TELAAH AKTIVITAS BAKTERI PENGHASIL LIPASE
YANG BERASOSIASI DENGAN TEMPE
NASWANDI NUR
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Telaah Aktivitas Bakteri Penghasil Lipase yang Berasosiasi dengan Tempe” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2015
RINGKASAN
NASWANDI NUR. Telaah Aktivitas Bakteri Penghasil Lipase yang Berasosiasi dengan Tempe. Dibimbing oleh ANTONIUS SUWANTO dan ANJA MERYANDINI.
Tempe dikenal sebagai produk makanan hasil fermentasi asal Indonesia. Selain sebagai bahan makanan bernilai gizi tinggi, tempe juga dikenal sebagai ekosistem bagi berbagai jenis mikroorganisme. Kadar lemak total pada tempe menunjukkan penurunan bila dibandingkan dengan kadar lemak pada biji kedelai tanpa fermentasi. Penurunan kadar lemak pada tempe ini menjadi indikator keberadaan mikroorganisme lipolitik di dalamnya. Adanya aktivitas lipolisis pada tempe ini menjadikan tempe lebih mudah dicerna dan memiliki aroma yang khas dari hasil hidrolisis asam lemak.
Sampel yang digunakan merupakan makanan tradisional asal Indonesia yaitu tempe. Tempe dikumpulkan dari berbagai pengrajin tempe dengan kategori pemilihan sampel tertentu. Pengamatan kondisi fisik, konsistensi dan aroma tempe dilakukan pada beberapa rentang waktu pengamatan setelah pengumpulan sampel. Sampel tempe yang telah diamati selanjutnya dienumerasi jumlah total populasi bakteri dan bakteri penghasil lipase (lipolitik). Metode yang digunakan ialah standard plate count dengan media kultivasi berupa plate count agar untuk total populasi bakteri dan media agar-agar Luria Bertani yang diemulsikan dengan minyak zaitun dan rhodamine B untuk bakteri lipolitik. Peranan bakteri lipolitik dalam pembentukan cita rasa tempe juga akan dikaji secara ringkas berdasarkan profil kandungan asam lemak bebas yang dianalisis dengan metode gas chromatrography with flame ionizer detector (GC-FID) .
Pengumpulan sampel tempe diperoleh dari tiga pengrajin tempe berbeda di kota Bogor, selanjutnya disebut dengan CMG, EMP dan RTI. Kriteria pembeda ketiga sampel tempe berdasarkan kebersihan selama proses produksi tempe. Perubahan kondisi fisik, konsistensi dan aroma tempe menunjukkan perbedaan pada tiap jenis tempe. Perubahan yang teramati selama pengamatan paling cepat terjadi pada tempe CMG diikuti oleh tempe EMG dan perubahan paling lambat terjadi pada tempe RTI. Hasil enumerasi total populasi bakteri menunjukkan kepadatan populasi tertinggi terjadi pada tempe CMG sekitar 2.31 × 109 cfu g-1 selanjutnya diikuti oleh tempe EMP sebesar 9.78 × 108 cfu g-1 dan tempe RTI sebesar 4.04 × 106 cfu g-1. Kepadatan populasi bakteri lipolitik yang teramati ialah seperseribu dari total populasi bakteri. Tempe CMG memiliki kepadatan populasi bakteri lipolitik tertinggi sebesar 1.04 × 106 cfu g-1 diikuti oleh tempe EMP sebesar 2.88 × 105 cfu g-1 dan tempe RTI sekitar 1.72 × 104 cfu g-1. Selain itu ditemukan adanya hubungan yang unik antara dinamika populasi bakteri lipolitik terhadap perubahan kondisi fisik, konsistensi dan aroma tempe yang terjadi selama pengamatan. Komposisi lemak bebas pada tempe CMG, EMP dan RTI didominasi oleh senyawa asam linoleat, oleat, palmitat, linolenat dan stearat. Kata kunci: tempe, pembentukan cita rasa, bakteri lipolitik, GC-FID, asam lemak
Associated with Tempe. Supervised by ANTONIUS SUWANTO and ANJA MERYANDINI.
Tempe is known as fermented food products the origin of Indonesia. In addition as high nutritional value of foodstuffs, tempe is also known as the ecosystem for many types of microorganisms. Total fat content on tempe shows a decrease when compared to the fat content in soybean without fermentation. A decrease in fat levels in tempe is becoming an indicator of existence of lipolytic microorganisms in it. The existence of lipolysis activity in tempe make it more easy to digest and have a distinctive scent of the hydrolysis of the fatty acids.
The sample was the traditional foods origin of Indonesia namely tempe. Tempe was collected from various of tempe craftsmen with specific of sample selection category. Observation of physical condition, consistency and aroma of tempe were done during several observation time span after sample collection. Samples of tempe have been observed subsequent of total bacterial population and lipase producing bacteria (lipolytic). The method is the standard plate count which plate count agar medium for enumeration of total bacterial population and Luria-Bertani agar medium emulsified with olive oil and rhodamine B for enumeration of lipolytic bacteria. The role of lipolytic bacteria in the flavour development of tempe also examined in briefly based on the content of free fatty acid profiles were analyzed by gas chromatrography with flame ionizer detector (GC-FID) method.
The collection of samples obtained from three different tempe craftsmen in Bogor, hereinafter referred to as CMG, EMP and RTI. The all of those three distinguishing criteria of tempe sample based on cleanliness during the process of tempe production. Changes in physical condition, consistency and aroma of the tempe shows the difference on each type of tempe. The changes observed during the fastest observations occurred in tempe CMG followed by tempe EMG and slowest changes occur in tempe RTI. Total bacteria and lipolytic bacterial populations were carried out employing standard plate count method and showed CMG harbored the highest bacterial population number 2.31 × 109 cfu g-1 followed by EMP 9.78 × 108 cfu g-1 and RTI 4.04 × 106 cfu g-1. Lipolytic bacteria were found in each tempe samples which were approximately 0.1 % of total bacterial population. CMG harbored the highest lipolytic bacterial population number 1.04 × 106 cfu g-1 followed by EMP 2.88 × 105 cfu g-1 and RTI 1.72 × 104 cfu g-1. Tempe aroma and texture showed strong correlation between the numbers of bacterial population during the period or stages of tempe maturation. Moreover, we also found a unique correlation between dynamic of lipolytic population and flavor development. Free faty acid composition on tempe CMG, EMP and RTI are dominated by linoleic, oleic, palmitic, linolenic and stearic acid respectively. Keywords: tempe, flavor development, lipolytic bacteria, GC-FID, free fatty
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
TELAAH AKTIVITAS BAKTERI PENGHASIL LIPASE
YANG BERASOSIASI DENGAN TEMPE
NASWANDI NUR
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Mikrobiologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan September 2014 hingga Juni 2015 ini ialah mikrobiologi tempe, dengan judul Telaah Aktivitas Bakteri Penghasil Lipase yang Berasosiasi dengan Tempe.
Terima kasih penulis ucapkan kepada Prof Dr Ir Antonius Suwanto, MSc, sebagai ketua komisi pembimbing dan Prof Dr Dra Anja Meryandini, MS, sebagai anggota komisi pembimbing, yang telah banyak memberikan nasehat, saran, motivasi, waktu konsultasi, serta solusi dari setiap permasalahan yang dihadapi penulis selama melaksanakan penelitian dan penyusunan karya ilmiah ini. Selainitu penulis ucapkan terima kasih kepada penguji luar komisi Prof Dr Ir Made Astawan, MS yang telah memberikan motivasi selama studi dan masukan pada saat ujian sidang tesis. Kepada kementerian RISTEK-DIKTI melalui Beasiswa Program Magister Doktor Sarjana Unggul (PMDSU) batch 1 2013 terima kasih atas kepercayaannya untuk memberikan beasiswa kuliah dan dana penelitian selama menempuh pendidikan pascasarjana di IPB.
