INDUKSI PEMBUNGAAN DI LUAR MUSIM PADA
TANAMAN JERUK KEPROK (
Citrus reticulata
)
MUHAMMAD DARMAWAN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
iii
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis “Induksi Pembungaan di Luar Musim pada Tanaman Jeruk Keprok (Citrus reticulata)” adalah karya saya
dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2014
Muhammad Darmawan
RINGKASAN
M. DARMAWAN. Induksi Pembungaan di Luar Musim pada Tanaman Jeruk Keprok (Citrus reticulata). Dibimbing oleh ROEDHY POERWANTO dan SLAMET SUSANTO.
Tujuan penelitian ini adalah (1) mempelajari efektivitas perlakuan induktor pembungaan (Prohexadion-Ca, Paclobutrazol, dan strangulasi) terhadap pembungaan tanaman jeruk keprok, (2) memperlajari efektivitas perlakuan zat pemecah dormansi (etephon, BAP, dan KNO3) terhadap pembungaan tanaman jeruk keprok, (3) mempelajari pengaruh interaksi antara perlakuan induktor pembungaan dan zat pemecah dormansi pada pembungaan tanaman jeruk keprok.
Penelitian ini dilakukan di kebun jeruk petani di Desa Tangkil, Kecamatan Caringin Kabupaten Bogor yang berlangsung dari bulan November 2012 sampai Juli 2013. Percobaan ini menggunakan rancangan blok terpisah (Split Block Design) yang terdiri dari dua faktor.Faktor pertama adalah induksi pembungaan yang terdiri atas empat taraf yaitu kontrol, Prohexadion Ca, Paclobutrazol, dan strangulasi, serta faktor kedua adalah pemberian zat pemecah dormansi yang terdiri atas empat taraf, yaitu kontrol, etepon, BAP dan KNO3. Terdapat 16 kombinasi perlakuan dan empat ulangan sehingga akan digunakan 64 tanaman sebagai satuan percobaan.
Tanaman yang digunakan dalam penelitian ini adalah tanaman jeruk yang berumur 5 tahun. Perlakuan induksi pembungaan, yaitu Prohexadion-Ca diaplikasikan pada daun sebanyak dua kali yaitu setelah trubus menjadi dewasa dan satu minggu setelah aplikasi pertama dengan konsentrasi 500 ppm/pohon dengan volume semprot 1 liter larutan. Paclobutrazol diaplikasikan pada tanah sebanyak satu kali setelah trubus menjadi dewasa dengan dosis 2 g/pohon dalam 1 liter larutan. Stragulasi dilakukan dengan pelilitan kawat berdiameter 2 mm pada pangkal pohon dan dibuka pada 14 hari setelah pelilitan. Kawat dililitkan pada ketinggian 30 cm pada pangkal pohon. Pelilitan kawat dilakukan setelah trubus
menjadi dewasa. Pemberian zat pemecah dormansi dilakukan pada 45 HSA (Hari Setelah Aplikasi) induksi pembungaan. Zat pemecah dormansi yang diberikan adalah pemberian etephon dengan konsentrasi 200 ppm/pohon, BAP 200 ppm/pohon, dan KNO3 40 g/ pohon dalam 1 liter larutan.
Hasil penelitian menunjukkan perlakuan penginduksi pembungaan (Prohexadion-Ca, Paclobutrazol, strangulasi) berpengaruh nyata dalam mempercepat pembungaan dan meningkatkan jumlah bunga dan buah tanaman jeruk. Perlakuan Prohexadion-Ca dapat mempercepat pembungaan yang lebih baik dari pada Paclobutrazol. Paclobutrazol, Prohexadion-Ca dan strangulasi meningkatkan jumlah buah dengan meningkatkan C/N rasio di daun. Perlakuan zat pemecah dormansi tidak memberikan pengaruh yang nyata pada percepatan pembungaan dan peningkatan jumlah bunga dan buah.
v
SUMMARY
M. DARMAWAN. The Off-Season Flowering Induction of „Keprok‟ Citrus Tree (Citrus reticulata). Supervised by ROEDHY POERWANTO and SLAMET SUSANTO.
The purposes of this study were to study (1) the effectiveness of flowering induction treatments (Prohexadion-Ca, Paclobutrazol, and strangulation) toward Keprok, (2) the effectiveness of dormancy-breaking substance treatments (etephon, BAP, and KNO3) on the flowering of Keprok, and (3) the interaction effect between flowering induction treatment and dormancy-breaking substances on the flowering of Keprok.
This research was conducted at orange farm in Tangkil, Caringin Bogor Sub-district which lasted from November 2012 until July 2013. This experiment used a separated block design (Split Block Design) which consisted of two factors. The first factor is the flowering induction consists of four levels i.e. control, Prohexadion-Ca, Paclobutrazol, and strangulation. Whereas, the second factor was the provision of dormancy -breaking substances consisted of four levels i.e. control, etephon, BAP and KNO3. From treatment combination, there were 16 trial units and 4 replications. As a result 64 plants were used.
Young citrus tres used in this research have similarity of age (5 years). Flowering induction treatment, namely, Prohexadion-Ca, was applied to the leaves twice. The first application after flush getting mature and the second was one week after the first application was done, with a concentration of 500 ppm/ plant in a liter of water. Paclobutrazol was applied to the soil once after flush getting mature with of 2 g/plant in a liter of water. Strangulation was done by a wire twist with diameter of 2 mm. The wire was twisted at a height of 30 cm. The wire twist was made after trubus getting mature. Dormancy-breaking substance treatment was applied 45 (HSA) flowering induction. Each dormancy-breaking substance was in liter of water added with 200 ppm ethepon, 200 ppm BAP, and 40 g KNO3 for each tree.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
vii
INDUKSI PEMBUNGAAN DI LUAR MUSIM PADA
TANAMAN JERUK KEPROK (
Citrus reticulata
)
MUHAMMAD DARMAWAN
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Agronomi dan Hortikultura
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ix
Judul Tesis : Induksi Pembungaan di Luar Musim pada Tanaman Jeruk Keprok (Citrus reticulata)
Nama : Muhammad Darmawan NIM : A252110231
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Roedhy Poerwanto, MSc Ketua
Prof Dr Ir Slamet Susanto, MSc Anggota
Diketahui oleh
Ketua Program Studi Agronomi dan Hortikultura
Dr Ir Maya Melati, MS, MSc Tanggal Ujian: 11 Maret 2014
Dekan Sekolah Pascasarjana
PRAKATA
Puji syukur penulis panjatkan kehadirat Allah SWT atas rahmat dan hidayah-Nya, sehingga penulis dapat menyelesaikan penulisan Tesis ini. Tesis yang disusun ini berjudul Induksi Pembungaan di Luar Musim pada Tanaman Jeruk Keprok (Citrus reticulta). Tesis ini merupakan salah satu syarat untuk mendapatkan gelar magister di Program Studi Agronomi dan Hortikultura, Sekolah Pascasarjana, Institut Pertanian Bogor.
Terima kasih Penulis sampaikan kepada Prof. Dr. Ir. Roedhy Poerwanto, M.Sc dan Prof. Dr. Ir. Slamet Susanto, M.Sc selaku pembimbing, serta Dr. Ir. Maya Melati, MS, M.Sc selaku ketua Program Studi Angronomi dan Hortikultura. Terima kasih pula Penulis sampaikan kepada para dosen Program Studi Agronomi dan Hortikultura, teman-teman mahasiswa pascasarjana Program Studi Agronomi dan Hortikultura dan teman-teman mahasiswa pascasarjana Sulawesi Selatan yang telah banyak memberi saran. Ungkapan terima kasih juga disampaikan kepada (alm) ayah, ibu, kakak dan seluruh keluarga besar atas segala doa dan kasih sayangnya selama ini. Penulis juga menyampaikan terima kasih Direktorat Jenderal Pendidikan Tinggi (DIKTI) atas pembiayaan biaya kuliah pada program Beasiswa Unggulan (BU) On Going. Penulis juga menyampaikan terima kasih atas pembiayaan penelitian dalam tesis ini melalui program Sistem Inovasi Nasional (SINAS) dengan No kontrak 38/SEK/INSINAS/PPK/I/2013 tanggal 14 Januari 2013 yang diketuai oleh Prof. Dr. Ir. Roedhy Poerwanto, M.Sc.
Semoga tulisan dan penelitian ini dapat memberi manfaat bagi dunia Pertanian dan pihak lain yang membutuhkannya.
Bogor, Mei 2014
xi
DAFTAR ISI
DAFTAR TABEL ix
DAFTAR GAMBAR ix
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis 3
2 TINJAUAN PUSTAKA 3
Taksonomi dan Morfologi Tanaman Jeruk 3
Syarat Tumbuh 3
Fisiologi Pembungaan 4
Pengaruh Prohexadion-Ca dalam Pembungaan 5
Pengaruh Paclobutrazol dalam Pembungaan 6
Strangulasi 7
Etephon 8
BAP (6-Benzyl Amino Purin) 8
Kalium Nitrat (KNO3) 9
3 METODE 10
Tempat dan Waktu Penelitian 10
Bahan dan Alat 10
Rancangan Percobaan 10
Pelaksanaan Penelitian 11
4 HASIL DAN PEMBAHASAN 15
Pertumbuhan Vegetatif 15
Pertumbuhan Generatif 20
Kandungan Karbohidrat, Nitrogen, dan Rasio C/N 24
Kualitas Buah 25
5 SIMPULAN DAN SARAN 27
Simpulan 27
Saran 27
DAFTAR PUSTAKA 28
LAMPIRAN 32
DAFTAR TABEL
1 Jumlah tunas per cabang tanaman jeruk keprok pada perlakuan induktor
pembungaan dan zat pemecah dormansi 15
2 Panjang per tunas tanaman jeruk keprok pada perlakuan induktor
pembungaan dan zat pemecah dormansi 18
3 Jumlah daun tanaman jeruk keprok pada perlakuan induktor pembungaan
dan zat pemecah dormansi 18
4 Tingkat kehijauan daun, luas daun dan kandungan klorofil daun pada perlakuan induktor pembungaan dan zat pemecah dormansi 19 5 Bunga pertama muncul, total bunga, fruit set, total buah pada perlakuan
induktor pembungaan dan zat pemecah dormansi 20
6 Gugur buah, jumlah buah panen, dan bobot panen pada perlakuan
induktor pembungaan dan zat pemecah dormansi 21
7 Pengaruh interaksi antara perlakuan induktor pembungaan dan zat pemecah dormansi terhadap kandungan karbohidrat, nitrogen, dan rasio
C/N daun. 25
8 Bobot buah, berat biji, berat daging buah, berat kulit, berat jus pada perlakuan induktor pembungaan dan zat pemecah dormansi 26 9 Edible portion, juice portion, padatan terlarut total, asam tertitrasi pada
perlakuan induktor pembungaan dan zat pemecah dormansi 27
DAFTAR GAMBAR
1 Perbandingan perlakuan faktor induktor pembungaan terhadap jumlah
bunga yang muncul 20
2 Perbandingan perlakuan faktor induktor pembungaan terhadap total buah
yang terbentuk 22
DAFTAR LAMPIRAN
1 Alur pelaksanaan penelitian di lapangan 32
2 Dena pengacakan percobaan di lahan 33
1
PENDAHULUAN
Latar Belakang
Jeruk merupakan salah satu komoditi tanaman buah-buahan yang tumbuh dan berkembang dengan baik di Indonesia. Buah jeruk sangat digemari oleh masyarakat karena rasanya enak, segar, dan mengandung vitamin C. Peluang pasar komoditi jeruk di dalam negeri terbuka cukup luas yang disebabkan oleh meningkatnya pertumbuhan penduduk, peningkatan pendapatan, dan kesadaran masyarakat akan pentingnya gizi di samping berkembangnya agroindustri. Saat ini Indonesia adalah negara pengimpor jeruk terbesar kedua di ASEAN setelah Malaysia. Selama kurun waktu 2005 sampai 2010 impor mencapai 550 809 ton per tahun dengan nilai mencapai US $ 650 128 774 selama lima tahun (BPS 2011).
