LALAT PENGOROK DAUN Liriomyza spp. (DIPTERA:
AGROMYZIDAE) DAN PARASITOIDNYA PADA SAYURAN
DATARAN TINGGI DENGAN PERHATIAN UTAMA PADA
PARASITOID Opius chromatomyiae BELOKOBYLSKIJ &
WHARTON (HYMENOPTERA: BRACONIDAE)
RUSLI RUSTAM
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Lalat Pengorok Daun
Liriomyza spp. (Diptera: Agromyzidae) dan Parasitoidnya pada Sayuran Dataran Tinggi dengan Perhatian Utama pada Parasitoid Opius chromatomyiae
Belokobylskij & Wharton (Hymenoptera: Braconidae) adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2009
Rusli Rustam
ii RUSLI RUSTAM. Leafminers Liriomyza spp. (Diptera: Agromyzidae) on Highland Vegetables and Their Parasitoids with The Emphasis on Opius chromatomyiae Belokobylskij & Wharton (Hymenoptera: Braconidae). Supervised by AUNU RAUF, NINA MARYANA, PUDJIANTO and DADANG.
Leafminers Liriomyza spp. are major pest on highland vegetables in Indonesia. The objectives of the research were to (1) evaluate leafminer and its parasitoids on higland vegetables, (2) study the intensity of parasitism of O. chromatomyiae in various host plants, (3) study the population dynamic of leafminer and its parasitoid in shallot, (4) evaluate the impact of three insecticides on parasitoid O. chromatomyiae, and (5) evaluate the demographic parameters and functional response of parasitoid O. chromatomyiae. Surveys of leafminer population and its parasitoid were undertaken by collecting 20-40 infested leaves. Leafminer flies and parasitoids emerged were collected and identified. Level of parasitism was calculated based on the number of parasitoids emerged divided by the total number of flies and wasps emerged. The population dynamic of leafminer and its parasitoids were studied on two varieties of green onion, by collecting 20 leaves and counting number of flies catched on yellow traps every week. The impact of insecticides on both leafminers and parasitoids was conducted by applying three different insecticides: abamectin, azadirachtin, and cartaphydrochloride once a week at the recommended dosage. Life table of O. chromatomyiae was studied to determine the population growth potential of the parasitoid, by calculating reproduction rate, intrinsic rate of increase, generation time, and stable age distribution. Functional response was evaluated by exposing
O. chromatomyiae on the third instar larvae of L. huidobrensis at eight different host densities (2, 4, 8, 10, 12, 16, 20, and 24) of. Our surveys revealed that
Liriomyza huidobrensis, L. sativae, L. chinensis and Chromatomyia horticola
were associated with various highland vegetables. Eight parasitoids emerged from leafminer-infested leaves: Hemiptarsenus varicornis, Asecodes deluchii, Neochrysocharis okazakii, N. formosa, Neocrhysocharis sp. Quadrastitchus liriomyzae, O. chromatomyiae and Gronotoma micromorpha. Parasitoid Opius chromatomyiae emerged from 13 different species of vegetables infested by leafminers.. Number of leafminers and parasitoids emerged, and crop damage were significantly different between two green onion varieties tested. Cartap hydrochloride was the most effective insecticide in term for reducing leafminer populations. Even though not as effective as cartap hydrochloride, abamectin and azadirachtin could reduce level of leafminer damage and had lower detrimental effects on natural enemies. Life table studies showed that longevity of females
Opius chromatomyiae (9.95 days) shorter than males (11.73 days). Females laid 104.73 eggs with the oviposition rate was 9.31 eggs/days. Reproductive output (R0) was 28.55 with generation time (T) was about 15.96 days and intrinsic rate of increase was 0.21. The stable age ditribution was 37.93% eggs, 24.92% larvae, 20.36% pupae and 16.78% adults. Functional response of O. chromatomyiae is type II (hyperbolic curve) with searching time was 0.0777/hour and handling time was 2.5818/hour.
iii RUSLI RUSTAM. Lalat Pengorok Daun Liriomyza spp. (Diptera: Agromyzidae) dan Parasitoidnya pada Sayuran Dataran Tinggi dengan Perhatian Utama pada Parasitoid Opius chromatomyiae Belokobylskij & Wharton (Hymenoptera: Braconidae). Dibimbing oleh AUNU RAUF, NINA MARYANA, PUDJIANTO, dan DADANG.
