotensi menghasilkan enzim
hidrolitik ... 46
4 Sidik ragam bakteri terhadap perkembangan radial miselium
fungi patogen ...47
BAB I
PENDAHULUAN
1.1 Latar belakang
Sebagai medium pertumbuhan tanaman, tanah mengandung organisme hidup dalam jumlah yang relatif besar dan termasuk komponen fundamental yang penting. Banyak aktivitas biota tanah yang berpengaruh terhadap kesuburan tanah dan pertumbuhan tanaman. Masing-masing organisme berperan dalam ekosistem, terutama terkait dengan aliran energi dan siklus unsur hara sebagai akibat dari aktivitas utama organisme hidup (Ahemad et al. 2009).
Mikoriza adalah asosiasi saling menguntungkan antara fungi dan akar tanaman yang membentuk struktur simbiotik. Melalui hubungan simbiosis dengan tanaman, mikoriza berperan penting dalam pertumbuhan tanaman, perlindungan penyakit, dan peningkatan kualitas tanah secara keseluruhan. Dengan demikian mikoriza sangat berperan dalam produktivitas tanaman (Siddiqui dan Pithtel 2008).
Fungi Mikoriza Arbuskula (FMA) adalah salah satu komponen mikroorganisme tanah yang banyak dijumpai pada kebanyakan agroekosistem. Budiman (2000) melaporkan bahwa Duabanga moluccana yang terinfeksi oleh
Glomus fasciculata memiliki pertumbuhan yang lebih baik dibandingkan dengan yang tidak terinfeksi. Fakta ini menunjukan bahwa FMA dapat berperan sebagai biofertiliser (Mulongoy et al. 1992), bioregulator (Lovato et al. 1994), dan bioprotektor (Linderman 1994).
Peranan FMA dan organisme pendukungnya (bakteri) berpotensi untuk diaplikasikan sebagai biofertiliser. Bakteri yang mampu meningkatkan perkembangan mikoriza diberi nama Mycorrhiza Helper Bacteria (MHB) (Garbaye 1994). Beberapa peneliti menemukan bahwa bakteri yang diisolasi dari fungi mikoriza dapat menstimulasi infeksi mikoriza, produksi spora dan juga perlawanan terhadap patogen tanaman (Garbaye 1994; Von Alten et al. 1993, Barea et al. 1998, Budi et al. 1998).
Jumlah bakteri di dalam tanah sangat banyak (satu sendok tanah produktif mengandung 100 juta sampai 1 milyar bakteri) (Handayanto dan Hairiah 2007) namun tidak semua dapat berperan sebagai MHB. Bakteri dinyatakan sebagai MHB sejati ketika bakteri tersebut benar-benar hidup berasosiasi dengan fungi mikoriza dan dapat menstimulasi perkembangan FMA (Garbaye dan Bowen 1989). Untuk mendapatkan bakteri yang berpotensi sebagai MHB perlu dilakukan eksplorasi dan pengujian. Eksplorasi bakteri yang berpotensi sebagai MHB merupakan upaya pengembangan potensi FMA.
1.2 Tujuan
Tujuan dari penelitian ini ialah:
1. Menemukan jenis-jenis bakteri pada spora FMA Gigaspora sp. dan Glomus sp.,
2. Mendapatkan bakteri yang mampu menstimulir perkembangan FMA, 3. Mendapatkan bakteri yang berpotensi menghasilkan enzim hidrolitik, dan 4. Menemukan bakteri yang bersifat antagonis terhadap patogen tanaman.
1.3 Hipotesis
1. Bakteri MHB dapat diisolasi dari spora FMA Gigaspora sp. dan Glomus sp., 2. Ada bakteri yang dapat menstimulir perkembangan FMA,
1.4 Manfaat
Manfaat penelitian ini ialah memberikan informasi tentang mikroorganisme (bakteri endofit) yang memiliki peran membantu aktivitas FMA sehingga dapat digunakan bersama-sama untuk diinokulasikan ke tanaman.
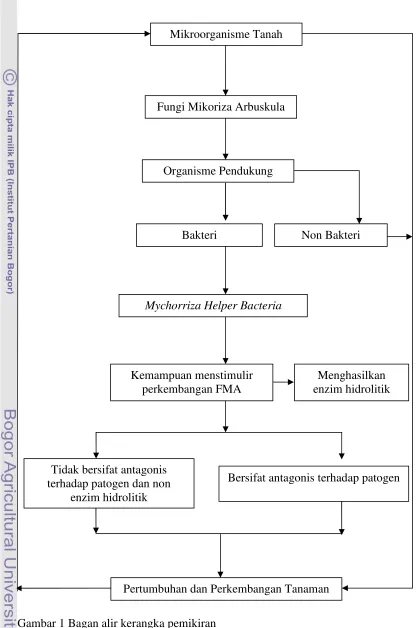
1.5 Kerangka Pemikiran
Di dalam tanah, masing-masing organisme memainkan peran penting dalam ekosistem. Aktivitas mikroorganisme tanah di sekitar rizosfer sangat membantu perbaikan kesuburan tanah dan berdampak pada peningkatan produktivitas tanaman. FMA adalah salah satu mikroorganisme tanah yang banyak menyediakan keutungan pada tanaman inang, antara lain: transfer unsur hara, membantu perkembangan tanaman dan dapat menjadi agen pengendali hayati terhadap patogen akar.
Gambar 1 Bagan alir kerangka pemikiran
Mikroorganisme Tanah
Fungi Mikoriza Arbuskula
Organisme Pendukung
Non Bakteri Bakteri
Mychorriza Helper Bacteria
Kemampuan menstimulir perkembangan FMA
Tidak bersifat antagonis terhadap patogen dan non
enzim hidrolitik
Bersifat antagonis terhadap patogen
Pertumbuhan dan Perkembangan Tanaman
BAB II
TINJAUAN PUSTAKA
2.1 Diversitas Bakteri Asal Mikoriza
Keberadaan mikroorganisme tanah di alam sangat melimpah dalam keragaman dan jumlah jenisnya. Setiap mikroorganisme memiliki peran spesifik terhadap pertumbuhan dan perkembangan tumbuhan. Beberapa mikroorganisme memiliki sifat parasit namun ada pula yang bersifat menguntungkan dan tidak sedikit pula yang keberadaannya bersifat netral.
Aktivitas mikroorganisme tanah di sekitar rizosfer sangat membantu perbaikan kesuburan tanah dan berdampak positif terhadap peningkatan kualitas dan kuantitas produksi tanaman. Penggunaan mikroorganisme untuk meningkatkan pertumbuhan tanaman merupakan hal menarik yang dalam beberapa tahun terakhir ini mendapat perhatian para peneliti. Sejalan dengan perkembangan ilmu bioteknologi, mikroorganisme tanah merupakan objek yang mempunyai prospek untuk dikembangkan (Barriuso et al. 2008).
Mikroba yang memiliki sifat asosiasi dengan tanaman inang, umumnya memiliki sifat kompatibilitas antara inang dan mikroba tersebut. Mikroba yang dapat berasosiasi dengan tanaman inang tertentu, belum tentu dapat berasosiasi dengan tanaman inang yang lain. Dengan demikian, perlu dilakukan suatu penelitian untuk mendapatkan jenis-jenis mikroba yang kompatibel dengan jenis tanaman yang diinginkan. Ada beberapa rangkaian kegiatan pengujian yang harus dilakukan hingga didapatkan isolat mikroorganisme yang efektif membantu pertumbuhan tanaman.
Keberadaan bakteri yang terlibat secara langsung dalam pembentukan mikoriza pertama kali dibuktikanoleh penelitian yang dilakukan oleh Bowen dan Theodorou (1979). Beberapa penelitian telah dilakukan dan ditemukan adanya bakteri yang berasosiasi dengan mikoriza. Dahm et al. (2005) mengisolasi bakteri dari beberapa jenis fungi ektomikoriza dan mendapatkan sejumlah bakteri dengan jumlah
jenis dan penampakan morfologi yang berbeda (Tabel 1). Budi et al. (1999) mengisolasi
bakteri asal sporokarp FMA Glomus mosseae dan mendapatkan delapan jenis
mengisolasi bakteri spora FMA tanpa disterilkan dari daerah mikorizosfer kelapa sawit dan menemukan 20 isolat bakteri.