Penulis juga mengucapkan terima kasih kepada seluruh Staf Pengajar Departemen Mikrobiologi dan staf Laboratorium serta seluruh teman-teman di Laboratorium Mikrobiologi IPB, atas dukungan, motivasi, dan bantuannya selama penelitian ini. Ucapan terima kasih tak terhingga juga penulis ucapkan kepada bapak, ibu, kakak dan adikku tercinta, serta sahabat-sahabatku atas doa, dukungan, kasih sayang, dan semangat yang diberikan. Terima kasih untuk teman-teman seperjuangan di Pascasarjana Mikrobiologi IPB angkatan 2013 serta seluruh pihak yang telah memberikan doa dan dukungannya, penulis ucapkan terima kasih.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2015
DAFTAR ISI
Tempe Sebagai Produk Fermentasi Asal Indonesia ... 4
Komunitas Mikroorganisme pada Tempe ... 5
Mikroorganisme Penghasil Lipase ... 5
Keberadaan Aktivitas Lipase pada Tempe... 11
METODE PENELITIAN ... 13
Kerangka Penelitian ... 13
Waktu dan Lokasi Penelitian ... 13
Observasi dan Pengumpulan Sampel ... 13
Pengamatan Kondisi Fisik, Konsistensi dan Aroma Tempe ... 14
Media Kultur ... 14
Teknik Penghitungan Jumlah Total Koloni ... 14
Enumerasi Total Populasi Bakteri dan Populasi Bakteri Lipolitik ... 15
Analisis Metil Ester Asam Lemak Mengguakan GCFID ... 15
Analisis Statistik ... 15
HASIL DAN PEMBAHASAN ... 16
Survei dan Pengumpulan Sampel ... 16
Pengamatan Kondisi Fisik, Konsistensi dan Perubahan Aroma Tempe .... 16
Penghitungan Jumlah Total Koloni ... 17
Enumerasi Total Populasi Bakteri... 18
Enumerasi Populasi Bakteri Lipolitik ... 19
Dinamika Total Populasi Bakteri dan Populasi Bakteri Lipolitik... 20
Hubungan Dinamika Populasi Lipolitik terhadap Perubahan Aroma Tempe... 23
Analisis Kandungan Asam Lemak dengan Metode GCFID ... 24
SIMPULAN DAN SARAN ... 29
Simpulan ... 29
Saran ... 29
DAFTAR PUSTAKA ... 30
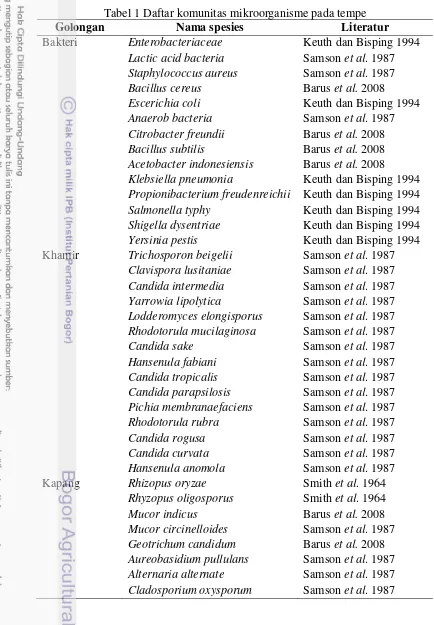
1 Daftar komunitas mikroorganisme pada tempe ... 6
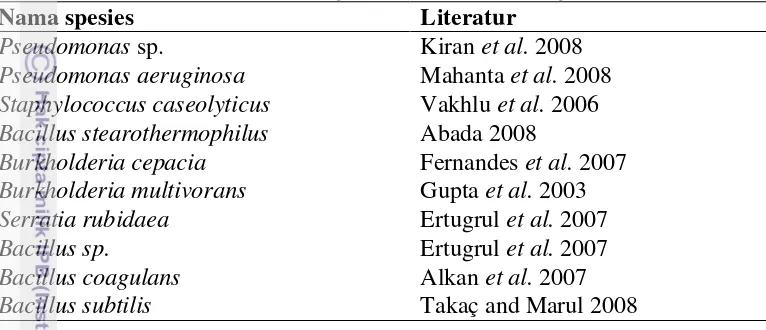
2 Daftar bakteri penghasil lipase dari berbagai literatur ... 8
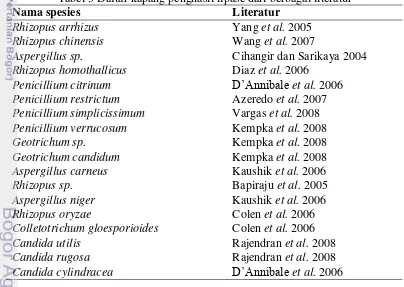
3 Daftar kapang penghasil lipase dari berbagai literatur ... 9
4 Daftar khamir penghasil lipase dari berbagai literatur ... 11
5 Pengamatan kondisi fisik, konsistensi dan perubahan aroma tempe ... 17
6 Total populasi bakteri pada media PCA dan bakteri lipolitik pada media LAOR yang diisolasi dari berbagai umur tempe dan diinkubasi pada suhu 38 0C selama 48 jam ... 18
7 Penghitungan jumlah total populasi bakteri pada media PCA yang diisolasi dari berbagai umur tempe dan diinkubasi pada suhu 38 0C selama 48 jam ... 19
8 Penghitungan jumlah populasi bakteri lipolitik pada media LAOR yang diisolasi dari berbagai umur tempe dan diinkubasi pada suhu 38 0C selama 48 jam ... 20
9 Profil proporsi kandungan asam lemak bebas tempe yang dianalisis pada berbagai umur tempe ... 25
DAFTAR GAMBAR
1 Ilustrasi tempe kedelai, dua hingga tiga hari setelah fermentasi ... 42 Diagram alir penelitian ... 13
3 Pengamatan total populasi bakteri pada media PCA serta populasi lipolitik pada media LAOR tanpa penyinaran UV dan dengan penyinaran UV 365 nm... 17
4 Dinamika total populasi bakteri yang berasosiasi dengan tempe CMG (□), tempe EMP (∆) dan tempe RTI (○) ... 21
5 Dinamika populasi bakteri lipolitik yang berasosiasi dengan tempe CMG (□), tempe EMP (∆) dan tempeRTI (○) ... 22
6 Hubungan antara dinamika bakteri lipolitik tempe CMG (■) dan EMP (▲) terhadap perubahan cita rasa aroma khas tempe (AKT) dan aroma tempe semangit (ATS) ... 23
7 Profil perbandingan komposisis asam lemak bebas dominan penyusun tempe CMG, EMP dan RTI yang diamati pada beberapa rentang waktu pengamatan ... 25
9 Perubahan komposisis asam lemak bebas dominan penyususun tempe CMG, EMP dan RTI yang diamati pada beberapa rentang waktu
pengamatan ... 27
DAFTAR LAMPIRAN
1 Survei Lokasi dan Pengumpulan Sampel... 352 Pengamatan Kondisi Fisik, Konsistensi dan Aroma Tempe ... 38
3 Enumerasi Total Populasi Bakteri dan Populasi Lipolitik ... 39
4 Media Kultur dan Reagen ... 45
5 Analisis Statistik Total Populasi Bakteri dengan Minitab ... 47
6 Analisis Statistik populasi Bakteri Lipolitik dengan Minitab ... 51
7 Analisis Asam Lemak dengan Metode GCFID ... 55
PENDAHULUAN
Latar Belakang
Tempe merupakan produk fermentasi kedelai menggunakan kapang Rhyzopus sp. tempe merupakan makanan tradisional yang berasal dari Indonesia dan telah dikenal sejak berabad-abad lamanya, terutama dalam tatanan budaya makan masyarakat jawa. Apabila dilihat dari segi nutrisi, tempe dikenal sebagai makanan berserat tinggi. Tempe juga mengandung berbagai jenis nutrisi penting seperti kalsium, vitamin B, mineral, zat besi dan sebagainya. Seiring berjalannya waktu, tempe kini menjadi makin popular tidak hanya di Indonesia melainkan di negara-negara lain seperti Jepang, Amerika Serikat, Kanada, Australia dan beberapa negara di Eropa Barat. Fakta tersebut menjadi motivasi tersendiri untuk lebih mengeksplorasi potensi yang ada pada tempe dan memperkenalkannya secara luas tidak hanya sebagai makanan murah bergizi tinggi namun juga sebagai agen bioteknologi yang memiliki nilai tinggi.
Tempe Indonesia merupakan ekosistem bagi banyak mikroorganisme dengan keragaman yang sangat tinggi (Barus et al. 2008). Selain Rhyzopus oligosporus, kehadiran bakteri juga memiliki peranan yang sangat penting selama proses fermentasi tempe. Beberapa diantara bakteri tersebut bahkan memegang peranan yang sangat penting dalam improvisasi rasa, aroma, nutrisi, serta kualitas tempe yang dihasilkan (Hagedorn dan Kaphammer 1994). Sebagai contoh, Citrobacter freundii dan Klebsiella pneumoniae meningkatkan kandungan vitamin B12 (Keuth dan Bisping 1994), Micrococcus atau Arthrobacter berperan dalam pembentukan senyawa isoflavones (Klus et al. 1993; Nakajima et al. 2005) dan R. oligosporus berperan dalam hidrolisis asam lemak (Yuliani et al. 2007).
Sama halnya dengan aroma dan nutrisi pada tempe, lipase tidak hanya dihasilkan oleh R. oligosporus namun beberapa mikrorganisme lain yang terdapat pada tempe juga mampu menghasilkan lipase. USDA National Nutrient Database for Standard reference telah merilis informasi yang menunjukkan jumlah lipid (lemak) yang terkandung pada tempe jauh lebih rendah jumlah lipid yang terkandung pada kedelai tanpa fermentasi. Data tersebut menunjukkan bahwa selama proses fermentasi kedelai, terjadi proses hidrolisis triacylglycerol menjadi asam lemak bebas oleh enzim lipase yang mengakibatkan turunnya kandungan lipid sebesar 25-26 % (Leitao et al. 1967; Astuti et al. 2000).
Hingga saat ini laporan mengenai mikroorganisme pada tempe yang memiliki aktivitas lipolitik terbatas pada Rhyzopus sp. dan Mucor sp. sementara masih banyak potensial lainnya yang belum diselidiki lebih lanjut. Penelitian ini diharapkan dapat mempelajari keragaman dan aktivitas mikroorganisme penghasil lipase dan kaitannya terhadap pembentukan rasa serta aroma pada tempe. Selain itu dalam jangka panjang diharapkan mikroorganisme lipolitik yang berasosiasi dengan tempe dapat dijadikan sebagai salah satu sumber baru untuk memperoleh enzim lipase yang ke depannya dapat diterapkan sebagai agen bioteknologi dan bioprospeksi.
Perumusan Masalah
1. Kelebihan tempe sebagai makanan hasil fermentasi multikultur adalah terdapatnya keragaman mikroorganisme di dalamnya beserta kelimpahan potensi yang ikut bersamanya.
2. Bakteri berperan penting dalam proses fermentasi tempe, kandungan lemak tempe yang jauh lebih rendah dibandingkan lemak pada biji kedelai menujukkan keberadaan bakteri lipolitik yang berasosiasi dengan tempe. 3. Bakteri lipolitik dikenal sebagai penghasil lipase yang menjanjikan dan
berperan sangat penting dalam industri bioteknologi.
4. Sejauh ini belum ada data mengenai dinamika bakteri lipolitik selama proses fermentasi tempe.
5. Keberadaan bakteri lipolitik ini juga diduga memberi rasa yang khas pada tempe selama proses fermentasi.
Tujuan Penelitian
Penelitian ini bertujuan untuk melakukan deteksi aktivitas bakteri lipolitik maupun total bakteri yang berasosiasi dengan tempe, membandingkan dinamika populasi bakteri terhadap populasi bakteri lipolitik pada tempe dan mendeteksi kandungan senyawa fatty acid methyl ester (FAME) pada tempe serta kaitannya dalam pembentukan aroma dan rasa.
Manfaat Penelitian
Hasil penelitian diharapkan memberikan informasi baru mengenai keragaman dan kepadatan populasi bakteri penghasil lipase yang berasosiasi pada tempe dan hubungannya dalam pembentukan rasa pada tempe. Manfaat prospeksi dari penelitian ini juga diharapkan dapat memberi data acuan dan referensi yang relevan dalam upaya menyelidiki potensi keberadaan bakteri lipolitik pada tempe sebagai agen bioprospeksi lipase di masa mendatang.