Tabuhan Opius chromatomyiae adalah parasitoid yang dominan berasosiasi dengan larva pengorok daun pada pertanaman sayuran dataran tinggi di Indonesia. Penelitian bertujuan untuk (1) mengidentifikasi spesies lalat pengorok daun pada pertanaman sayuran dataran tinggi serta parasitoid yang berasosiasi, (2) mengetahui tingkat parasitisasi O. chromatomyiae pada berbagai jenis tanaman inang, (3) mengkaji dinamika populasi lalat pengorok daun dan parasitoidnya pada tanaman bawang daun, (4) mengevaluasi selektifitas beberapa insektisida terhadap parasitoid O. chromatomyiae (5) mengkaji potensi keefektifan parasitoid O. chromatomyiae berdasarkan parameter demografi yang meliputi laju pertambahan intristik (r), reproduksi bersih (Ro), masa generasi (T) dan proporsi persebaran usia stabil (px), (6) mengkaji potensi keefektifan parasitoid O. chromatomyiae berdasarkan tanggapnya terhadap peningkatan kelimpahan inang, serta menetapkan laju pelacakan inang (a) dan masa penanganan inang (Th).
Survei lalat pengorok daun dan parasitoidnya dilakukan dengan mengambil 20-40 helai daun tanaman sayuran yang memperlihatkan gejala korokan lalat pengorok daun. Imago lalat pengorok daun dan parasitoid yang muncul dari daun diidentifikasi. Tingkat parasitisasi dihitung dengan membandingkan jumlah parasitoid dengan lalat pengorok daun yang muncul. Percobaan dinamika populasi lalat pengorok daun dan parasitoidnya dilakukan pada dua varietas tanaman bawang daun. Pengamatan dilakukan setiap minggu dengan mengambil 20 helai daun terserang dan pemasangan perangkap kuning. Percobaan aplikasi insektisida pada tanaman bawang daun dan pengaruhnya terhadap parasitoid dilakukan pada tiga jenis insektisida, yaitu abamektin, azadirakhtin, dan kartap dengan dosis sesuai anjuran pada label kemasan insektisida. Aplikasi insektisida dilakukan setiap minggu. Pengamatan dilakukan dengan mengambil 15 helai daun tanaman terserang dan pemasangan perangkap kuning. Data dianalisis dengan pengamatan data berulang. Sedangkan percobaan parameter demografi O. chromatomyiae dilakukan pada 50 individu cohort awal dengan mengamati pertumbuhan dan perkembangan parasitoid mulai dari telur sampai imago mati. Parameter yang diamati adalah keperidian, laju pertumbuhan intrinsik, masa generasi, dan persebaran umur stabil. Percobaan tanggap fungsional O. chromatomyiae dilakukan di laboratorium dengan menggunakan larva lalat pengorok daun instar-3 sebagai serangga inangnya. Percobaan tanggap fungsional dilakukan pada kerapatan inang 2, 4, 8, 12, 16, 20, dan 24 dengan analisis regresi logistik dan regresi non linear.
iv
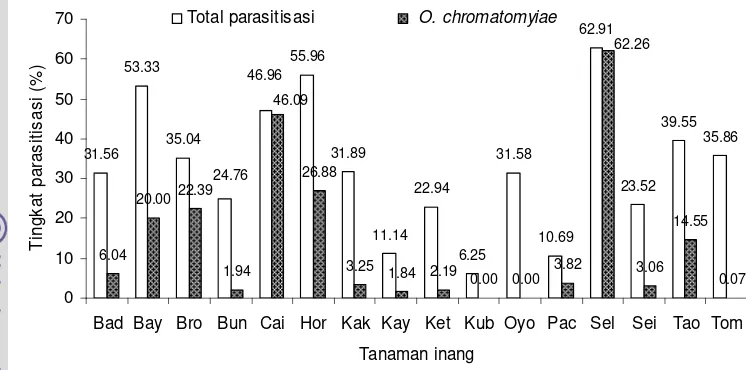
liriomyzae dan masing-masing satu spesies dari famili Braconidae dan Eucoilidae yaitu O. chromatomyiae dan Gronotoma micromorpha. Parasitoid O. chromatomyiae merupakan parasitoid yang dominan pada tingkat ketinggian 1001-1300 dan 1301-1600 m di atas permukaan laut dengan tingkat parasitisasi berkisar antara 1.84 – 62.26%. Dari 16 spesies tanaman yang diinfestasi oleh lalat pengorok daun, O. chromatomyiae dapat memparasitisasi lalat pengorok daun pada 13 tanaman yang terserang.