Tabel 1 Jumlah dan morfologi jenis bakteri yang diisolasi dari sporokarp fungi ektomikoriza
Fungi Jumlah
isolat yang dijumpai Jumlah gram negatif (batang) Jumlah gram positif (basil) Jumlah gram positif (kokus) Jumlah bakteri variabel pelomorfik Amanita muscaria Hebeloma
crustuliniforme (1) Hebeloma
crustuliniforme (2) Hebeloma
crustuliniforme (3) Laccaria amethistina Laccaria laccata (1) Laccaria laccata (2) Laccaria laccata (3) Laccaria proxima1 Lycoperdon sp. Scleroderma sp. Suillus grevillei Suillus luteus
Thelephora terrestris Xerocomus sp.
3 30 30 30 29 30 29 30 30 30 30 30 30 30 29 3 21 18 28 5 30 29 27 27 30 30 20 28 30 27 – 6 – – – – – 1 – – – 1 2 – 2 – 2 12 – 24 – – – – – – 9 – – – – 1 – 2 – – – – 3 – – – – – –
Sumber : Dahm et al. (2005)
Tabel 2 Jenis bakteri yang ditemukan dari sporokarp Glomus mosseae
Nomor isolat Jenis-jenis bakteri yang ditemukan B1 Methylobacterium sp.
B2 Paenibacillus azotofixans
B3 Bacillus megatorium
B4 Bacillus firmus
B5 Bacillus circulans
B6 Bacillus polymixa
B7 Bacillus sp.
B8 Paenibacillus aspiarius
Tabel 3 Jenis–jenis bakteri yang ditemukan dari sporokarp Suillus grevillei dan ektomikoriza
Bakteri asal Sporokarp Bakteri asal Ektomikoriza
Bacillus bravis Bacillus cereus Bacillua megaterium Bacillus mycoides Bacillus polymixa Bacillus purnilus Bacillus subtilis
Enterobacter aglomerans Flavobacterium oryzihabitans Micrococcus luteus
Pseudomonas aureofaciens Pseudomonas cepacia Pseudomonas chloraphis Pseudomonas fluorocens Serratia fonticola Streptomyces spp.
Acinobacter calcoaceticus Aeromonas hydrophyla Agrobacterium radiabacter Bacillus cereus
Bacillus mycoides
Enterobacter aglomerans 3 Enterobacter aglomerans 4 Enterobacter sakazakii Klebseiella pneumoniae Klebseiella ozaenae Micrococcus luteus Pseudomonas cepacia Pseudomonas aeruginosa Pseudomonas putida Pseudomonas choraphis Serratia liquefaciens Serratia marcescens Serratia rubidae Streptomyces sp.
2.2 Fungi Mikoriza Arbuskula (FMA)
Mikoriza adalah asosiasi saling menguntungkan antara fungi dan akar tanaman yang membentuk struktur simbiotik dan menghasilkan sifat morfologi yang baru. Melalui hubungan simbiosis dengan tanaman, mikoriza berperan penting dalam pertumbuhan tanaman, perlindungan penyakit, dan peningkatan kualitas tanah secara keseluruhan. Dari tujuh jenis mikoriza yang dijelaskan dalam literatur ilmiah saat ini (FMA, ektomikoriza, ektendomikoriza, arbutoid, monotropoid, mikoriza ericoid dan mikoriza orchid), FMA dan Ektomikoriza adalah yang paling banyak diteliti.
Mikoriza dapat berfungsi sebagai alat untuk pertanian berkelanjutan karena mempunyai kemampuan dalam meningkatkan pertumbuhan sistem perakaran tanaman, meningkatkan vigor tanaman dan kualitas tanah. Hifa dari mikoriza memperluas bidang perakaran serta mengeluarkan enzim, membantu penyerapan nutrisi secara efisien dan dapat berperan sebagai kontrol patogen. Dengan demikian, mikoriza sangat berperan dalam produktivitas tanaman (Siddiqui dan Pithtel 2008).
Fungi pembentuk mikoriza efektif mendukung pertumbuhan tanaman setelah terbentuk kolonisasi pada akar tumbuhan inangnya. Kolonisasi merupakan jembatan untuk transfer timbal balik antara fungi pembentuk mikoriza dan tanaman inangnya. Untuk dapat berkolonisasi pada tumbuhan simbion, mikobion harus mampu hidup, tumbuh dan berkembang pada rizosfer tumbuhan simbion. Dengan demikian, kemampuan beradaptasi mikroba di lingkungannya merupakan kunci keberhasilan simbiosisnya.
Perkembangan hifa yang ekstensif yang berasosiasi dengan akar tanaman inang menyebabkan terbentuknya apresorium. Ketika tidak terjadi sinyalisasi, morfogenesis dan apresorium tidak terbentuk. Studi interaksi simbiotik dan patogen menunjukkan bahwa pembentukan apresorium dapat dianggap sebagai tanda keberhasilan pengenalan tanaman inang sebelum terjadi infeksi ke inang. Apresorium merupakan ujung hifa yang berbentuk elips, rata dan terbentuk pada permukaan akar inang.
Berbeda dengan patogen, proses infeksi fungi endofit dicirikan dengan produksi enzim pendegradasi dinding sel dalam jumlah kecil dan terkontrol. FMA hanya mengkolonisasi jaringan akar spesifik seperti epidermis dan korteks sedangkan jaringan lainnya seperti pembuluh dan meristem resisten terhadap kolonisasi.
2.3 Mycorrhiza Helper Bacteria (MHB)
MHB merupakan istilah yang digunakan bagi bakteri yang dapat membantu mikoriza menjalankan perannya. Bakteri dikatakan MHB ketika bakteri itu bersifat endofit dengan kata lain bakteri tersebut harus berada di salah satu bagian tubuh mikoriza, dan berperan terhadap perkembangan mikoriza. Simbiosis mikoriza bukan hanya hubungan antara fungi pembentuk mikoriza dan tanaman inang namun melibatkan organisme pendukung lainnya seperti bakteri.
Hubungan antara fungi, tanaman inang dan MHB membentuk mikorizosfer yaitu daerah sekitar perakaran dimana fungi, bakteri dan akar tanaman saling berinteraksi. Akar bermikoriza dan hifa fungi mikoriza turut membentuk komposisi jenis bakteri akibat eksudasi dan pergantian akar atau hifa (Bowen 1993; Morgan et al. 2005). Keberadaan mikorizosfer dapat menyebabkan peningkatan nutrisi tanah dan membantu tanaman bertahan terhadap serangan patogen (Linderman 1988; Frey-Klett et al. 2005).
Eksudat MHB sering kali merangsang perkecambahan spora fungi. Xavier dan Germida (2003) mengamati bahwa sebagian besar bakteri dari dinding sel spora FMA mampu meningkatkan perkecambahan spora G. clarum ketika terjadi kontak langsung antara spora dan bakteri, sementara sebagian isolat bakteri menghambat perkecambahan spora dengan menghasilkan volatile antagonistic.