Produksi jeruk bersifat musiman karena tanaman jeruk hanya dapat berbuah dalam beberapa bulan saja setiap tahunnya. Sifat ini tidak menguntungkan karena pada musim panen pasokan buah jeruk berlebih. Pasokan buah jeruk impor pada saat tidak musim buah menyebabkan terjadi yang stabilitas harga jual buah jeruk lokal di pasaran tidak terjamin. Untuk mengatasi permasalahan tersebut diperlukan teknik budidaya yang dapat mengatur produksi buah di luar musim, sehingga pasokan buah jeruk diharapkan tersedia sepanjang tahun.
Aktivitas fisiologi yang berperan dalam mempengaruhi perubahan pembungaan antara lain kandungan karbohidrat, kandungan nitrogen, dan rasio C/N yang terdapat dalam tanaman (Vemmos 1995). Berbagai upaya perlu dilakukan untuk meningkatkan pembungaan dan produksi antara lain rekayasa terhadap tanaman dan lingkungan tumbuhnya. Langkah-langkah yang diperlukan untuk induksi pembungaan seperti pengaturan suhu (Poerwanto et al. 1994), stress air (Susanto et al. 1993), strangulasi (Thamrin 2008, Yamanishi et al. 1993), dan aplikasi zat pengatur tumbuh (Efendi 1994, Poerwanto dan Susanto 1996).
Zat pengatur tumbuh yang komersil dan banyak digunakan untuk produksi buah di luar musim adalah Paclobutrazol. Paclobutrazol menghambat biosintesis giberelin sehingga dapat mengalihkan pertumbuhan vegetatif menuju ke pertumbuhan reproduktif. Paclobutrazol dapat menginduksi produksi buah di luar musim pada tanaman mangga (Efendi 1994, Susanto dan Poerwanto 1999), jeruk (Poerwanto dan Inoue 1994), dan manggis (Rai et al. 2004). Penggunaan Paclobutrazol untuk menginduksi pembungaan di luar musim sangat efektif tetapi Paclobutrazol meninggalkan residu pada tanah. Residu Paclobutrazol pada perkebunan mangga bisa sampai 2 tahun yang mengakibatkan pertumbuhan vegetatif tanaman terhambat dan mengganggu lingkungan (data tidak dipublikasikan). Diduga penggunaan Paclobutrazol secara berlebihan akan mengakibatkan terjadinya pencemaran air dan tanah sehingga pertumbuhan tanaman akan terganggu (Adiel et al. 2011). Terhambatnya pertumbuhan vegetatif oleh residu Paclobutrazol akan mengganggu produksi pada tahun-tahun berikutnya.
biosintesis giberelin. Prohexadion-Ca adalah bahan kimia yang dapat dimetabolisme dengan cepat dalam jaringan tanaman (Evans et al. 1999) dan sangat efektif dalam menghambat pertumbuhan tunas tanaman apel (Medjdoub dan Blanco 2003). Prohexadione-Ca saat ini digunakan untuk menekan pertumbuhan vegetatif dari buah pohon pome dan juga mengontrol tinggi tanaman dipohon buah-buahan lainnya, sayuran dan biji-bijian (Kofidis et al. 2008). Hasil penelitian Adiel et al. (2011) menunjukan bahwa Prohexadion-Ca dapat menginduksi pembungaan pada mangga. Prohexadion-Ca efektif dalam menekan pertumbuhan vegetatif, meningkatkan jumlah pembungaan, dan meningkatkan hasil dan kualitas buah pada rasberry (Poledica et al. 2012). Aplikasi Prohexadion-Ca juga telah efektif digunakan untuk mempercepat pembungaan pada tanaman apel (Owens dan Stiver 1999), ceri (Elfving et al. 2003), dan mangga (Adiel et al. 2011). Prohexadione-Ca adalah pengatur tumbuh yang relatif baru, dengan efek residu pendek yang berlangsung hanya beberapa minggu (Adil
et al. 2011). Dengan demikian tidak seperti Paclobutrazol, penggunaan Prohexadion-Ca dapat menghambat pertumbuhan tanaman vegetatif dan menginduksi bunga tanpa menyebabkan resiko pada tanah.
Pengaturan pembungaan dapat pula dilakukan secara fisik yaitu dengan strangulasi. Susanto et al. (2002) menyatakan bahwa strangulasi pada tanaman jeruk pamelo mampu menginduksi tanaman untuk berbunga dan membentuk buah. Penelitian Putra (2002) menyatakan bahwa strangulasi batang utama dengan penggunaan kawat 2.0 mm dalam waktu tiga bulan mampu meningkatkan pembungaan jeruk pamelo „Nambangan‟. Hasil penelitian lainnya menunjukan bahwa strangulasi pada tanaman jeruk pamelo menghambat translokasi fotosintat dari tajuk ke akar sehingga terjadi peningkatan akumulasi karbohidrat di bagian tajuk yang akan merangsang tanaman jeruk untuk berbunga dan membentuk buah (Susanto 2002, Yamanishi et al. 1995).
Aplikasi Paclobutrazol pada tanaman mangga menyebabkan mata tunas menjadi dorman dan pecah tunas akan terjadi setelah beberapa bulan aplikasi Paclobutrazol. Pemberian etephon, BAP, atau KNO3 dapat mempercepat pecah tunas dan pembentukan bunga (Poerwanto et al. 1995). Etephon merupakan salah satu zat pengatur tumbuh sintetik yang mampu mengatasi dormansi tunas generatif, antara lain pada mangga dan jeruk (Syahbudin 1999). BAP adalah salah satu sitokinin sintetik yang dapat mendorong pembelahan sel, morfogenesis, pertunasan, pembelahan kloroplas, serta menghambat senesen dan absisi. KNO3 dapat menyerempakkan pecah tunas pada tanaman mangga (Efendi 1994).
Tujuan Penelitian
1 Mempelajari efektivitas perlakuan induktor pembungaan (Prohexadion-Ca, Paclobutrazol, dan strangulasi) terhadap pembungaan tanaman jeruk keprok 2 Mempelajari efektivitas perlakuan zat pemecah dormansi (etephon, BAP, dan
KNO3) terhadap pembungaan tanaman jeruk keprok
3
Hipotesis
1. Terdapat satu atau lebih perlakuan penginduktor pembungaan yang mempercepat pembungaan tanaman jeruk keprok.
2. Terdapat salah satu atau lebih zat pemecah dormansi yang dapat memecah dormansi pada tanaman jeruk keprok.
3. Terdapat interaksi antara perlakuan induktor pembungaan dan zat pemecah dormansi pada pembungaan tanaman jeruk keprok.
2
TINJAUAN PUSTAKA
Taksonomi dan Morfologi Tanaman Jeruk
Jeruk merupakan tanaman yang termasuk dalam famili Rutaceae, sub famili
Aurantioideae. Famili Rutaceae terdiri dari 130 genus dan yang paling banyak dibudidayakan adalah genus Citrus. Genus Citrus terdiri dari dua sub genus yaitu
Papeda dan Eucitrus. Papeda memiliki tangkainya panjang, pangkal daun bersayap, dan buahnya tidak dapat dimakan karena adanya tetes-tetes minyak pedas yang banyak dalam kantong juice, sedangkan Eucitrus merupakan jenis tanaman jeruk yang paling banyak dan paling luas dibudidayakan karena buahnya enak untuk dimakan. Jeruk keprok (Citrus reticulata) merupakan salah satu spesies yang termasuk di dalam subgenus Eucitrus (Samson 1980).
Genus Citrus pada umumnya berupa pohon atau perdu, posisi daun berhadap-hadapan atau berseling, bentuk daun bisa berupa daun tunggal atau majemuk. Jeruk berdaun menyirip dan beranak daun satu (unifoliatus) dan dahan yang lebih dewasa biasanya tidak berduri. Pohon jeruk mempunyai akar tunggang dan mempunyai beberapa rambut akar. Bunga tumbuh pada ketiak daun, tunggal atau dalam rangkaian. Bunga jeruk berbau sangat harum bila membuka penuh, kelopak berbentuk segi tiga yang runcing atau tumpul, dan berbulu halus. Dari luar bunga jeruk berwarna putih hijau kekuningan atau putih kekuningan dengan bintik kelenjar kuning muda sedangkan dari dalam berwarna putih. Buah merupakan jenis beri yang spesial yang disebut hesperidium, lapisan luar kaku dan mengandung banyak kelenjar minyak atsiri. Kulit buah mula-mula berwarna hijau tetapi setelah masak warna berubah menjadi kuning atau jingga. Lapisan kulit ini disebut flavedo. Lapisan tengah bersifat seperti spon, terdiri atas jaringan bunga karang yang biasanya berwarna putih yang disebut dengan albedo. Biji berisi satu atau lebih embrio (poliembrioni) (Harjadi 1996).
