Latar Belakang
Dalam kurun waktu 14 tahun terakhir ini, pertanaman sayuran di Indonesia diinvasi oleh tiga hama eksotik yang tergolong genus Liriomyza (Diptera: Agromyzidae). Invasi diawali oleh L. huidobrensis (Blanchard) sekitar tahun 1994 (Rauf 1995), dan dua tahun kemudian diikuti oleh L. sativae Blanchard (Rauf et al. 2000). Jenis yang ke tiga adalah L. chinensis Kato yang dilaporkan pertama kali di Indonesia tahun 2000 (Rauf & Shepard 2001). Sebelumnya telah terdapat satu spesies lalat pengorok daun yang keberadaannya sudah lama di Indonesia, yakni Chromatomyiae horticola (Goureau) yang dikenal sebagai hama penting pada tanaman kacang kapri (Kalshoven 1981).
Asal lalat pengorok daun Liriomyza spp. adalah Amerika Selatan (Spencer 1973) dan diperkirakan masuk ke Indonesia melalui perdagangan bunga potong dan produk sayuran segar (Rauf 1995). Sekarang hama ini telah menyebar hampir ke seluruh wilayah sayuran di Indonesia, menjadi hama utama pada pertanaman sayuran dataran rendah sampai dataran tinggi. Menurut Rauf dan Shepard (1999), hama ini ditemukan pada 45 spesies tanaman dari Famili Cruciferae, Liliaceae, Cucurbitaceae, Umbelliferae, Compositae, Amaranthaceae, Chenopodiaceae, Solanaceae, Euphorbiaceae, Convolvulaceae, Basellaceae, dan Labitaceae.
Kerusakan yang ditimbulkan ketiga spesies hama ini adalah sama, yaitu kerusakan jaringan mesofil daun akibat aktifitas makan larva sehingga menyebabkan kemampuan tanaman dalam melakukan fotosintesis menurun (Minkenberg & van Lenteren 1986). Bekas tusukan ovipositor imago tampak berupa bercak-bercak putih pada daun, sedangkan korokan yang dibuat larva berbentuk seperti terowongan kecil yang berliku dan larvanya berada pada ujung liang. Serangan berat dapat mengakibatkan daun mengering dan gugur sebelum waktunya, sehingga menurunkan kuantitas dan kualitas hasil. Serangan hama ini dapat menurunkan hasil antara 30% sampai 70% (Rauf & Shepard 1999).
serangan karena lalat pengorok daun yang menyebar ini diduga berasal dari populasi yang telah resisten (Parrella & Keil 1984), dan karena larva berada dalam jaringan tanaman sehingga terlindung dari pengaruh insektisida (Parrella 1987). Di pihak lain, penggunaan insektisida yang berlebihan dikhawatirkan dapat menimbulkan dampak buruk seperti resistensi dan resurgensi hama, terbunuhnya musuh alami, dan pencemaran lingkungan secara umum (CEQ 1972).
Untuk mengatasi kerugian pada produksi pertanian dan kesulitan dalam pengendalian hama pengorok daun karena tidak efektifnya insektisida yang digunakan serta untuk mencegah atau mengurangi dampak penggunaan insektisida seperti matinya musuh alami, perlu dicari alternatif pengendalian hama tersebut.
Pendekatan Masalah
Di antara alternatif yang ada, pengendalian hama pengorok daun yang dapat dipilih adalah pemanfaatan musuh alami. Musuh alami merupakan salah satu komponen dalam pengendalian hama terpadu (PHT) yang aman terhadap lingkungan sehingga keberhasilan parasitoid dalam menekan populasi hama akan mengurangi penggunaan insektisida dalam pengendalian hama.
endoparasitoid larva-pupa (Rustam 2002), sedangkan H. varicornis merupakan ektoparasitoid larva. Informasi parasitoid H. varicornis sebagai agens hayati lalat pengorok daun di Indonesia telah banyak diteliti oleh Hidrayani (2003 ) dan Thamrin (2004).
O. chromatomyiae adalah endoparasitoid yang umum ditemui memarasit hama pengorok daun. Menurut Rauf dan Shepard (1999), tingkat parasitisasi parasitoid sangat dipengaruhi oleh tanaman inang dan ketinggian tempat. Oleh karena itu, survei lalat pengorok daun dan parasitoidnya, khususnya O. chromatomyiae, menjadi sangat berguna dalam pemanfaatan parasitoid tersebut sebagai agens pengendalian hayati. Lalat pengorok daun mudah beradaptasi dengan lingkungan sehingga penyebaran lalat pengorok cepat terjadi di lapangan. Penyebaran lalat pengorok daun akan diikuti oleh penyebaran parasitoidnya.
Dari survei pendahuluan yang dilakukan, bawang daun merupakan tanaman yang dominan terserang lalat pengorok daun dan pengorok ini menimbulkan kerugian yang besar. Informasi tentang dinamika populasi lalat pengorok daun maupun parasitoidnya pada bawang daun menjadi sangat penting untuk diketahui sehingga pengendalian lalat pengorok daun ini dapat dilakukan dengan efisien dan efektif.
O. chromatomyiae merupakan parasitoid yang diandalkan dapat mengendalikan lalat pengorok daun di daerah dataran tinggi. Namun dalam pemanfaatan di lapangan perlu kajian yang lebih dalam seperti bioekologi parasitoid dan hubungan parasitoid dengan tanaman inang, sehingga pelepasan di lapangan akan lebih tepat dan bermakna. Kajian bioekologi parasitoid seperti neraca kehidupan yang meliputi laju reproduksi, laju pertumbuhan instristik, persebaran umur stabil, dan waktu generasi serta kajian tanggap parasitoid terhadap kelimpahan inang (tanggap fungsional) menjadi sesuatu yang utama dalam pemanfaatan parasitoid ini.
insektisida dapat berakibat buruk bagi keberadaan musuh alami lalat pengorok daun (Johnson et al. 1980). Oleh karena itu, diperlukan penelitian tentang insektisida yang dapat mengendalikan lalat pengorok daun, tetapi aman terhadap musuh alami.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengidentifikasi spesies lalat pengorok daun pada pertanaman sayuran dataran tinggi serta parasitoid yang berasosiasi, (2) mengetahui tingkat parasitisasi parasitoid khususnya O. chromatomyiae pada berbagai jenis tanaman inang, (3) mengkaji dinamika populasi lalat pengorok daun dan parasitoidnya pada tanaman bawang daun (4) mengevaluasi selektifitas beberapa jenis insektisida terhadap parasitoid khususnya O. chromatomyiae (5) mengevaluasi potensi keefektifan parasitoid O. chromatomyiae berdasarkan parameter demografi yang meliputi laju pertambahan intristik (r), reproduksi bersih (Ro), dan masa generasi (T), proporsi usia stabil (px), dan (6) mengevaluasi potensi keefektifan parasitoid O. chromatomyiae berdasarkan tanggapnya terhadap peningkatan kelimpahan inang, serta menetapkan laju pelacakan inang (a) dan masa penanganan inang (Th).
Manfaat Penelitian
TINJAUAN PUSTAKA
Lalat Pengorok Daun
Menurut Parrella (1987), lalat pengorok daun mempunyai lebih dari 300 spesies yang sebagian besar tersebar di daerah beriklim sedang dan beberapa spesies ditemukan di daerah tropis. Spencer dan Steyskal (1986) menyebutkan terdapat tiga spesies lalat pengorok daun yang bersifat polifag yaitu Liriomyza huidobrensis (Blanchard), L. sativae Blanchard dan L. trifolii (Burgess).
Di Indonesia, lalat pengorok daun diperkirakan sudah ada sejak tahun 1990-an, namun pertama kali ditemukan dan diketahui sebagai hama sekitar bulan Sepetember 1994 di daerah Cisarua, Bogor (Rauf 1995). Tiga spesies yang telah dilaporkan banyak menimbulkan kerugian di berbagai daerah di Indonesia adalah L. huidobrensis, L. sativae dan L. chinensis (Rauf & Shepard 2001).
Liriomyza huidobrensis (Blanchard)
Lalat pengorok daun, L. huidobrensis berasal dari Amerika Selatan. Hama ini tersebar di Eropa, Afrika, dan Asia. Persebaran di Asia meliputi Malaysia, Indonesia, Filipina, Vietnam, Thailand, Srilangka, India, Pakistan, Laos, China, dan Banglades (Weintraub & Horowitz 1994; Rauf & Shepard 2001). Di Indonesia L. huidobrensis dilaporkan telah ditemukan di Jawa, Sumatra dan Sulawesi Selatan (Rauf 1995).
Imago L. huidobrensis berukuran 1.7-2.3 mm. Imago betina umumnya berukuran lebih besar dari imago jantan, dan muncul dari puparium yang berukuran lebih besar (Parrella 1987). Lama hidup imago betina berkisar antara 6-14 hari, sedang jantan 3-9 hari (Supartha 1998).
Liriomyza sativae Blanchard
Menurut Spencer (1973), negara asal L. sativae adalah Amerika. Di Indonesia, L. sativae telah tersebar di dataran rendah Jalur Pantura mulai dari Karawang sampai Brebes. Selain itu, L. sativae juga tersebar di daerah Bogor-Jawa Barat dan Banjar-Kalimantan Selatan (Rauf & Shepard 2001; Susilawati 2002).
Imago L. sativae berukuran kecil yaitu panjang 1.5 mm dengan warna hitam kecoklatan dan terdapat bintik kuning pada tubuhnya. Imago betina lebih besar dibandingkan dengan imago jantan. Lama hidup imago berkisar antara 10-20 hari tergantung pada kondisi lingkungan dengan kemampuan menghasilkan telur 600-700 butir (Mau & Kessing 1991; Capinera 2001).
Telur L. sativae bewarna putih, berbentuk lonjong dengan panjang 0.23 mm dan lebar 0.13 mm. Telur diletakkan satu per satu di dalam jaringan mesofil, di bawah permukaan daun. Tiga hari kemudian telur akan menetas menjadi larva. Larva terdiri dari tiga instar dan masing-masing instar memerlukan waktu pertumbuhan 2-3 hari (Mau & Kessing 1991; Capinera 2001). Larva instar akhir akan keluar dari daun dan menjatuhkan diri ke tanah untuk membentuk puparium (Parrella 1987). Periode puparium berlangsung selama 5-12 hari (Mau & Kessing 1991)
Liriomyza chinensis (Kato)
L. chinensis berasal dari daerah Asia bagian Timur (Taiwan dan Jepang) dan sekarang telah tersebar di Eropa (Spencer 1973; Dempewolf 2004). L. chinensis diduga masuk ke Indonesia melalui sayuran yang diimpor dari wilayah Asia bagian Timur (Rauf 2001).