Duponnois dan Garbaye (1990) menganalisis bagaimana MHB mempengaruhi konsentrasi senyawa antagonistik yang diproduksi oleh fungi mikoriza. Mereka mendapati bahwa bakteri tersebut mampu mendetoksifikasi media cair dari metabolit fungi yang bersifat menghambat. Bakteri MHB kemungkinan juga dapat menekan produksi senyawa toksik oleh mikroba tanah. Vivas et al. (2005) melaporkan bahwa bakteri MHB memiliki dampak positif yang kuat terhadap perkecambahan spora dan pertumbuhan fungi prasimbiosis dalam larutan yang terkontaminasi logam berat. Inokulasi bakteri bukan hanya menurunkan kerusakan hifa G. mosseae tetapi bahkan menghasilkan peningkatan pertumbuhan akar dari 95% (tanpa Cd) menjadin 254% (dengan larutan Cd). Pengaruh ini sama kuatnya dengan pada perlakuan Zn di mana pertumbuhan miselium berkisar dari 125% (tanpa Zn) hingga 232% (dengan larutan Zn).
Produksi akar secara lateral dapat dipengaruhi secara positif oleh MHB (Garbaye 1994; Schrey et al. 2005), kemungkinan karena produksi auksin atau senyawa yang berhubungan dengan auksin oleh bakteri. Pembentukan ujung akar baru dapat menyebabkan pembentukan lebih banyak mikoriza, karena kerapatan situs kolonisasi per volume tanah meningkat. Paenibacillus sp. EJP73 dan
Burkholderia sp. EJP67, dan dua isolat strain dari mikoriza Lactarius rufus, ditemukan meningkatkan percabangan dikotomus pada semai Pinus sylvestris.
Beberapa laporan menyatakan bahwa kombinasi inokulasi dengan Plant Growth Promoting Bacteria (PGPB) dan fungi mikoriza dapat menghasilkan pengaruh positif secara sinergis terhadap pertumbuhan tanaman. Inokulasi dengan Azospirillum brasilense dan FMA Gigaspora margarita atau Glomus fasciculatum menyebabkan peningkatan biomassa pucuk dan akar pada
Pennisetum americanum, sebagai akibat dari peningkatan penyerapan fosfor (Rao
pertumbuhan tanaman timun. Gamalero et al. (2003) menduga bahwa alasan yang dapat menjelaskan hal ini adalah kuatnya laju produksi IAA oleh bakteri.
Untuk mengilustrasikan kompleksitas interaksi di dalam mikorizosfer, Gamalero et al. (2004) menunjukkan bahwa penggunaan dua strain bakteri bersama dengan satu FMA secara kuat meningkatkan pertumbuhan tomat. Strain MHB P. fluorescens 92 digunakan bersama dengan strain PGPB P. fluorescens
P190r, dan kombinasi kedua bakteri ini dengan G. mossaeae BEG12 menyebabkan peningkatan pertumbuhan tanaman secara nyata.
Dalam sebuah laporan tentang tanah yang terkontaminasi timah, Vivas et al. (2003) menunjukkan bahwa isolat Bacillus brevis mampu meningkatkan pembentukan mikoriza dan nodulasi, mampu menurunkan jumlah Pb yang diserap tanaman, mampu meningkatkan biomassa tanaman, dan akumulasi N dan P dalam
Trifolium pretense. Sebagai akibat dari perubahan nyata arsitektur akar, diduga bahwa produksi IAA oleh Bacillus brevis kemungkinan cukup tinggi.
BAB III
METODE PENELITIAN
3.1 Tempat dan Waktu
Penelitian ini dilaksanakan di Laboratorium Silvikultur Fakultas Kehutanan Institut Pertanian Bogor dari bulan Maret 2010 – Februari 2011. 3.2 Bahan dan Alat
Bahan-bahan yang digunakan pada penelitian ini yaitu: alkohol, spirtus, aquades, tissue, parafilm, kertas saring, aluminium foil, kertas label, zeolit, media
Potato Dextros Agar (PDA), media Nutrient Broth (NB), media Nutrient Agar
(NA), media enzim protease, media enzim selulase, media enzim pektinase, Kloramin-T 2%, Streptomycin 0.02%, spora FMA Glomus sp., spora FMA
Gigaspora sp., fungi patogen (Rhizoctonia sp., Sclerotium sp. dan Ganoderma
sp.).
Alat-alat yang digunakan pada peneitian ini yaitu: saringan bertingkat,
Laminar Air Flow Cabinet, mikroskop, autoklaf, timbangan analitik, hot plate dan stirer, oven, shaker, inkubator, sentrifus, refrigerator, labu ukur, labu Erlenmeyer, gelas piala, cawan Petri, bunsen, spatula, jarum inokulasi, pipet, pipet mikro, jarum suntik, botol film, ependof, botol semprot, gunting, stopwatch, kamera digital.
3.3 Prosedur Penelitian
3.3.1 Eksplorasi Bakteri yang Berasal dari Spora FMA 3.3.1.1 Isolasi spora FMA
Spora yang digunakan berasal dari jenis Gigaspora sp. dan Glomus sp. yang diperoleh dari koleksi Laboratorium Silvikultur, Fakultas Kehutanan IPB. Isolasi spora dilakukan dengan menggunakan metode Gardeman dan Nicholson (1963). Spora yang telah ditemukan dimasukan ke dalam botol 10 ml, masing-masing sebanyak 50 spora per botol dan disimpan di dalam refrigerator.
3.3.1.2 Isolasi bakteri dari spora FMA
selanjutnya disebar di atas media NA dan disimpan di dalam inkubator pada suhu 300C selama tiga hari, untuk mendapatkan koloni bakteri.
3.3.1.3 Karakterisasi dan identifikasi bakteri
Koloni yang tumbuh dikarakterisasi secara morfologi (warna, permukaan koloni, tipe koloni) kemudian dimurnikan. Identifikasi bakteri dilakukan di Laboratorium Teknologi Hayati, Institut Teknologi Bandung. Metode yang digunakan adalah metode Analitycal Profile Index (API) (Buchanan dan Gibbons (1974); Holt et al. (1994); Cappuccino dan Sherman (2005)). Secara garis besar analisis menggunakan metode ini dilakukan dengan melihat bentuk makroskopis koloni, bentuk mikroskopis sel, motilitas dan reaksi biokimia dari bakteri.
3.3.2 Uji Stimulasi Mikoriza
Uji stimulasi spora FMA dilakukan secara in vitro menggunakan spora
Gigaspora sp. Pengujian dilakukan sesuai dengan metode Budi et al. (1999). Sterilisasi permukaan spora dilakukan sebelum pemberian perlakuan. Perkembangan spora dan hifa awal diukur terlebih dahulu dengan mengambil 100 spora secara acak untuk dilakukan pengamatan dan pengukuran. Rata-rata dari 100 spora tersebut digunakan sebagai pendekatan untuk mengetahui perkembangan spora dan hifa sebelum diberi perlakuan. Spora yang diberi perlakuan ditempatkan pada kertas saring steril yang dijenuhkan di dalam 100 ml suspensi bakteri dan dimasukan ke dalam cawan Petri berisi media zeolit dengan jumlah spora sebanyak 10 spora untuk setiap cawan Petri. Cawan Petri direkatkan dengan parafilm, kemudian diinkubasi dalam keadaan gelap pada suhu 300C selama 21 hari.
Pengamatan dilakukan dengan mengukur panjang hifa dibandingkan dengan kontrol. Isolat bakteri yang dapat menstimulasi pertumbuhan mikoriza dapat dipertimbangkan sebagai MHB.
Uji stimulasi mikoriza dilakukan menggunakan Rancangan Acak Lengkap dengan 13 perlakuan dan diulang sebanyak 3 kali. Data yang diperoleh diolah menggunakan perangkat statistik SAS dan dilanjutkan dengan uji Duncan.
3.3.3 Uji Enzimatik
enzim yang akan diuji yaitu enzim selulase, protease, dan pektinase. Uji aktivitas enzim selulase dan pektinase dilakukan dengan metode Teather dan Wood (1982), sedangkan aktivitas enzim protease menggunakan metode Dunne et al. (1997). Pengamatan yang dilakukan ialah: adanya zona bening (hallo) pada media. Bakteri yang menunjukan respon terhadap media selanjutnya akan ditandai, dan diukur diameter zona beningnya.