Ruang Lingkup Penelitian
TINJAUAN PUSTAKA
Tempe Sebagai Produk Fermentasi Asal Indonesia
Tempe merupakan makanan fermentasi tradisional paling popular di Indonesia yang terbuat dari bahan baku kedelai yang telah direbus kemudian dirangkai oleh jalinan-jalinan hifa halus seperti kapas dari kapang Rhizopus dalam suatu bentuk yang padat dan menyerupai kue berwarna putih (Shurtleff dan Aoyagi 1979). Tempe diperkirakan telah dikenal masyarakat Indonesia sejak tahun 1800an bahkan beberapa orang meyakini bahwa tempe telah ada jauh sebelum itu (Shurtleff dan Aoyagi 1979). Di Indonesia, tempe dikonsumsi oleh hampir semua tingkatan masyarakat hampir di seluruh Indonesia terutama di Jawa dan Bali. Penyajian kedelai menjadi tempe adalah unik dibandingkan dengan berbagai bentuk penyajian sebagai pangan yang lain. Keunikan tersebut ialah karena sebagai tempe, kedelai dikonsumsi utuh, berbeda dengan tahu atau susu kedelai misalnya, yang dikonsumsi hanya sebagai ekstrak protein saja
Gambar 1 Ilustrasi tempe kedelai, dua hingga tiga hari setelah fermentasi Selain jenis tempe kedelai ada jenis tempe yang lain, yakni tempe leguminosa non kedelai dan tempe non leguminosa. Tempe leguminosa non kedelai diantaranya adalah tempe benguk, tempe kecipir, tempe kedelai hitam, tempe lamtoro, tempe kacang hijau, tempe kacang merah dan sebagainya. Sedangkan jenis tempe non leguminosa diantaranya tempe gandum, tempe sorghum, tempe campuran beras dan kedelai, tempe ampas tahu, tempe bongkrek, tempe ampas kacang, dan tempe tela.
menggunakan kapang murni, laru juga dapat digunakan sebagai starter dalam pembuatan tempe. Ciri tempe yang “berhasil” adalah ada lapisan putih di sekitar kedelai dan pada saat di potong, tempe tidak hancur. Beberapa indikator ketidak”berhasil”an pada proses pembuatan tempe diantaranya adalah tempe tetap basah, jamur tumbuh kurang baik, tempe berbau busuk, ada bercak hitam dipermukaan tempe, dan jamur hanya tumbuh baik di salah satu tempat.
Komunitas Mikroorganisme pada Tempe
Tempe seperti yang telah diketahui sebelumnya adalah makanan tradisional di Indonesia yang dibuat melalui proses fermentasi, terutama menggunakan Rhizopus oligosporus. Selain R. oligosporus, kehadiran bakteri lain juga sangat penting dalam produksi Tempe, karena beberapa dari mereka berperan dalam peningkatan kualitas tempe. Pembuatan tempe secara tradisional, starter kultur biasanya disiapkan dari hasil sporulating fermentasi tempe sebelumnya dan oleh karena itu terdiri dari campuran ragi, cendawan dan bakteri, dengan Rhizopus sebagai genus dominan (Samson et al. 1987).
Smith et al. (1964) melaporkan bahwa tempe sebagai makanan asal Indonesia dibuat dengan memfermentasi kedelai menggunakan spesies dari Rhizopus (R. oligosporus, R. oryzae). Tempe sebagai makanan fermentasi memiliki kandungan protein yang tinggi. Tikus yang diberi makan dengan tempe menunjukkan penurunan kecil dalam pertumbuhan dan protein efisiensi dibandingkan dengan dengan yang diberi makan dengan bungkil dan kedelai mentah.
Barus et al. (2008) mengisolasi beberapa jenis bakteri pada media PCA yang diidentifikasi menggunakan gen 16S rRNA. Strategi yang digunakan untuk menentukan bakteri ini dilakukan berdasarkan sequent gen 16S rRNA. Bakteri dominan pada tempe yang teridentifikasi meliputi Acetobacter indonesiensis, Klebsiella pneumoniae, Bacillus subtilis, Flavobacterium sp., Klebsiella sp., Brevundimonas sp., Pseudomonas putida, Bacillus sp. dan Acinetobacter sp. Mereka juga mengisolasi jenis cendawan yang tumbuh di Tempe meliputi R. oligosporus, Mucor sp., dan Geotrichum candidum., dengan kelimpahan cendawan dari 3.0 x 105 CFUg-1 sampai 4.0 x 103 CFUg-1.
Pada pekerjaan mereka tentang vitamin B12 di Tempe, Keuth dan Bisping (1994) melaporkan munculnya Citrobacter freundii, Propionibacterium freudenreichii dan Bacillus cereus di Tempe disiapkan dengan tradisional direndam, nonautoclaved kedelai dapat dijelaskan oleh kondisi nonsterile proses fermentasi Tempe, yang meniru proses yang dilakukan di Indonesia. Hasilnya menunjukkan bahwa bakteri yang hadir selama perendaman kedelai dapat ditransfer ke SSF. Keluarga Enterobacteriaceae seperti Salmonella typhi, Shigella dysenteriae, K pneumoniae dan Yersinia pestis, galur E. coli dan C. freundii juga terdapat di Tempe
Mikroorganisme Penghasil Lipase
Tabel 1 Daftar komunitas mikroorganisme pada tempe
Golongan Nama spesies Literatur
Bakteri Enterobacteriaceae Keuth dan Bisping 1994 Lactic acid bacteria Samson et al. 1987 Staphylococcus aureus Samson et al. 1987
Bacillus cereus Barus et al. 2008
Escerichia coli Keuth dan Bisping 1994
Anaerob bacteria Samson et al. 1987 Citrobacter freundii Barus et al. 2008 Bacillus subtilis Barus et al. 2008 Acetobacter indonesiensis Barus et al. 2008
Klebsiella pneumonia Keuth dan Bisping 1994 Propionibacterium freudenreichii Keuth dan Bisping 1994
Salmonella typhy Keuth dan Bisping 1994
Shigella dysentriae Keuth dan Bisping 1994
Yersinia pestis Keuth dan Bisping 1994
Khamir Trichosporon beigelii Samson et al. 1987 Clavispora lusitaniae Samson et al. 1987 Candida intermedia Samson et al. 1987 Yarrowia lipolytica Samson et al. 1987 Lodderomyces elongisporus Samson et al. 1987 Rhodotorula mucilaginosa Samson et al. 1987
Candida sake Samson et al. 1987
Hansenula fabiani Samson et al. 1987 Candida tropicalis Samson et al. 1987 Candida parapsilosis Samson et al. 1987 Pichia membranaefaciens Samson et al. 1987 Rhodotorula rubra Samson et al. 1987
Candida rogusa Samson et al. 1987
Candida curvata Samson et al. 1987
Hansenula anomola Samson et al. 1987
Kapang Rhizopus oryzae Smith et al. 1964
Rhyzopus oligosporus Smith et al. 1964
Mucor indicus Barus et al. 2008
bioteknologi. Microbial lipase memiliki kemampuan enzimatis serta spesifitas pengenalan substrat yang sangat beragam sehingga menjadikan enzim ini sangat atraktif untuk penerapan industri (Arpigny dan Jaeger 1999). Dalam beberapa tahun terakhir ini penggunaan lipase meningkat sangat pesat dan merambah berbagai jenis industri terutama industri detergen dan kosmetik. Lipase kini perlahan-lahan mulai digunakan pula sebagai katalis biologi dalam pembuatan produk olahan makanan dan proses pembuatan bahan kimia yang lebih baik. Produksi lipase di industri pada umumnya dilakukan melalui teknologi DNA rekombinan. Penggunaan lipase menempati posisi ketiga sebagai enzim yang paling banyak digunakan di industri (setelah protease dan karbohidrase) berdasarkan total volume penjualannya yang mencapai angka penjualan hingga miliaran dolar (Jaeger et al. 1994)
Golongan Bakteri Penghasil Lipase
Dari banyak jenis bakteri penghasil lipase yang dieksploitasi, kebanyakan berasal dari anggota Bacillus yang menunjukkan sifat menarik sehingga menjadikan mereka calon yang tepat dalam aplikasi bioteknologi. Bacillus subtilis, Bacillus pumilus, Bacillus licheniformis, Bacillus coagulans, Bacillus alcalophilus dan Bacillus stearothermophilus merupakan bakteri penghasil lipase yang banyak digunakan. Selain itu, Pseudomonas sp., Pseudomonas aeruginosa, Burkholderia multivorans, Burkholderia cepacia, dan Staphylococcus caseolyticus juga dilaporkan sebagai bakteri produsen lipase.
Ertugrul et al. (2007) mengisolasi 17 galur bakteri yang bisa tumbuh pada media yang mengandung air limbah pabrik minyak (OMW) dan dipilih strain yang paling menjanjikan untuk produksi lipase. Setelah penapisan dalam media tributyrin agar, strain Bacillus sp. diidentifikasikan sebagai produser lipase terbaik. Setelah optimasi lebih lanjut, aktivitas intraselular yang ditemukan adalah 168 UmL−1. Kiran et al. (2008) mengisolasi 57 bakteri heterotrof dari marine spons Dendrodoris nigra, yang 37% menghasilkan zona bening di sekitar koloni-koloni di tributyrin agar untuk produser lipase. Pseudomonas MSI057 diamati besar zona bening di sekitar koloni-koloni. Kemudian, strain ini dipilih untuk studi lanjut, dan setelah optimasi, aktivitas lipase maksimum yang ditemukan sebesar 750 UmL−1.
Carvalho et al. (2008) mengisolasi galur bakteri dari tanah terkontaminasi oleh frompetroleum dan diberi kode sebagai Biopetro-4. Setelah penyelidikan dari beberapa inducers aktivitas lipase, nilai maksimal yang diperoleh adalah 1,675 UmL−1 setelah 120 jam fermentasi. Abada (2008) memproduksi lipase yang dihasilkan dari galur B. stearothermophilus AB-1 diisolasi dari udara dan memperoleh aktivitas lipase maksimum 1,585 UmL−1 dalam 48 jam fermentasi. Takaç dan Marul (2008) kultur mikrob yang diisolasi dari tanah yang diperkaya secara subkultur ditumbuhkan dalam kaldu nutrisi yang mengandung 1% (v/v) tributyrin. Proses isolasi sampel dilakukan melalui pengenceran berseri pada tributyrin agar (TBA) plate. Bacillus sp. dipilih berdasarkan pembentukan zona halo terbesar.