Imago L. chinensis berukuran 2.00-2.39 mm. Imago betina mempunyai ukuran yang lebih besar dari imago jantan (Nawin 2003). Imago yang baru muncul bersifat fototaksis positif dan naik ke bagian atas tanaman (Parrella 1987). Lama hidup imago berkisar antara 8-12 hari dengan kemampuan menghasilkan telur 17-95 butir (Nawin 2003).
kekuningan yang mengecil pada bagian depan. Larva terdiri atas empat instar (Parrella 1987). Larva instar akhir akan keluar dari rongga daun untuk membentuk puparium di dalam tanah. Puparium berwarna kuning hingga coklat kekuningan (Nawin 2003).
Chromatomyia horticola Goureau
C. horticola tersebar di Eropa, Afrika Utara, Mesir, Libia, Maroko, Afrika Selatan, Madagaskar, Kamerun, Kenya, Turki, India, Cina, Jepang, Iran, Filipina, Malaysia, Korea dan Nepal (Spencer 1973; Dempewolf 2004). Hama ini telah lama dilaporkan keberadaanya di Indonesia (Kalshoven 1981).
Seekor betina C. horticola dapat menghasilkan 100-500 telur, dan setiap harinya mampu meletakkan 50 butir telur (Dempewolf 2004). Telur diletakkan oleh betina dalam permukaan atas dan permukaan bawah daun (Tandon 1965 dalam Dempewolf 2004). Telur menetas menjadi larva instar awal 1-4 hari setelah peletakan. Periode larva berkisar antara 5-6 hari. Larva mengorok daun secara tidak teratur.
Larva berkembang menjadi puparium dengan periode puparium selama 9-5 hari. Pupa tetap tinggal di dalam jaringan daun. Puparium bewarna abu-abu. Imago bewarna hitam dengan panjang sayap 2.2 – 2.7 mm (Dempewolf 2004).
Gejala serangan lalat pengorok daun dimulai pada daun bagian bawah, kemudian daun bagian tengah, dan yang lebih parah pada bagian pucuk (Cisneros & Mujica 2000). Chavez dan Raman (1987) melaporkan bahwa di Amerika Serikat dan Peru, serangan L. huidobrensis dapat mengakibatkan kehilangan hasil pada kentang sekitar 35% sedang di Lembang kehilangan hasil sekitar 34% (Soeriaatmadja & Udiarto 1996). Lebih lanjut dikemukakan oleh Rauf et al. (2000) bahwa berdasarkan hasil survei di Bandung dan Garut (Jawa Barat), Banjarnegara dan Wonosobo (Jawa Tengah), Alahan Panjang (Sumatera Barat), dan Karo (Sumatera Utara), petani setempat melaporkan kehilangan hasil akibat serangan Liriomyza pada tanaman kentang sekitar 30 – 70%.
Faktor yang Mempengaruhi Perkembangan Populasi Pengorok Daun Naik turunnya populasi organisme di suatu ekosistem dipengaruhi oleh faktor-faktor lingkungan fisik, inang atau sumber pakan, ruang populasi itu sendiri, dan spesies atau organisme lain. Faktor-faktor tersebut berperan dalam menentukan laju natalitas, mortalitas dan migrasi (Dent 1995).
Faktor lingkungan fisik antara lain adalah suhu dan kelembaban, baik pada tajuk tanaman maupun di permukaan tanah. Lama perkembangan Liriomyza lebih singkat pada suhu yang tinggi. Pada suhu 20 °C, lama perkembangan berkisar antara 23.7-27.0 hari, sedang pada suhu 25 °C berkisar antara 15.0-20.9 hari (Parrella 1987). Perkembangan L. trifolii dengan tingkat mortalitas rendah membutuhkan suhu optimum sekitar 25-30 °C (Leibee 1984). Pada tanaman krisan, imago yang keluar dari puparium pada suhu 15.6, 21.2, 26.7, 32.2, dan 37.8 °C masing-masing adalah 68, 80, 92.5, 75.5, dan 0 % (Minkenberg & van Lenteren 1986). Pada suhu 15 °C kematian L. trifolii tinggi terutama pada fase telur dan larva (Parrella 1987; Minkenberg 1988).
pada kelembaban 76-100% dapat dihasilkan puparium sebanyak 65-88% (Parrella 1987).
Perkembangan populasi Liriomyza spp. dipengaruhi oleh jenis, kuantitas, dan kualitas tanaman inang sebagai sumber makanan. Distribusi dan kerapatan trikoma, kandungan fenolat dan nutrisi tanaman mempengaruhi pemilihan tanaman inang oleh imago Liriomyza spp. (Fagoonee & Toory 1983). Kandungan N (nitrogen) daun tanaman inang berkorelasi positif dengan kelimpahan populasi Liriomyza di lapangan (Minkenberg & van Lenteren 1986; Parrella 1987). Pada kandungan nitrogen daun yang lebih tinggi, aktifitas makan dan keperidian Liromyza meningkat (Minkenberg & van Lenteren 1986). Ketersediaan berbagai jenis tanaman inang di lapangan selain membantu pertumbuhan dan perkembangan serangga, juga membantu pemencarannya. Selain itu, sifat polifag yang dimiliki Liriomyza spp. memungkinkan memencar lebih cepat (Supartha 1998).
Faktor spesies atau organisme lain meliputi pesaing dan musuh alami. Musuh alami Liriomyza yang banyak ditemukan adalah predator dan parasitoid (Minkenberg & van Lenteren 1986).
Liriomyza dapat dimangsa oleh berbagi jenis serangga predator (Coleoptera, Hemiptera, Diptera, Dermaptera, Hymenoptera) dan laba-laba (Cisneros & Mujica 2000; Harwanto et al. 2002). Hampir semua predator bersifat generalis, kecuali Dolichopodidae dan Empididae (Diptera) yang bersifat spesialis. Kemampuan predator dalam mempengaruhi perkembangan populasi Liriomyza sulit untuk dievaluasi (Cisneros & Mujica 2000).
Ragam spesies parasitoid Liriomyza spp. tidak selalu sama pada masing-masing jenis tanaman inang dan daerah geografis yang berbeda (Minkenberg & van Lenteren 1986). Terdapat 40 spesies parasitoid yang memarasit Liriomyza spp. pada fase larva dan puparium (Oatman 1959 dalam Supartha 1998), diantaranya ada empat famili yang berasosiasi dengan L. trifolii dan L. sativae pada tanaman tomat, yaitu Braconidae, Cynipidae, Eulophidae, dan Pteromalidae (Schuster & Wharton 1993).
Braconidae (Rauf et al. 2000; Susilawati 2002). Hemiptarsenus varicornis (Eulophidae) dan Opius chromatomyiae (Braconidae) merupakan spesies yang dominan ditemukan pada pertanaman sayuran dataran tinggi (Purnomo 2003).
Pengendalian Lalat Pengorok Daun
Teknik pengendalian yang umum dilakukan terhadap lalat pengorok daun di negeri asalnya adalah cara budidaya, kimiawi, dan pengendalian hayati. Cara budidaya yang biasa diterapkan adalah sanitasi, penyiangan gulma, pemangkasan bagian daun yang terserang dan pemasangan penutup tanah dari plastik hitam. Penggunaan perangkap kuning berperekat di rumah-rumah kaca juga hasilnya dinilai kurang memuaskan sehingga kurang layak dimasyarakatkan (Minkenberg & van Lenteren 1986). Walaupun demikian, cara pengendalian yang disebut terakhir di Peru dinilai efektif mengendalikan L. huidobrensis (Chaves & Raman 1987; Raman 1988; Raman & Radcliffe 1992).
Insektisida yang mengandung zat pengatur tumbuh (ZPT) serangga lebih efektif dibandingkan dengan insektisida lain yang berspektrum luas yang telah digunakan dalam usaha pengendalian Liriomyza spp. Dua jenis ZPT tersebut adalah Tripene ZR-619 dan Kinoprene ZP-777. Kedua ZPT tersebut dilaporkan sangat berbahaya bagi parasitoid penting seperti Opius dimiatus Ash. (Minkenberg & van Lenteren 1986).
ekonominya berdasarkan keadaan populasi lalat dan musuh alaminya di lapangan (Trumble 1985).
Pengendalian hayati Liriomyza spp. dengan menggunakan parasitoid telah banyak dilakukan. Semenjak tahun1980, pengendalian hayati itu telah dilakukan di Belanda pada areal seluas lebih kurang 30 ha per tahun. Dalam tahun 1985 dengan pelepasan sebanyak 10 000-20 000 per ha dengan perbandingan kira-kira satu parasitoid per empat tanaman, telah menunjukkan keberhasilan pengendalian Liriomyza spp. selama musim tanam. Dacnusa sibirica Telenga (Hymenoptera: Braconidae) diketahui efektif mengendalian L. trifolii di Belanda, sedangkan di Inggris dan Swedia, pengendalian dengan D. sibirica tidak berhasil (Minkenberg 1990). Hal ini dapat disebabkan oleh beberapa faktor, di antaranya adalah kemungkinan terjadinya imigrasi lalat pengorok, waktu pelepasan parasitoid yang tidak tepat, jumlah parasitoid yang dilepas tidak mencukupi dan mungkin kualitas parasitoid tidak bagus (Minkenberg 1990).
Faktor yang Mempengaruhi Keberadaan Musuh Alami
Agroekosistem tanaman semusim merupakan ekosistem ephemeral, yang berarti usianya singkat dan sering mengalami gangguan. Jenis gangguan yang sering terjadi pada agroekosistem adalah praktek budidaya misalnya pengolahan tanah, penyiangan, pemakaian insektisida dan pemanenan. Praktek budidaya tersebut secara ekologi sering tidak mendukung kehidupan musuh alami. Aplikasi insektisida menyebabkan musuh alami terbunuh, dan praktek budidaya bersih menyebabkan tanaman inang dan sumber daya tambahan berkurang ketersediannya (DeBach 1973). Keadaan tersebut pada akhirnya berdampak terhadap agroekosistem (Altieri & Whitcomb 1979).
Pemanfaatan parasitoid sebagai agens pengendalian hayati hama dapat diterapkan melalui metode konservasi dan augmentasi inundatif. Metode konservasi diarahkan untuk mempertahankan pelepasan musuh alami agar tetap lestari, sedangkan augmentasi inundatif merupakan pelepasan musuh alami dalam jumlah yang relatif besar dengan harapan segera dapat mengendalikan populasi hama dalam waktu yang singkat (Sosromarsono 1999). Sebelum metode tersebut dilaksanakan, terlebih dahulu harus diketahui karakter biologi, perilaku, serta kondisi ekologis yang diperlukan agar musuh alami dapat bekerja dengan baik di lapangan.
Pemilihan Inang oleh Parasitoid
Sebagian besar parasitoid Hymenoptera dapat memarasit beberapa jenis inang dan hanya sedikit spesies yang spesifik memarasit satu spesies inang. Parasitoid yang spesifik tersebut, pada kondisi laboratorium bahkan juga sering dapat dipelihara pada inang lain yang secara alamiah bukan merupakan inang karena adanya hambatan waktu dan ruang yang memisahkannya. Kenyataan bahwa parasitoid dapat dibiakkan di laboratorium dengan serangga bukan inang alamiah menjadi penting dalam pembiakkan masal parasitoid (Doutt 1959).