Penelitian dilakukan dengan menggunakan Rancangan Acak Lengkap. Perlakuannya ialah 12 bakteri hasil isolasi, dan diulang sebanyak 3 kali. Data yang diperoleh diolah menggunakan perangkat statistik SAS dan dilanjutkan dengan uji Duncan.
3.3.4 Uji Antagonis
Uji antagonis dilakukan secara in vitro untuk melihat adanya sifat antagonis dari bakteri hasil isolasi terhadap jenis-jenis fungi patogen tular tanah. Patogen yang digunakan ialah Rhizoctonia sp. Ganoderma sp. dan Sclerotium sp. Patogen-patogen tersebut diperoleh dari laboratorium Penyakit Hutan, Fakultas Kehutanan IPB.
Pengujian dilakukan sesuai dengan metode Varese et al. (1996). Patogen ditumbuhkan pada cawan Petri yang berisi media PDA. Koloni dari bakteri ditempatkan pada jarak 1.5 cm dari patogen. Cawan Petri direkatkan dengan parafilm dan diinkubasikan dalam keadaan gelap pada suhu 250 C dengan durasi waktu hingga penuhnya kontrol pada media PDA.
Pengamatan dilakukan dengan mengukur pertumbuhan fungi yang diberi perlakuan secara radial dibandingkan dengan kontrol. Data pengamatan digunakan untuk menghitung perkembangan radial miselium dan persen penghambatan bakteri terhadap patogen. Persen penghambatan dihitung dengan menggunakan rumus:
Persen penghambatan = 100 % - Persen (%) total pertumbuhan fungi. Persen total pertumbuhan fungi = Diameter perkembangan fungi X 100 %. Total diameter cawan Petri
BAB IV
HASIL
4.1 Keragaman Jenis Bakteri Asal Spora FMA
Sebanyak 12 jenis bakteri telah diisolasi dan dimurnikan dari spora
Gigaspora sp. dan Glomus sp. Jenis-jenis bakteri tersebut tersaji pada Tabel 4. Tabel 4 Jenis-jenis bakteri asal spora FMA
Spora FMA Bakteri yang ditemukan
Gigaspora sp. 1. Bacillus subtilis 2. Bacillus licheniformis 3. Bacillus cereus 4. Bacillus brevis
5. Pseudomonas diminuta 6. Bacillus laterosporus 7. Bacillus pasteurii Glomus sp. 1. Bacillus circulans
2. Proteus penneri
3. Enterobacter hormaechei 4. Bacillus firmus
5. Bacillus cereus
Jumlah bakteri yang ditemukan dari spora Gigaspora sp. sebanyak 7 jenis dan
Glomus sp. sebanyak 5 jenis. Hanya satu jenis bakteri yang dijumpai pada kedua spora FMA tersebut yaitu B. cereus. Penampilan dari bakteri-bakteri yang ditemukan dapat dilihat pada Gambar 2. Bakteri yang berasal dari genus Bacillus
sebanyak 9 jenis dan untuk genus Pseudomonas, Enterobacter dan Proteus
4.2 Uji Stimulasi Mikoriza
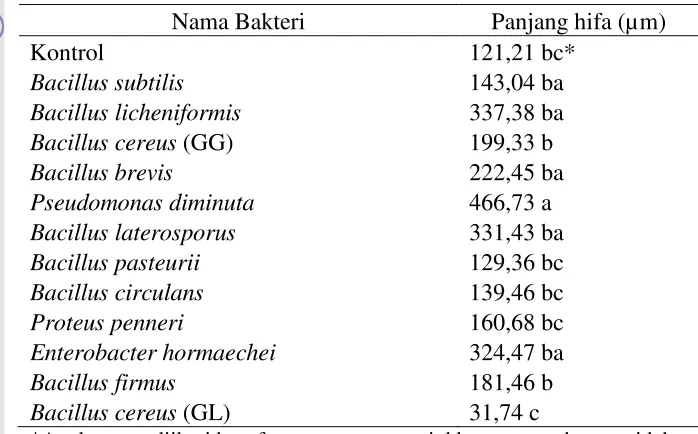
[image:32.595.111.460.178.395.2]Hasil pengujian isolat-isolat bakteri terhadap kemampuannya dalam menstimulasi perkembangan hifa spora Gigaspora sp. disajikan pada Tabel 5.
Tabel 5 Perkembangan hifa mikoriza
Nama Bakteri Panjang hifa (µm)
Kontrol 121,21 bc*
Bacillus subtilis 143,04 ba
Bacillus licheniformis 337,38 ba
Bacillus cereus (GG) 199,33 b
Bacillus brevis 222,45 ba
Pseudomonas diminuta 466,73 a
Bacillus laterosporus 331,43 ba
Bacillus pasteurii 129,36 bc
Bacillus circulans 139,46 bc
Proteus penneri 160,68 bc
Enterobacter hormaechei 324,47 ba
Bacillus firmus 181,46 b
Bacillus cereus (GL) 31,74 c
*Angka yang diikuti huruf yang sama menunjukkan pengaruh yang tidak berbeda nyata berdasarkan uji lanjut Duncan pada tingkat kesalahan 5%
Berdasarkan data pada Tabel 5, terlihat bahwa 8 jenis bakteri yaitu: P. diminuta, B. licheniformis, B. laterosporus, E. hormaechei, B. brevis,B. subtilis, B. cereus (GG), dan B. firmus mampu menstimulir perkembangan hifa mikoriza lebih tinggi dibandingkan dengan kontrol. Bakteri yang menstimulir perkembangan hifa Gigaspora sp. lebih tinggi dibanding dengan bakteri yang lain yaitu P. diminuta. Hasil pengujian statistik menunjukan bahwa 3 bakteri memberikan pengaruh yang sama dengan kontrol dan 1 bakteri yaitu B. cereus
121.21 143.04 337.38
199.
Gambar 3 Grafik per
Grafik pada menunjukkan bakteri yang berwarna hijau perkembangan hifa. S
199.33 222.45 466.73
331.43
129.36 139.46 160.68 324.47
erpanjangan hifa Gigaspora sp.
da Gambar 3 terlihat bahwa poligon be ri yang menstimulir perpanjangan hifa paling jau memperlihatkan kemampuan bakteri yan . Sedangkan poligon yang berwarna putih merup
47
181.46
31.74
Gambar 4 Perkemban
4.3 Uji Bakteri yang Dua belas isola enzim hidrolitik selul dengan adanya zona b Hasil uji enzimatik da masing bakteri disajika
Hifa
bangan hifa mikoriza Gigaspora sp.
yang Berpotensi Menghasilkan Enzim Hidroliti olat bakteri telah diuji kemampuannya dalam
lulase, protease, dan pektinase (Tabel 6). Hal ona bening (halo) di sekeliling koloni bakteri (G k dan ukuran diameter zona bening yang terben
jikan pada Gambar 5 dan 7. Spora
olitik
masing-Tabel 6 Aktivitas enzimatik (diameter halo (cm) ) bakteri yang diisolasi dari spora
Gigaspora sp. dan Glomus sp.
No Isolat bakteri
Aktivitas enzimatik (ø zona bening, cm) Selulase Protease Pektinase 1 Bacillus subtilis 4,80 cd* 4,80 ba 0,00 2 Bacillus licheniformis 2,83 e 0,00 c 0,00 3 Bacillus cereus (GG) 5,70 b 6,50 a 0,00 4 Bacillus brevis 0,00 f 5,77 a 0,00 5 Pseudomonas diminuta 5,10 cb 0,00 c 0,00 6 Bacillus laterosporus 5,87 b 6,20 a 0,00 7 Bacillus pasteurii 7,17 a 5,83 a 0,00 8 Bacillus circulans 0,00 f 0,00 c 0,00 9 Proteus penneri 5,07 cb 4,73 ba 0,00 10 Enterobacter hormaechei 2,97 e 0,00 c 0,00 11 Bacillus firmus 3,23 e 3,73 b 0,00 12 Bacillus cereus (GL) 4,10 d 4,73 ba 0,00 * Angka yang diikuti huruf yang sama menunjukkan pengaruh yang tidak berbeda nyata
berdasarkan uji lanjut Duncan pada tingkat kesalahan 5%
Ada 7 bakteri yang mempunyai potensi aktivitas enzimatik selulase dan protease yaitu: B. subtilis, B. cereus (GG), B. laterosporus, B. pasteurii, P. penneri, B. firmus, dan B. cereus (GL). Bakteri yang mempunyai aktivitas selulase tertinggi yaitu B. pasteurii, dan yang mempunyai aktivitas protease tertinggi dijumpai pada B. cereus (GG).