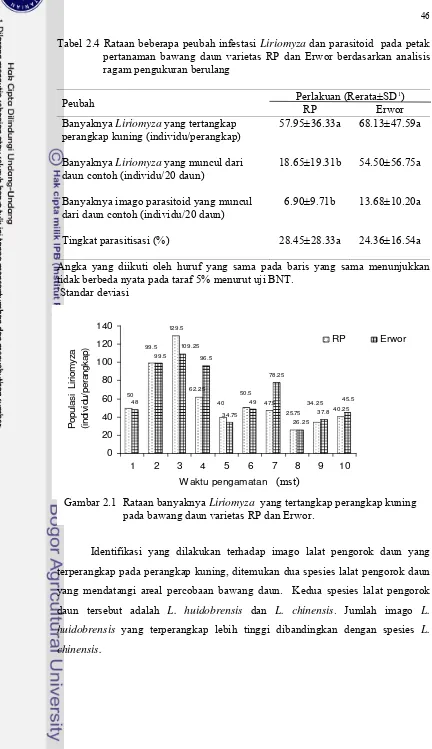
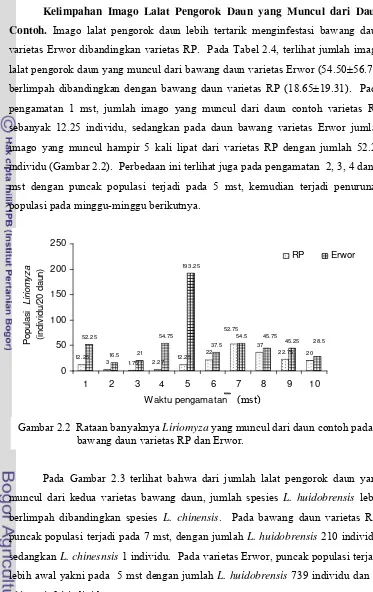
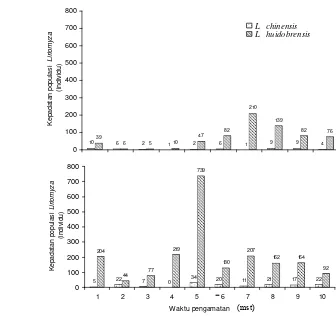
Analisis pengamatan berulang mingguan menunjukkan bahwa perbedaan varietas bawang daun tidak mempengaruhi jumlah Liriomyza yang tertangkap perangkap kuning namun mempengaruhi jumlah Liriomyza dan parasitoid yang muncul dari daun contoh. Terdapat dua spesies pengorok daun yang menyerang pertanaman bawang daun, yaitu L. huidobrensis dan L. chinensis dengan rerata jumlah Liriomyza yang muncul dari bawang daun varietas Erwor dan RP berturut-turut sebanyak 54,50 individu/20 daun dan 18.65 individu/20 daun. Jumlah parasitoid yang muncul dari bawang daun varietas Erwor (13.68 individu/20 daun) jauh berlimpah dibandingkan varietas RP (6.9 individu/20 daun). Spesies parasitoid yang muncul dari daun contoh kedua varietas adalah H. varicornis dan
O. chromatomyiae dengan tingkat parasitisasi kedua spesies hampir sama. Tingkat kerusakan tanaman lebih berat terjadi pada bawang daun varietas Erwor namun tidak mempengaruhi jumlah anakan karena jumlah anakan pada varietas Erwor dua kali lipat dibandingkan varietas RP.
Aplikasi insektisida abamektin, azadirahtin dan kartap tidak berpengaruh terhadap jumlah imago Liriomyza yang terperangkap perangkap kuning namun berpengaruh terhadap jumlah Liriomyza yang muncul dari daun, tingkat kerusakan tanaman dan parasitoid yang berasosiasi dengan lalat ini. Insektisida kartap lebih efektif dalam mengendalikan lalat pengorok daun pada tanaman bawang daun sehingga tingkat kerusakan tanaman jauh lebih rendah dibandingkan dua insektisida lainnya. Namun demikian aplikasi insektisida kartap yang merupakan insektisida sintetis menyebabkan jumlah parasitoid O. chromatomyiae dan H. varicornis yang muncul dari daun lebih rendah dibandingkan pada perlakuan insektisida abamektin dan azadiraktin. Aplikasi abamektin dan azadirakhtin cukup mampu menurunkan tingkat serangan lalat pengorok daun dan aman terhadap musuh alami. Aplikasi insektisida tidak berpengaruh terhadap bobot kotor bawang daun dan jumlah anakan.