Menurut Doutt (1959), terdapat empat tahapan yang harus dilewati agar parasitoid berhasil memarasit inangnya, yaitu 1) penemuan habitat inang, 2) penemuan inang, 3) penerimaan inang, dan 4) kesesuaian inang. Selanjutnya Vinson (1981) menambahkan pengaturan inang sebagai tahap yang kelima karena keberhasilan parasitisme juga ditentukan oleh kemampuan parasitoid dalam mengatur fisiologi inangnya.
Penemuan inang oleh parasitoid dipandu oleh rangsangan fisik dan kimia yang dikeluarkan oleh inang. Rangsangan fisik yang berperan terutama suara dan gerakan. Rangsangan kimia dapat dibagi menjadi dua kelompok. Pertama, rangsangan kimia yang dapat diterima dari jarak jauh misalnya bau inang. Rangsangan yang diterima memungkinkan parasitoid untuk melokalisasi areal pencarian inang. Kedua, rangsangan kimia yang dapat dideteksi hanya dari jarak dekat, yaitu setelah terjadi kontak fisik. Rangsangan ini biasanya berasal dari senyawa-senyawa padat atau cair misalnya kotoran inang, sekresi dari kelenjar labium inang, produk inang lain dan bekas parasitoid lain. Adanya rangsangan ini memungkinkan terjadinya kontak antara parasitoid dengan inangnya yang dicirikan oleh perilaku pengujian oleh parasitoid berupa pergerakan memutar dengan cepat dan perubahan kecepatan pergerakan. Faktor lain yang ikut menentukan penemuan inang adalah pengalaman dan perilaku orientasi parasitoid (Weseloh 1981).
Penerimaan inang atau pengenalan inang adalah proses diterima atau ditolaknya inang untuk peletakkan telur setelah terjadi kontak (Arthur 1981). Proses tersebut dibagi dalam empat fase yaitu: 1) kontak dan pemeriksaan, 2) penusukan dengan ovipositor, 3) pemasukan ovipositor dan 4) peletakan telur. Keempat fase tersebut harus lengkap dan berurutan sehingga bila terjadi hambatan pada salah satu fase, proses dimulai lagi dari awal.
Kesesuaian inang yang menentukan keberhasilan perkembangan parasitoid sampai menjadi imago tergantung pada beberapa faktor, yaitu: 1) kemampuan parasitoid dalam menghindari atau melawan sistim pertahanan inang, 2) kompetisi dengan parasitoid lain, 3) adanya toksin yang mengganggu atau merusak telur atau larva parasitoid, dan 4) kesesuaian makanan parasitoid. Faktor lain yang berpengaruh adalah infeksi patogen, kerentanan inang, faktor lingkungan dan pengaruh hormon-hormon pengendali serangga (Vinson & Iwantsch 1980).
Biologi Opius sp.
Parasitoid Opius sp. merupakan endoparasit larva-pupa. Studi biologi dan morfologi O. dissitus telah diteliti pada inang L. trifolii oleh Bordat et al. (1995), O. melleus Gahan pada larva lalat buah Famili Tephritidae (Lathrop & Newton 1933) dan Opius sp. pada inang L. huidobrensis (Rustam 2002).
Telur O. dissitus berbentuk lonjong dengan warna putih dan tembus pandang. Ukuran telur rata-rata 0.22 mm dan stadium telur 1-3 hari (Bordat et al. 1995), sedangkan stadium telur O. melleus lebih lama yakni 3-6 hari (Lathrop & Newton 1933). Pada inang L. huidobrensis, stadium telur Opius sp. adalah 2 hari dengan ukuran telur lebih besar dibandingkan dengan O. dissitus yakni 0.26 mm (Rustam 2002). Setelah telur diletakkan, telur mengalami pertumbuhan dan perkembangan di dalam tubuh inang.
Larva O. dissitus terdiri atas dua instar, dengan ukuran masing-masing 0.47 mm dan 0.99 mm, sedangkan O. melleus, Diachasma tryoni (Hymenoptera: Braconidae), larva terdiri atas empat instar (Pemberton & Willard 1918; Lathrop & Newton 1933; Bordat et al. 1995). Larva O. chromatomyiae terdiri atas dua instar dengan ukuran yang lebih besar dibandingkan dengan larva O. dissitus dan O. melleus (Rustam 2002). Larva instar satu mempunyai bagian kapsul kepala yang kokoh dengan dua pengait yang runcing, tubuhnya ramping dan pada ujungnya terlihat kasar. Larva instar dua berbentuk bulat, dimana pengaitnya telah hilang dan berada dalam puparium inang yang panjang dengan warna putih krem (Bordat et al. 1995; Rustam 2002) dan bersifat tidak aktif (Lathrop & Newton 1933).
kepala, toraks dan abdomen dapat terlihat jelas. Pada awalnya pupa berwarna kuning pucat dan lama kelamaan berwarna gelap. Tubuh pupa yang berumur lanjut berwarna hitam dengan ukuran 1.62 mm (Bordat et al. 1995; Rustam 2002), pembentukan pupa terjadi di dalam puparium inang (Pemberton & Willard 1918) dan masa perkembangan pupa berlangsung selama 5.93 hari (Rustam, 2002). Imago O. chromatomyiae berwarna coklat kehitaman dengan ukuran yang hampir sama antara jantan dan betina, yaitu rata-rata 1.72 mm dan 1.80 mm. Jantan dan betina sulit dibedakan dengan mata biasa, namun dengan bantuan kaca pembesar terlihat betina mempunyai ovipositor sebagai alat untuk peletakan telur (Rustam 2002). Antena imago O. dissitus panjang, hitam, dan panjangnya hampir sama dengan tubuhnya (Bordat et al. 1995). Pada O. chromatomyiae jantan dan betina sudah bisa dibedakan berdasarkan jumlah ruas antenanya dimana antena jantan terdiri atas 23 ruas sedangkan betina terdiri atas 25 ruas (Rustam 2002). Pada O. melleus ovipositor relatif panjang dan merupakan ciri khasnya (Lathrop & Newton 1933).
Imago parasitoid keluar dengan cara merobek puparium inang. Opius betina yang baru muncul dapat melakukan parasitisasi dengan menusukkan ovipositornya ke dalam rongga tubuh larva inang tanpa mematikan atau melumpuhkannya. Larva inang terparasit tetap hidup dan mengorok daun hingga menjadi pupa. Parasitoid tetap berkembang di dalam tubuh inang sampai keluar menjadi imago (Bordat et al. 1995; Rustam 2002). Clausen (1940) melaporkan bahwa imago betina O. fulvicornis dapat langsung meletakkan telur setelah keluar dari puparium inang.
Menurut Rustam (2002), umumnya imago jantan muncul lebih awal dibandingkan imago betina. Sebanyak 85.71% dari imago yang muncul pada hari pertama adalah jantan. Lebih lanjut Rustam (2002) menjelaskan bahwa seekor imago betina mempunyai keperidian berkisar antara 49-187 butir telur dengan nisbah kelamin keturunan yang bias betina yakni sebesar 73.47 %.
Parameter Pertumbuhan Populasi
(1997), menyatakan bahwa dari angka kelahiran dan angka kematian dapat disusun suatu neraca hidup yang menjelaskan perubahan–perubahan kuantitatif dari suatu populasi selama satu generasi.
Ada dua tipe neraca kehidupan yaitu yang bersifat spesifik umur (age spesific) atau tabel kehidupan horizontal, serta yang bersifat spesifik waktu (time spesific) atau tabel kehidupan vertikal. Neraca kehidupan yang berspesifik waktu menganalisis data yang diambil pada suatu kejadian tunggal, ketika diasumsikan bahwa semua generasinya sudah saling lingkup dengan sempurna, oleh karena itu kelas umur secara simultan sama. Neraca kehidupan yang bersifat spesifik umur mencakup perhitungan yang berulang terhadap suatu kelompok (cohort) tunggal yang terdiri dari individu yang sama umurnya sepanjang waktu. Tabel ini sering digunakan dalam entomologi (Bellow & van Driesche 1992).
Menurut Poole (1974), perumusan neraca kehidupan merupakan langkah pertama dalam menghitung laju pertumbuhan intrinsik (r). Dua data utama yang dibutuhkan dalam perhitungan tersebut adalah lx, peluang bertahan pada umur x; dan mx, rataan jumlah keturunan betina yang dihasilkan dalam satu unit waktu oleh seekor betina berumur x (Birch 1948). Disamping nilai r, dari data tersebut dapat juga dihitung laju reproduksi kotor (GRR), laju reproduksi bersih (Ro), dan waktu generasi (T) (Birch 1948; Price 1997).
Neraca kehidupan merupakan riwayat perkembangan cohort yang bersifat dinamis, mulai dari umur 0 sampai umur dimana semua individu dalam populasi mati. Dari neraca kehidupan dapat diketahui laju reproduksi bersih (Ro), lama waktu generasi (T) dan laju pertambahan instrinsik (r) (Tarumingkeng 1992).
Keefektifan musuh alami dalam mengendalikan hama sasaran biasanya diukur berdasarkan beberapa ciri biologinya, di antaranya adalah (a) kemampuan mencari inang yang tinggi, terutama pada saat kelimpahan inang rendah, (b) kekhususan terhadap inang tertentu, (c) potensi reproduksi yang tinggi, (d) kisaran toleransi terhadap lingkungan yang lebar serta kemampuan memarasit berbagai instar inang (DeBach 1973).
kelamin (Carey 1993). Nilai r merupakan salah satu kriteria yang penting untuk mengevaluasi keefektifan atau potensi dari agens pengendalian hayati (Lee & Ahn 2000), serta dapat digunakan untuk menduga potensi pertumbuhan populasi musuh alami (Lysyk 2000).
Respon Pemangsaan
Preferensi makanan, selera parasitoid terhadap inangnya, kerapatan inang, kualitas makanan dan adanya makanan alternatif merupakan faktor lingkungan yang berperan dalam menentukan laju pemangsaan oleh suatu predator terhadap mangsanya atau parasitoid terhadap inangnya. Solomon (1949 dalam Tarumingkeng 1992) mengemukakan teori mengenai respon pemangsaan terhadap kerapatan inang dalam dua tipe, yaitu tanggap fungsional dan tanggap numerikal.
Tanggap fungsional merupakan salah satu ukuran untuk menentukan keefektifan suatu predator atau parasitoid sebagai agens dalam pengendalian hayati (Doutt 1973). Menurut Solomon (1949 dalam Hassel 2000) tanggap fungsional merupakan perubahan jumlah mangsa yang diserang oleh individu predator/parasitoid akibat perubahan kepadatan populasi mangsa per satuan waktu.