1,084 Ugds−1 menggunakan toleran pelarut P. aeruginosa galur PseA. Alkan et al. (2007) lipase ekstraseluler yang diproduksi oleh B. coagulans dan diperoleh aktivitas lipase maksimum 149 Ugds−1 setelah 24 jam fermentasi. Fernandes et al. (2007) memperoleh aktivitas lipase maksimum 108 Ugds−1 setelah 72 jam fermentasi oleh B. cepacia.
Tabel 2 Daftar bakteri penghasil lipase dari berbagai literatur
Nama spesies Literatur
Pseudomonas sp. Kiran et al. 2008
Pseudomonas aeruginosa Mahanta et al. 2008 Staphylococcus caseolyticus Vakhlu et al. 2006 Bacillus stearothermophilus Abada 2008
Burkholderia cepacia Fernandes et al. 2007
Burkholderia multivorans Gupta et al. 2003
Serratia rubidaea Ertugrul et al. 2007
Bacillus sp. Ertugrul et al. 2007
Bacillus coagulans Alkan et al. 2007
Bacillus subtilis Takaç and Marul 2008
Golongan Kapang Penghasil Lipase
Vargas et al. (2008) mempelajari produksi lipase oleh Penicillium simplicissimum dan memperoleh kegiatan 30 Ugds−1. P. verrucosum dan P. simplicissimum yang diisolasi dari industri minyak babassu.
Perbandingan kuantitatif antara SmF dan SSF sulit karena perbedaan dalam metode yang digunakan untuk menentukan aktivitas lipase. Sebagai contoh, lipase ekstraseluler diperoleh menggunakan Rhizopus homothallicus antara SSF dan SmF, masing-masing dengan aktivitas lipase 1.500 Ugds−1 dan 50 UmL−1 (Diaz et al. 2006). Azeredo et al. (2007) dengan mengkultur Penicillium restrictum masing-masing pada SSF dan SmF, memperoleh aktivitas lipase 17 Ugds−1 dan 12 UmL−1. Beberapa laporan mengenai imobilisasi cendawan dalam mengolah biomassa telah banyak diterbitkan. Imobilisasi menguntungkan karena dapat menghindarkan biomassa dari penurunan harga akibat tingginya faktor pengenceran. Konsentrasi tinggi sel dalam reaktor juga dapat dicapai dan pemisahan biomassa dari media (Elitol dan Ozer 2000). Wolski et al. (2008) melaporkan penggunaan respon permukaan metodologi untuk mengoptimalkan produksi lipase melalui fermentasi tenggelam menggunakan imobilisasi biomassa Penicillium SP. Pada kondisi eksperimental yang dioptimalkan, aktivitas lipase mencapai sekitar 21 UmL−1, lebih tinggi dari aktivitas yang diperoleh oleh mikroorganisme yang sama sebelum Imobilisasi.
Tabel 3 Daftar kapang penghasil lipase dari berbagai literatur
Nama spesies Literatur
Rhizopus arrhizus Yang et al. 2005
Rhizopus chinensis Wang et al. 2007
Aspergillus sp. Cihangir dan Sarikaya 2004
Rhizopus homothallicus Diaz et al. 2006
Penicillium citrinum D’Annibale et al. 2006
Penicillium restrictum Azeredo et al. 2007 Penicillium simplicissimum Vargas et al. 2008
Penicillium verrucosum Kempka et al. 2008
Geotrichum sp. Kempka et al. 2008
Geotrichum candidum Kempka et al. 2008
Aspergillus carneus Kaushik et al. 2006
Rhizopus sp. Bapiraju et al. 2005
Aspergillus niger Kaushik et al. 2006
Rhizopus oryzae Colen et al. 2006
Colletotrichum gloesporioides Colen et al. 2006
Candida utilis Rajendran et al. 2008
Candida rugosa Rajendran et al. 2008
Candida cylindracea D’Annibale et al. 2006
total biomassa R. arrhizus dan tingkat produksi lipase adalah konstan melalui beberapa eksperimen batch.
Golongan khamir penghasil lipase
Menurut Vakhlu dan Kour (2006), spesies utama dari ragi yang mampu menghasilkan lipase adalah: Candida rugosa, Candida tropicalis, Candida Antartika, Candida cylindracea, Candida parapsilopsis, Candida deformans, Candida curvata, Candida berlaku, Yarrowia lipolytica, Rhodotorula glutinis, Rhodotorula pilimornae, Pichia bispora, Pichia mexicana, Pichia sivicola, Pichia xylosa, Pichia burtonii, Saccharomycopsis crataegenesis, Torulaspora globosa, dan Trichosporon asteroid. Gen yang menyandikan lipase dalam Candida sp., Geotrichum sp., Trichosporon sp. dan Y. lipolytica telah dikloning dan diovereksprsikan (Wang et al. 2007). Meskipun lipase dari C. rugosa dan C. antartika telah secara ekstensif digunakan di berbagai bidang, ada beberapa publikasi terbaru yang melaporan produksi lipase oleh ragi lainnya.
Potumarthi et al. (2008) mengumpulkan sampel tanah asal laut dekat platform ekstraksi minyak di laut Arab. Setelah isolasi, koloni ditransfer ke cawan berisi tributyrin 2% dan diinkubasi pada suhu 35° C selama 3-4 hari. Koloni-koloni yang menunjukkan hidrolisis berupa zona bening terbesar diseleksi. Galur yang paling efektif untuk produksi lipase diidentifikasikan sebagai Rhodotorula mucilaginosa MTCC 8737 berdasarkan karakteristik fenotipiknya. Kumar dan Gupta (2008) mengisolasi 15 ragi dari minyak dan minyak sludge daerah di Delphi (India). Isolat tersebut yang dimurnikan dan diperiksa untuk potensi lipolitik mereka. Dari semua galur ragi, satu galur dipilih untuk studi lanjut, berdasarkan halo lipolisis terbesar. Berdasarkan urutan homologi, galur ini diidentifikasi sebagai Rhodotorula mucilaginosa dengan homologi 99% menurut database yang sudah ada.
Ciafardini et al. (2006) telah menemukan bahwa minyak zaitun yang baru diproduksi terkontaminasi oleh banyak mikro-flora, mampu mengkondisikan secara fisikokimia maupun organoleptik karakteristik minyak, melalui produksi enzim. Antara mikroorganisme yang diisolasi dari minyak ini, beberapa galur ragi diidentifikasi sebagai Saccharomyces cerevisiae, Candida wickerhamii, Williopsis eschscholzia, dan Candida boidinii, yang mana S. cerevisiae dan W. eschscholzia menunjukkan potensi yang baik untuk menghasilkan lipase. Aktivitas lipase dalam S. cerevisiae tercatat intraseluler dan ekstraseluler di W. eschscholzia. Proses ekstraksi minyak zaitun tiga fase menghasilkan limbah berwarna gelap, biasanya disebut air limbah pabrik minyak zaitun (OMW). D'Annibale et al. (2006) menyelidiki OMW valorization yang digunakan sebagai media pertumbuhan untuk produksi mikrob penghasil lipase ekstraseluler. Di antara galur yang diuji, galur yang paling menjanjikan adalah C. cylindracea dan Candida sp. merupakan produsen lipase paling potensial dari semua ragi yang dilaporkan dalam literatur.
substrat dan kondisi operasional. Berbeda dengan aktivitas tinggi dicapai dalam tulisan yang disebutkan di atas, Rajendran et al. (2008) melaporkan aktivitas lipase optimal C. rugosa hanya 3,8 UmL−1.
Tabel 4 Daftar khamir penghasil lipase dari berbagai literatur
Species Reference
Rhodotorula mucilaginosa Potumarthi et al. 2008 Rhodotorula mucilaginosa Kumar and Gupta 2008 Yarrowia lipolytica Ciafardini et al. 2006 Aureobasidium pullulans Ciafardini et al. 2006 Saccharomyces cerevisiae Ciafardini et al. 2006 Williopsis californica Ciafardini et al. 2006
Keberadaan Aktivitas Lipolitik pada Tempe
Lipase adalah biocatalysts serbaguna yang dapat melakukan banyak reaksi yang berbeda. Sifat mereka enantioselektif, chemoselective, dan stereoselektif menjadikannya sebagai alat penting dalam bidang sintesis organik. Tidak seperti glukosida yang bekerja dalam fasa air, lipase unik karena mereka bertindak di antarmuka minyak/air. Selain lipolitik, lipase juga memiliki aktivitas esterolytic dan dengan demikian memiliki berbagai berbagai substrat. Lipase dapat dilihat baik sebagai enzim lipolitik dan esterolytic, menjadi katalis untuk sejumlah besar Ester. Aktivitas lipase diuji melalui pemantauan pelepasan asam lemak atau gliserol dari trigliserida atau Ester asam lemak. Lebih jauh lagi, karena lipase bertindak pada interface minyak/air, perubahan dalam sifat-sifat interface (nomor kandungan lemak/minyak) adalah kriteria penting untuk penentuan aktivitas lipolisis.
Kandungan lemak pada biji kedelai
Kandungan lipid pada tempe adalah 61% lemak tak jenuh ganda dan 24% lemak tak jenuh tunggal yang sebanding dengan kandungan lemak tak jenuh total dari minyak nabati lainnya (~85%). Seperti minyak nabati lainnya, minyak kedelai tidak mengandung kolesterol.