Opius chromatomyiae merupakan endoparasitoid yang memperlihatkan lama hidup imago betina (9.95 hari) lebih singkat dibandingkan imago jantan (11.73 hari). Keperidian imago betina 104.73 individu dengan laju peneluran 9.31 butir/hari, sementara itu, laju reproduksi bersih (Ro) 28.55 dengan masa generasi (T) 15.96 hari dan laju pertumbuhan intrinsik sebesar 0.21 pada proporsi persebaran usia stabil 37.93% telur, 24.92% larva, 20.36% pupa dan 16.78% imago. O. chromatomyiae mempunyai tanggap fungsional tipe II dengan laju pencarian inang 0.0777/jam dan masa penanganan inang 2.5818 jam. Jumlah inang yang diparasit O. chromatomyie meningkat dengan bertambahnya kerapatan inang.
v
©
Hak Cipta Milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebahagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
vi
AGROMYZIDAE) DAN PARASITOIDNYA PADA SAYURAN
DATARAN TINGGI DENGAN PERHATIAN UTAMA PADA
PARASITOID Opius chromatomyiae BELOKOBYLSKIJ &
WHARTON (HYMENOPTERA: BRACONIDAE)
RUSLI RUSTAM
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi/Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
vii Penguji pada Ujian Tertutup : Dr. Ir. R. Yayi Munara Kusumah, M.S.
Penguji pada Ujian Terbuka : Prof. Dr. Ir. Utomo Kartosuwondo, M.S.
viii dan Parasitoidnya pada Sayuran Dataran Tinggi dengan Perhatian Utama pada Parasitoid Opius chromatomyiae
Belokobylskij & Wharton (Hymenoptera: Braconidae) Nama : Rusli Rustam
NIM : A461040021
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Aunu Rauf, M.Sc. Dr. Ir. Nina Maryana, M.S. Ketua Anggota
Dr. Ir. Pudjianto, M.S. Dr. Ir. Dadang, M.Sc. Anggota Anggota
Mengetahui
Ketua Program Studi Dekan Sekolah Pascasarjana Entomologi/Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
ix Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia- Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Januari 2006 ini adalah parasitoid, dengan judul Lalat Pengorok Daun Liriomyza spp. (Diptera: Agromyzidae) dan Parasitoidnya pada Sayuran Dataran Tinggi dengan Perhatian Utama pada Parasitoid Opius chromatomyiae Belokobylskij & Wharton (Hymenoptera: Braconidae.
Disertasi ini memuat satu judul yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal ilmiah. Judul 1, Lalat pengorok daun dan parasitoidnya pada pertanaman sayuran dataran tinggi di Kabupaten Cianjur-Bogor, Jawa Barat telah diterbitkan (JNI 11: 1 tahun 2008).
Terima kasih penulis ucapkan kepada Prof. Dr. Ir. Aunu Rauf, M.Sc., Dr. Ir. Nina Maryana, M.S., Dr. Ir. Pudjianto, M.S., dan Dr. Ir. Dadang, M.Sc. yang telah membimbing penulis dengan sabar dan hati yang ikhlas mulai dari penyusunan proposal sampai penulisan disertasi ini. Penulis juga mengucapkan terima kasih kepada Dekan Fakultas Pertanian Universitas Riau (UNRI) dan Bapak Rektor UNRI, atas izin dan kesempatan yang diberikan kepada penulis untuk mengikuti program Doktor (S3) di IPB. Terima kasih disampaikan pula kepada Direktur Program pascasarjana IPB dan Tim Pengelola Beasiswa BPPS Depdiknas atas kesempatan dan dukungan biaya yang telah diberikan sehingga proses penyelesaian studi penulis dapat berjalan dengan baik. Kepada Departemen Proteksi Tanaman, Fakultas Pertanian IPB, khususnya Program Studi Entomologi-Fitopatologi penulis sangat berterima kasih atas kesungguhan hati para dosennya dalam mentransfer ilmu pengetahuan melalui perkuliahan, diskusi, dan seminar.