Holling (1959 dalam Price 1997) mengkategorikan tanggap fungsional menjadi tiga tipe: 1) Tipe respon fungsional linear (Tipe I), laju pemangsaan oleh predator meningkat atau menurun sebanding dengan kerapatan mangsa dan mencapai tahap kejenuhan. Tipe I ini biasanya ditemukan pada predator yang bersifat pasif seperti laba-laba, 2) Tipe respon fungsional hiperbolik (Tipe II), laju pemangsaan secara progresif semakin menurun. Respon ini sangat umum di antara serangga-serangga predator dan parasitoid dan 3) Tipe respon fungsional sigmoid (Tipe III), perubahan pemangsaan berlangsung lambat, diikuti dengan peningkatan kemudian mendatar (asimptotik).
banyak dilakukan seperti yang diteliti oleh Wang dan Ferro (1998); Buchori (2003); Hidrayani (2003).
Respon pemangsaan yang kedua adalah tanggap numerik, yaitu pemangsaan menyebabkan perubahan kerapatan predator atau parasitoid pada suatu luasan pemangsaan tertentu. Ada dua mekanisme yang mendorong terjadinya tanggap numerik yaitu 1) peningkatan laju reproduksi predator atau parasitoid bila mangsa berlimpah dan 2) keterpikatan predator pada tempat begerombolnya mangsa, dikenal juga dengan tanggap agregasi.
Hubungan antara Inang, Parasitoid, dan Tanaman
Perilaku parasitoid terhadap inangnya tidak hanya dipengaruhi oleh senyawa-senyawa kimia yang dihasilkan oleh serangga inang, tetapi juga oleh makanan inang itu sendiri (Vinson 1984). Menurut Takabayashi et al. (1998), parasitoid merupakan musuh alami yang agak khusus yang menyerang sejumlah spesies herbivora tertentu. Kekhususan itu tidak hanya tergantung pada herbivoranya, tetapi juga tanaman inangnya. Parasitoid mungkin hanya dapat menemukan inang yang makan pada satu atau beberapa spesies tanaman saja, sedangkan diketahui bahwa serangga tersebut mempunyai kisaran inang yang luas.
Tingkat parasitisasi dan distribusi beberapa parasitoid pengorok daun tidak merata pada berbagai tanaman inang, dan berbeda-beda menurut jenis tanaman dan lokasi ditemukan. Selain itu parasitoid Liriomyza spp. dapat dikelompokkan sesuai dengan spesies serangga inang, tanaman inang, dan daerah geografis tempat tanaman inang tumbuh (Johnson & Hara 1987; Rauf et al. 2000).
Di Indonesia, parasitoid yang dominan memarasit L. huidobrensis adalah H. varicornis dan O. chromatomyaie (Hidrayani 2003; Purnomo 2003), sedangkan spesies parasitoid yang dominan memarasit L. sativae adalah Acecodes deluchii (Rauf et al. 2000; Susilawati 2002; Tapahillah 2002). Di California, spesies parasitoid yang dominan menyerang L. sativae adalah Chrysocharis parksi ( Zehnder & Trumble 1984).
ditemukan pada tanaman bawang daun. Distribusi parasitoid Liriomyza spp. pada berbagai tanaman inang tidak merata. Pengendalian biologi yang efektif memerlukan adanya kesesuaian antara spesies parasitoid dengan serangga inang dan tanaman inangnya.
Aplikasi Insektisda dalam Pengendalian Liriomyza
Penggunaan insektisida sintetik sangat beresiko karena dapat menimbulkan dampak negatif, khususnya terhadap parasitoid sebagai serangga musuh alami lalat pengorok daun. Menurut Purnomo (2003), jika insektisida terpaksa digunakan, hendaknya insektisida tersebut bersifat selektif dan tidak berdampak negatif bagi aktifitas musuh alami.
Penggunaan insektisida kimiawi seperti organoklorin, organofosfat, dan karbamat yang berspektrum luas sudah sejak lama direkomendasikan sebagai cara pengendalian hama, walaupun cara ini dinilai terlalu berbahaya. Penggunaan insektisida telah banyak memacu ledakan hama sekunder, karena Liriomyza menjadi resisten terhadap sejumlah insektisida yang digunakan. Oleh sebab itu penggunaan insektisida harus atas hasil pengamatan agroekosistem untuk mengetahui ambang ekonominya berdasarkan keadaan populasi lalat dan musuh alaminya (Cardova & Karel 1990).
Penggunaan bioinsektisida merupakan salah satu alternatif yang tepat untuk mengurangi dampak negatif yang ditimbulkan oleh insektisida sintetik. Menurut Copping dan Menn (2000), istilah bioinsektisda mencakup berbagai bahan pengendalian hama seperti: mikroorganisme (virus, bakteri, cendawan), nematoda entomopatogen, insektisida botani, metabolit sekunder yang dihasilkan mikroorganisme, dan feromon serangga. Penelitian mengenai pemanfaatan bahan-bahan alami untuk perlindungan tanaman semakin gencar dilakukan selama tiga dekade terakhir. Hingga saat ini terdapat tiga sumber insektisida alami yang penting dan memiliki prospek yang baik untuk dikembangkan lebih lanjut, yaitu tumbuhan, mikroorganisme tanah, dan organisme laut (Prijono 1999).
perkembangan larva, mengurangi terbentuknya puparium, menyebabkan pergantian kulit abnormal pada Musca autumnalis (Diptera: Muscidae) (Schmutterer 1995).
1. LALAT PENGOROK DAUN DAN PARASITOIDNYA PADA
PERTANAMAN SAYURAN DATARAN TINGGI DI
KABUPATEN CIANJUR-BOGOR, JAWA BARAT
Abstrak
Lalat pengorok daun Liriomyza spp. adalah hama yang banyak menimbulkan kerusakan pada berbagai pertanaman sayuran dataran tinggi di Indonesia. Penelitian ini bertujuan untuk mengidentifikasi spesies Liriomyza yang menyerang pertanaman sayuran dataran tinggi, parasitoid yang berasosiasi dengan pengorok dan tingkat parasitisasi Opius chromatomyiae pada pertanaman sayuran dataran tinggi. Survei dilakukan dengan mengambil 20-40 helai daun tanaman sayuran yang mempelihatkan gejala korokan lalat pengorok daun. Imago lalat pengorok daun dan parasitoid yang muncul dari daun contoh diidentifikasi. Parasitisasi dihitung dengan membandingkan jumlah parasitoid dan imago lalat pengorok daun. Terdapat empat spesies lalat pengorok daun yang menginfestasi pertanaman sayuran dataran tinggi, yakni Liriomyza huidobrensis, L. chinensis, L. sativae, dan Chromatomyia horticola. Diketahui delapan spesies parasitoid yang berasosiasi dengan lalat pengorok daun, enam spesies dari famili Eulophidae, yaitu Hemiptarsenus varicornis, Asecodes deluchii, Neochrysocharis okazakii, N. formosa, Neochrysocharis sp., Quadrastichus liriomyzae dan masing-masing satu spesies dari famili Braconidae dan Eucoilidae yaitu Opius chromatomyiae dan Gronotoma micromorpha. Parasitoid O. chromatomyiae merupakan parasitoid yang dominan pada skala ketinggian 1001-1300 m dan 1301-1600 m dari permukaan laut dengan tingkat parasitisasi berkisar antara1.84 – 62.26%. Dari 16 spesies tanaman yang diinfestasi oleh lalat pengorok daun, O. chromatomyiae dapat memarasit lalat pengorok daun pada 13 tanaman.
Kata kunci: lalat pengorok daun, Liriomyza spp., parasitoid, sayuran
Abstract
asl in which the parasitism reached to approximatelly 1.84–62.26%. Among 16 crop spesies, O. chromatomyiae could be found on 13 host plants.
Keywords: Leafminers, Liriomyza spp., parasitoid, vegetables
Pendahuluan
Hama lalat pengorok daun, Liriomyza spp. merupakan hama utama pada pertanaman sayuran. Sejak ditemukan pertama kali tahun 1994 di daerah Cisarua, Bogor (Rauf 1995), hingga saat ini, serangan hama lalat pengorok daun masih menjadi kendala utama bagi petani, khususnya petani sayuran. Hama ini bersifat polifag, serangannya pada tanaman kentang dapat menurunkan hasil hingga 70% (Rauf & Shepard 1999).
Lalat Liriomyza diketahui berasal dari Amerika Selatan (Rauf 1995). Genus Liriomyza terdiri dari 300 spesies dan diketahui ada tiga spesies yang bersifat polifag yang menimbulkan kerugian di berbagai negara (Spencer & Steyskal 1986). Ketiga spesies tersebut adalah L. trifolii (Burgess), L. sativae Blanchard dan L. huidobrensis (Blanchard). Menurut Rauf (1999), L. huidobrensis merupakan spesies yang pertama kali masuk ke Indonesia.
Aktifitas peneluran dan makan imago serta aktifitas makan larva pengorok daun dapat menyebabkan rusaknya sel-sel jaringan daun tanaman dengan gejala berupa bintik-bintik putih serta liang korokan larva yang berkelok-kelok dan bewarna keperakan. Akibatnya luas bagian daun yang berfotosintesis berkurang sehingga menurunkan kualitas dan kuantitas hasil (Parrella 1987; Cardova & Karel 1990), menurunkan nilai estetika, tanaman lebih mudah tertular penyakit, dan daun lebih cepat gugur (Supartha 1998).
chromatomyiae merupakan endoparasitoid yang dilaporkan dominan menyerang lalat pengorok daun (Rustam 2002). Hasil survei yang dilakukan ini, belum mengungkapkan tingkat parasitisasi O. chromatomyiae dan penyebaran inangnya pada berbagai ketinggian tempat.
Pemanfaatan parasitoid ini belum banyak dilakukan karena kajian tentang O. chromatomyaie ini masih sangat terbatas. Hubungan antara parasitoid dengan inangnya, serta faktor-faktor lingkungan yang mempengaruhi interaksinya di lapangan, misalnya ketinggian lokasi dan tanaman inang merupakan aspek-aspek yang perlu diketahui untuk merumuskan strategi pengendalian hayati yang tepat.
Penelitian bertujuan untuk mengidentifikasi spesies lalat Liriomyza spp. pada pertanaman sayuran dataran tinggi serta parasitoid yang berasosiasi dengan lalat pengorok daun. Selain itu, penelitian juga bertujuan untuk mengetahui tingkat parasitisasi O. chromatomyiae pada berbagai ketinggian dan jenis tanaman inang.
Bahan dan Metode
Tempat dan Waktu Penelitian
Penelitian dilaksanakan di pertanaman sayuran pada berbagai ketinggian tempat di Kabupaten Cianjur dan Bogor (Tabel 1.1). Pemeliharaan dan identifikasi lalat pengorok daun dan parasitoidnya dilaksanakan di Laboratorium Lapangan Agropolitan, Cianjur dan Laboratorium Ekologi Serangga, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian berlangsung dari bulan Juni sampai Desember 2006.