Bakteri yang tidak menunjukkan aktivitas selulase yaitu B. brevis dan B. circulans, untuk aktivitas protease dijumpai 8 jenis bakteri. B. circulans
[image:35.595.104.518.125.320.2]4.8
2.83 5.
Gambar 5 Grafik dia selulase
Grafik pada Gam yang terbentuk pada bakteri tertinggi terl terendah tampak pada
Gambar 6 Penampaka 5.7
0 5.1
5.87 7.17
0
5.07
2.97
Zona bening
diameter zona bening (cm) akibat aktivitas ba
ambar 5 menunjukkan perbedaan luasan diame a media akibat aktivitas bakteri penghasil sel erlihat pada poligon berwarna merah seda da poligon yang berwarna hijau.
pakan zona bening enzim selulase
3.23 4.1
bakteri penghasil
4.8
0 6.5
Gambar 7 Grafik dia protease
Grafik pada Gam yang terbentuk pada bakteri tertinggi terl terendah tampak pada aktivitas enzim protea
Gambar 8 Penampaka 6.5
5.77
0 6.2
5.83
0
4.73
0
diameter zona bening (cm) akibat aktivitas ba
ambar 7 menunjukkan perbedaan luasan diame a media akibat aktivitas bakteri penghasil prot erlihat pada poligon berwarna merah seda ada poligon yang berwarna hijau. Bakteri yang otease tampak pada poligon yang berwarna putih.
kan zona bening enzim protease Zona Bening
3.73 4.73
bakteri penghasil
4.4 Uji Antagonis terhadap Fungi Patogen
Pengujian isolat-isolat bakteri terhadap kemampuannya menghambat perkembangan hifa patogen dilakukan secara in vitro. Jenis-jenis patogen yang digunakan ialah patogen akar yang menyebabkan peyakit busuk akar dan lodoh kecambah (damping-off), yaitu Rhizoctonia sp. (Rhi), Sclerotium sp. (Scl) , dan
Ganoderma sp. (Gano). Hasil pengujian penghambatan perkembangan patogen disajikan pada Tabel 7, 8 dan 9. Perbedaan respon penghambatan patogen dari setiap pemberian bakteri tersaji pada Gambar 9, 11 dan 13. Penampakan antagonis bakteri terhadap patogen terdokumentasi pada Gambar 10, 12 dan 14.
Tabel 7 Perkembangan radial miselium dan persen penghambatan patogen
Sclerotium sp.
Isolat bakteri Perkembangan radial miselium (cm)
Persen penghambatan (%)
Kontrol 9,00 a* 0,00
Bacillus subtilis 1,70 d 81,11
Bacillus licheniformis 9,00 a 0,00
Bacillus cereus (GG) 9,00 a 0,00
Bacillus brevis 9,00 a 0,00
Pseudomonas diminuta 1,88 d 79,11
Bacillus laterosporus 9,00 a 0,00
Bacillus pasteurii 9,00 a 0,00
Bacillus circulans 9,00 a 0,00
Proteus penneri 3,00 c 66,67
Enterobacter hormaechei 3,86 b 57,11
Bacillus firmus 9,00 a 0,00
Bacillus cereus (GL) 9,00 a 0,00
[image:38.595.117.511.317.527.2]9
1.7 9
Gambar 9 Grafik panj
Poligon berwar menunjukkan empat fungi Sclerotium sp. memiliki kemampuan m
9 9
1.88
9 9
9
3
3.86
anjang radial miselia patogen fungi Sclerotium
arna kuning, hijau, merah dan ungu pada Ga pat bakteri yang memiliki kemampuan mengh p. poligon berwarna putih menunjukkan bakt puan menghambat patogen.
9 9
um sp.
Gambar 10 Penampilpilan penghambatan patogen Sclerotium sp. ole Miselium
Bakteri
Fungi
Fungi Bakteri
Fungi
p. oleh bakteri Bakteri
9
4.18
9 9
Tabel 8 Perkemban
Rhizoctoni
Isolat bakteri
Kontrol
Bacillus subtilis Bacillus licheniform Bacillus cereus (GG
Bacillus brevis Pseudomonas diminut Bacillus laterosporus Bacillus pasteurii Bacillus circulans Proteus penneri Enterobacter hormae Bacillus firmus Bacillus cereus (GL *Angka yang diikuti hur
berdasarkan uji lanju
Gambar 11 Grafik pa
9 9 4.35 9 9 6.65 4.16 4.86 bangan radial miselium dan persen pengham
onia sp.
Perkembangan radial
miselium (cm) pengh 9,00 a*
4,18 d
ormis 9,00 a
G) 9,00 a
9,00 a
inuta 4,35 d
porus 9,00 a 9,00 a 4,65 b 4,16 d
maechei 4,86 c 9,00 a
L) 9,00 a
uti huruf yang sama menunjukkan pengaruh yang tida njut Duncan pada tingkat kesalahan 5%
k panjang radial miselia patogen fungi Rhizoctoni
9 9
ambatan patogen
Persen nghambatan (%) 0,00 53,56 0,00 0,00 0,00 51,67 0,00 0,00 48,33 53,78 46,00 0,00 0,00
tidak berbeda nyata
Poligon berwarna kuni menunjukkan lima ba
[image:42.595.99.513.185.565.2]Rhizoctonia sp. Polig memiliki kemampuan m
Gambar 12 Penampil Miselium
Fungi
kuning, hijau, merah, ungu dan biru pada Ga bakteri yang memiliki kemampuan menghamb oligon yang berwarna putih menunjukkan bakt puan menghambat fungi patogen.
pilan penghambatan patogen Rhizoctonia sp. ol Bakteri
Fungi
Gambar 7 di atas bat patogen fungi bakteri yang tidak
9
1.04 9 Tabel 9 Perkemban
Ganoderm
Isolat bakteri
Kontrol
Bacillus subtilis Bacillus licheniform Bacillus cereus (GG
Bacillus brevis Pseudomonas diminut Bacillus laterosporus Bacillus pasteurii Bacillus circulans Proteus penneri Enterobacter hormae Bacillus firmus Bacillus cereus (GL *Angka yang diikuti hur
berdasarkan uji lanju
Gambar 13 Grafik
9 9 1.13 9 9 9 0.93 3.68 bangan radial miselium dan persen pengham
rma sp.