Ahli gizi menyarankan membatasi konsumsi lemak maksimal 30% atau kurang dari jumlah kalori harian dan membatasi lemak jenuh maksimal 10% atau kurang. Asam lemak jenuh meningkatkan kolesterol darah yang dapat menebalkan dinding arteri dan meningkatkan risiko penyakit jantung. Dalam uji klinis dan studi populasi, lemak tak jenuh ganda dalam diet menunjukkan gejala aktif menurunkan kadar kolesterol. Penelitian lainnya yang dikumpulkan selama bertahun-tahun dari seluruh dunia telah menunjukkan bahwa populasi dengan diet rendah lemak jenuh memiliki tingkat kematian yang terendah. Akibatnya, penggantian lemak jenuh dengan jumlah yang wajar dari lemak tak jenuh ganda, seperti yang ditemukan dalam minyak kedelai sangat direkomendasikan.
prekursor untuk hormon yang mengatur kontraksi otot polos, tekanan darah, dan pertumbuhan sel-sel. Minyak kedelai murni mengandung sekitar 50% asam linoleat dan asam linolenat 8%.
Hidrogenasi digunakan untuk memperkuat minyak kedelai dalam pembuatan margarin. Proses ini meningkatkan stabilitas minyak dan untuk meningkatkan titik lebur. Hidrogenasi perubahan komposisi kimia dan sifat fisik dari minyak dan mempengaruhi nilai gizi. Tingkat perubahan dalam nilai gizi tergantung pada jumlah hidrogenasi yang diperlukan untuk menghasilkan produk akhir dan pengurangan polyunsaturates yang terjadi. Proses hidrogenasi juga menciptakan asam lemak trans dari cis unsaturates dengan mengatur hydrogens di sekitar ikatan ganda asam lemak.
Kandungan lemak pada tempe
Wagenknecht et al. (1961) melaporkan perubahan-perubahan yang dibawa oleh Rhizopus oryzae selama produksi tempe. Starter kultur memiliki aktivitas lipase kuat dan menyebabkan hidrolisis dari lebih dari sepertiga dari lemak netral tempe kedelai selama fermentasi. Lemak netral terdiri dari palmitic, stearat dan oleic, linoleic dan asam linolenic dengan asam linoleat mendominasi.
METODE PENELITIAN
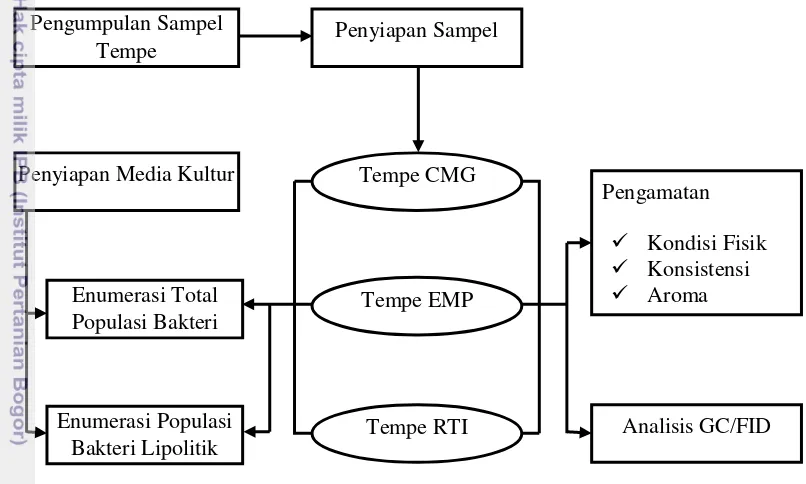
Kerangka Penelitian
Kerangka Penelitian di bawah (Gambar 2) meliputi pengumpulan sampel, teknik penghitungan koloni bakteri, analisis konsistensi dan aroma tempe, analisis dinamika kepadatan total populasi dan populasi bakteri lipolitik serta analisis senyawa asam lemak dengan metode GCFID.
Gambar 2. Diagram alir penelitian Waktu dan Lokasi Penelitian
Penelitian ini dilaksanakan dari bulan September 2014 hingga Juni 2015. Penelitian dilaksanakan di Laboratorium Mikrobiologi, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor. Analisis asam lemak menggunakan GCFID dilakukan di Laboratorium Kesehatan Daerah DKI Jakarata (LABKESDA DKI).
Observasi dan Pengumpulan Sampel
Sampel berupa makanan fermentasi tradisional asal Indonesia yaitu tempe kedelai. Sampel tempe yang akan digunakan dikumpulkan dari beberapa pengrajin tempe berdasarkan teknik pengumpulan sampel purposive sampling. Jumlah sampel tempe yang akan diambil untuk tiap lokasi tersebut masing-masing sebanyak minimal 100 gram dalam berat basah. Sampel tempe diambil sebanyak lima kali berdasarkan variasi usia inkubasi tempe yaitu 12 jam, 24 jam, 48 jam, dan 72 jam setelah tempe jadi (tempe segar).
Pengamatan Kondisi Fisik, Konsistensi dan Aroma Tempe
Pengamatan dilakukan pada beberapa rentang waktu sejak tempe menjadi tempe segar. Rentang pengamatan dimulai pada jam ke-0, jam ke-12, jam ke-24, jam ke-48 hingga jam ke-72. Pengamatan kondisi fisik tempe meliputi tekstur tempe (padat, lunak dan/atau berair) dan biji kedelai (warna dan tekstur) yang menyusunnya serta kondisi fisik dan jalinan hifa (warna, kepadatan dan sporulasi) yang mengikat tempe.
Media Kultur
Media selektif yang digunakan untuk mendeteksi keberadaan mikroorganisme penghasil lipase yaitu media agar-agar Luria Bertani yang diemulsikan bersama Olive oil dan Rhodamine B (Kumar et al 2012). Media ini tumbuh disesuiakan pH 7.0±2, dilakukan sterilisasi menggunakan autoklaf (121 0C, 15 menit) dan didinginkan hingga suhu 60 0C. Emulsi lipid dibuat sebanyak 30 ml dengan mensuspensikan secara aseptik 7.5 ml olive oil ke dalam 22.5 ml polyvinyl alcohol 2%, dihomogenkan dengan blender jar selama dua kali dua menit dengan jeda 30 detik tiap dua menit. 50 ml rhodamine B 0.1% dipersiapkan secara aseptic dan ditambahkan kedalam media tumbuh beserta dengan emulsi lipid. Tambahkan dH2O steril hingga volume akhir 250 ml. Cycloheximide 20 ppm ditambahkan untuk menekan pertumbuhan fungi secara umum.
Teknik Penghitungan Jumlah Total Koloni cawan menggunakan teknik cawan sebar untuk memperoleh data pengenceran optimum pada proses penghitungan jumlah koloni (CFU). Pengenceran terbaik selanjutnya digunakan untuk menghitung jumlah koloni bakteri pada masing-masing perlakuan. Tiap unit perlakuan dilakukan pengulangan sebanyak lima kali di luar kontrol pengamatan. Hasil pengenceran pada media cawan diinkubasi selama 24 – 48 jam pada suhu 38 0C (±1). Jumlah koloni yang terbentuk (colony forming unit /CFU) dihitung menggunakan persamaan;
�� −1=� ℎ � � �
Enumerasi Total Populasi Bakteri dan Populasi Bakteri Lipolitik
Jumlah total populasi bakteri dan bakteri lipolitik dihitung sesuai metode perhitungan jumlah total koloni dan menggunakan media kultur yang telah dijelaskan sebelumnya. Jumlah populasi total bakteri pada media PCA diamati secara langsung berdasarkan jumlah koloni tampak (viable cell) yang terbentuk (Cappucino dan Sherman 2001), sementara jumlah populasi lipolitik pada media LAOR diamati berdasarkan jumlah koloni yang membentuk zona berpendar apabila diamati melalui pencahayaan UV transiluminator. Masa inkubasi untuk penghitungan jumlah koloni kedua media dilakukan secara aerobik selama 24-48 jam pada suhu 38 0C (±1).
Analisis Metil Ester Asam Lemak Menggunakan GC-FID
Sebanyak 1 g tempe dihomogenkan bersama 10 ml H2O untuk memperoleh bubur tempe. Sebanyak 100 mg bubur tempe selanjutnya dilarutkan dalam methanol-benzen 4:1 (v/v). Esterifikasi dilakukan dengan menambahkan 200 µl asetil klorida perlahan-lahan sambal dikocok dengan vortex. Larutan yang terbentuk pada tabung diinkubasi selama 60 menit pada suhu 100 0C dengan perlakuan vortex tiap selang inkubasi 15 menit. Tabung didinginkan pada suhu 4 0C selama 10 menit, kemudian ditambahkan 5 ml larutan 6 % K2CO3 secara perlahan-lahan untuk menghentikan reaksi dan menetralkan larutan. Rotari evaporasi dilakukan selama 15 menit sebelum ditambahkan 4 ml heksan dan di rotari evaporasi kembali selama 30 menit. Ekstrak yang diperoleh selanjutnya disentrifugasi 3000 rpm selama 10 menit dan dilakukan pemisahan fasa organik. Analisi dilakukan dengan menyuntikkan 2 µl hasil esterifikasi asam lemak pada alat kromatografi gas (Agilent Technologies 6890) yang dilengkapi dengan Flame Ionizer Detector (FID), HP-Innowax, capillary column, 30 m × 0.250 mm dilapisi dengan film dengan ketebalan 0.25 µm. gas pembawa yang digunakan berupa hydrogen dengan split rasio 8:1. Suhu injektor dan detektor diatur pada 250 0C. suhu kolom dipertahankan pada 60 0C selama 30 detik selanjutnya dinaikkan hingga 210 0C selama 15 menit (Ansorena et al. 2001).
Analisis Statistik
HASIL DAN PEMBAHASAN
Survei dan Pengumpulan Sampel
Observasi awal pengumpulan sampel dilakukan pada beberapa pengrajin tempe yang tersebar di Kota Bogor. Observasi dilakukan untuk mencari produsen tempe dengan proses produksi dan bahan baku pembuatan serta metode pembuatan yang homogen. Hasil observasi selanjutnya menghasilkan tiga produsen tempe yang sesuai sebagai objek penelitian. Ketiga produsen tempe dipilih berdasarkan kriteria kebersihan selama produksi dan fermentasi tempe. Produsen tempe pertama merepresentasikan produksi tempe secara tradisional dan memiliki kadar kontaminasi mikrob paling tinggi yang selanjutnya disebut dengan tempe CMG, produsen tempe kedua juga merepresentasikan produksi tempe secara tradisional dan memiliki kadar kontaminasi mikrob cukup tinggi, selanjutnya disebut dengan tempe EMP dan produsen tempe terakhir merepresentasikan produksi tempe secara tradisional namun diolah secara modern melalui suatu control produksi tertentu sehingga memiliki kadar kontaminasi mikrob rendah dan selanjutnya disebut dengan tempe RTI. Sampel yang digunakan sebagai objek penelitian adalah tempe (tempe kedelai). Sampel tempe dikumpulkan melalui teknik purposive sampling, dengan beberapa kriteria yang telah ditentukan. Jumlah sampel yang dikumpulkan dari masing-masing sumber sesuai besaran kemasan yang diproduksi (>100 g). Pengambilan sampel dilakukan secara ketat dengan langsung memasukkan sampel kedalam plastik steril dan disimpan di dalam kotak pendingin.