Pengambilan Daun Contoh
Geografis Position System (GPS) dan ketinggian dikelompokkan atas tiga skala yaitu 700-1000 m, 1001-1300, dan 1301-1600 m dpl.
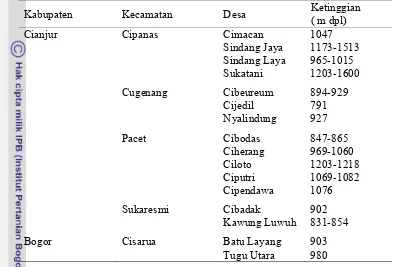
Tabel 1.1 Lokasi pengambilan sampel daun
Kabupaten Kecamatan Desa Ketinggian
( m dpl) Cianjur Bogor Cipanas Cugenang Pacet Sukaresmi Cisarua Cimacan Sindang Jaya Sindang Laya Sukatani Cibeureum Cijedil Nyalindung Cibodas Ciherang Ciloto Ciputri Cipendawa Cibadak Kawung Luwuh Batu Layang Tugu Utara 1047 1173-1513 965-1015 1203-1600 894-929 791 927 847-865 969-1060 1203-1218 1069-1082 1076 902 831-854 903 980
Pengamatan di Laboratorium
Daun contoh yang dikoleksi dibersihkan dari serangga- serangga lain dan kotoran yang menempel pada daun dengan menggunakan kuas, kemudian dimasukkan ke dalam wadah plastik berbentuk silinder dengan diameter 25 cm dan tinggi 15 cm. Pada bagian tengah wadah dipasang sekat kawat kasa sebagai penyangga daun, dan pada bagian tutup wadah dibuat ventilasi udara berbentuk bujur sangkar dengan ukuran 15 cm x 15 cm. Jenis dan jumlah imago pengorok daun dan parasitoid yang muncul diamati dan dihitung setiap hari sampai seluruh imago keluar.
Imago Liriomyza dan parasitoid yang muncul dimasukkan ke dalam botol film yang berisi alkohol 70% untuk dikoleksi serta diidentifikasi. Tingkat parasitisasi parasitoid dihitung dengan rumus:
Jumlah imago parasitoid
Tingkat parasitisasi = x 100%
Identifikasi
Imago Liriomyza spp. diidentifikasi berdasarkan ciri morfologi dengan menggunakan kunci determinasi Spencer (1973). Parasitoid diidentifikasi dengan menggunakan Program Lucid Key: Liriomyza Parasitoid of South East Asia (Fisher et al. 2006).
Hasil dan Pembahasan Hasil
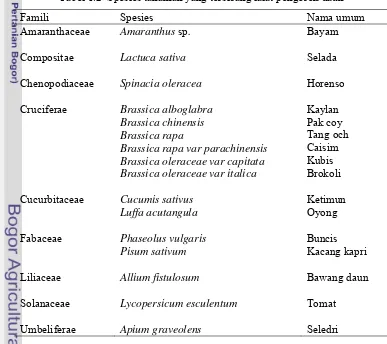
Lalat Pengorok Daun dan Tanaman Inangnya. Berdasarkan pengumpulan daun contoh yang menunjukkan gejala korokan, didapatkan 16 jenis tanaman dari 23 jenis tanaman. Tanaman tersebut terdiri atas 9 famili tanaman yakni Amaranthaceae, Compositae, Chenopodiaceae, Cruciferae, Cucurbitaceae, Fabaceae, Liliaceae, Solanaceae, dan Umbeliferae (Tabel 1.2).
Tabel 1.2 Spesies tanaman yang terserang lalat pengorok daun
Famili Spesies Nama umum
Amaranthaceae
Compositae
Chenopodiaceae
Cruciferae
Cucurbitaceae
Fabaceae
Liliaceae
Solanaceae
Umbeliferae
Amaranthus sp.
Lactuca sativa
Spinacia oleracea
Brassica alboglabra Brassica chinensis Brassica rapa
Brassica rapa var parachinensis Brassica oleraceae var capitata Brassica oleraceae var italica
Cucumis sativus Luffa acutangula
Phaseolus vulgaris Pisum sativum
Allium fistulosum
Lycopersicum esculentum
Apium graveolens
Bayam
Selada
Horenso
Kaylan Pak coy Tang och Caisim Kubis Brokoli
Ketimun Oyong
Buncis Kacang kapri
Bawang daun
Tomat
Spesies tanaman yang tidak terserang lalat pengorok daun adalah cabai (Capsicum annum), bit (Beta vulgaris), kucai (Allium sp.), lobak (Raphanus sativus), selada indian (Lactuca indica), terong (Solanum melongena), dan wortel (Daucus carota). Di lapangan ditemukan juga tanaman liar yang terserang lalat pengorok daun, yaitu babadotan (Ageratum conyzoides).
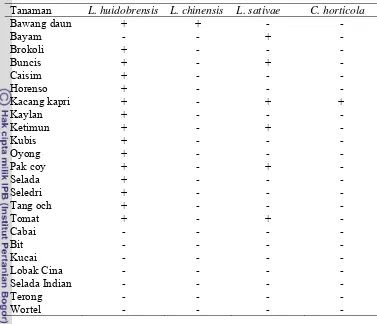
Spesies pengorok daun yang muncul dari daun contoh dengan gejala korokan bervariasi tergantung pada jenis tanaman inang. Ditemukan empat spesies lalat pengorok daun Famili Agromyzidae yang menyerang tanaman sayuran dataran tinggi, yaitu L. huidobrensis, L. chinensis, L. sativae, dan Chromatomyiae horticola (Lampiran 1.1). Dari empat spesies lalat pengorok daun tersebut, dua spesies bersifat polifag yaitu L. huidobrensis dan L. sativae. Kedua spesies ini dapat menyerang berbagai spesies tanaman dari famili yang berbeda, sedangkan dua spesies lainnya yaitu L. chinensis ditemukan hanya menyerang bawang daun dan C. horticola menyerang kacang kapri (Gambar 1.1, 1.2, Tabel 1.3)
L. huidobrensis merupakan spesies yang dominan ditemukan di dataran tinggi. Pada tanaman bawang daun populasi serangga ini semakin berlimpah dengan meningkatnya ketinggian tempat. Hampir semua lalat pengorok daun yang muncul dari daun terserang adalah spesies L. huidobrensis. Tanaman bawang daun hanya diinfestasi oleh dua spesies lalat pengorok daun yaitu L. huidobrensis dan L. chinensis. Beberapa tanaman seperti seledri dan bawang daun hanya diinfestasi oleh L. huidobrensis walaupun L. sativae mampu hidup pada ketinggian tanaman tersebut tumbuh.
Jenis Parasitoid, Kelimpahan dan Tingkat Parasitisasi. Dari survei pada berbagai jenis tanaman sayuran yang terserang lalat pengorok daun, diperoleh delapan spesies parasitoid yang berasosiasi dengan lalat pengorok daun (Tabel 1.4). Delapan spesies parasitoid tersebut terdiri dari masing-masing satu spesies dari famili Braconidae dan Eucoilidae dan enam spesies dari famili Eulophidae (Tabel 1.4, Lampiran 1.2)
Gambar 1.1 Kelimpahan populasi lalat pengorok daun pada tanaman bawang daun, brokoli, seledri, ketimun dan kacang kapri pada berbagai ketinggian tempat. 0 100 200 300 400 500 600 700
791 901 903 927 932 965 1011 1060 1082 1200 1205 1256 1333 1359 1430 1489 1501 1570 1592
Ketinggian tempat (m dpl)
P o pul as i l a la t pe ng or o k da un (i nd iv id u/ 40 da un )
L. huidob rensis L. chinensis
Bawang daun 0 200 400 600 800 1000
1203 1218 1447 1496 1506 1582 Ketinggian tempat (m dpl)
P op ul as i l al at p eng or ok da un ( in di v id u/20 dau
n) L. huidobrensis
Brokoli 0 50 100 150 200
865 969 1003 1007 1007 1184 1203
Ketinggian tempat (m dpl)
P opulas i lalat pengo rok daun (i ndiv idu/ 20 da
un) L. huidobrensis
Seledri 0 200 400 600 800 1000
847 854 932 1010 1076 1359
Ketinggian tempat (m dpl)
P o pul a s i l al a t pe ngo rok da un (i nd iv id u/ 20 da un)
L. huidobrensis L. sativae
Ketimun 0 100 200 300 400 500 600 700
838 839 847 8561011 1173 1174 1205
Ketinggian tempat (m dpl)
P op ul as i l ala t pe ng or ok da un (i n di v id u/ 4 0 d aun )
L. huidobrensis C. horticola
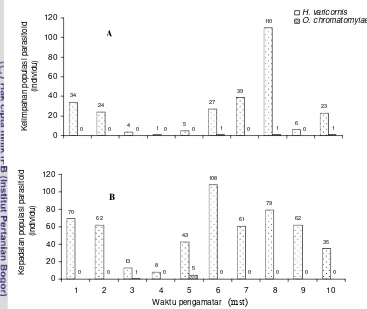
Kelimpahan parasitoid pada 3 kisaran daerah ketinggian memperlihatkan bahwa H. varicornis merupakan spesies parasitoid yang paling dominan di berbagai daerah ketinggian. O. chromatomyiae menempati urutan kedua terbanyak ditemukan. Semakin tinggi tempat maka proporsi O. chromatomyiae semakin meningkat. Sementara itu, spesies parasitoid lain hanya sedikit ditemukan, yakni pada kisaran proporsi 0.00- 3.23% pada berbagai daerah ketinggian (Gambar 1.3).