Perkembangan radial
miselium (cm) pengh 9,00 a*
1,04 c
ormis 9,00 a
G) 9,00 a
9,00 a
inuta 1,13 c
porus 9,00 a 9,00 a 9,00 a 0,93 d
maechei 3,68 b 9,00 a
L) 9,00 a
uti huruf yang sama menunjukkan pengaruh yang tida njut Duncan pada tingkat kesalahan 5%
ik panjang radial miselia patogen fungi Ganode
9 9
ambatan patogen
Persen nghambatan (%) 0,00 88,44 0,00 0,00 0,00 87,45 0,00 0,00 0,00 89,67 59,11 0,00 0,00
tidak berbeda nyata
Poligon berwar menunjukkan empat fungi Ganoderma sp. memiliki kemampuan m
Gambar 14 Penampil
Ada 4 bakteri y yang memiliki kemam diujikan. Persen peng bakteri dan antar pe pertumbuhan Sclerot hormaechei memiliki namun keduanya mem memberikan respon pe penghambatan hampi
arna kuning, hijau, merah dan ungu pada Ga pat bakteri yang memiliki kemampuan mengh
sp. poligon berwarna putih menunjukkan bakt puan menghambat patogen.
pilan penghambatan patogen Ganoderma sp. ol yaitu B. subtilis, P. diminuta, P. penneri, dan ampuan untuk menghambat pertumbuhan 3 jeni
nghambatan dari ke-4 bakteri tersebut berva perlakuan patogen. B. subtilis dan P. diminut
rotium sp. sebesar 79-80%. Bakteri P. pe
liki peran yang sama dalam penghambatan emberikan respon penghambatan yang berbeda pon penghambatan 57% lebih kecil dari P. penne
pir mencapai 67%. Bakteri
Fungi
ambar 13 di atas nghambat patogen bakteri yang tidak
p. oleh bakteri
dan E. hormaechei
jenis patogen yang variasi antar jenis
nuta menghambat
Dalam pengujian penghambatan Rhizoctonia sp. oleh bakteri yang dicobakan, 5 isolat bakteri (B. subtilis, P. diminuta, B. circulans, P. penneri, dan
E. hormaechei) menunjukan respon penghambatan. Bakteri-bakteri tersebut, kecuali B. circulans dan E. hormaechei, memberikan respon penghambatan > 50%. Bakteri B. circulans dan E. hormaechei memperlihatkan respon penghambatan < 50%. Fenomena ini terlihat dalam uji analisis statistik, ke-3 bakteri kecuali B. circulans dan E. hormaechei memberikan respon yang sama.
BAB V
PEMBAHASAN
Keberadaan bakteri yang terlibat secara langsung dalam pembentukan mikoriza pertama kali dibuktikanoleh penelitian yang dilakukan oleh Bowen dan Theodorou (1979). Pada penelitian ini 75 % jenis bakteri yang dijumpai adalah
Bacillus. Jenis yang sama dilaporkan paling banyak dijumpai pada penelitian Bakhtiar et al. (2010) yang mengisolasi bakteri dari spora FMA tanpa disterilkan dari daerah mikorizosfer kelapa sawit dan menemukan 20 isolat bakteri. Varese et al. (1996), dalam penelitiannya terkait jenis bakteri pada sporokarp Suillus grevillei berhasil menemukan 16 jenis bakteri. Tiga dari bakteri yang ditemukannya juga dijumpai pada penelitian ini, yaitu B. subtilis, B. cereus dan B. brevis.
Jenis bakteri yang dijumpai dari spora Gigaspora sp. lebih banyak dibandingkan dengan yang berasal dari Glomus sp. (Tabel 1). Hal ini kemungkinan disebabkan oleh adanya perbedaan ukuran kedua spora FMA tersebut. Ukuran spora Gigaspora sp. 100–300 µm dan Glomus sp. 20–200 µm. Dengan ukuran yang lebih besar diasumsikan bahwa bakteri yang terdapat di dalam spora Gigaspora sp. lebih banyak dibandingkan dengan Glomus sp.
Berdasarkan hasil identifikasi semua bakteri yang ditemukan memiliki ciri dapat menghasilkan endospora (Lampiran 1 dan 2). Kemampuan menghasilkan endospora memungkinkan bakteri dapat bertahan pada kondisi kering. Dengan memiliki karakteristik ini maka prospek dari bakteri-bakteri yang dijumpai untuk dikembangkan dalam bentuk inokulum bersama FMA dapat diaplikasikan.
Secara umum proses infeksi FMA pada tanaman melalui empat tahapan. Proses infeksi diawali dengan induksi perkecambahan spora dan pertumbuhan hifa, kontak antara hifa dan permukaan akar yang menyebabkan pengenalan dan pembentukan apresorium, penetrasi hifa ke dalam akar dan perkembangan struktur arbuskula internal yang selanjutnya terjadi simbiosis yang fungsional. Sebelum terbentuk kolonisasi, tumbuhan mengeluarkan sinyal dalam bentuk senyawa-senyawa tertentu untuk mengundang mikoriza mendekat dan mengkolonisasi akar tanaman.
Bakteri berperan membantu mikoriza melalui dua cara yaitu secara langsung dan tidak langsung. Aktivitas bakteri secara langsung bagi FMA terlihat ketika bakteri turut serta aktif membantu pertumbuhan dan perkembangan FMA sedangkan bantuan tidak langsung dengan menciptakan kondisi lingkungan yang nyaman sehingga mikoriza dapat menjalankan siklus hidupnya dengan baik.
Bakteri dengan kemampuan yang dapat membantu mikoriza ini dikenal dengan sebutan MHB. Garbaye dan Bowen (1989), menyatakan bahwa bakteri dapat benar-benar disebut sebagai MHB jika bakteri tersebut hidup dan berasosiasi dengan mikoriza di bagian dalam atau luar tubuh fungi tersebut (spora, hifa, vesikel, arbuskula dan bagian tubuh yang lainnya) dan menstimulir perkembangan mikoriza. Selain itu beberapa bakteri MHB dapat pula menghasilkan enzim-enzim hidrolitik dan menghambat pertumbuhan patogen.
Berdasarkan hasil penelitian terdapat 8 jenis bakteri yaitu: P. diminuta, B. licheniformis, B. laterosporus, E. hormaechei, B. brevis, B. subtilis, B. cereus
menunjukkan bahwa beberapa isolat bakteri dapat meningkatkan dan sebagian lainnya menghambat kolonisasi akar Pinus radiata oleh Rhizopogon luteolus.
Proses pengenalan antara tanaman inang dengan fungi mikoriza melibatkan penerimaan sinyal tanaman oleh miselium fungi, pertumbuhan memanjang hifa kemotropik ke arah situs yang memiliki prospek infeksi, dan perubahan karakteristik dalam morfologi miselium dan hifa. Xie et al. (1995) menunjukkan bahwa MHB kemungkinan dapat meningkatkan produksi sinyal perangsangan yang mengarahkan pertumbuhan miselium menuju akar.
Meyer dan Linderman (1986), melaporkan bahwa bakteri rizosfer
Pseudomonas putida dapat meningkatkan infeksi mikoriza dan pertumbuhan tanaman semanggi. Hasil penelitian Von alten et al. (1993) menunjukan bahwa
Bacillus subtilis dapat merangsang pembentukan spora dari FMA. Pseudomonas
sp. strain F113 dilaporkan memberikan stimulasi yang nyata pada perkembangan miselia dari spora Glomus mosseae dan terlibat dalam proses pembentukan asosiasi mikoriza dalam tanah (Barea 1998).
Telah diketahui dengan baik bahwa bakteri mikorizosfer, termasuk MHB, dapat meningkatkan nutrisi tanaman, dan bahwa kandungan P dan N tanah mempengaruhi besarnya tanggap tanaman terhadap beberapa inokulasi mikroba (Barea et al. 2005; Morgan et al. 2005). Barea dan kawan-kawan telah menemukan rhizobakteria yang meningkatkan pembentukan FMA dan meningkatkan kelarutan P dari batuan fosfat (Toro et al. 1997). Inokulasi bawang dengan Enterobacter sp., Bacillus subtilis dan FMA memiliki pengaruh positif terhadap pertumbuhan tanaman, status P dan N. B. subtilis pada khususnya adalah yang paling effektif. Inokulasi isolat bakteri ini secara nyata meningkatkan simbiosis FMA, berat kering pucuk, kandungan P pucuk, dan kandungan N pucuk. Hal ini menunjukan bahwa MHB berperan penting dalam penyerapan nutrisi dan pertumbuhan tanaman.
Bowen 1989; Garbaye dan Duponnois 1992; Gryndler dan Vosatka 1996). Berdasarkan hasil pengujian stimulasi perkembangan mikoriza, 8 bakteri yang dijumpai mempunyai potensi sebagai MHB.