Ucapan terima kasih penulis sampaikan pula kepada Bapak Saodik, Bapak Wawan Laboran Laboratorium Ekologi Serangga dan Ibu Aisyah Laboran Laboratorium Biosistematika Serangga yang telah banyak membantu penulis selama melaksanakan penelitian. Kepada saudara Acep yang telah membantu penulis dalam melaksanakan penelitian di Cipanas, penulis ucapkan terima kasih.
Kepada rekan di program studi Entomologi/Fitopatologi dan rekan-rekan di laboratorium Ekologi Serangga dan Biosistematika Serangga, penulis mengucapkan terima kasih atas budi baik dan kerjasama yang telah penulis rasakan selama ini. Rekan-rekan telah banyak memberi bantuan melalui kegiatan diskusi dan tukar pikiran yang bermanfaat bagi penulis. Akhirnya penulis menyampaikan terima kasih kepada seluruh keluarga, khususnya kepada istri tercinta Jum’atri Yusri atas bantuan moril dan materil serta keihklasannya selama penulis mengikuti pendidikan di IPB. Disertasi yang tidak luput dari kekurangan dan kelemahan ini mudah-mudahan dapat bermanfaat bagi kita semua.
Bogor, Januari 2009
x Penulis dilahirkan di Padang pada tanggal 11 November 1969 sebagai anak keenam dari sembilan bersaudara dari pasangan Rustam (alm.) dan Rosna Zein (almh.). Pendidikan sarjana ditempuh di Jurusan Hama dan Penyakit Tumbuhan, Fakultas Pertanian, Universitas Andalas Padang, lulus tahun 1993. Pada tahun 2000, penulis diterima di Program Studi Entomologi-Fitopatologi, Program Pascasarjana IPB dan menamatkannya pada tahun 2002. Kesempatan untuk melanjutkan ke program doktor pada program studi dan pada perguruan tinggi yang sama diperoleh pada tahun 2004. Beasiswa pendidikan pascasarjana diperoleh dari Departemen Pendidikan Nasional Republik Indonesia.
Penulis bekerja sebagai staff pengajar di Program Studi Hama dan Penyakit Tumbuhan, Jurusan Agronomi, Fakultas Pertanian, Universitas Riau sejak tahun 1999. Mata kuliah yang menjadi tanggung jawab penulis adalah Dasar-Dasar Perlindungan Tanaman.
Selama mengikuti program S3, penulis menjadi anggota Perhimpunan Entomologi Indonesia (PEI). Karya ilmiah berjudul Lalat Pengorok Daun dan Parasitoidnya pada Pertanaman Sayuran Dataran Tinggi di Kabupaten Cianjur dan Bogor, Jawa Barat telah disajikan pada Seminar Nasional PEI Bogor di Cibinong-Bogor pada bulan Maret 2008. Studi parasitoid Opius chromatomyiae
xi
xii 3. APLIKASI BEBERAPA JENIS INSEKTISIDA UNTUK MENGEN-
DALIKAN LALAT PENGOROK DAUN DAN PENGARUH SAM-PINGNYA TERHADAP PARASITOID
Abstrak ... 59
Abstract ... 59
Pendahuluan ... 60
Bahan dan Metode ... 61
Hasil dan Pembahasan ... 64
Kesimpulan ... 73
Daftar Pustaka ... 73
4. PARAMETER DEMOGRAFI PARASITOID Opius chromatomyiae (HYMENOPTERA: BRACONIDAE) PADA LALAT PENGOROK DAUN Liriomyza huidobrensis (DIPTERA: AGROMYZIDAE) Abstrak ... 75
Abstract ... 75
Pendahuluan ... 75
Bahan dan Metode ... 76
Hasil dan Pembahasan ... 79
Kesimpulan ... 85
Daftar Pustaka ... 85
5. TANGGAP FUNGSIONAL Opius chromatomyiae (HYMENOPTERA: BRACONIDAE) PADA LALAT PENGOROK DAUN Liriomyza huidobrensis (DIPTERA: AGROMYZIDAE) Abstrak ... 88
Abstract ... 88
Pendahuluan ... 88
Bahan dan Metode ... 89
Hasil dan Pembahasan ... 92
Kesimpulan ... 95
Daftar Pustaka ... 95
PEMBAHASAN UMUM ... 97
KESIMPULAN DAN SARAN ... 102
xiii
Halaman