Gambar 1.2 Kelimpahan populasi lalat pengorok daun pada tanaman kaylan, tomat, horenso, buncis, tang och, bayam, dan caisim di berbagai ketinggian tempat 0 100 200 300 400
1332 1339 1389 1509 1584 1600 Ketinggian tempat (m dpl)
P opu las i l a lat p eng or o k daun (i ndi v idu/ 20 d aun )
L. huidobrensis
Kaylan 0 100 200 300 400 500
969 980 1174 1184 1399
Ketinggian tempat (m dpl)
P o pul a s i l a lat pen go ro k da un (i nd iv idu /2 0 da un ))
L. huidobrensis L. sativae
Tomat 0 100 200 300 400
1332 1339 1389 1509 1584 1600 Ketinggian tempat (m dpl)
P opu las i l a lat p eng or o k daun (i ndi v idu/ 20 d aun
) L. huidobrensis
Horenso 0 50 100 150 903 1007
Ketinggian tempat (m dpl)
P opul as i l a lat pengo rok daun ( in di v id u/ 2 0 d
aun) L. huidobrensis L. sativae
Pak coy 0 20 40 60 80 100 120 140 903 1007
Ketinggian tempat (m dpl)
P o pu la s i l a la t p e n g o rok dau n (i nd iv id u/2 0 da un )
L. huidobrensis L. sativae
Buncis 0 20 40 60 80 100 894 1076 Ketinggian tempat (m dpl)
P o pul as i l al at pen go ro k d au n ( ind iv idu /20 da un
) L. huidobrensis L. sativae
Tabel 1.3 Spesies lalat pengorok daun pada berbagai jenis tanaman inangnya
Tanaman L. huidobrensis L. chinensis L. sativae C. horticola
Bawang daun + + - -
Bayam - - + -
Brokoli + - - -
Buncis + - + -
Caisim + - - -
Horenso + - - -
Kacang kapri + - + +
Kaylan + - - -
Ketimun + - + -
Kubis + - - -
Oyong + - - -
Pak coy + - + -
Selada + - - -
Seledri + - - -
Tang och + - - -
Tomat + - + -
Cabai - - - -
Bit - - - -
Kucai - - - -
Lobak Cina - - - -
Selada Indian - - - -
Terong - - - -
Wortel - - - -
Tabel 1.4 Parasitoid yang menyerang lalat pengorok daun
Famili Spesies Braconidae Opius chromatomyiae
Eucoilidae Gronotoma micromorpha
Eulophidae Asecodes deluchii
Hemiptarsenus varicornis Neochrysocharis okazakii Neochrysocharis formosa Neocrhysocharis sp. Quadrastichus liriomyzae
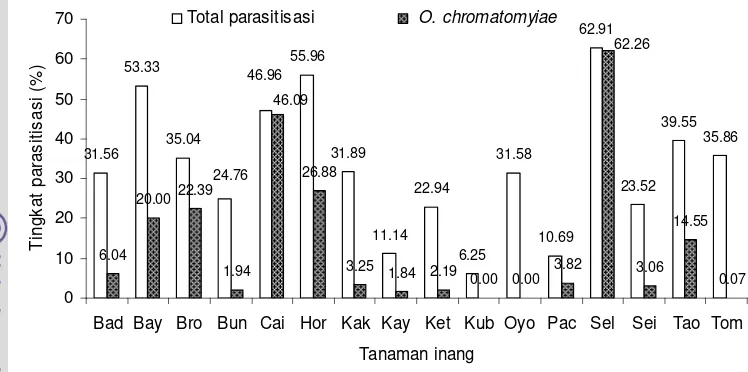
chromatomyiae mendominasi tingkat parasitisasi seperti terlihat pada tanaman brokoli, caisim, horenso, dan selada.
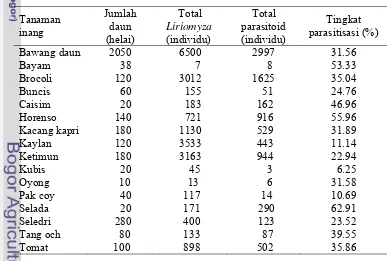
Tabel 1.5 Kelimpahan lalat pengorok daun dan tingkat parasitisasi parasitoid pada berbagai jenis tanaman inang
Tanaman inang Jumlah daun (helai) Total Liriomyza (individu) Total parasitoid (individu) Tingkat parasitisasi (%)
Bawang daun 2050 6500 2997 31.56
Bayam 38 7 8 53.33
Brocoli 120 3012 1625 35.04
Buncis 60 155 51 24.76
Caisim 20 183 162 46.96
Horenso 140 721 916 55.96
Kacang kapri 180 1130 529 31.89
Kaylan 120 3533 443 11.14
Ketimun 180 3163 944 22.94
Kubis 20 45 3 6.25
Oyong 10 13 6 31.58
Pak coy 40 117 14 10.69
Selada 20 171 290 62.91
Seledri 280 400 123 23.52
Tang och 80 133 87 39.55
Tomat 100 898 502 35.86
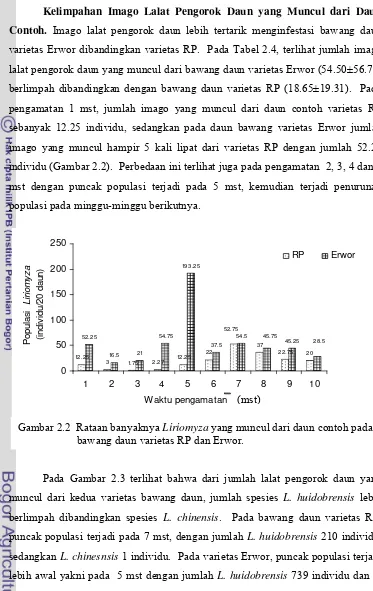
[image:30.612.119.506.405.666.2]Kelimpahan Lalat Pengorok Daun dan Parasitoidnya pada Bawang Daun. Pada pengamatan di lapangan, bawang daun merupakan tanaman yang Gambar 1.3 Komposisi parasitoid menurut ketinggian tempat.
30.90
38.97
0.54 0.00 0.00
87.52
68.49
60.97
0.99 2.96 0.00 0.08 0.00 0.00
1.71 1.26 1.80 3.23 0.41 0.12 0.00 0.04 0.02 0.00
0 10 20 30 40 50 60 70 80 90 100
700-1000 1001-1300 1301-1600
Ketinggian tempat (m dpl)
P ropor s i par a s it oi d (% )
O. chromatomyiae G. micromorpha Asecodes deluchii
H. varicornis N. okazakii N. formosa
Neochrysocharis sp. Q.liriomyzae
dominan ditanam pada pertanaman sayuran dataran tinggi di Kabupaten Cianjur dan Bogor. Tanaman ini mendapat serangan lalat Liriomyza dengan tingkat serangan tinggi.
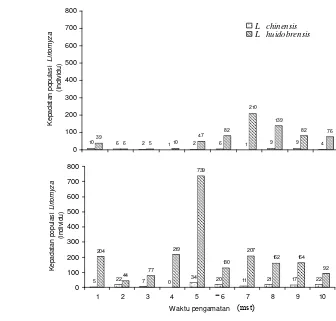
Pengamatan jumlah spesies lalat pengorok daun dan parasitoidnya pada tanaman bawang daun memperlihatkan bahwa pada satu helai daun bawang dapat diinfestasi oleh dua spesies Liriomyza (Tabel 1.6). Kedua spesies yang menyerang tanaman bawang daun tersebut adalah L. huidobrensis dan L. chinensis dengan rata-rata jumlah imago yang muncul dari daun korokan berturut-turut 19.02 dan 1.12 individu per daun. Parasitoid yang muncul dari korokan daun adalah H. varicornis dan O. chromatomyiae dengan tingkat parasitisasi masing-masing 8.81 % dan 3.11% pada kedua spesies Liriomyza.
Pembahasan
[image:31.612.136.510.86.272.2]Hasil penelitian ini menunjukkan bahwa ditemukan empat spesies lalat pengorok daun yang menyerang tanaman sayuran di dataran tinggi Kabupaten Cianjur dan Bogor. L. huidobrensis tercatat sebagai spesies lalat pengorok daun yang paling banyak menyerang berbagai jenis tanaman sayuran. Spesies L. huidobrensis ini bersifat polifag (Parrella 1987; Murphy & LaSalle 1999).
Gambar 1.4. Tingkat parasitisasi Opius chromatomyiae:
Bawang daun (Bad), Bayam (Bay), Brokoli (Bro), Buncis (Bun), Caisim (Cai), Horenso (Hor), Kacang kapri (Kak), Kaylan (Kay), Ketimun (Ket), Kubis (Kub), Oyong (Oyo), Pak coy (Pac), Selada (Sel), Seledri (Sei), Tang och (Tao) dan Tomat (Tom).
31.56 53.33 35.04 55.96 31.89 11.14 22.94 6.25 31.58 10.69 62.91 23.52 39.55 35.86 6.04 20.0022.39 1.94 26.88
3.25 1.84 2.19 0.00 0.00 3.82 3.06 14.55 0.07 24.76 46.96 62.26 46.09 0 10 20 30 40 50 60 70
Bad Bay Bro Bun Cai Hor Kak Kay Ket Kub Oyo Pac Sel Sei Tao Tom
Tanaman inang T in g ka t p a ra s it isa si ( % )
Tabel 1.6 Kelimpahan spesies Liriomyza dan parasitoid per daun pada tanaman bawang daun
Parameter Rerata jumlah individu per daun dan kisaran
Pupariun Liriomyza Jenis Liriomyza
Liriomyza huidobrensis Liriomyza chinensis Jenis parasitoid
Hemiptarsenus varicornis Opius chromatomyiae
Persentase parasitisasi parasitoid Hemiptarsenus varicornis Opius chromatomyiae
26.13 (7-48)
19.02 (4-39) 1.12 (0-7)
1.60 (0-7) 0.67 (0-4)
8.81% (0-50%) 3.11% (0-13.64%)
Rauf et al. (2000) melaporkan bahwa L. huidobrensis dapat menyerang 45 spesies tanaman yang tergolong ke dalam 20 famili tanaman. Banyaknya tanaman inang memungkinkan L. huidobrensis dengan cepat memencar dari satu wilayah ke wilayah lain (Setiawati 1998).
L. huidobrensis merupakan spesies yang pertama kali ditemukan di Indonesia pada dataran tinggi (Rauf 1999). Dari hasil penelitian ini, spesies L. huidobrensis sangat dominan di daerah tempat pengamatan. Semakin tinggi lokasi, proporsi populasi L. hudobrensis semakin meningkat. Hampir semua tanaman sayuran dataran tinggi dapat diinfestasi oleh L. huidobrensis. Kelimpahan L. huidobrensis pada dataran tinggi erat kaitannya dengan asal lalat pengorok daun ini. Lalat pengorok daun ini di daerah asalnya (Amerika Selatan) yang menyerang tanaman di daerah dataran tinggi juga.
Lalat pengorok spesies L. sativae lebih banyak pada daerah ketinggian yang lebih rendah (kisaran 700-1000 m dpl). L. sativae dapat menyerang berbagai jenis tanaman, namun kisaran tanaman inangnya lebih sempit dibandingkan L. huidobrensis. Hama ini berasal dari daerah dataran rendah dan di Indonesia juga banyak menyerang sayuran di dataran rendah (Rauf 1999, Susilawati 2002; Herlinda et al. 2006). Kerusakan pada tanaman ketimun di dataran rendah dapat mencapai 60% (Rauf & Shepard 2001).
di dataran rendah (Tapahillah 2002). Hama ini pernah mengalami ledakan populasi (out break) pada tahun 2000 dengan kehilangan hasil yang cukup tinggi (Rauf & Shepard 2001). Dalam penelitian ini pertama kali dilaporkan L. chinensis ditemukan pada ketinggian 1500 m dpl. Nampaknya L. chinensis mengalami adaptasi terhadap tanaman inang yang tumbuh di dataran yang lebih tinggi.