Pengamatan Kondisi Fisik, Konsistensi dan Perubahan Aroma Tempe
Kondisi fisik awal dari semua sampel menunjukkan kondisi miselia yang putih bersih tanpa sporulasi dan terlihat kuat mengikat biji kedelai, sementara kondisi biji kedelai terlihat bening dengan tekstur yang padat (Gunawan-Puteri et al. 2015). Perubahan kondisi fisik tempe (miselia dan biji kedelai) paling cepat terjadi pada tempe CMG yaitu pada umur tempe jam ke-24 selanjutnya diikuti oleh tempe EMP pada jam ke-48 dan tempe RTI antara jam ke-48 dan jam ke-74. Tingkat kebersihan selama produksi tempe berkorelasi positif terhadap lama daya simpan tempe. Kebersihan selama proses produksi berpengaruh sangat besar terhadap jumlah mikroorganisme yang berhasil menginvasi tempe. Beberapa mikroorganisme memang telah diketahui memiliki peranan yang sangat besar dalam proses pembusukan makanan. Daya simpan tempe pada kondisi moderat secara umum hanya dapat bertahan beberapa hari saja (Babu et al. 2009).
(Tabel 5). Aroma kurang sedap terkait hidrolisis trigliserida umumnya terjadi pada senyawa asam lemak rantai pendek (C2-C4) yang umumnya dihidrolisis oleh enzim hidrolase golongan esterase.
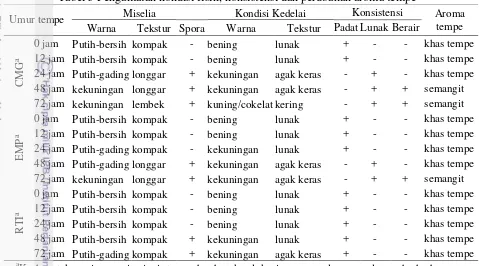
Tabel 5 Pengamatan kondisi fisik, konsistensi dan perubahan aroma tempe
Umur tempe Miselia Kondisi Kedelai Konsistensi Aroma
tempe
Warna Tekstur Spora Warna Tekstur Padat Lunak Berair
C
aKode untuk masing-masing jenis tempe berdasarkan lokasi pengumpulan sampel yang berbeda.
Penghitungan Jumlah Total Koloni
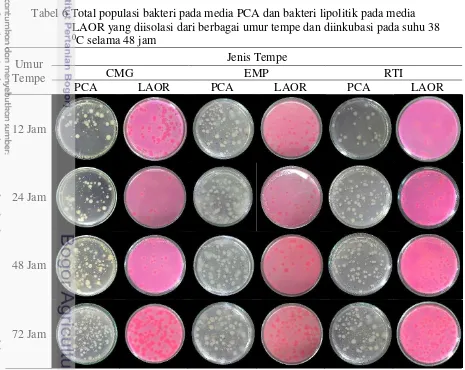
Jumlah koloni yang terbentuk dihitung untuk mendapatkan nilai kepadatan bakteri yang terkandung dalam tiap gram tempe (cfu g-1). Data enumerasi populasi total bakteri maupun bakteri lipolitik diperoleh setelah masa inkubasi cawan 24 jam dan 48 jam. Jumlah ulangan untuk tiap perlakuan sebanyak lima kali dengan keragaman yang homogen. Data enumerasi yang diperoleh dari semua ulangan dirata-ratakan untuk memperoleh jumlah populasi pada tiap titik pengamatan. Koloni total bakteri pada media PCA diidentifikasi sebagai semua jenis kenampakan bakteri yang terbentuk (Gambar 3). Koloni bakteri lipolitik tidak mencakup seluruh koloni namun hanya terbatas pada koloni yang menunjukkan zona berpendar jingga hingga merah di sekitar koloni (Gambar 3).
Gambar 3 Pengamatan total populasi bakteri pada media PCA (kiri) serta populasi lipolitik pada media LAOR tanpa penyinaran UV (tengah) dan dengan
Peristiwa lipolisis atau pemecahan lemak dapat diketahui melalui berbagai metode. Pengamatan secara visual melalui fenomena flouresensi (pendaran) merupakan metode yang paling efisien untuk digunakan pada pengamatan yang tidak membutuhkan adanya kuantifikasi (Jensen 1983), salah satu contohnya yaitu pengamatan pada media cawan. Pendaran yang terbentuk pada media LAOR disebabkan karena adanya reaksi antara pewarna rhodamine B terhadap asam lemak bebas yang terbentuk dari proses hidrolisis lemak (olive oil) oleh enzim lipase yang dihasilkan oleh bakteri lipolitik. Perubahan derajat keasaman akibat akumulasi asam lemak bebas menyebakan terjadinya penumpukan kation yang memicu terbentuknya kompleks uranyl-faty acid yang dapat berpendar jingga hingga merah jika dieksitasi dengan sinar pada penjang gelombang 350nm (Kouker dan Jaeger 1987). Kecerahan dan besarnya zona pendaran di sekitar koloni menunjukkan kemampuan lipolitik dari koloni tersebut. Mekanisme yang sama sebenarnya ditunjukkan pula oleh senyawa Victoria Blue B, dimana perubahan keasaman memicu reaksi perubahan warna secara langsung, namun fungsinya sebagai indikator terkadang menghasilkan hasil yang false positive terhadap asam yang dihasilkan dari hidrolisis karbohidrat (Alford dan Steinle 1967).
Tabel 6 Total populasi bakteri pada media PCA dan bakteri lipolitik pada media LAOR yang diisolasi dari berbagai umur tempe dan diinkubasi pada suhu 38 0C selama 48 jam
Umur Tempe
Jenis Tempe
CMG EMP RTI
PCA LAOR PCA LAOR PCA LAOR
12 Jam
24 Jam
48 Jam
72 Jam
Ket: CMG, EMP dan RTI menunjukkan inisial jenis tempe yang berbeda. PCA merupakan media
Enumerasi Total Populasi Bakteri
Hasil enumerasi (penghitungan) total populasi bakteri yang berasosiasi dengan tempe dilakukan secara manual. Teknik penghitungan dilakukan dengan mengidentifikasi dan menghitung keseluruhan koloni yang tumbuh pada media kultur. Media kultur yang digunakan dalam enumerasi total populasi bakteri yaitu media plate count agar (PCA). Media PCA merupakan media dengan kandungan nutrisi yang lengkap yang dapat mendukung pertumbuhan bakteri secara umum. Hasil enumerasi total populasi bakteri yang berasosiasi dengan tempe ditunjukkan pada tabel 7.
Tempe CMG merupakan jenis tempe yang memiliki kepadatan bakteri tertinggi yaitu mencapai 1.02 × 1010 cfu g-1, diikuti oleh tempe EMP sebesar 6.92 × 109 cfu g-1 dan yang paling rendah yaitu tempe RTI dengan kepadatan populasi sebesar 4.72 × 105 (Tabel 7). Jumlah total populasi bakteri tempe CMG pada jam ke-12 menunjukkan kepadatan populasi bakteri sebesar 2.31 × 109 cfu g-1. Kenaikan total populasi bakteri tempe CMG terjadi secara linear pada jam ke-24, jam ke-48 dan jam ke-72 berturut-turut sebesar 3.96 × 109 cfu g-1, 6.22 × 109 cfu Peningkatan jumlah kepadatan populasi terjadi secara signifikan setelah umur tempe jam ke-48 yaitu sebesar 3.26 × 109 cfu g-1 dan 6.92 × 109 cfu g-1 pada tempe jam ke-72.
Total kepadatan bakteri tempe RTI pada jam ke-12 hanya sebesar 4.04 × 105 cfu g-1 (Tabel 7). Jumlah tersebut jauh lebih kecil seperseribu kali dibanding tempe EMP dan CMG yang berada di kisaran 108 cfu g-1. Secara berturut-turut kenaikan total bakteri tempe RTI yaitu 5.72 × 105 cfu g-1 pada jam ke-24, 8.54 × 105 cfu g-1 pada jam ke-48 dan 1.26 × 106 cfu g-1 pada jam ke-72.