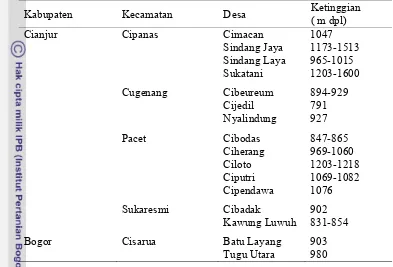
1.1 Lokasi pengambilan sampel daun ... 24 1.2 Spesies tanaman yang terserang lalat pengorok daun ... 25 1.3 Spesies lalat pengorok daun pada berbagai jenis tanaman inangnya . 29 1.4 Parasitoid yang menyerang lalat pengorok daun ... 29 1.5 Kelimpahan lalat pengorok daun dan tingkat parasitisasi parasitoid
pada berbagai jenis tanaman inang ... 30 1.6 Kelimpahan spesies Liriomyza dan parasitoid per daun pada
tanaman bawang daun ... 32 2.1 Deskripsi tanaman bawang daun ... 42 2.2 Kriteria skor kerusakan daun akibat serangan Liriomyza ... 44 2.3 Perbandingan beberapa peubah infestasi Liriomyza pada petak
pertanaman bawang daun varietas RP dan Erwor berdasarkan sidik ragam pengukuran berulang ... 45 2.4 Rataan beberapa peubah infestasi Liriomyza dan parasitoid pada
petak pertanaman bawang daun varietas RP dan Erwor berdasarkan analisis ragam pengukuran berulang ... 46 2.5 Perbandingan beberapa peubah parasitoid Liriomyza pada petak
pertanaman bawang daun varietas RP dan Erwor berdasarkan sidik ragam pengukuran berulang ... 49 3.1 Kriteria skor kerusakan daun akibat serangan lalat pengorok daun ... 63 3.2 Perbandingan beberapa peubah infestasi Liriomyza pada petak
pertanaman bawang daun yang diaplikasi insektisida dan petak kontrol berdasarkan sidik ragam pengukuran berulang ... 64 3.3 Rataan beberapa peubah infestasi Liriomyza pada petak pertanaman
bawang daun yang diaplikasi insektisida dan petak kontrol berdasarkan analisis ragam pengukuran berulang ... 66 3.4 Perbandingan kelimpahan parasitoid dan tingkat parasitisasi pada
pertanaman bawang daun yang diaplikasi beberapa jenis insektisida dan pada petak kontrol berdasarkan analisis ragam pengukuran berulang ... 68 3.5 Rataan beberapa peubah parasitoid pada pada petak pertanaman
bawang daun yang diaplikasi insektisida dan petak kontrol berdasarkan analisis ragam pengukuran berulang ... 68 3.6 Tingkat serangan, bobot bawang daun dan jumlah anakan pada
xiv 4.2 Parameter demografi Opius chromatomyiae ... 82 4.3 Proporsi berbagai fase perkembangan Opius chromatomyiae pada
persebaran umur stabil ... 84 5.1 Hasil analisis regresi logistik proporsi larva Liriomyza huidobrensis
yang terparasit oleh Opius chromatomyiae pada berbagai kerapatan inang ... 93 5.2 Nilai penduga parameter laju pencarian (a) dan masa penanganan
xv
Halaman
1.1 Kelimpahan populasi lalat pengorok daun pada tanaman bawang daun, brokoli, seledri, ketimun, dan kacang kapri pada berbagai ketinggian tempat ... 27 1.2 Kelimpahan populasi lalat pengorok daun pada tanaman kaylan,
tomat, horenso, buncis, tang och, bayam, dan caisim di berbagai ketinggian tempat ... 28 1.3 Komposisi parasitoid menurut ketinggian tempat ... 30 1.4 Tingkat parasitisasi Opius chromatomyiae ... 31 2.1 Rataan banyaknya Liriomyza yang tertangkap perangkap kuning
pada bawang daun varietas RP dan Erwor ... 46 2.2 Rataan banyaknya Liriomyza yang muncul dari daun contoh pada
bawang daun varietas RP dan Erwor ... 47 2.3 Kelimpahan populasi Liriomyza huidobrensis dan Liriomyza
chinensis pada bawang daun varietas RP dan Erwor ... 48 2.4 Kelimpahan populasi Hemiptarsenus varicornis dan Opius