Kemampuan L. chinensis beradaptasi dengan dataran tinggi akan membuat kerusakan tanaman bawang daun semakin parah. Dari survey yang dilakukan, lalat pengorok daun ini ditemukan menyerang seluruh sampel bawang daun. Pengamatan intensif terhadap tanaman bawang daunmenemukan bahwa, ternyata satu helai daun dapat diserang bersama-sama oleh L. chinensis dan L. huidobrensis, namun populasinya masih didominasi oleh spesies L. huidobrensis. Spesies keempat yang menyerang tanaman sayuran dataran tinggi adalah C. horticola yang hanya menyerang tanaman kacang kapri. C. horticola telah sejak lama ada di Indonesia dan dikenal sebagai hama pengorok daun kapri (Kalshoven 1981). Sejak masuknya L. huidobrensis ke Indonesia, kedudukan C. horticola sebagai hama penting pada tanaman kapri tampaknya telah tergeser. Hampir semua imago pengorok yang keluar dari daun kacang kapri adalah L. huidobrensis.
Parasitoid yang berasosiasi dengan lalat pengorok daun yang ditemukan dalam penelitian ini meliputi delapan spesies dari tiga famili. Parasitoid-parasitoid tersebut adalah O. chromatomyiae (famili Braconidae), G. micromorpha (famili Eucoilidae) serta H. varicornis, A. deluchii, N. okazakii, N. formosa, Neochrysocharis sp., dan Q. liriomyzae (famili Eulophidae). Survei yang dilakukan oleh Rauf dan Shepard (1999) menemukan delapan spesies parasitoid yang berasosiasi dengan lalat pengorok daun yang terdiri atas famili Eulophidae dan Eucoilidae. Susilawati (2002) menemukan 18 spesies parasitoid yang menyerang lalat pengorok daun di daerah Bogor dan Banjar yang terdiri atas famili Eulophidae, Braconidae dan Eucoilidae. Famili Braconidae yang ditemukan tersebut berasal dari daerah Bogor.
H. varicornis, Zagrammosoma sp., dan satu spesies dari famili Pteromalidae (Supartha 1998). Dalam penelitian yang dilakukan ini sudah ditemukan delapan spesies parasitoid. Keberadaan parasitoid lalat pengorok daun yang beragam di lapangan saat ini, diduga berasal dari parasitoid yang berasosiasi dengan lalat pengorok daun C. horticola. Lalat pengorok daun ini merupakan hama utama pada tanaman kacang kapri (Kalshoven 1981) yang keberadaanya sudah lama di Indonesia sebelum kedatangan spesies Liriomyza spp.. Menurut Spencer (1973), Hemiptarsenus sp., Opius sp., dan Chrysocharis sp., merupakan tiga spesies diantara parasitoid yang berasosiasi dengan C. horticola pada tanaman kacang kapri.
H. varicornis dan O. chromatomyiae merupakan dua spesies parasitoid yang populasinya berlimpah ditemukan di lapangan. Hasil penelitian ini sesuai dengan hasil penelitian Hidrayani (2003) dan Purnomo (2003) yang melaporkan bahwa H. varicornis dan O. chromatomyiae merupakan parasitoid yang dominan di lapangan. Dalam penelitian ini, H. varicornis menempati proporsi paling tinggi (87.52%) pada skala ketinggian 700-1000 m dpl, sedangkan dengan peningkatan ketinggian tempat proporsi H. varicornis menurun. Sementara itu, terjadi peningkatan pada proporsi O. chromatomyiae dari 1.71% pada skala 700-1000 m dpl menjadi 38.97% pada 1301-1600 m dpl. Tampaknya O. chromatomyiae lebih dapat beradaptasi pada daerah yang lebih tinggi yakni pada daerah suhu yang lebih rendah dan kelembaban yang tinggi. Proporsi parasitoid lainnya relatif rendah dan hanya mencapai kisaran maksimal 3.23%.
Tingkat parasitisasi O. chromatomyiae lebih tinggi pada tanaman selada, caisim, dan horenso. Diduga morfologi daun yang lebar dan bentuk susunan daun yang saling tumpang tindih ketiga jenis tanaman tersebut memberikan kondisi iklim mikro yang sesuai bagi parasitoid O. chromatomyiae untuk meletakkan telurnya pada larva lalat pengorok daun. Tanaman yang berdaun lebar yang sangat disukai oleh parasitoid, khususnya O. chromatomyiae, dapat digunakan dalam manipulasi habitat untuk mendukung keberadaan parasitoid di lapangan, sehingga diharapkan dapat efektif menekan populasi lalat pengorok daun.
Kesimpulan
Tanaman sayuran dataran tinggi Kabupaten Cianjur dan Bogor telah diinfestasi oleh empat spesies lalat pengorok daun, yakni L. huidobrensis, L. sativae, L. chinensis, dan C. horticola. Dua spesies pertama, L. huidobrneisis dan L. sativae bersifat polifag, sedangkan L. chinensis dan C. horticola ditemukan hanya menyerang masing-masing pada tanaman bawang daun dan kacang kapri. Semakin tinggi ketinggian tempat jumlah L. huidobrensis semakin berlimpah sedangkan populasi L. chinensis semakin menurun.
Parasitoid yang ditemukan berasosiasi dengan lalat pengorok daun adalah O. chromatomyiae (Braconidae), G. micromorpha (Eucoilidae), H. varicornis, A. deluchii, N. okazakii, N. formosa, Neochrysocharis sp., dan Q. liriomyzae (Eulophidae). Parasitoid O. chromatomyiae ditemukan memarasit lalat pengorok daun yang menginfestasi hampir semua spesies tanaman yang dikoleksi dengan tingkat parasitisasi mencapai 62.26% dan dapat beradaptasi dengan baik pada daerah dataran tinggi.
Tanaman gulma berbunga dan tanaman yang sangat disukai oleh parasitoid, khususnya O. chromatomyiae, dapat digunakan untuk mengkonservasi parasitoid di lapangan.
Daftar Pustaka
Cardova C, Karel AK. 1990. Insect and other invertebrate pest of beans. In S.R. Singh. Insect Pest of Food Legumes. Jhon Willey & Sons Ltd. 157-191. Fisher N, Ubaidillah R, Reina P, La Salle J. 2006. Liriomyza Parasitoids of South
Herlinda S, Purwanti E, Pujiastuti Y, Rauf A. 2006. Preferensi Liriomyza sativae (Balnchard) (Diptera: Agromyzidae) pada ketimun dan tomat. Agritop 25(1): 7-11.
Hidrayani. 2003. Bioekologi Hemiptarsenus varicornis (Girault) (Hymenoptera: Eulophidae), parasitoid Liriomyza huidobrensis (Blanchard) (Dipetara: Agromyzidae) [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Revised by PA. van der Laan. PT. Ichtiar Baru. Van Hoeve. Jakarta. 701 p.
Murphy ST, LaSalle J. 1999. Balancing biological control strategies in the IPM of new world invasive Liriomyza leafminers in field vegetable crops. Biocontrol News and Information 20: 91-104.
Parrella MP. 1987. Biology of Liriomyza. Annu Rev Entomol 32:201-224.
Purnomo. 2003. Liriomyza huidobrensis (Blanchard) (Diptera: Agromyzidae): Kesesuaian inang, perkembangan populasi, dan pengaruh insektisida translamina [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor. Rauf A. 1995. Liriomyza: hama pendatang baru di Indonesia. Bul HPT 8(1):
46– 48.
Rauf A. 1999. Persepsi dan tindakan petani kentang terhadap lalat pengorok daun, Liriomyza huidobrensis (Blanchard) (Diptera: Agromyzidae). Buletin HPT 11(1):1-13.
Rauf A, Shepard BM. 1999. Leafminers in vegetables in Indonesia: surveys of host crops. Species composition, parasitoids and control practices. Worhshop of Leafminers of Vegetables in Southeast Asia; Selangor, 2-5 February 1999. Selangor: CAB International, Southeast Asia Regional Center.
Rauf A, Shepard BM. 2001. Current status on the biology, ecology and management of Liriomyza spp. in Indonesia with emphasis on L. huidobrensis. Paper presented at seminar on invasive arthropod pests of vegetables and economic food crops, Kualalumpur Malaysia, 13-14 March 2001.
Rauf A, Shepard BM, Johnson MW. 2000. Leafminers in vegetables, ornamental plants and weeds in Indonesia: surveys of host crops, species composition and parasitoids. Intern J Pest Manage 46(4):257-266.
Rustam R. 2002. Biologi Opius sp.(Hymenoptera: Braconidae) parasitoid lalat pengorok daun kentang [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Setiawati W. 1998. Liriomyza huidobrensis hama baru pada tanaman kentang. Bandung: Balai Tanaman Sayuran
Spencer KA. 1973. Agromyzidae (Diptera) of Economic Importance. Junk BVW, editor. The Hague, 418 p.
Supartha IW. 1998. Bionomi lalat pengorok daun, Liriomyza huidobrensis (Blanchard) (Diptera: Agromyzidae), pada pertanaman kentang [disertasi]. Bogor; Program Pascasarjana , Institut Pertanian Bogor.
Susilawati. 2002. Komposisi dan kelimpahan parasitoid lalat pengorok daun Liriomyza sativae Blanchard (Diptera: Agromyzidae) [thesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Lampiran 1.1 Spesies lalat pengorok daun yang menyerang pertanaman sayuran pada dataran tinggi (skala pembanding adalah 1 mm)
Liriomyza huidobrensis Liriomyza sativae
Lampiran 1.2 Spesies parasitoid lalat pengorok daun dataran tinggi (skala pembanding adalah 1 mm)
Opius chromatomyiae Hemiptarsenus varicornis
Asecodes deluchii Neochrysocharis formosa
Neochrysocharis okazakii Neochrysocharis sp.
2.
DINAMIKA POPULASI
Liriomyza
spp. (DIPTERA:
AGROMYZIDAE) DAN PARASITOIDNYA PADA DUA
VARIETAS TANAMAN BAWANG DAUN
(
Allium fistulosum
L.)