pematangan tempe selama fermentasi, keberadaan bakteri pada suatu makanan telah banyak dilaporkan terkait dalam proses pembusukan (Huis in’t Veld 1996; Gram et al. 2002)
Enumerasi Populasi Bakteri Lipolitik
Hasil enumerasi (penghitungan) populasi bakteri lipolitik yang berasosiasi dengan tempe juga dilakukan secara manual. Teknik penghitungan dilakukan dengan mengidentifikasi dan menghuitung keseluruhan koloni yang tumbuh pada media kultur. Media kultur yang digunakan dalam enumerasi total populasi bakteri yaitu media agar-agar Luria Bertani yang diemulsikan dengan rhodamine dan olive oil (LAOR). Rhodamine B pada media LAOR bertindak sebagai pewarna yang mendeteksi pelepasan asam lemak bebas akibat aktivitas lipolitik. Tabel 8 Penghitungan jumlah populasi bakteri lipolitik pada media LAOR yang
Kepadatan populasi bakteri lipolitik tempe CMG sebesar 1.04 × 106 cfu g-1 pada jam ke-12, menjadi hanya 7.7 × 105 cfu g-1 pada jam ke-24 (Tabel 8). Selanjutnya terjadi lonjakan kepadatan secara bertahap dengan kepadatan 1.18 × 106 cfu g-1 pada jam ke-48 dan 1.74 × 106 cfu g-1 pada jam ke-72. Kepadatan bakteri lipolitik pada tempe EMP jam ke-12 sebesar 2.88 × 105 cfu g-1 (Tabel 8). Kepadatan ini berangsur-angsur menurun pada tempe jam ke-24 dan jam ke-48 yaitu 2.18 × 105 cfu g-1 dan 1.88 × 105 cfu g-1. Pada saat tempe mencapai jam ke-72, kepadatan populasi lipolitik meningkat secara drastis mencapai 4.72 × 105 cfu g-1. Pada pengamatan jam ke-12, kepadatan populasi lipolitik hanya sebesar 1.5 × 104 cfu g-1 atau 100 kali lebih kecil bila dibandingkan dengan tempe CMG maupun EMP. kepadatan tersebut meningkat menjadi 2.82 × 105 cfu g-1, seterusnya melandai pada jam ke-48 sebesar 2.8 × 105 cfu g-1 dan 3.06 × 105 cfu g-1 pada jam ke-72 (Tabel 8). Pada pengamatan tempe RTI jam ke-12, kepadatan populasi lipolitik hanya sebesar 1.5 × 104 cfu g-1 atau 100 kali lebih kecil bila dibandingkan dengan tempe CMG maupun EMP. kepadatan tersebut meningkat menjadi 2.82 × 105 cfu g-1, seterusnya melandai pada jam ke-48 sebesar 2.8 × 105 cfu g-1 dan 3.06 × 105 cfu g-1 pada jam ke-72 (Tabel 8)
metil ester asam lemak banyak dilakukan oleh mikroorganisme penghasil enzim lipase. Mikroorganisme ini sering disebut mikroorganisme lipolitik. Pada tempe CMG dan EMP jumlah populasi bakteri lipolitik tergolong sangat tinggi sehingga proses perubahan aroma tempe berlangsung sangat cepat. Hal berbeda terjadi pada tempe RTI, meskipun terjadi perubahan aroma namun berjalan sangat lambat.
Dinamika Total Populasi Bakteri dan Populasi Bakteri Lipolitik
Dinamika total populasi bakteri pada tempe CMG secara umum menunjukkan kenaikan jumlah total populasi bakteri dari waktu ke waktu. Kepadatan populasi bakteri tempe CMG pada awal pengamatan tergolong cukup tinggi yang kemuadian meningkat secara linear selama waktu pengamatan. Pola dinamika total populasi bakteri yang ditunjukkan tempe EMP terlihat serupa dengan pola dinamika yang ditunjukkan oleh tempe CMG (Gambar 3). Perbedaan yang teramati dari kedua jenis tempe tersebut terletak pada kepadatan jumlah total populasi bakteri CMG yang berada pada kisaran lebih besar bila dibandingkan kepadatan total populasi bakteri tempe EMP. Pada awal pengamatan kepadatan populasi bakteri tempe EMP terlihat konstan (jam ke-12 hingga jam ke-24), kemudian meningkat secara drastis pada sisa waktu pengamatan. Meskipun menunjukkan pola dinamika yang sama terhadap kedua jenis tempe lainnya, namun kepadatan jumlah bakteri pada tempe RTI masih berada jauh di bawah level kepadatan bakteri pada tempe CMG maupun EMP (Gambar 3). Total kepadatan bakteri tempe RTI pada jam ke-12 hanya sebesar 4.04 × 105 cfu g-1 (Tabel 7). Jumlah tersebut jauh lebih kecil seperseribu kali dibanding tempe EMP dan CMG yang berada di kisaran 108 cfu g-1.
Gambar 4 Dinamika total populasi bakteri yang berasosiasi dengan tempe CMG (□), tempe EMP (∆) dan tempe RTI (○)
keadaan yang stabil dan menguntungkan, pertumbuhan mikrob terjadi secara cepat begitupula dengan proses pembelahan sel. Bakteri memperbanyak diri melalui mekanisme pembelahan biner. Pada kasus pembelahan biner, materi sel dari sel induk akan dibagikan sama banyaknya kepada kedua sel anakan hasil pembelahan. Ketika kondisi ingkungan menjadi sangat menguntungkan dan pertumbuha bakteri menjadi sangat cepat, maka jumlah kepadatan suatu bakteri pada lingkungan tertentu akan meningkat secra logaritmik dari waktu ke waktu.
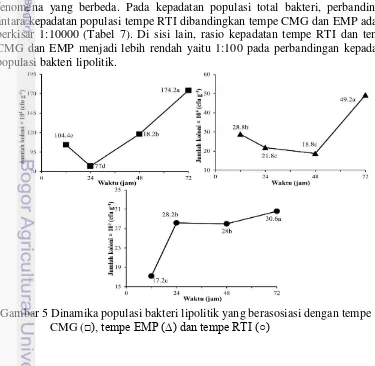
Kepadatan populasi bakteri lipolitik tempe CMG awalnya menurun dari pada jam ke-12 kemudian meningkat pada jam ke-24 (Gambar 4). Selanjutnya terjadi lonjakan kepadatan secara bertahap pada jam ke-48 jam ke-72 yang bahkan jauh lebih tinggi dibandingkan kepadatan populasi pada awal pengamatan. Pola yang hampir sama dengan tempe CMG ditunjukkan oleh dinamika populasi bakteri lipolitik yang berasosiasi dengan tempe EMP (Gambar 4). Pola dinamika yang pada awal pengamatan menunjukkan penurunan populasi bakteri lipolitik kemudian diikuti oleh peningkatan jumlah kepadatan populasi. Di lain sisi, tempe RTI menunjukkan pola dinamika lipolitik yang jauh berbeda dari kedua jenis tempe sebelumnya (Gambar 4). Pola dinamika tempe RTI menunjukkan peningkatan diawal pengamatan dan berangsur-angsur melandai pada pengamatan selanjutnya.
Faktor yang mempengaruhi perbedaan pola dinamika pada tempe RTI adalah kepadatan total populasi bakteri tempe RTI yang sejak awal memang sangat rendah jika dibandingkan dengan tempe CMG maupun EMP yang memiliki kepadatan populasi bakteri 1000 kali lebih banyak. Hasil perbandingan Rasio bakteri lipolitik terhadap total bakteri pada tempe RTI menunjukkan fenomena yang berbeda. Pada kepadatan populasi total bakteri, perbandingan antara kepadatan populasi tempe RTI dibandingkan tempe CMG dan EMP adalah berkisar 1:10000 (Tabel 7). Di sisi lain, rasio kepadatan tempe RTI dan tempe CMG dan EMP menjadi lebih rendah yaitu 1:100 pada perbandingan kepadatan populasi bakteri lipolitik.
Gambar 5 Dinamika populasi bakteri lipolitik yang berasosiasi dengan tempe
Pola dinamika populasi akan terjadi dalam berbagai pola yang berbeda-beda bergantung terhadap faktor pembatas pada lingkungan. Pada saat terjadinya keterbatasan nutrisi atau adanya penggunaan bersama suatu nutrisi pada suatu lingkungan, pertumbuhan mikrob tidak selamanya akan meningkat secara eksponensial. Lipid mengacu pada golongan senyawa hidrokarbon alifatik nonpolar dan hidrofobik. istilah lipid kadang-kadang digunakan sebagai sinonim dari lemak. Lipid juga meliputi molekul-molekul seperti asam lemak dan turunan-turunannya (termasuk tri-, di-, dan monogliserida dan fosfolipid, juga metabolit yang mengandung sterol, seperti kolesterol. Trigliserida (atau lebih tepatnya triasilgliserol atau triasilgliserida) adalah sebuah gliserida, yaitu ester dari gliserol dan tiga asam lemak. Trigliserida merupakan penyusun utama minyak nabati dan lemak hewani. Panjang rantai asam lemak pada trigliserida yang terdapat secara alami dapat bervariasi, namun panjang yang paling umum adalah 16, 18, atau 20 atom karbon. Lipase adalah enzim yang dapat larut dalam air dan bekerja dengan mengkatalisis hidrolisis ikatan ester dalam substrat lipid yang tidak larut air seperti trigliserida dengan asam lemak berantai panjang. Spesifitas lipase sangat bergantung dari jenis suatu lipase.
Beberapa mikroorganisme yang berbeda akan menghasilkan jenis lipase dengan spesifitas yang berbeda pula. Golongan bakteri lipolitik dengan lipase yang sesuai terhadap komponensubstrat yang dominan akan sintas terlebih dahulu dibandingkan bakteri lipolitik dengan spesisifitas substrat minoritas. Pada kasus ini bakteri lipolitik pada awal pengamatan akan sangat tinggi dan setelahnya menurun secara perlahan. Pada saat senyawa asam lemak dominan berangsur habis, maka kompetisi nutrisi akan berbalik pada bakteri dengan substrat minoritas dan mengutilisasi substratnya secara cepat. Pada tahaop inidinamika populasi bakteri akan kembali meningkat namun dengan keragaman bakteri yang jauh berbeda dibandingkan sebelumnya.
Hubungan Dinamika Populasi Lipolitik Terhadap Perubahan Aroma Tempe
Hubungan dinamika populasi bakteri lipolitik terhadap perubahan aroma pada tempe diamati pada tempe CMG dan EMP. Kedua jenis tempe ini menunjukkan pola dinamika yang sama yaitu terdapat fase jumlah populasi lipolitik yang menurun dan fase jumlah populasi lipolitik yang meningkat.
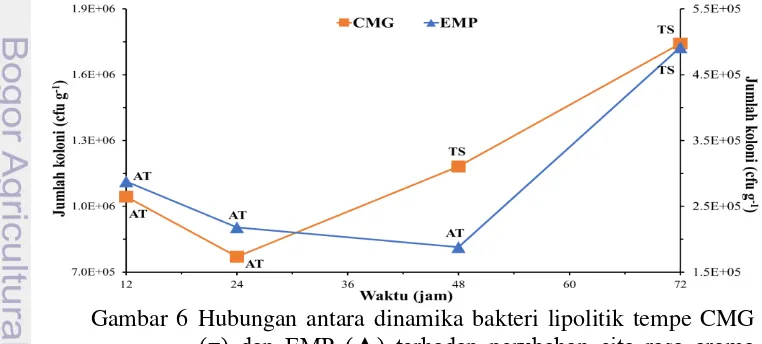
Tempe CMG mengalami penurunan jumlah populasi lipolitik pada jam ke-12 hingga jam ke-24 dan mengalami kenaikan jumlah populasi lipolitik pada jam ke-24 hingga jam ke-72 (Gambar 5). Terdapat suatu titik terjadinya pergantian pola dari populasi yang menurun menjadi populasi yang meningkat. Titik tersebut berada pada umur tempe jam ke-24. Hasil pengamatan sebelumnya menunjukkan bahwa pada jam ke-24 terjadi perubahan aroma tempe yang dari aroma khas tempe (AT) menjadi aroma tempe semangit (TS).