chromatomyiae pada bawang daun variatas RP dan Erwor ... 50 2.5 Rataan tingkat parasitisasi parasitoid pada bawang daun varietas RP
dan Erwor ... 51 2.6 Perkembangan tingkat kerusakan tanaman pada bawang daun
varietas RP dan Erwor ... 51 2.7 Rerata jumlah anakan bawang daun per rumpun ... 52 3.1 Rataan banyaknya imago lalat pengorok daun yang tertangkap
perangkap kuning (ekor/perangkap) pada pertanaman bawang daun yang diaplikasikan insektisida ... 65 3.2 Jumlah Liriomyza huidobrensis (A) dan Liriomyza chinensis (B)
yang muncul dari tanaman bawang daun yang di aplikasikan abamektin, azadiraktin, kartap hidroklorida, dan kontrol ... 67 3.3 Skor kerusakan tanaman bawang daun akibat serangan Liriomyza
pada petak yang mendapat perlakuan abamektin, azadiraktin, kartap hidroklorida, dan kontrol ... 67 3.4 Rataan banyaknya Opius chromatomyiae (A) dan Hemiptarsenus
varicornis (B) yang muncul dari daun contoh yang dikoleksi dari tanaman bawang daun yang diaplikasikan abamektin, azadiraktin, kartap dan kontrol ...
xvi
chromatomyiae ... 84 5.1 Kurva tanggap fungsional (tipe II) Opius chromatomyiae pada
xvii
Halaman
1.1
Spesies lalat pengorok daun yang menyerang pertanaman sayuran pada dataran tinggi (skala pembanding adalah 1 mm) ... 38 1.2 Spesies parasitoid lalat pengorok daun dataran tinggi (skala
Latar Belakang
Dalam kurun waktu 14 tahun terakhir ini, pertanaman sayuran di Indonesia
diinvasi oleh tiga hama eksotik yang tergolong genus Liriomyza (Diptera:
Agromyzidae). Invasi diawali oleh L. huidobrensis (Blanchard) sekitar tahun
1994 (Rauf 1995), dan dua tahun kemudian diikuti oleh L. sativae Blanchard
(Rauf et al. 2000). Jenis yang ke tiga adalah L. chinensis Kato yang dilaporkan
pertama kali di Indonesia tahun 2000 (Rauf & Shepard 2001). Sebelumnya telah
terdapat satu spesies lalat pengorok daun yang keberadaannya sudah lama di
Indonesia, yakni Chromatomyiae horticola (Goureau) yang dikenal sebagai hama
penting pada tanaman kacang kapri (Kalshoven 1981).
Asal lalat pengorok daun Liriomyza spp. adalah Amerika Selatan (Spencer
1973) dan diperkirakan masuk ke Indonesia melalui perdagangan bunga potong
dan produk sayuran segar (Rauf 1995). Sekarang hama ini telah menyebar
hampir ke seluruh wilayah sayuran di Indonesia, menjadi hama utama pada
pertanaman sayuran dataran rendah sampai dataran tinggi. Menurut Rauf dan
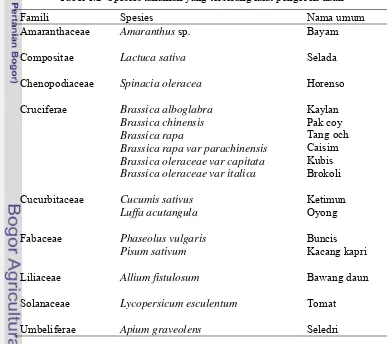
Shepard (1999), hama ini ditemukan pada 45 spesies tanaman dari Famili
Cruciferae, Liliaceae, Cucurbitaceae, Umbelliferae, Compositae, Amaranthaceae,
Chenopodiaceae, Solanaceae, Euphorbiaceae, Convolvulaceae, Basellaceae, dan
Labitaceae.
Kerusakan yang ditimbulkan ketiga spesies hama ini adalah sama, yaitu
kerusakan jaringan mesofil daun akibat aktifitas makan larva sehingga
menyebabkan kemampuan tanaman dalam melakukan fotosintesis menurun
(Minkenberg & van Lenteren 1986). Bekas tusukan ovipositor imago tampak
berupa bercak-bercak putih pada daun, sedangkan korokan yang dibuat larva
berbentuk seperti terowongan kecil yang berliku dan larvanya berada pada ujung
liang. Serangan berat dapat mengakibatkan daun mengering dan gugur sebelum
waktunya, sehingga menurunkan kuantitas dan kualitas hasil. Serangan hama ini
dapat menurunkan hasil antara 30% sampai 70% (Rauf & Shepard 1999).