Abstrak
Tanaman inang sebagai sumber makanan serangga merupakan salah satu faktor yang mempengaruhi perkembangan populasi serangga di lapangan. Tujuan penelitian adalah untuk mempelajari perkembangan populasi Liriomyza spp. dan parasitoidnya pada dua varietas tanaman bawang daun di lapangan. Percobaan dinamika populasi lalat pengorok daun dan parasitoidnya dilakukan dengan mengambil 20 helai daun terserang lalat pengorok daun dan pemasangan perangkap kuning setiap minggu. Dari analisis pengamatan data berulang terhadap data pengamatan mingguan, ternyata perbedaan varietas tanaman bawang daun tidak mempengaruhi jumlah Liriomyza yang tertangkap perangkap kuning namun mempengaruhi jumlah Liriomyza dan parasitoid yang muncul dari daun contoh. Terdapat dua spesies pengorok daun yang menyerang pertanaman bawang daun, yaitu L. huidobrensis dan L. chinensis dengan rerata jumlah Liriomyza yang muncul dari bawang daun varietas Erwor dan RP berturut-turut sebanyak 54.50 individu dan 18.65 individu. Jumlah parasitoid yang muncul dari bawang daun varietas Erwor (13.68 individu) berlimpah dibandingkan varietas RP (6.90 individu). Spesies parasitoid yang muncul dari daun contoh kedua varietas adalah Hemiptarsenus. varicornis dan Opius chromatomyiae dengan tingkat parasitisasi kedua spesies hampir sama pada kedua varietas bawang daun. Tingkat kerusakan tanaman lebih berat terjadi pada bawang daun varietas Erwor
Kata kunci: populasi, lalat pengorok daun, parasitoid, bawang daun
Abstract
In general,host plant plays an important role in affecting the development of insect populating. The study aimed to investigate the population growth of leafminers and its parasitoids on two varieties of shallot in the field. To obtain the aim, 20 damage leaves were collected and 5 yellow traps were placed. The observation was conducted every week. The result showed that shallot varieties did not affect the number of leafminers trapped on yellow sticky trap, but they significantly influenced the number of leafminers and its parasitoids. Two species were found, L. huidobrensis and L. Chinensis. In Erwor and RP variety, the average of individual reached 54.50 and 18.65 respectivelly, in consequences, crop damage was also significantly higher in Erwor rather than in RP variety. In term of parasitoids, only two species were found on both Erwor and RP varieties. They were Hemiptarsenus varicornis and Opius chromatomyiae. Host plant influenced the number of of parasitoid population in which the population was higher in Erwor than RP variety reaching 13.68 and 6.90 individual per 20 damage respectively. However, host plant did not affect to parasitim rate.
Pendahuluan
Bawang daun merupakan tanaman utama pada pertamanan sayuran dataran tinggi di Kabupaten Cianjur. Namun dalam upaya peningkatan produksinya petani menghadapi kendala karena serangan hama pengorok daun Liriomyza spp. Hal ini merupakan masalah yang sangat penting pada tanaman bawang daun karena yang diserang adalah bagian tanaman yang langsung dipasarkan sehingga menurunkan harga jual.
Sampai saat ini telah diketahui tiga spesies Liriomyza yang menginfestasi pertanaman sayuran di Indonesia. Liriomyza huidobrensis (Blanchard) spesies yang pertama kali ditemukan pada sekitar tahun 1994 di Cisarua (Rauf 1995), dua tahun kemudian diikuti oleh L. sativae Blanchard (Rauf et al. 2000). Jenis yang ketiga adalah L. chinensis Koto yang tiba di Indonesia pada tahun 2000 (Rauf & Shepard 2001). Spesies L. huidobrensis dan L sativae bersifat polifag dan menyerang sekitar 45 spesies tanaman (Rauf et al. 2000), sedangkan L. chinensis hanya menyerang pada jenis tanaman bawang.
Kerusakan pada tanaman yang terserang oleh ketiga spesies pengorok ini adalah karena tusukan ovopisitor imago pada daun yang menurunkan nilai produksi daun dan korokan larva pada jaringan daun yang menyebabkan menurunya kemampuan fotosintesis tanaman. Serangan berat menyebabkan daun mengering dan gugur sebelum waktunya. Menurut Rauf dan Shepard (1999), serangan hama pengorok ini dapat menurunkan hasil panen sampai 70%. Pada keadaan populasi tinggi, serangan dapat menyebabkan kegagalan panen atau kematian pada tanaman (Spencer 1973).
dengan insektisida kurang efektif. Dalam PHT, informasi tentang dinamika populasi hama yang akan dikendalikan sangat diperlukan.
Tabel 2.1 Deskripsi tanaman bawang daun
Kategori Varietas bawang daun
RP Erwor Panjang batang semu (cm) 18-25 15-17
Panjang daun (cm) 30-35 40-55 Diameter batang (cm) 1.7-2.25 1.5-2.0 Warna pangkal batang Putih orange
Batang semu kokoh agak kokoh
Warna daun Hijau tua hijau muda
Ketebalan daun Tebal kurang tebal
Anakan sedikit banyak
Sampai saat ini, belum pernah dilaporkan tentang kajian dinamika populasi hama lalat pengorok daun dan parasitoidnya pada tanaman bawang daun. Oleh karena itu, kajian ekologi hama pengorok daun pada tanaman bawang daun perlu dilakukan termasuk parasitoidnya yang meliputi populasi imago dan tingkat kerusakan daun oleh serangan hama pengorok daun, spesies parasitoid yang menyerang hama pengorok daun pada tanaman bawang dan menganalisis hubungan antara kelimpahan larva hama penggorok daun dan parasitoidnya.
Bahan dan Metode
kandang yang diberikan sebanyak 20 ton/ha, Urea 300 kg/ha, TSP 400 kg/ha dan KCl 200 kg/ha.
Percobaan disusun dalam bentuk Rancangan Acak Kelompok dengan dua perlakuan dan empat ulangan. Peubah yang diamati mencakup populasi imago lalat Liriomyza yang terperangkap, jumlah Liriomyza dan parasitoid yang muncul, tingkat parasitisasi parasitoid, dan tingkat kerusakan tanaman.
Pengamatan Populasi Imago Liriomyza spp.
Untuk memantau populasi imago pengorok daun digunakan perangkap kuning seperti yang dilakukan oleh Robin dan Mitcheli (1987). Perangkap kuning berukuran 15 cm x 10 cm yang terbuat dari lembaran plastik bewarna kuning. Plastik transparan yang telah dioles tipis dengan lem tikus Cap Gajah yang bewarna bening ditempelkan pada kedua sisi lembaran perangkap kuning. Perangkap kuning dipasang pada ketinggian 100 cm dari permukaan tanah selama 24 jam. Pemasangan perangkap dilakukan setiap minggu dan dilakukan pada saat tanaman berumur 1 minggu setelah tanam (mst) hingga panen (10 mst). Jumlah perangkap kuning yang dipasang pada tiap subpetak contoh adalah sebanyak 5 buah dan tersebar sesuai arah diagonal.
Pengamatan Kelimpahan Liriomyza spp. dan Parasitoid
Perhitungan Tingkat Parasitisasi
Jumlah imago parasitoid
Tingkat parasitisasi = x 100%
Jumlah imago parasitoid + imago Liriomyza
Pengamatan Tingkat Kerusakan Tanaman
Kerusakan tanaman bawang daun akibat serangan Liriomyza diamati secara langsung pada daun tanaman contoh. Pengamatan dilakukan pada 20 tanaman contoh untuk setiap subpetak percobaan yang tersebar secara diagonal. Pengamatan ini dilakukan setiap minggu, sejak tanaman berumur 1 – 10 mst.
Penentuan tingkat kerusakan dilakukan dengan menilai kerusakan tanaman menggunakan skala 0-4 berdasarkan kriteria seperti pada Tabel 2.2.
Tabel 2.2 Kriteria skor kerusakan daun akibat serangan Liriomiyza Skor Kondisi serangan/gejala
0
1
2
3
4
Tidak ada gejala korokan pada daun
Kerusakan rendah, gejala korokan hanya terbatas pada daun bagian bawah
Kerusakan sedang, gejala korokan terbatas pada daun bagian bawah dan tengah tanaman
Gejala korokan hampir pada seluruh daun tanaman kecuali daun pucuk
Gejala korokan ditemukan pada semua daun tanaman
Analisis Data
Data penelitian selama kurun waktu pengamatan yang meliputi populasi imago lalat pengorok daun, banyaknya parasitoid yang muncul, tingkat parasitisasi, dan tingkat kerusakan tanaman, dianalisis dengan sidik ragam pengamatan berulang dengan Program SAS 9.0.
Hasil dan Pembahasan Hasil
jumlah Liriomyza yang muncul dari daun contoh (F=14.38, db=1,3, P=0.0005). Pengaruh perbedaan waktu pengamatan berpengaruh nyata terhadap kedua peubah tersebut (Tabel 2.3).
Tabel 2.3 Perbandingan beberapa peubah infestasi Liriomyza pada petak pertanaman bawang daun varietas RP dan Erwor berdasarkan sidik ragam pengukuan berulang
Pengaruh Peubah F db P
Perlakuan Banyaknya Liriomyza yang tertangkap perangkap kuning
Banyaknya Liriomyza yang muncul dari daun contoh
5.13
14.38
1,30 0.2970
0.0005
Waktu Banyaknya Liriomyza yang tertangkap perangkap kuning
Banyaknya Liriomyza yang muncul dari daun contoh
26.30
3.21
9,27 <0.0001
0.0060
Perbedaan varietas bawang daun tidak mempengaruhi jumlah imago Liriomyza yang mendatangi areal pertanaman bawang daun. Rataan jumlah imago Liriomyza yang tertangkap perperangkap kuning sebanyak 57.95±36.33 pada varietas RP dan 68.13±47.59 pada varietas Erwor (Tabel 2.4). Namun pengaruh waktu menyebabkan jumlah lalat pengorok daun yang tertangkap perangkap kuning terlihat berbeda dari minggu ke minggu.
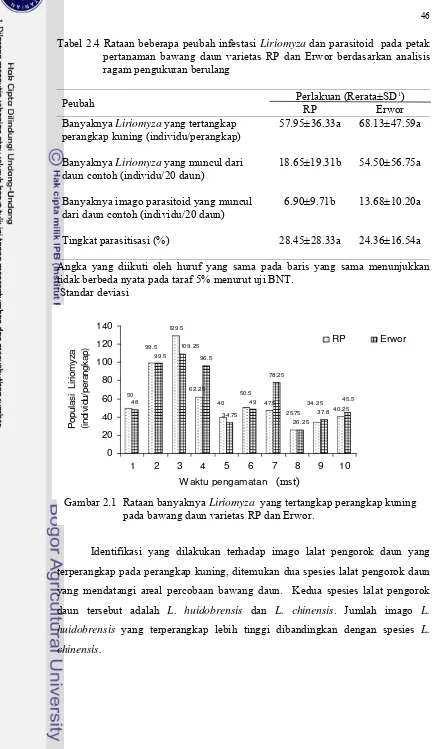
Tabel 2.4 Rataan beberapa peubah infestasi Liriomyza dan parasitoid pada petak pertanaman bawang daun varietas RP dan Erwor berdasarkan analisis ragam pengukuran berulang
Peubah Perlakuan (Rerata±SD
1)
RP Erwor Banyaknya Liriomyza yang tertangkap
perangkap kuning (individu/perangkap)
Banyaknya Liriomyza yang muncul dari daun contoh (individu/20 daun)
57.95±36.33a
18.65±19.31b
68.13±47.59a
54.50±56.75a
Banyaknya imago parasitoid yang muncul dari daun contoh (individu/20 daun)
6.90±9.71b 13.68±10.20a
Tingkat parasitisasi (%) 28.45±28.33a 24.36±16.54a
Angka yang diikuti oleh huruf yang sama pada baris yang sama menunjukkan tidak berbeda nyata pada taraf 5% menurut uji BNT.
<