Syarat Tumbuh
Tanaman jeruk tumbuh baik pada tipe tanah yang berdrainase baik, memiliki porositas yang baik, dan bersolum dalam seperti jenis tanah aluvial. Kisaran pH yang optimal untuk pertumbuhan jeruk adalah 5-8. Air diperlukan untuk pertumbuhan, pembungaan, dan produksi jeruk, namun jeruk peka terhadap kondisi tergenang dan kelembaban yang tinggi karena dapat menimbulkan penyakit (Samson 1980).
Fisiologi Pembungaan
Pohon buah mengalami siklus tumbuh dari embrio (dalam biji), kecambah, yuwana (juvenil) dan dewasa. Tanaman akan mengalami perubahan secara perlahan-lahan dari masa muda ke masa dewasa. Masa muda tanaman ditandai dengan belum adanya kemampuan untuk berbunga pada tanaman tersebut. Bagian bawah yang jauh dari akar telah menjadi dewasa, sedangkan bagian yang dekat dari akar biasanya masih muda. Pada saat tanaman mulai mampu berbunga disebut sebagai masa transisi. Masa transisi pada tanaman tidak berlangsung serentak. Transisi dari juvenil ke dewasa melalui suatu tahapan dimana bagian-bagian bawah tetap mempertahankan karakteristik juvenil sementara bagian-bagian atas pohon mulai berbunga (Poerwanto 2003).
Bagian batang yang berbatasan dengan akar juga tetap pada fase juvenil. Beberapa ahli fisiologi menghubungkan fenomena ini dengan produksi hormon akar yang ditranslokasikan ke atas. Perubahan peningkatan hormon ini dalam meristem apikal bertanggung jawab untuk transisi dari juvenil ke tahap dewasa. Pucuk apikal juvenil dan dewasa mempunyai tipe RNA yang berbeda dengan kode genetik suatu klon tetap. Perubahan ini mencerminkan represi dan ekspresi gen yang dibawa hormon (Poerwanto 2003)
Giberelin merupakan zat pengatur tumbuh (ZPT) yang berperan dalam proses pembungaan. Aplikasi ZPT memberikan respon yang berbeda pada setiap jenis tanaman, demikian juga dengan giberelin. Tidak semua tanaman yang diberi giberelin dapat berbunga. Goldlochmidt dan Monselise (1972) menyatakan bahwa penurunan giberelin diperlukan untuk menginduksi pembungaan pada tanaman jeruk dan tanaman berkayu lainnya. Goldschmidt dan Monselise (1972) selanjutnya juga menyatakan bahwa penghambatan pembungaan oleh giberelin mungkin merupakan pengaruh langsung yang disebabkan oleh peningkatan pertumbuhan vegetatif.
Lang (1952) menyatakan bahwa proses pembentukan bunga dibagi menjadi empat tahap. Pertama, induksi atau inisiasi bunga dan diferensiasi primordia bunga. Kedua, penyusunan/organisasi bunga dan diferensiasi bagian-bagian bunga secara individu. Ketiga, pematangan bunga dan biasanya terjadi bersamaan dengan proses pertumbuhan bagian-bagian bunga, diferensiasi jaringan sporogen, meiosis, tepung sari dan perkembangan kantung embrio. Keempat, antehis atau bunga mekar. Induksi bunga merupakan suatu tahapan ketika kuncup vegetatif dirangsang secara biokimia dan berubah menjadi pucuk reproduktif. Tidak ada perubahan morfologi yang menandakan tahap ini terjadi (Poerwanto 2003).
5
pada tanaman. Nisbah C/N yang tinggi dapat menginduksi pembungaan, sedangkan bila nisbah C/N rendah tanaman akan lebih mengarah pada pertumbuhan vegetatif. Guardiola (1981) menyatakan inisiasi bunga merupakan tahap paling selektif terhadap faktor hormon dan lingkungan, jika kondisinya tidak sesuai maka perkembangan tunas bunga menjadi terhambat.
Induksi bunga adalah fase yang paling penting dalam proses pembungaan. Pada fase ini terjadi perubahan fisiologis atau biokimia pada mata tunas dari pertumbuhan vegetatif mengarah pada pertumbuhan generatif. Induksi pembungaan merupakan fase yang penting karena perubahan hanya nampak pada kuncub bunga, sedangkan perubahan secara morfologis tidak ada. Induksi pembungaan berkaitan dengan beberapa faktor. Faktor tersebut adalah faktor eksternal, internal dan manipulasi oleh manusia. Faktor eksternal yaitu suhu, stress air, dan panjang hari; faktor internal yaitu kandungan nitrogen, karbohidrat, asam amino, dan hormon serta faktor manipulasi oleh manusia seperti girdling/ringing, pemangkasan, pengeringan, pamangkasan akar, pelengkungan cabang, dan pemberian zat pengatur tumbuh.
Aplikasi zat pengatur tumbuh yang berpotensi menurunkan aktivitas giberelin endogen diharapkan mampu meningkatkan pembungaan pada tanaman jeruk. Menurut Wattimena (1987), Paclobutrazol adalah senyawa aktif yang menghambat produksi giberelin, sehingga secara tidak langsung menyediakan sebagian besar fotosintat untuk pertumbuhan generatif. Prohexadion-Ca adalah senyawa yang dapat menyebabkan penuruan pertumbuhan dengan cara memblokir biosintesis giberelin, sehingga dapat menghambat pertumbuhan vegetatif (Adiel et al. 2011)
Mengacu pada penelitian Goldsmidt dan Monselise (1972) bahwa dalam pengaturan pembungaan digunakan zat yang dapat menekan biosintesis giberelin yang disebut sebagai zat penghambat tumbuh. Penggunaan zat penghambat tumbuh secara langsung menekan pertumbuhan vegetatif sehingga akan meningkatkan nisbah C/N dan mendorong terjadinya inisiasi tunas bunga. Penggunaan retardan dalam mengatur pembungaan kadang-kadang menyebabkan dormansi tunas, sehingga tunas-tunas yang sudah terinisiasi tidak dapat muncul dan berkembang menjadi tunas bunga. Menurut Ryugo (1988) pemecahan dormansi dari tunas-tunas bunga tersebut tergantung pada keberadaan cadangan karbohidrat pada tanaman.
Pengaruh Prohexadione-Ca dalam Pembungaan
Akhir-akhir ini telah ditemukan zat penghambat tumbuh yang mempunyai sifat fisiologi mirip dengan Paclobutrazol tetapi tidak meninggalkan residu setelah aplikasi. Zat pengatur tumbuh tersebut ialah Prohexadion-Ca. Prohexadione-Ca menyebabkan penurunan pertumbuhan dengan cara memblokir biosintesis giberelin. Prohexadione-Ca adalah zat pengatur tumbuh yang relatif baru, dengan efek residu pendek yang berlangsung hanya beberapa minggu (Adil et al. 2011).
1999) dan sangat efektif dalam menghambat pertumbuhan tunas tanaman apel (Medjdoub dan Blanco 2003). Prohexadion-Ca efektif dalam menekan pertumbuhan vegetatif, meningkatkan pembungaan, serta meningkatkan hasil dan kualitas buah pada rasberry (Poledica et al. 2012). Aplikasi Prohexadion-Ca juga telah efektif digunakan untuk mempercepat pembungaan pada tanaman apel (Owens dan Stiver 1999), ceri (Elfving et al. 2003), dan mangga (Adiel et al.
2011).
Beberapa tahun terakhir, Prohexadione-Ca telah digunakan sebagai alternatif untuk pengganti Dominozide. Dominozide menimbulkan masalah dalam pencemaran lingkungan dan penggunaannya saat ini sangat dibatasi. Prohexadione-Ca dianggap aman karena tidak memiliki efek terhadap mamalia dan berpotensi rendah untuk bioakumulasi dalam lingkungan. Prohexadione-Ca saat ini digunakan untuk menekan pertumbuhan vegetatif dari buah pohon pome dan juga mengontrol tinggi tanaman di pohon buah-buahan lainnya, sayuran, dan biji-bijian (Kofidis et al. 2008). Hasil penelitian Adiel et al. (2011) menunjukkan bahwa Prohexadion-Ca dapat menginduksi pembungaan pada mangga. Prohexadion-Ca diaplikasikan pada tanaman melalui daun. Hasil penelitian menunjukkan bahwa pemberian melalui daun lebih efektif jika dilakukan beberapa kali dengan dosis yang rendah. Prohexadion-Ca dengan dosis 0.5 g per liter air menyebabkan tanaman mangga berbunga lebih cepat 30 hari dibandingkan tanaman kontrol (Adiel et al. 2011), sedangkan dengan dosis 0.2-0.4 g per liter Prohexadion-Ca efektif menghambat pertumbuhan tanaman apel sebesar 27%-36% dibandingkan kontrol. Dengan demikian tidak seperti Paclobutrazol, penggunaan Prohexadion-Ca dapat menghambat pertumbuhan vegetatif tanaman dan induksi bunga tanpa menyebabkan residu pada tanah.
Pengaruh Paclobutrazol dalam Pembungaan
Paclobutrazol dapat diaplikasikan pada tanaman melalui daun atau tanah. Hasil penelitian menunjukkan bahwa perlakuan melalui tanah lebih efektif dibandingkan perlakuan melalui daun, sedangkan penyemprotan melalui daun akan lebih efektif jika dilakukan beberapa kali dengan dosis yang rendah (Voon
et al. 1992).
Paclobutrazol diserap oleh tanaman melalui daun, pembuluh batang, atau akar, kemudian translokasikan secara akropetal melalui xylem ke bagian tanaman yang lain. Pada meristem sub apikal senyawa ini akan menghambat biosintesis giberelin yang selanjutnya akan menyebabkan penurunan laju pembelahan sel sehingga menghambat pertumbuhan vegetatif dan secara tidak langsung akan mengalihkan fotosintat ke pertumbuhan reproduktif yang diperlukan untuk membentuk bunga dan buah (Susanto dan Poerwanto 1999)
7
Penghambatan biosintesis giberelin oleh Paclobutrazol yaitu pada proses oksidasi kauren menjadi asam kaurenat, berakibat meningkatnya biosintesis asam absisat (ABA) yaitu hormon yang berperan dalam dormansi tunas (Wattimena 1987). Terhambatnya biosintesis giberelin menyebabkan perpanjangan sel pada meristem apikal berjalan lambat sehingga menekan pertumbuhan vegetatif pada tanaman (Susanto dan Poerwanto 1999).