Sebagai bagian dari pertahanan inang, dinding sel akar tanaman tersusun dari komponen-komponen selulosa, protein dan lignin yang menyebabkan mikroorganisme sulit menembus jaringan akar. Untuk dapat menembus jaringan akar tanaman, selain secara mekanis FMA membutuhkan enzim hidrolitik dalam konsentrasi rendah untuk melunakkan jaringan akar. Ketersediaan enzim hidrolitik seperti selulase dan protease sangat berguna di awal proses kolonisasi.
Pengujian lebih lanjut terhadap bakteri-bakteri yang ditemukan menunjukan bahwa terdapat 7 bakteri yang mempunyai potensi aktivitas enzimatik selulase dan protease yaitu: B. subtilis, B. cereus (GG), B. laterosporus, B. pasteurii, P. penneri, B. firmus, dan B. cereus (GL). Dengan kemampuan menghasilkan enzim hidrolitik, ke-7 bakteri tersebut dapat membantu FMA lebih cepat mengkolonisasi tanaman inang. Kecepatan FMA mengkolonisasi tanaman inang dapat diamati dari persen kolonisasi FMA pada akar tanaman inang. Persen kolonisasi akar tinggi menunjukan FMA tersebut telah mengkolonisasi dan berkembang jauh lebih lama pada perakaran tanaman inang. Bakteri-bakteri penghasil enzim hidrolitik yang ditemukan berpotensi untuk digunakan dalam inokulum FMA guna membantu mikoriza mempercepat proses kolonisasi.
Keberadaan bakteri endosimbiotik yang mampu menghasilkan enzim hidrolitik berpotensi membantu simbiosis mikoriza dengan inang. Mosse (1962) menginformasikan bahwa beberapa bakteri yang termasuk ke dalam genus
Beberapa bakteri MHB memiliki peran sebagai pengendali patogen tular tanah. Saat ini upaya pengendalian patogen akar telah mendapat perhatian serius dibidang pertanian maupun kehutanan. Fokus perhatian utama lebih ditujukan pada kematian semai di pembibitan yang dilakukan secara konvensional. Salah satu upaya yang dilakukan adalah meningkatkan aplikasi metode pengendalian hayati termasuk penggunaan fungi ektomikoriza dan FMA dalam pengendalian penyakit hutan (Barea et al. 2005). MHB dapat digunakan untuk promosi fungi simbiotik tertentu dan sekaligus untuk menghambat fungi yang bersifat patogen bagi tanaman. Antagonisme in vitro terhadap fitopatogen telah sering kali diamati dengan MHB (Barea et al. 1998; Budi et al. 1999).
Pada penelitian ini ditemukan 4 jenis bakteri yaitu B. subtilis, P. diminuta, P. Penneri dan E. hormaechei mempunyai sifat antagonis terhadap patogen yang diujikan. Nawangsih (2006) melaporkan bahwa bakteri B. subtilis memiliki kemampuan sebagai biokontrol dalam mengendalikan penyakit layu bakteri karena meningkatkan aktivitas enzim perioksidase pada akar. Kemampuan bakteri dalam menghambat perkembangan miselium fungi patogen kemungkinan berhubungan dengan adanya aktivitas enzimatik yang dihasilkan oleh bakteri. Enzim hidrolitik yang dihasilkan oleh bakteri dapat mendegradasi didinding sel fungi patogen.
yang ditemukan memiliki prospek untuk dikembangkan sebagai biokontrol bagi fungi patogen akar.
Peran bakteri MHB penghambat patogen memiliki prospek untuk dikembangkan dalam meningkatkan kualitas produksi inokulum FMA. Proses produksi inokulum FMA saat ini masih dilakukan secara konvensional yaitu dengan menggunakan metode kultur pot terbuka. Kelebihan dari metode ini adalah mudah dalam pengerjaannya namun inokulum yang dihasilkan tidak steril dari patogen tular tanah. Inokulum yang terkontaminasi dengan patogen akan berdampak merugikan ketika diaplikasikan ke tanaman di persemaian. Dengan ditemukannya ke-4 isolat bakteri yang berpotensi menghambat patogen penyerang akar tanaman, maka bakteri-bakteri tersebut dapat diaplikasikan sewaktu memproduksi inokulum FMA sehingga kualitas inokulum dapat ditingkatkan.
Bakteri disebut MHB ketika bakteri tersebut merupakan bakteri endofit dan dapat menstimulir perkembangan mikoriza. Beberapa bakteri selain bersifat MHB dapat juga mempunyai kemampuan menghasilkan enzim hidrolitik yang berguna membantu mikoriza di awal proses kolonisasi dan memiliki sifat antagonis terhadap patogen. Bakteri dengan kemampuan menstimulir perkembangan mikoriza, memiliki aktivitas enzimatik dan mempunyai sifat antagonis terhadap patogen dapat ditetapkan sebagai MHB plus.
Berdasarkan penelitian terdapat 3 jenis bakteri Bacillus subtilis,
Pseudomonas diminuta dan Enterobacter hormaechei berpotensi menjadi MHB
BAB VI
KESIMPULAN DAN SARAN
6.1 Kesimpulan1. Ada 12 bakteri yang berhasil diisolasi dari spora FMA, 7 bakteri dari
Gigaspora sp. dan 5 bakteri dari Glomus sp.
2. Ada 8 jenis bakteri yaitu : P. diminuta, B. licheniformis, B. laterosporus, E. hormaechei, B. brevis, B. subtilis, B. cereus (GG), dan B. firmus mampu menstimulir perkembangan hifa mikoriza.
3. Ada 7 bakteri yang mempunyai potensi aktivitas enzimatik selulase dan protease yaitu: B. subtilis, B. cereus (GG), B. laterosporus, B. pasteurii, P. penneri, B. firmus, dan B. cereus (GL).
4. Ada 4 bakteri (B. subtilis, P. diminuta, P. penneri, dan E. hormaechei) yang dapat menghambat pertumbuhan patogen Rhizoctonia sp., Sclerotium sp., dan
Ganoderma sp.
6.2 Saran
DAFTAR PUSTAKA
Ahemad M, Almas Z, Md Saghir K, Mohammad O. 2009. Factors affecting the variation of microbial communities in different agro–ecosystems. Microbial Strategis for Crop Improvement. Springer. Hlm 301
Baktiar Y, Yahya S, Sumaryono W, Sinaga MS, Budi SW, Tajudin T. 2010. Isolation and identification of mycorrhizosphere bacteria and their antagonistic effects towards Ganoderma boniense in vitro. J Microbiol Indones 4(2)
Barea JM, Andrade G, Biancioto V, Dowling D, Lohkre S, Bonfante P, O’Gara F, Azcon-Aguilar C. 1998. Impact on arbuscular mycorrhiza formation on Pseudomonands strains used as inoculants for biocontrol of soil-borne fungal plant pathogens. App Envir Microbiol 64:2304-2307
Barea JM, Pozo MJ, Azcon R, Azcon-Aguilar C. 2005. Microbial co-operation in the rhizosphere. J Exp Bot 56:1761-1778
Barriuso J, Beatriz RS, Jose AL, Agustin PL, Ana GV, Francisco JGM. 2008. Ecology, genetic diversity and screening strategies of plant growth promoting rhizobacteria (PGPR). Di dalam: Plant-bacteria interactions strategies and techniques to promote plant growth. Ahmad I, Pichtel J, Hayat S, eds. Weinheim: Wiley-VCH Verlag GmbH and Co.KgaA. Hlm 1-13
Buchanan RE, Gibbons E. 1974. Bergey’s Manual of Determinative Bacteriology, 8th ed. USA: The Williams & Wilkins Co., Inc.
Bowen, Theodore. 1979. Interactions between bacteria and ectomycorrhizal fungi.
Soil Biochem 11:119–126
Bowen GB. 1993. The ecology of ectomycorrhiza formation and functioning.