Tempe EMP menunjukkan pola dinamika lipolitik yang pada dasarnya hampir sama dengan tempe CMG. Penurunan jumlah populasi lipolitik terjadi pada jam ke-12 hingga jam ke-48 dan mengalami kenaikan pada jam ke-48 hingga jam ke-72 (Gambar 5). Titik pergantian pola untuk tempe EMP terjadi pada jam ke-48 yang secara otomatis menjadikan rentang waktu pola populasi yang menurun pada tempe EMP menjadi lebih panjang dibanding tempe CMG. Bersamaan dengan pergantian pola dinamika tempe EMP yaitu pada jam ke-48 terjadi pula perubahan aroma tempe dari aroma khas tempe menjadi tempe semangit.
Lemak atau lipid menjadi salah satu faktor penting dalm pembentukan aroma dan cita rasa tempe. Aroma didefenisikan berdasarkan hubungan antara respon sensorik terhadap jenis bau. Asam lemak bebas atau metil ester asam lemak (khususnya rantai sedang) berperan sangat esensial dalam pembentukan aroma (Ritvanen et al. 2005). Asam lemak bebas dihasilkan melalui proses lipolisis lemak. Peristiwa lipolisis salah satunya berupa reaksi kimia enzim lipase yang dihasilkan oleh mikroorganisme seperti bakteri lipolitik.
Berdasarkan data yang diperoleh terlihat adanya hubungan antara perubahan aroma dan dinamika populasi bakteri lipolitik. Fakta tersebut tidak memberi alasan yang kuat untuk menyatakan perubahan aroma yang terjadi adalah akibat peranan bakteri lipolitik. Perubahan aroma melibatkan komunitas dan banyak faktor yang ada dalam tempe. Bahkan jika diamati secara seksama, bukan tidak mungkin perubahan aroma yang terjadi merupakan faktor yang mempengaruhi pola dinamika bakteri lipolitik itu sendiri.
Analisis Kandungan Asam Lemak Tempe dengan Metode CGFID
Tabel 9 Profil kandungan asam lemak bebas dominan pada tempe yang dianalisis lemak bebas dominan yang menyusun per 100 gram ketiga jenis tempe yaitu asam linoleat dengan komposisi terbesar (3.51 - 10.29 g) dikuti oleh asam oleat (1.44 – 3.96 g), asam palmitat (0.91 – 2.53 g), asam γ-linolenat (0.31 – 1.24 g), asam linolenat (0.27 – 0.92 g) dan yang terendah asam stearat (0.32 – 0.78 g) (Tabel 9).
Gambar 7 Profil perbandingan komposisis asam lemak bebas dominan penyusun tempe CMG, EMP dan RTI yang diamati pada beberapa rentang waktu
Kandungan asam linoleat yang tinggi menjadi nilai tambah pada nutrisi tempe. Secara alamiah tubuh tidak dapat mensintesis asam lemak esensial, sehingga kebutuhan asam lemak ini umumnya diperoleh melalui asupan nutrisi. Asam linoleat merupakan salah satu jenis asam lemak esensial. Asam linoleat diketahui sangat berperan dalam membantu pertumbuhan, pemeliharaan membran sel, pengaturan metabolisme, menurunkan tekanan darah dan sebagainya. Dalam keadaan masih berupa biji kedelai tanpa fermentasi, komposisi asam lemak bebas pada biji kedelai sangat rendah karena umumnya masih berada dalam bentuk senyawa trigliserida. Masing-masing trigliserida yang terkandung pada biji kedelai membawa masing-masing tiga jenis asam lemak yang ketiganya bisa saja sama maupun berbeda satu sama lain. Berdasarkan analisis trigliserida, komponen umum penyusun rantai asam lemak pada biji kedelai berturut-turut didominasi oleh linoleat, oleat, palmitat, linolenat dan stearat (Diers dan Shoemaker 1992).
segi nutrisi sebagai sumber asam lemak bebas essensial maupun dari sisi bioprospeksinya karena mengindikasikan aktivitas lipase yang tinggi. Tempe RTI merupakan kebalikan dari tempe CMG, jumlah populasi yang rendah mengakibatkan melipahnya nutrisi yang terkandung pada tempe. Populasi bakteri yang ada di dalamnya akan lebih menggunakan karbohidrat sebagai sumber nutrisi paling utama sebelum beralih menggunakan asam lemak bebas yang tersedia. Hal tersebut menyebabkan tingginya kandungan asam lemak bebas pada tempe RTI.
Asam lemak bebas yang terkandung pada masing-masing tempe tidak hanya dipengaruhi oleh aktivitas lipolitik yang dilakukan oleh mikroorganisme di dalamnya. Komposisi lemak biji kedelai yang digunakan pada tiap-tiap jenis tempe juga menjadi factor yang mempengaruhi komposisi asam lemak bebas yang dihasilkan. Jenis kedelai yang berbeda akan memiliki kandungan lemak yang berbeda pula sehingga asam lemak yang dihasilkan selama proses lipolisis akan ikut berbeda. Selain jenis kedelai, modifikasi inokulum yang digunakan pada tempe CMG dan EMP juga dapat mempengaruhi aktivitas lipolisis selama fermentasi tempe.
Gambar 9 Perubahan komposisis asam lemak bebas dominan penyususun tempe CMG, EMP dan RTI yang diamati pada beberapa rentang waktu pengamatan
SIMPULAN DAN SARAN
Simpulan
Pola dinamika total populasi bakteri pada ketiga jenis tempe CMG, EMP dan RTI menunjukkan pola yang sama namun dengan kepadatan yang berbeda. Kepadatan populasi bakteri yang teramati mulai yang terbesar hingga terkecil berturut-turut tempe CMG, EMP dan RTI. Terdapat hubungan antara dinamika total populasi bakteri terhadap konsistensi dan perubahan aroma pada tempe. Dinamika populasi bakteri lipolitik pada tempe CMG dan EMP menunjukkan pola dinamika yang serupa namun tempe RTI tidak menunjukkan pola yang sama. Dinamika populasi tempe CMG dan EMP menunjukkan hubungan yang unik antara pola dinamika yang terbentuk terhadap konsistensi dan perubahan aroma pada tempe. Profil asam lemak tempe dominan terdiri atas asam linoleat, asam oleat, asam palmitat, asam γ-linolenat, asam linolenat dan asam stearat. Konsistensi senyawa asam lemak bebas pada tempe tidak menunjukkan adanya hubungan yang kuat terhadap pembentukan aroma khas tempe.
Saran
DAFTAR PUSTAKA
Abada EAE. 2008. Productastutuion and characterization of a mesophilic lipase isolated from Bacillus stearothermophilus AB-1. Pak J Biol Sci .11:1100– 1106.
Alford JA, Steinle EE. 1967. A double layered plate method for the detection of microbial lipolysis. J Appl Bacteriol. 30(3):488–494.
Alkan H, Baysal Z, Uyar F, Dogru M. 2007. Production of lipase by a newly isolated Bacillus coagulans under solid-state fermentation using melon waste. Appl Biochem Biotechnol. 136:183–192.
Ansorena D, Gimeno O. Astiasaran I, Bello J. 2001. Analysis of volatile compounds by GC-MS of a dry fermented sausage: chorizo de Pamplona. Food Res Intern. 34(1): 67-75. Doi 10.1016/S0963-9969(00)00133-2 Arpigny JL, Jaeger KE. 1999. Bacterial lipolytic enzymes, classification and
properties. Biochem J. 343:177-183.
Astuti M, Meliala A, Dalais FS, Wahlqvist ML. 2000. Tempe, a nutritious and healthy food from Indonesia. Asia Pacific J Clin Nutr. 9(4):322–325. Azeredo LAL, Gomes PM, Sant’Anna G, Castilho Jr LR, Freire DG. 2007.
Production and regulation of lipase activity from Penicillium restrictum in submerged and solid-state fermentations. Curr Microbiol. 54:361–365 Bapiraju KVVSN, Sujatha P, Ellaiah P, Ramana T. 2005. Sequential parametric
optimization of lipase production by a mutant strain Rhizopus sp. BTNT-2”, Braz J Chem Eng. 45(4):257–273.
Barus T, Suwanto A, Wahyudi AT, Wijaya H. 2008. Role of bacteria in tempe bitter taste formation microbiological and molecular biological analysis based on 16S rRNA gene. Microbiol Indones. 2(1):17-21.
Bavia ACF, da Silva CE, Ferreira MP, Santos Leite R, Mandarino JMG, Carrao-Panizzi MC. 2012. Chemical composition of tempe from soybean cultivars specially developed for human consumption. Ciênc. Tecnol Aliment Campinas. 32(3):613-620.doi: 10.1590/S0101-20612012005000085. Cappuccino JG, Sherman N. 2001. Microbiology A Laboratory Manual. New
York (US): Benjamin Cummings.
Carvalho NB, Souza RL, Castro HF, Zanin GM, Lima AS, Soares CML. 2008. Sequential production of amylolytic and lipolytic enzymes by bacterium strain isolated from petroleum contaminated soil. Appl Biochem Biotechnol. 150:25–32.
Ciafardini G, Zullo BA, Iride A. 2006. Lipase production by yeasts from extra virgin olive oil. Food Microbiol. 23:60–67.
Cihangir N, Sarikaya E. 2004. Investigation of lipase production by a new isolated of Aspergillus sp.. World J Microbiol Biotechnol. 20:193–197.
Colen G, Junqueira RG, Moraes-Santos T. 2006. Isolation and screening of alkaline lipase-producing fungi from Brazilian savanna soil. World J Microbiol Biotechnol. 22:881–885.