Pembungaan berhubungan dengan kandungan giberelin dalam tanaman. Kandungan giberelin yang tinggi akan menekan kandungan karbohidrat pada pucuk yang aktif tumbuh sehingga menghambat pembungaan. Pemberian Paclobutrazol sebagai zat penghambat tumbuh yang meghambat biosintesis giberelin dapat menginduksi pembungaan (Sach 1977)
Aplikasi Paclobutrazol secara nyata berperan menginduksi pembungaan tanaman mangga di luar musim pada tanaman. Bunga muncul pada 61-71 hari setelah aplikasi Paclobutrazol dengan persentase pembungaan 83.3%-100%, sementara pada kontrol tidak berbunga sama sekali. Selain itu, Paclobutrazol secara nyata menghambat pertumbuhan vegetatif tanaman mangga, yaitu menurunkan total tunas dan memperpendek panjang tunas. Induksi pembungaan terjadi pada 61-71 hari setelah perlakuan Paclobutrazol pada tanaman mangga (Susanto dan Poerwanto 1999). Konsentrasi pemberian Paclobutrazol dan efek penggunaan Paclobutrazol pada setiap tanaman bervariasi, hal ini dipengaruhi oleh faktor genetik dan faktor lingkungan (Voon et al. 1992). Lontoh et al.
(1989) menyatakan bahwa penggunaan Paclobutrazol memiliki efek residu pada tahun berikutnya, hal ini diperkuat oleh Lyer dan Kurian (1991) yang menyatakan bahwa, efek Paclobutrazol dapat bertahan selama 2 tahun. Dari beberapa penelitian penggunaan Paclobutrazol terlihat bahwa konsentrasi dan waktu pemberian Paclobutrazol harus tepat sehingga ada jaminan akan keberhasilan pembungaan.
Strangulasi
Thamrin (2008) menyatakan bahwa strangulasi dengan cara melilitkan kawat dengan diameter 3.0 mm pada batang utama, menekan kawat ke batang sedalam 3.0 mm, dan dilepaskan tiga bulan setelah aplikasi dapat mempercepat terbentuknya kuncup bunga, meningkatkan bunga mekar, buah terbentuk, dan
fruit set per pohon, jika dibandingkan dengan tanpa perlakuan strangulasi. Strangulasi dapat meningkatkan kandungan karbohidrat daun dan nisbah C/N dibandingkan tanaman kontrol, tetapi kandungan nitrogen pada daun akan menurun dibandingkan kontrol. Tanaman yang diberi perlakuan strangulasi memiliki panjang tunas yang lebih pendek dan memiliki jumlah tunas yang lebih sedikit dibandingkan dengan tanpa perlakuan strangulasi. Tanaman yang diberikan perlakuan strangulasi dapat mempercepat pembungaan, meningkatkan jumlah kuncup bunga, dan jumlah cluster. Tanaman yang diberikan perlakuan strangulasi juga memiliki jumlah karbohidrat yang lebih lebih tinggi dibandingkan dengan tanpa perlakuan strangulasi (Ramda 2005).
Hasil percobaan pada Rambutan di Bogor menunjukan bahwa perlakuan ringing pada bulan Mei, Juni, atau Juli dapat menyebabkan pohon rambutan berbunga lebih awal daripada kontrol (Poerwanto 2001). Percobaan Poerwanto (2001) menunjukan bahwa tanaman yang tidak diperlakukan (cek) berbunga pada awal Oktober, sedangkan tanaman yang diringing berbunga mulai pertengahan Juni. Dengan mengatur waktu ringing dapat diatur pula waktu berbunga rambutan, sehingga waktu panen rambutan juga dapat diatur (Poerwanto 2003).
Etephon
Etephon (asam 2-kloroetil fosfonat) merupakan salah satu zat pengatur tumbuh sintetik yang mampu mengatasi dormansi tunas generatif antara lain pada leci (Poerwanto et al. 1997), mangga (Efendi 1994), dan jeruk keprok siem (Sostenes 1996). Fungsi etephon setelah pemberian Paclobutrazol adalah sebagai zat pemecah dormansi mata tunas generatif, karena menurut Mehouachi et al.
(1996), aplikasi Paclobutrazol juga meningkatkan biosintesis asam absisat dan hal itu menyebabkan pucuk yang telah terinduksi berbunga mengalami dormansi. Menurut Moore (1979), etephon dalam jaringan tanaman terhidrolisis menghasilkan etilen, ion klor, dan fosfat. Etilen disamping berfungsi mendorong pemecahan dormansi tunas, juga mendorong terjadinya absisi.
Etephon adalah senyawa yang secara spontan melepas etilen setelah kontak dengan air. Etephon mampu merangsang pembungaan nanas dan mempercepat pembungaan pada jeruk keprok dibandingkan dengan kontrol. Muchjajib dan Espino (1991) menambahkan bahwa aplikasi Paclobutrazol yang diberikan bersamaan dengan etephon mampu meningkatkan persentase pembungaan pada tanaman rambutan. Penelitian yang dilakukan oleh Chandraparnih et al. (1992) pada tanaman lychee yang berumur 15 tahun menunjukkan bahwa pemberian Paclobutrazol 1000 ppm (foliar sprays) yang diikuti dengan aplikasi etephon 400 ppm ternyata dapat meningkatkan pembungaan sampai 300%.
BAP (6-Benzyl Amino Purin)
9
Tidak seperti auksin dan giberelin, sitokinin hanya sedikit yang ditranslokasikan melalui jaringan yang hidup pada tanaman. Sitokinin dibawa secara pasif sepanjang aliran transpirasi pada xylem dari akar, yang merupakan sumber utama produksinya, menuju ke bagian-bagian lain dalam tanaman (Krishnamoorty 1981).
Krishnamoorty (1981) menyatakan bahwa selama pecah tunas, terjadi perubahan-perubahan dalam tunas, yaitu peningkatan pada kandungan sitokinin, asam nukleat, protein, poliamin, respirasi produksi etilen dan sebagainya. Chandraparnih et al. (1992) menambahkan bahwa total kandungan sitokinin meningkat dalam xylem 30 hari sebelum pembentukan tunas bunga dan maksimal selama pembentukan bunga dan bunga mekar (antesis). Hasil penelitian Chandraparnih et al. (1992) menunjukkan bahwa kandungan sitokinin yang tinggi berkorelasi positif dengan pembentukan bunga. Sitokinin berfungsi mendorong pemecahan tunas bunga yang mengalami dormansi. Watimmena (1987) menyatakan bahwa pemberian BAP 100 ppm dapat mempercepat pecah tunas serta meningkatkan jumlah tunas pecah. Penelitian Sostenes (1996) menyatakan bahwa pemberian zat pemecah dormansi BAP dengan dosis 100 ppm yang diaplikasikan tiga bulan setelah pemberian Paclobutrazol berpengaruh dalam meningkatkan jumlah tunas, panjang tunas, dan jumlah daun dibandingkan dengan kontrol.
Kalium Nitrat (KNO3)
Kalium nitrat (KNO3) mengandung dua unsur hara esensial yang dibutuhkan tanaman, yaitu kalium dan nitrogen. Peranan utama kalium dalam metabolisme tanaman adalah sebagai katalisator, terlibat dalam sintesis protein dari asam-asam amino, dan metabolisme karbohidrat. Ion kalium dalam sel tanaman mempunyai peranan dalam transportasi karbohidrat fotosintesis dan meningkatkan permeabilitas membran. Nitrogen dalam tanaman berperan pada sintesis asam amino saat pembentukan protein (Prahardini et al. 1989).
KNO3 mampu berperan sebagai zat pemecah dormansi pada tanaman jeruk. Penelitan yang dilakukan oleh Erez et al. (1971) membuktikan bahwa KNO3 efektif dalam memecah dormansi pada peach dan beberapa tanaman decidious. Bondan dan Linsangan (1979) menyatakan bahwa zat pemecah dormansi KNO3 dapat menginduksi pembungaan pada mangga. Lebih lanjut Subhandrabandhu dan Tongumpai (1990) menambahkan bahwa KNO3 efektif dalam memecahkan dormansi beberapa varietas mangga di Thailand. Menurut Efendi (1994), KNO3 yang diaplikasikan setelah pemberian Paclobutrazol pada mangga mampu mempercepat dan meningkatkan persen pembungaan dibandingkan perlakuan Paclobutrazol secara mandiri.
METODE
Tempat dan Waktu Penelitian
Percobaan dilakukan di kebun jeruk petani di Desa Tangkil, Kecamatan Caringin, Kabupaten Bogor yang terletak pada ketinggian 700 m dari permukaan laut dan mempunyai suhu rata-rata harian maksimum 30±2oC dan minimum 21±2oC. Penelitian ini berlangsung dari November 2012 sampai Juli 2013. Pengujian karbohidrat dan nitrogen dilakukan di Laboratorium Balai Besar Penelitian dan Pengembangan Pascapanen Pertanian, Bogor. Pengujian kandungan klorofil dilakukan di Laboratorium Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor. Pengujian kualitas buah di lakukan di Laboratorium Pasca Panen Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor.
Bahan dan Alat
Penelitian menggunakan tanaman jeruk keprok berumur 5 tahun yang memiliki kesamaan diameter batang dan ukuran tajuk, hal ini bertujuan untuk menyeragamkan kondisi tanaman. Bahan-bahan yang digunakan antara lain pupuk kandang 60 kg/tanaman, Prohexadion-Ca, Paclobutrazol, etephon, BAP, KNO3, tali, dan label. Alat-alat yang digunakan adalah tangga, alat penyiram, jangka sorong, piloks untuk menandai, ember plastik, pengaduk, kawat 2 mm untuk strangulasi, cool box, tang, dan Chlorophyll Meter (SPAD-502).
Bahan yang digunakan untuk analisis buah adalah jeruk keprok yang diperoleh dari hasil panen, fenoftalein, NaOH 0.1 N, dan aquades. Peralatan yang digunakan untuk analisis kualitas buah terdiri dari timbangan analitik, hand reftaktometer, dan alat titrasi.
Rancangan Percobaan
Percobaan ini menggunakan Rancangan Blok Terpisah (Split Block Design). Faktor pertama adalah induktor pembungaan yang terdiri atas empat taraf yaitu, kontrol (I1), aplikasi Prohexadione-Ca (I2), Paclobutrazol (I3), dan strangulasi (I4). Faktor kedua adalah aplikasi pemberian zat pemecah dormansi yang terdiri atas empat taraf yaitu, kontrol (D1), etepon (D2), BAP (D3), dan KNO3 (D4). Terdapat 16 kombinasi perlakuan dan empat ulangan sehingga digunakan 64 tanaman sebagai satuan percobaan. Data yang diperoleh diuji dengan uji DMRT (Duncan‟s Multiple Range Test) pada taraf nyata 5%.