Plant Soil 159:61–67
Budi SW, Causannel JP, Gianinazzi S. 1998. The biotechnology of mycorrhizas.
In “ Microbiaal interaction in Agriculture and Forestry” Vol I, N.S. Subba Rao and Y.R. Dommergues, eds. New Delhi India: Oxford and IBH Publishing co Ltd. Hlm 149-162
Budi SW, Van Tuinen D, Negrel J, Martinotti G, Gianinazzi S. 1998. Isolation from the mycorrhizosphere of sorghum bicolor of a bacterioum compatible with arbuscular mycorrhizal development and antagonist against soil borne fungal pathogen. Di dalam: Gianinazzi S, Schueep H, eds. Arbuscular Mycorrhizas in Sustainable Soil-Plant Systems. Luxemburg: European Commision. Hlm 138-139
lands”, S. Scannerini, A. Baker, B.V. Charlwood, C. Domiano, C. fraanz and S. Giaaninazzi, eds. Acta Horticulturae 457:71-75
Budi SW, Blal B, Gianinazzi S. 1999. Surface sterilization of Glomus mosseae
sporocarpes for studying endomycorrhization in vitro. J of Mycorrhizae 8: 15-18
Budiman S. 2000. Pengaruh cendawan VAM dan kompos kasting terhadap pertumbuhan semai Duabanga moluccana BL. pada tanah bekas tambang emas. [skripsi]. Bogor: Jurusan Manajemen Hutan, Fakultas Kehutanan IPB Brulé C, Frey-Klett P, Pierrat JC, Courrier S, Gérard F, Lemoine MC, Rousselet
JL, Sommer J, Garbaye J. 2001. Survival in the soil of the ectomycorrhizal fungus Laccaria bicolor and effect of a mycorrhiza helper Pseudomonas fluorescens. Soil Biol Biochem 33: 1683–1694
Cappuccino JG, Sherman N. 2005. Microbiology : A Laboratory Manual, 5nd ed., USA: The Benjamin/Cummings Publ. Co.,Inc.
Dahm H, Wrótniak W, Strzelczyk E, Li C-Y, Bednarska E. 2005. Diversity of culturable bacteria associated with fruiting bodies of ectomycorrhizal fungi.
Phytopathol 38:51-62
Dunne C, Crowly JJ, Moenne-Loccoz Y, Dowling DN, FJ de Bruijn, O’Gara F. 1997. Biological control of Pythium ultinum by Stenotromonas maltophilia
W18 is mediated by extra cellular proteolytic activity. Microbiology
143:3921-3939
Duponnois R, Garbaye J. 1990. Some mechanisms involved in growth stimulation of ectomycorrhizal fungi by bacteria. Can J Bot 68:2148–2152
Foster, Marks. 1966. The Fine Structure of The Mycorrhizas of Pinus Radiata.
Australian J Biol Sci 19:1027-1038
Frey-Klet, Garbaye. 2005. Mychorrhiza helper bacteria: a Promising Model for the Genomic Analysis of Fungal – Bacterial Interactions. New Phytol 168:4-8
Gamalero E, Fracchia L, Cavaletto M, Garbaye J, Frey-Klett P, Varese GC, Martinotti MG. 2003. Characterization of functional traits of two fluorescent pseudomonads isolated from basidiomes of ectomycorrhizal fungi. Soil Biol Biochem 35:55–65
Gamalero E, Trotta A, Massa N, Copetta A, Martinotti MG, Berta G. 2004. Impact of two fluorescent pseudomonads and an arbuscular mycorrhizal fungus on tomato plant growth, root architecture and P acquisition.
Mycorrhiza 14:229–234
Garbaye J. 1994. Helper bacteria: a new dimension to the mycorrhizal symbiosis.
New Phytol 128:197-210
Garbaye J, Bowen GD. 1989. Stimulation of mycorrhizal infection of Pinus radiata by some microorganisms associated with the mantle of ectomycorrhizas. New Phytol 112:383–388
Garbaye J, Duponnois R. 1992. Specificity and function of mycorrhization helper bacteria (MHB) associated with the Pseudotsuga menziesii-Laccaria laccata
Gardemann JW, Nicholson TH. 1963. Spores of mycorrhizal endogones species extracted from soil by wet-sieving and decanting. Trans Br Mycol Soc 46:235-244
Gryndler M, Vosatka M. 1996. The response of Glomus fistulosum-maize mycorrhiza to treatments with culture fractions from Pseudomonas putida.
Mycorrhiza 6:207–211
Hendayanto E, Hairiah K. 2007. Biologi Tanah – Landasan Pengelolaan Tanah Sehat. Yogyakarta: Pustaka Adipura
Holt JG, Krieg NR, Sneath PHA, Staley JT, Williams ST. 1994. Bergey’s Manual of Determinative Bacteriology, 9th ed. USA: The Williams & Wilkins Co., Inc.
Linderman RG. 1988. Mychorrizal Interaction With the Rhizosphere microflora-the Mycorirhizosphere effect. Phytopatology 78:366-371
Linderman RG. 1994. Role of VAM fungi in biocontrol. Di dalam: Pfleger FL Linderman RG, ed. Mycorrhizae and Plant Health. St Paul, MN: APS Press. Hlm 1-26
Lovato PS, Trouvelot AH, Gianinazzi S. 1994. Application of arbuscular mycorrhizal fungi (AMF) in orchards and ornamental plants. In: Mycorrhizas; structure, function, molecular Biology and Biotechnology, Varma, A and hock, B. eds, Verlag Heidelberg: Springer
Meyer JR, Lindermann RG. 1986. Selective influence on populations of rhizosphere or rhizoplane bacteria and actinomycetes by mycorrhizas formed by Glomus fasciculatum.Soil Biol Bioch 18:191-196
Morgan JAW, Bending GD, White PJ. 2005. Biological costs and benefits to plant–microbe interactions in the rhizosphere. J Exp Bot 56:1729–1739 Mosse B. 1962. The establishment of vesicular-arbuscular mycorrhiza under
aseptic conditions. J Gen Microbiol 27:509–520
Mulongoy K, Gianinazzi S, Roger PA, Dommergues Y. 1992. Biofertilizers: agronomic and environmental impacts and economics. In: Microbial Technology: Economic and Social Aspects, Da Silva, E,. Sasson, A and Ratledge, C. Cambrige: Cambridge university Press
Nawangsih AA. 2006. Seleksi dan karakterisasi bakteri biokontrol untuk mengendalikan penyakit layu bakteri Ralstonia solanacearum pada tomat.. [Disertasi]. Bogor: Sekolah Pascasarjana IPB
Rao NSS, Tilak KVBR, Singh CS. 1985. Synergistic effect of vesicular-arbuscular mycorrhizae and Azospirillum brasiliense on the growth of barley in pots. Soil Biol Biochem 1:121–129
Rao MB, Tangsale AM, Ghasge MS, Deshapande VV. 1998. Molecular and biotechnological aspects of microbial protease. Microb Mol Biol Rev
62:597-635
Schrey SD, Schellhammer M, Ecke M, Hampp R, Tarkka MT. 2005. Mycorrhiza helper bacterium Streptomyces AcH 505 induces differential gene expression in the ectomycorrhizal fungus Amanita muscaria. New Phytol
Siddiqui AZ, Pichtel. 2008. Mycorrhizae: an Overview, Mychorrhizae: Sustainable Agriculture and Forestry. Verlag Berlin Heidelberg: Springer Teather RM, Wood JP. 1982. Use of Congo red polysacharide interaction in
enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol 43:777-780
Toro M, Azcon R, Barea JM. 1997. Improvement of arbuscular mycorrhiza development by inoculation of soil with phosphate- solubilizing rhizobacteria to improve rock phosphate bioavailability (32P) and nutrient cycling. Appl Environ Microbiol 63:4408–4412
Varese GC, Portinaro S, Trotta A, Scannerini S, Luppi-Mosca AM, Martinotti MA.