Model linier dari rancangan blok terpisah secara umum dapat dituliskan sebagai berikut :
Yijk = µ + Kk+ αi+ ik+ j + ik+ (α )ij+ ijk Dimana :
Yijk : Nilai pengamatan pada faktor induksi taraf ke-i, faktor zat
pemecah dormansi taraf ke-j dan blok ke-k µ : Komponen aditif dari rataan
Kk : Pengaruh blok ke-k
11
j : Pengaruh utama faktor zat pemecah dormansi ke-j
(α )ij : Komponen interaksi dari faktor induksi ke-i dan faktor pemecah dormansi ke-j.
ik : Komponen acak dari faktor induksi yang menyebar normal (0,ϭδ2) ik : Komponen acak dari faktor pemecah dormansi yang menyebar
Tanaman dipilih sebanyak 64 tanaman berdasarkan ukuran tajuk, umur tanaman, dan kondisi tanaman. Setiap perlakuan diberikan label sesuai dengan jenis perlakuan. Untuk keperluan pengamatan vegetatif pada setiap percobaan dipilih 4 tunas berdasarkan arah mata angin per tanaman.
Pemupukan awal dilakukan sebelum perlakukan dasar dengan menggunakan pupuk kandang sapi dengan dosis 60 kg/pohon yang diberikan sebelum perlakuan dimulai. Pemangkasan (prunning) dilakukan sebelum perlakuan dimulai, hal ini bertujuan untuk menghilangkan cabang-cabang yang kering dan terserang oleh hama dan penyakit. Selain itu, dilakukan pembersihan gulma, penyemprotan insektisida, dan penyemprotan fungisida.
Perlakuan Penelitian
Prohexadione-Ca diaplikasi pada daun dengan konsentrasi 500 ppm/pohon dalam 1 liter air dan diaplikasikan sebanyak 2 kali. Aplikasi pertama dilakukan setelah trubus menjadi dewasa (9 Desember 2012) dan aplikasi yang kedua dilakukan pada 1 minggu setelah perlakukan pertama dilakukan (16 Desember 2012). Paclobutrazol diaplikasikan pada tanah sebanyak 1 kali dengan dosis 2 g/pohon dalam 1 liter air yang diaplikasikan ketika trubus menjadi dewasa (9 Desember 2012). Strangulasi dilakukan dengan cara melilitkan kawat berdiameter 2 mm pada pangkal pohon. Pelilitan dilakukan pada ketinggian 30 cm dari pangkal pohon. Pelilitan dilakukan sekuat-kuatnya sampai kulit batang terluka. Strangulasi diaplikasikan pada tanggal 9 Desember 2012 dan kawat di lepaskan 2 minggu setelah proses pelilitan (23 Desember 2012). Pemberian zat pemecah dormansi dilakukan pada 45 HSA (Hari Setelah Aplikasi) induksi pembungaan yaitu tanggal 23 Januari 2013. ZPT yang diberikan adalah etephon dengan konsentrasi 200 ppm/pohon dengan volume 1 liter air, BAP dengan konsentrasi 200 ppm/pohon dengan volume 1 liter air, dan KNO3 40 g/pohon dalam 1 liter air.
menggunakan metode Dan Sims (2002). Pengambilan sampel buah untuk analisis kualitas buah dilakukan pada saat buah sudah siap untuk dipanen.
Pengamatan
Pada tiap tanaman sebanyak 4 ranting contoh diberi tanda untuk pengamatan vegetatif. Ranting di luar contoh digunakan untuk sampel analisis kandungan karbohidrat dan kandungan nitrogen. Sampel daun untuk analisis kandungan karbohidrat dan nitrogen diambil dari daun yang sudah dewasa (berkembang penuh dan berwarna hijau tua). Peubah yang diamati adalah:
Pertumbuhan vegetatif 1. Jumlah tunas per cabang
Pengamatan dilakukan setelah tunas pecah dan keluar bakal daun diamati setiap minggu.
2. Panjang tunas (cm)
Pengamatan diukur dari pangkal tunas sampai pada titik tumbuh tunas terminal dan diamati setiap minggu.
3. Jumlah daun per tunas
Pengamatan dilakukan setelah tunas pecah dan diamati setiap minggu. Untuk pengolahan data, jumlah daun dikelompokan dalam selang waktu 2 minggu.
4. Luas daun contoh (cm2)
Luas daun diukur dengan menggunakan alat Leaf Area Meter. Daun yang diamati adalah daun ke-5 dari tajuk yang sudah dewasa. Pengamatan dilakukan pada akhir penelitian.
5. Kehijauan daun (unit)
Pengamatan dilakukan dengan menggunakan alat Chlorophyll Meter
(SPAD-502) Minolta, dengan mengambil daun dari tajuk yang sudah dewasa. Pengamatan tingkat kehijauan daun dilakukan pada akhir penelitian yaitu 17 Februari 2013.
6. Analisis kandungan klorofil (mg/g)
Analisis kandungan klorofil dilakukan berdasarkan metode Dan Sims (2002). Sampel daun yang diambil adalah daun ke-5 yang dihitung dari atas (daun yang paling muda). Sampel daun ditimbang dengan berat ±0.02 g. Daun tersebut dihaluskan dan ditambahkan acetris (85% aseton + 15% tris) sebanyak 1 ml. Daun yang sudah halus dimasukan ke dalam microtube 2 ml, sisa sampel dibilas dengan acetris sampai microtube penuh 2 ml. Setelah itu disentrifugasi dengan kecepatan 14 000 rpm selama 10 detik. Supernatan diambil 1 ml kemudian dimasukkan ke dalam tabung reaksi dan ditambahkan acetris 3 ml ke dalam tabung reaksi dan ditutup dengan kaleng kemudian dikocok dengan menggunakan vortex. Absorbansi diukur dengan spektrophotometer pada panjang gelombang 470 nm, 537 nm, 647 nm, dan 663 nm.
13
Pembungaan
1. Waktu bunga pertama muncul (HSAIP).
Saat pertama berbunga adalah saat pertama muncul tunas bunga pada tanaman setelah perlakuan induksi pembungaan.
2. Total bunga mekar.
Total bunga mekar adalah jumlah total bunga mekar di setiap tanaman dihitung setiap satu minggu sekali setelah perlakuan.
Buah
1. Fruit set (%)
Fruit set diamati setelah fase bunga mekar, dihitung dari jumlah buah yang terbentuk. Fruit set dihitung setiap dua minggu sekali dengan menggunakan rumus :
� � = Jumlah Bakal Buah Terbentuk
Jumlah Total Bunga Terbentuk x 100 %
2. Jumlah buah
Jumlah buah yang terbentuk dihitung pada setiap tanaman, dilakukan setiap satu minggu sekali setelah perlakuan.
3. Persentase gugur buah (%)
Persentase gugur buah dihitung di akhir penelitian. Perhitungan gugur buah dihitung dengan menggunakan rumus :
Gugur buah = Jumlah buah gugur
Jumlah buah terbentuk x 100 %
4. Jumlah buah panen
Jumlah buah panen dihitung pada akhir penelitian. 5. Bobot panen (kg)
Bobot panen dihitung pada akhir penelitian. Analisis Kimia
1. Analisis karbohidrat (%)
Analisis kandungan karbohidrat dilakukan dengan menggunakan metode Luff-Schoorl (Lampiran 3)
2. Analisis nitrogen (%)
Analisis kandungan nitrogen dilakukan dengan menggunakan metode Semimikro Kjeldhal (Lampiran 4)
3. Rasio C/N (%)
Perhitungan rasio C/N yang dilakukan dengan menggunakan rumus: :
Rasio C/N =Kandungan karbohidrat
Kandungan nitrogen
Kualitas Buah Jeruk Keprok 1. Bobot buah (g)
Pengukuran dilakukan dengan menimbang setiap sampel buah hasil panen setiap tanaman.
2. Kulit buah (g)
3. Daging buah (g)
Pengukuran dilakukan dengan menimbang bobot daging buah yang telah dipisahkan dengan kulit buah dan biji .
4. Biji (g)
Pengukuran dilakukan dengan menimbang bobot biji buah yang telah dipisahkan dengan daging buah dan kulit buah.
5. Jus (g)
Pengukuran dilakukan dengan menimbang total jus yang diperoleh dari setiap buah.
6. Edible portion (%)
Pengukuran edible portion mengacu pada Muchtadi dan Sugiyono (1989) dengan menimbang bobot daging buah dan membandingkan dengan bobot total buah. Pengamatan menggunakan timbangan analitik dan dihitung dengan rumus :
BDD = Bobot Daging Buah
Bobot Total Buah x 100 %
7. Juice portion (%)
Pengukuran juice portion dengan menimbang bobot jus buah dan membandingkan dengan total buah. Pengamatan menggunakan timbangan analitik dan dihitung dengan rumus :
JP = Bobot Jus Buah
Bobot Total Buah x 100 %
8. Padatan Terlarut Total (%)
Prosedur mengacu pada AOAC (1995), dimana padatan terlarut total diukur dengan refraktometer dan dinyatakan dalam oBrix. Daging buah dihancurkan, kemudian diambil sarinya menggunakan kain saring. Selanjutnya sari buah tersebut diletakkan pada prisma reftraktometer, kemudian ditutup dan dilakukan pembacaan di tempat terang. Sebelum dan sesudah digunakan, prisma refraktometer dibersihkan dengan aquades dan tisu.
9. Asam Tertitrasi (%)
Prosedur mengacu pada AOAC (1995) dimana kandungan asam diukur dengan menghitung persen asam tertitrasi. Pengukuran kadar asam dilakukan dengan menimbang sari jeruk sebesar 10 g kemudian diencerkan pada labu takar 250 ml. Larutan dipipet 25 ml, dimasukan ke dalam erlenmeyer 125 ml yang dilakukan sebanyak dua kali. Pengukuran dilakukan dengan metode titrasi basa dengan menggunakan NaOH 0.1 N dan indikator fenoftalein (tiga tetes). Titrasi dilakukan sampai larutan berwarna merah muda stabil. Kandungan asam titrasi dihitung menggunakan rumus :
Asam tertitrasi =ml NaOH x N NaOH x fp x 64
bobot bahan mg x 100%
Keterangan :
15
HASIL DAN PEMBAHASAN
Pertumbuhan Vegetatif
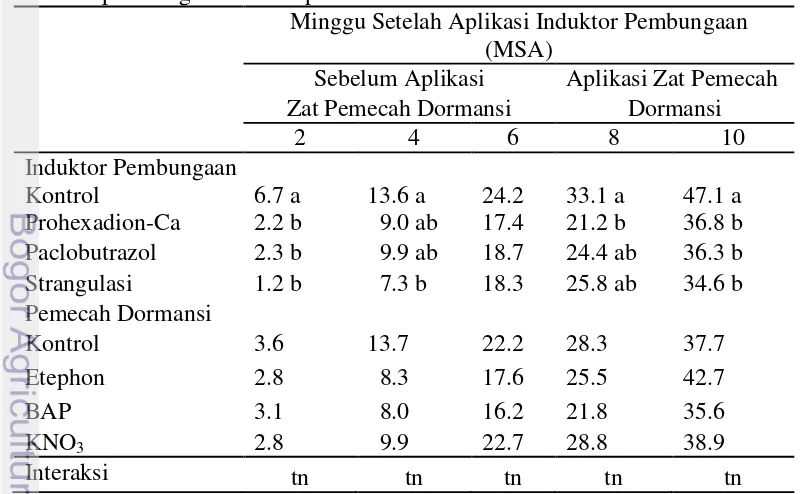
Perlakuan Prohexadion-Ca, Paclobutrazol, dan strangulasi secara nyata menurunkan jumlah tunas per cabang dibandingkan dengan kontrol. Jumlah tunas vegetatif pada tanaman kontrol jauh lebih banyak yaitu 47.1 tunas dibandingkan dengan tanaman yang mendapatkan perlakuan penginduktor pembungaan. Jumlah tunas vegetatif per cabang yang paling rendah terdapat pada perlakuan strangulasi yaitu 34.6 tunas, sedangkan perlakuan Paclobutrazol dan Prohexadion-Ca rata-rata jumlah tunas yang terbentuk yaitu 36.3 tunas dan 36.8 tunas pada 10 MSA (Minggu Setelah Aplikasi) induktor pembungaan (Tabel 1). Tekanan pertumbuhan tunas pada perlakuan Prohexadion-Ca dan Paclobutrazol disebabkan karena adanya penghambatan biosintesis giberelin. Menurut Wattimena (1988) pengaruh giberelin terutama di dalam perpanjangan ruas tanaman yang disebabkan oleh ukuran sel bertambah besar. Selain pemanjangan batang, giberelin juga memperbesar luas daun dari berbagai jenis tanaman, mempengaruhi proses perkecambahan, dan mengakhiri masa dorman pada biji. Menurut Poerwanto (2003) menunjukan bahwa perlakuan strangulasi dapat menekan munculnya tunas vegetatif, panjang tunas, dan jumlah daun karena terjadi penghambatan aliran fotosintat ke akar yang menyebabkan akar kekurangan energi dalam penyerapan mineral terutama nitrogen, air, dan dalam sistesis hormon di antaranya giberelin sehingga dapat menekan pertumbuhan vegetatif pada tanaman. Davies (1995) mengemukakan bahwa kandungan giberelin yang menurun akan mengakibatkan terjadinya penurunan laju pertumbuhan sel pada meristem sub-apikal sehingga pertumbuhan vegetatif akan terhambat.
Tabel 1 Jumlah tunas per cabang tanaman jeruk keprok pada perlakuan induktor pembungaan dan zat pemecah dormansi
Minggu Setelah Aplikasi Induktor Pembungaan (MSA)
Sebelum Aplikasi Aplikasi Zat Pemecah Zat Pemecah Dormansi Dormansi
Perlakuan penginduktor pembungaan secara nyata menekan panjang tunas pada tanaman jeruk dibandingkan perlakuan kontrol. Panjang tunas pada tanaman kontrol 11.63 cm pada 10 MSA induktor pembungaan lebih panjang dibandingkan dengan tanaman yang mendapat perlakukan penginduktor pembungaan. Panjang tunas vegetatif pada perlakuan penginduktor pembungaan berturut-turut adalah Paclobutrazol 6.16 cm, Prohexadion-Ca 7.88 cm, dan strangulasi 8.03 cm pada 10 MSA induktor pembungaan (Tabel 2). Penghambatan pertumbuhan panjang tunas pada perlakuan Paclobutrazol dan Prohexadion-Ca terjadi karena adanya penghambatan biosintesis giberelin dan mengakibatkan terhambatnya pertumbuhan vegetatif pada tanaman. Arteca (1996) menyatakan bahwa giberelin adalah faktor endogen yang menghambat pembungaan tanaman buah karena merangsang pertumbuhan vegetatif. Wattimena (1987) menyatakan bahwa giberelin memiliki pengaruh terutama di dalam pemanjangan ruas tanaman yang disebabkan oleh bertambah besar dan bertambahnya jumlah sel-sel pada ruas-ruas tanaman. Prawiranata et al. (1992) menyatakan bahwa penghambatan terhadap biosintesis giberelin akan merangsang biosintesis hormon lainnya seperti asam absisi (ABA), dimana ABA merupakan hormon tumbuhan yang berpengaruh secara fisiologis yaitu menyebabkan tunas menjadi dorman sehingga pertumbuhan vegetatif menjadi terhambat, yang ditunjukkan dengan lebih rendahnya jumlah tunas vegetatif yang dihasilkan dan tunas berukuran lebih pendek dibandingkan kontrol. Pada perlakuan strangulasi adanya tekanan pertumbuhan vegetatif pada tanaman jeruk disebabkan karena terhambatnya translokasi fotosintat dari tajuk ke akar. Novita (2007) dan Yamanishi et al. (1993) menyatakan bahwa tunas vegetatif pada tanaman yang distrangulasi akan lebih rendah dibandingkan dengan tanaman tidak distrangulasi, hal ini disebabkan karena terjadinya penghambatan translokasi dari tajuk ke akar sehingga terjadi penghambatan pertumbuhan tunas vegetatif dan menyebabkan terjadi peningkatan karbohidrat pada daun.
Tabel 2 Panjang tunas tanaman jeruk keprok pada perlakuan induktor pembungaan dan zat pemecah dormansi
Minggu Setelah Aplikasi Induktor Pembungaan (MSA) Sebelum Aplikasi Zat
17
Perlakuan penginduktor pembungaan secara nyata menekan pertumbuhan jumlah daun pada tanaman jeruk dibandingkan perlakuan kontrol. Jumlah daun yang terbentuk pada tanaman dengan perlakuan penginduktor pembungaan lebih sedikit dibandingkan dengan kontrol (Tabel 3). Perlakuan yang paling efektif dalam menekan terbentuknya jumlah daun adalah perlakuan Prohexadion-Ca dan Paclobutrazol yaitu 7.8 daun dibandingkan dengan kontrol yaitu 14.2 daun pada 10 MSA induktor pembungaan. Perlakuan strangulasi menunjukkan jumlah daun yang muncul per tunas yaitu 8.5 daun lebih sedikit dibandingkan dengan perlakuan kontrol. Adanya tekanan pertumbuhan vegetatif pada perlakuan Prohexadion-Ca dan Paclobutrazol disebabkan karena adanya penghambatan biosintesis giberelin. Penelitian John (2002) menunjukkan bahwa aplikasi Prohexadion-Ca menekan pertumbuhan panjang tunas pada tanaman. Terhambatnya biosintesis giberelin oleh kedua zat pengatur tumbuh tersebut secara langsung akan menghambat pertumbuhan vegetatif tanaman. Pada beberapa pohon buah-buahan dewasa seperti jeruk (Poerwanto dan Susanto 1996), manggis (Rai et al. 2006) dan mangga (Efendi 1994), penghambatan pertumbuhan vegetatif terjadi dengan aplikasi Paclobutrazol. Pada tanaman yang mendapatkan perlakuan strangulasi, pertumbuhan vegetatif terhambat karena rendahnya suplai air dan hara nitrogen. Strangulasi menyebabkan aliran hasil fotosintesis ke akar berkurang, sehingga akar kekurangan energi untuk menyerap hara dan air.
Tabel 3 Jumlah daun tanaman jeruk keprok pada perlakuan induktor pembungaan dan zat pemecah dormansi.
Minggu Setelah Aplikasi Induktor Pembungaan (MSA) Sebelum Aplikasi Aplikasi Zat Pemecah Zat Pemecah Dormansi Dormansi
2 4 6 8 10
Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5% (uji selang berganda Duncan)
yang muncul pada tanaman jeruk keprok. Susanto dan Poerwanto (1999) juga menyatakan bahwa perlakuan zat pemecah dormansi tidak berpengaruh terhadap waktu munculnya bunga karena tanaman mangga berbunga sebelum diaplikasikan zat pemecah dormansi.
Perlakuan strangulasi menunjukkan kehijauan daun yang berbeda nyata dibandingkan Prohexadion-Ca, Paclobutrazol, dan kontrol (Tabel 4). Strangulasi menyebabkan aliran hasil fotosintesis ke akar berkurang sehingga akar kekurangan energi untuk menyerap hara dan air. Kurangnya energi dalam menyerap nitrogen akan menurunkan kandungan nitrogen pada tanaman yang menyebabkan kandungan klorofil akan berkurang sehingga warna daun akan berubah menjadi kuning. Selain itu, strangulasi menyebabkan kandungan karbohidrat pada daun meningkat. Karbohidrat pada daun disimpan pada chloroplas. Pati yang tidak terpigmentasi akan disimpan pada amioplas sehingga warna daun akan berubah menjadi kuning. Menurut pendapat Gardner et al. (1991) menyatakan bahwa gejala kekurangan N mengakibatkan berkurangnya warna hijau dari dedaunan (chlorosis). Daun akan menjadi lebih pucat, menguning, dan akan mengakibatkan kematian.
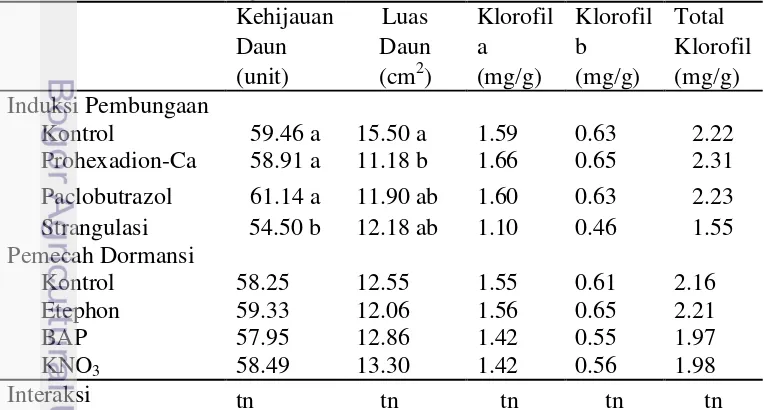
Perlakuan Prohexadion-Ca secara nyata menekan perkembangan luas daun pada tanaman jeruk keprok dibandingkan kontrol, namun tidak berbeda nyata dengan perlakuan Paclobutrazol dan strangulasi (Tabel 4). Luas daun pada perlakuan Prohexadion-Ca yaitu 11.18 cm2 lebih rendah dibandingkan kontrol (15.50 cm2), Paclobutrazol (11.90 cm2), dan strangulasi (12.18 cm2). Hal ini diduga, pada perlakuan Prohexadion-Ca ukuran sel daun tidak memanjang, sehingga fungsi untuk memperluas daun juga terhambat. Terhambatnya pembelahan sel pada tepi daun berhubungan dengan terhambatnya biosintesis giberelin pada tanaman. Wattimena (1987) mendefinisikan zat penghambat tumbuh (retardan) sebagai senyawa-senyawa organik sintetik yang bila diberikan kepada tanaman yang responsif akan menghambat perpanjangan sel pada meristem sub apikal, mengurangi laju perpanjangan batang.
Tabel 4 Kehijauan daun, luas daun, dan kandungan klorofil daun pada perlakuan induktor pembungaan dan zat pemecah dormansi
Kehijauan Luas Klorofil Klorofil Total
19
Pemberian perlakuan penginduktor pembungaan, zat pemecah dormansi, dan interaksi antara penginduktor pembungaan dengan zat pemecah dormansi tidak berpengaruh nyata terhadap pengamatan kandungan klorofil a, klorofil b, dan total klorofil. Kandungan klorofil pada masing-masing perlakuan secara berturut-turut, yaitu Prohexadion-Ca 2.31 mg/g, Paclobutrazol 2.23 mg/g, kontrol 2.22 mg/g, dan strangulasi 1.55 mg/g (Tabel 4).
Pertumbuhan Generatif
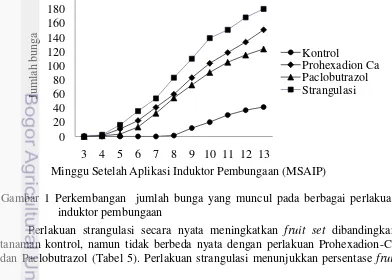
Perlakuan penginduktor pembungaan secara nyata mempercepat waktu berbunga dibandingkan tanaman kontrol. Prohexadion-Ca dan strangulasi mengakibatkan waktu mulai berbunga lebih cepat yaitu pada 17.50 HSAIP (Hari Setelah Aplikasi Induktor Pembungaan) dan 16.50 HSAIP dibandingkan dengan kontrol (39.00 HSAIP). Perlakuan Paclobutrazol secara nyata juga mempercepat waktu berbunga yaitu 23.38 HSAIP dibandingkan kontrol tetapi tidak sebaik perlakuan Prohexadion-Ca dan strangulasi (Tabel 5).
Total bunga secara nyata meningkat dengan perlakuan Prohexadion-Ca, Paclobutrazol, dan strangulasi dibandingkan kontrol. Jumlah bunga pada perlakuan Prohexadion-Ca yaitu 150.88 bunga secara nyata lebih banyak dari kontrol (41.75 bunga) namun tidak berbeda nyata dengan perlakuan Paclobutrazol (123.81 bunga) dan strangulasi (180.50 bunga) (Tabel 5).
Perlakuan penginduksi pembungaan meningkatkan jumlah bunga pada tanaman jeruk keprok dibandingkan kontrol Strangulasi menunjukan jumlah bunga yang paling banyak yaitu 180.5 bunga dibandingkan perlakuan Prohexadion-Ca (123.8 bunga), Paclobutrazol (150.9 bunga), dan kontrol (41.8 bunga) pada 13 MSA induksi pembungaan (Gambar 1).
Perlakuan Prohexadion-Ca dan Paclobutrazol secara nyata mempercepat waktu pembungaan dan meningkatkan jumlah bunga karena Prohexadion-Ca dan Paclobutrazol mengakibatkan terhambatnya biosintesis giberelin pada tanaman. Sach (1977) menyatakan bahwa pembungaan berhubungan dengan kandungan giberelin dalam tanaman. Kandungan giberelin yang tinggi akan meningkatkan pertumbuhan vegetatif sehingga menghambat pembungaan. Dengan demikian, pemberian Paclobutrazol sebagai zat penghambat tumbuh yang menghambat biosintesis giberelin dapat menginduksi pembungaan. Terhambatnya biosintesis giberelin mengakibatkan terhambatnya pertumbuhan vegetatif pada tanaman sehingga aktivitas pertumbuhan akan dialihkan ke pertumbuhan bunga atau buah dan akan meningkatkan rasio C/N pada tanaman. Bernier et al. (1984) menyatakan bahwa pembungaan berhubungan dengan kandungan giberelin dalam tanaman. Kandungan giberelin yang tinggi pada beberapa pohon buah-buahan seperti jeruk, mangga, dan manggis akan memacu pertumbuhan vegetatif dan menghambat pembungaan. Bioaktivitas giberelin endogen pada daun jeruk dan manggis pada fase induksi pembungaan lebih rendah dibandingkan pada fase pertumbuhan vegetatif (Poerwanto dan Inoue 1990, Rai et al. 2004). Paclobutrazol sebagai zat penghambat tumbuh yang menghambat biosintesis giberelin dapat menginduksi pembungaan beberapa pohon buah-buahan (Voon et al. 1992). Adiel
0
berpengaruh dalam menghambat pertumbuhan vegetatif tanaman mangga dan menginduksi pembungaan mangga di luar musim, hal tersebut diduga berhubungan dengan cara kerja Paclobutrazol dalam menghambat biosintesis giberelin.
Strangulasi akan merusak jaringan floem, sehingga menghambat translokasi hasil fotosintesis dari bagian atas tanaman ke bagian akar. Akibatnya terjadi penumpukan karbohidrat di bagian pucuk dan akar kekurangan fotosintat (karbohidrat). Akar yang kekurangan fotosintat akan mengalami penurunan aktivitas dalam serapan air dan hara, serta biosintesis giberelin dan sitokinin. Berkurangnya serapan air menyebabkan stress air yang dapat menginduksi pembungaan. Rendahnya serapan nitrogen dan tingginya akumulasi karbohidrat di bagian pucuk tanaman akan meningkatkan nisbah C/N yang merupakan syarat terjadinya induksi pembungaan. Rendahnya biosintesis giberelin juga mengakibatkan terjadinya induksi pembungaan yang disebabkan oleh perlakuan strangulasi. Pada rambutan perlakuan ringing dapat meningkatkan jumlah bunga dan buah, bahkan dapat menginduksi tanaman rambutan pada off year (tahun tidak berbunga) (Poerwanto dan Irdiastuti 2005). Menurut Ryoko (1988), kerat batang dapat menekan gerakan fotosintat (karbohidrat) dari daun ke akar sehingga akan terjadi akumulasi karbohidrat di daun yang selanjutnya digunakan untuk pembungaan. Karbohidrat pada daun akan dialokasikan untuk pembentukan bunga dan buah pada tanaman. Thamrin (2008) menyatakan bahwa tingginya jumlah bunga yang terbentuk diduga merupakan hasil penekanan strangulasi yang memblokir gerakan fotosintesis dari daun ke akar, sehingga terjadi penumpukan karbohidrat yang selanjutnya digunakan untuk pembungaan. Putra (2002) menyatakan bahwa strangulasi akan menghambat translokasi fotosintat dari tajuk ke akar, peningkatan karbohidrat tersebut akan menyebabkan rasio C/N pada tajuk akan tinggi sehingga dapat menginduksi pembungaan.
Perlakuan strangulasi secara nyata meningkatkan fruit set dibandingkan tanaman kontrol, namun tidak berbeda nyata dengan perlakuan Prohexadion-Ca dan Paclobutrazol (Tabel 5). Perlakuan strangulasi menunjukkan persentase fruit
21
set yaitu 65.42% lebih tinggi dibandingkan dengan Prohexadion-Ca (49.40%), Paclobutrazol (51.85%), dan kontrol 34.46%. Fruit set yang tinggi diperoleh dari semakin banyaknya jumlah bunga yang berdeferensiasi menjadi buah pada tanaman. Ryugo (1988) menyatakan bahwa produksi buah tergantung pada tunas yang berdeferensiasi menjadi bunga, bunga mekar yang mengalami penyerbukan, dan bunga yang berkembang menjadi buah.
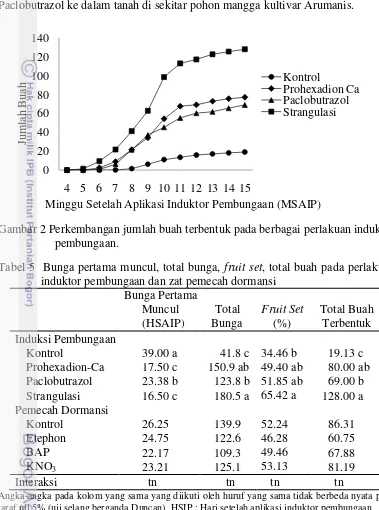
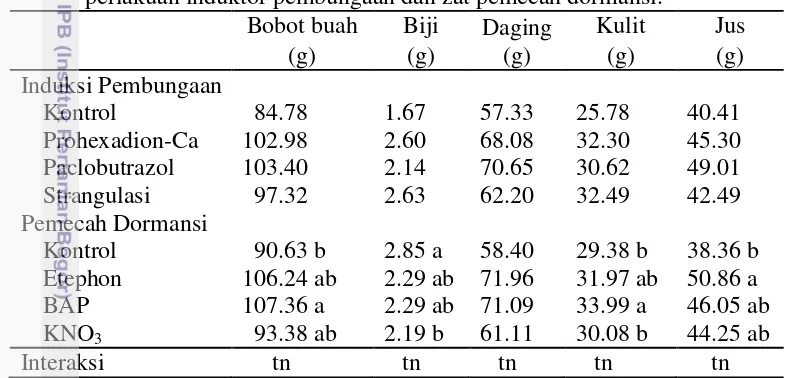
Total buah yang terbentuk, buah yang dipanen, dan bobot buah panen secara nyata meningkat tajam dengan perlakuan penginduktor pembungaan (Tabel 5) (Tabel 6). Hal ini terjadi karena selain adanya peningkatan jumlah bunga pada perlakuan penginduktor pembungaan (Tabel 5), peningkatan fruit set (Tabel 5), peningkatan kandungan karbohidrat (Tabel 7), peningkatan rasio C/N (Tabel 7) juga disebabkan karena penurunan persentase gugur buah (Tabel 6). Perhitungan persentase gugur bunga menunjukkan perlakuan kontrol memiliki tingkat gugur yang lebih tinggi yaitu 63.49% dibandingkan dengan perlakuan penginduktor pembungaan. Perlakuan penginduktor pembungaan memiliki persentase gugur buah yaitu Prohexadion-Ca 36.03%, Paclobutrazol 43.65 %, dan strangulasi 39.07%. Gugur bunga disebabkan karena adanya faktor lingkungan seperti angin, hujan, dan serangan hama dan penyakit. Hal ini sesuai pendapat Effendi (1994) yang menyatakan bahwa curah hujan yang tinggi sangat mempengaruhi tanaman dalam proses pembungaan dan pembuahan pada tanaman mangga. Laporan Thamrin (2008) dalam penelitiannya, menunjukan bahwa tingginya kuncup bunga dan bunga mekar tidak diikuti dengan jumlah buah yang terbentuk, hal ini disebabkan pada saat menjelang bunga mekar intesitas curah hujan tinggi sehingga menyebabkan banyak bunga yang rontok sebelum berbuah. Lebih jauh Rukayah et al. (1996) menyatakan bahwa selain faktor genetik, penyebab kerontokan buah adalah curah hujan, angin, serangan hama dan penyakit serta defisiensi hara dan hormonal.
dengan kemampuan dalam menurunkan kandungan giberelin dan miningkatkan kandungan gula serta rasio C/N pada daun. Wattimena (1987) menyatakan bahwa Paclobutrazol dapat menyebabkan pembungaan dan tentunya pembuahan yang lebih awal, lebih serempak, dan lebih banyak dengan perlakuan penyiraman Paclobutrazol ke dalam tanah di sekitar pohon mangga kultivar Arumanis.
.
Gambar 2 Perkembangan jumlah buah terbentuk pada berbagai perlakuan induktor pembungaan.
Tabel 5 Bunga pertama muncul, total bunga, fruit set, total buah pada perlakuan induktor pembungaan dan zat pemecah dormansi
Bunga Pertama
Muncul Total Fruit Set Total Buah (HSAIP) Bunga (%) Terbentuk Induksi Pembungaan
Kontrol 39.00 a 41.8 c 34.46 b 19.13 c Prohexadion-Ca 17.50 c 150.9 ab 49.40 ab 80.00 ab Paclobutrazol 23.38 b 123.8 b 51.85 ab 69.00 b Strangulasi 16.50 c 180.5 a 65.42 a 128.00 a Pemecah Dormansi
Kontrol 26.25 139.9 52.24 86.31 Etephon 24.75 122.6 46.28 60.75 BAP 22.17 109.3 49.46 67.88 KNO3 23.21 125.1 53.13 81.19 Interaksi tn tn tn tn
Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5% (uji selang berganda Duncan). HSIP : Hari setelah aplikasi induktor pembungaan
0 20 40 60 80 100 120 140
4 5 6 7 8 9 10 11 12 13 14 15
Juml
ah
B
ua
h
Minggu Setelah Aplikasi Induktor Pembungaan (MSAIP) Kontrol
23
Tabel 6 Gugur buah, jumlah buah panen, dan bobot panen pada perlakuan induktor pembungaan dan zat pemecah dormansi
Gugur Buah Jumlah Buah Bobot Penen
(%) Panen (kg)
Induksi Pembungaan
Kontrol 63.49 a 7.06 b 0.64 b
Prohexadion-Ca 36.03 b 53.31 a 5.60 a Paclobutrazol 43.65 ab 43.69 ab 4.43 ab
Strangulasi 39.07 b 78.69 a 7.46 a
Pemecah Dormansi
Kontrol 37.46 53.31 4.85
Etephon 42.02 38.13 4.13
BAP 45.60 40.81 4.51
KNO3 57.18 50.50 4.64
Interaksi tn tn tn
Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5% (uji selang berganda Duncan).
Zat pemecah dormansi tidak berpengaruh nyata dalam mempercepat waktu munculnya bunga, meningkatkan total bunga, fruit set, meningkatkan total buah, gugur buah, jumlah buah panen, dan bobot panen. Tidak ada interaksi antara perlakuan induksi pembungaan dan zat pemecah dormansi (Tabel 5) (Tabel 6). Hal ini disebabkan karena sebelum pengaplikasian zat pemecah dormansi, sebagian tanaman telah berbunga sehingga aplikasi zat pemecah dormansi tidak berpengaruh nyata terhadap pengamatan waktu munculnya bunga, total bunga,
fruit set, total buah, buah panen, dan bobot buah panen. Susanto dan Poerwanto (1999) juga menyatakan bahwa perlakuan zat pemecah dormansi tidak berpengaruh terhadap waktu munculnya bunga karena tanaman mangga berbunga sebelum diaplikasikan zat pemecah dormansi.
Kandungan Karbohidrat, Nitrogen, dan Rasio C/N
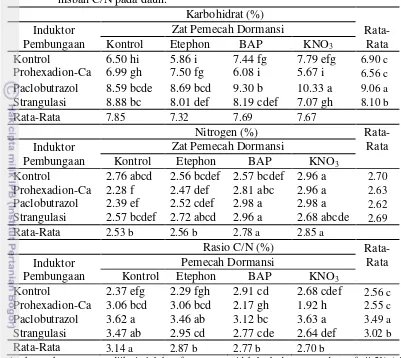
Hasil analisis kandungan karbohidrat, nitrogen, dan rasio C/N menunjukan terdapat interaksi yang nyata antara perlakuan penginduksi pembungaan dengan pemberian zat pemecah dormansi. Interaksi perlakuan Paclobutrazol dengan KNO3 (10.33%) menunjukkan nilai kandungan karbohidrat yang tertinggi dibandingkan interaksi antara perlakuan yang lainnya. Interaksi antara Paclobutrazol dengan BAP (9.30%) juga menunjukkan kandungan karbohidrat yang tinggi namun tidak berbeda nyata dengan interaksi antara perlakuan Paclobutrazol dengan etephon (8.69%), Paclobutrazol tanpa pemberian zat pemecah dormansi (8.59%), dan strangulasi tanpa pemberian zat pemecah dormansi (8.88%) (Tabel 7).
dormansi (2.57%), dan pemberian etephon dan BAP tanpa perlakuan induksi pembungaan yaitu 2.56 % dan 2.57 % (Tabel 7).
Interaksi antara perlakuan Paclobutrazol dengan KNO3 (3.63 %) menunjukan rasio C/N yang lebih tinggi dibandingkan kontrol (2.37%) namun tidak berbeda nyata dengan perlakuan Paclobutrazol tanpa pemberian zat pemecah dormansi (3.62%), interaksi antara Paclobutrazol dan etephon (3.46%), dan strangulasi tanpa pemberian zat pemecah dormansi (3.47%) (Tabel 7).
Paclobutrazol dan Prohexadion-Ca merupakan senyawa kimia yang bersifat penghambat biosintesis giberelin. Paclobutrazol dan Prohexadion-Ca berpengaruh terhadap penghambatan pertumbuhan vegetatif melalui penghambatan biosintesis giberelin. Adanya penghambatan biosintesis giberelin mengakibatkan kandungan karbohidrat dan rasio C/N meningkat dan akan merangsang pertumbuhan reproduksi tanaman (ICI 1986). Penghamatan biosintesis giberelin pada tanaman jeruk keprok menyebabkan terhambatnya pertumbuhan vegetatif. Terhambatnya pertumbuhan vegetatif pada tanaman akan meningkatkan kandungan karbohidrat dan rasio C/N pada tajuk tanaman jeruk keprok. Hal ini disebabkan karena karbohidrat hasil dari proses fotosintesis tidak dapat dialokasikan untuk pertumbuhan vegetatif pada tanaman. Menurut Harjadi (1996) tanaman menggunakan sebagian besar karbohidrat yang dibentuk dalam suatu fase vegetatif perkembangan tanaman. Barnier et al. (1985) menyatakan bahwa rasio C/N tinggi merupakan faktor pendorong tanaman untuk berbunga.
Kandungan karbohidrat dan rasio C/N yang tinggi pada tanaman dengan perlakuan strangulasi disebabkan terhambatnya translokasi fotosintat dari tajuk ke akar sehingga terjadi penumpukan karbohidrat dibagian tajuk. Akumulasi karbohidrat daun di bagian tajuk yang diikuti dengan berkurangnya absorsi hara mineral terutama nitrogen yang akan meningkatkan rasio C/N tajuk. Terhambatnya translokasi karbohidrat ke akar mengakibatkan akar kekurangan fotosintat dan respirasi akar menurun sehingga mengakibatkan aktivitas akar dalam mengabsorsi hara, mineral, dan air terganggu. Dalam penelitian ini terhambatnya translokasi fotosintat ditunjukan oleh tingginya kandungan karbohidrat, sedangkan terganggunya serapan hara ditunjukkan oleh turunnya kandungan N total daun sehingga rasio C/N pada perlakuan strangulasi tinggi. Thamrin (2008) menyatakan bahwa perlakukan strangulasi dapat meningkatkan rasio C/N dan mendorong pembungaan pada tanaman jeruk. Yamanishi et al.
25
Tabel 7 Pengaruh interaksi antara perlakuan induktor pembungaan dan zat pemecah dormansi terhadap kandungan karbohidrat, nitrogen, dan nisbah C/N pada daun.
Angka-angka yang sama diikuti oleh huruf yang sama tidak berbeda nyata pada taraf uji 5% (uji selang berganda Duncan)
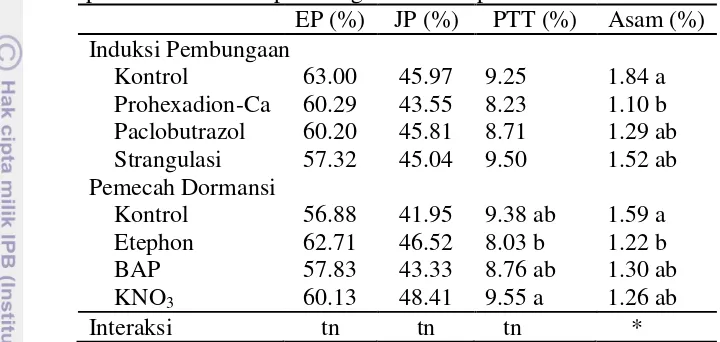
Kualitas Buah