Upaya pengendalian yang umum dilakukan oleh petani adalah penggunaan
insektisida profenofos, dengan frekuensi penyemprotan 2-3 kali per minggu (Rauf
serangan karena lalat pengorok daun yang menyebar ini diduga berasal dari
populasi yang telah resisten (Parrella & Keil 1984), dan karena larva berada dalam
jaringan tanaman sehingga terlindung dari pengaruh insektisida (Parrella 1987).
Di pihak lain, penggunaan insektisida yang berlebihan dikhawatirkan dapat
menimbulkan dampak buruk seperti resistensi dan resurgensi hama, terbunuhnya
musuh alami, dan pencemaran lingkungan secara umum (CEQ 1972).
Untuk mengatasi kerugian pada produksi pertanian dan kesulitan dalam
pengendalian hama pengorok daun karena tidak efektifnya insektisida yang
digunakan serta untuk mencegah atau mengurangi dampak penggunaan
insektisida seperti matinya musuh alami, perlu dicari alternatif pengendalian hama
tersebut.
Pendekatan Masalah
Di antara alternatif yang ada, pengendalian hama pengorok daun yang
dapat dipilih adalah pemanfaatan musuh alami. Musuh alami merupakan salah
satu komponen dalam pengendalian hama terpadu (PHT) yang aman terhadap
lingkungan sehingga keberhasilan parasitoid dalam menekan populasi hama akan
mengurangi penggunaan insektisida dalam pengendalian hama.
Parasitoid adalah salah satu kelompok musuh alami yang dapat
dimanfaatkan dalam pengendalian hama terpadu. Di antara rangkaian penelitian
yang telah dilakukan untuk mendukung pengendalian hama terpadu lalat pengorok
daun adalah survei musuh alami. Hingga kini di Indonesia dilaporkan terdapat 18
jenis parasitoid yang berasosiasi dengan larva Liriomyza spp. yaitu Asecodes
deluchii (Baucek), Chrysocharis sp., Cirropillus ambiguus (Hanson & LaSalle),
Closterocerus sp., Hemiptarsenus varicornis (Girault), Neochrysocharis formosa
(Westwood), Neochrysocharis sp., Pnigalio sp., Quadrastichus sp.,
Zagrammosoma sp., Stenomesius sp., Pediobius sp. (semuanya Hymenoptera:
Eulophidae), Gronotoma sp., Kleidotoma sp. dan Norlanderia sp. (Hymenoptera:
Eucoilidae), Opius chromatomyiae Belokobylskij & Wharton (Hymenoptera:
Braconidae) dan Sphegigaster sp. (Hymenoptera: Pteromalidae) (Supartha 1998;
Rauf et al. 2000; Susilawati 2002; Purnomo 2003). Pada pertanaman sayuran
dataran tinggi, jenis parasitoid yang paling dominan adalah H. varicornis dan O.
endoparasitoid larva-pupa (Rustam 2002), sedangkan H. varicornis merupakan
ektoparasitoid larva. Informasi parasitoid H. varicornis sebagai agens hayati lalat
pengorok daun di Indonesia telah banyak diteliti oleh Hidrayani (2003 ) dan
Thamrin (2004).
O. chromatomyiae adalah endoparasitoid yang umum ditemui memarasit
hama pengorok daun. Menurut Rauf dan Shepard (1999), tingkat parasitisasi
parasitoid sangat dipengaruhi oleh tanaman inang dan ketinggian tempat. Oleh
karena itu, survei lalat pengorok daun dan parasitoidnya, khususnya O.
chromatomyiae, menjadi sangat berguna dalam pemanfaatan parasitoid tersebut
sebagai agens pengendalian hayati. Lalat pengorok daun mudah beradaptasi
dengan lingkungan sehingga penyebaran lalat pengorok cepat terjadi di lapangan.
Penyebaran lalat pengorok daun akan diikuti oleh penyebaran parasitoidnya.
Dari survei pendahuluan yang dilakukan, bawang daun merupakan
tanaman yang dominan terserang lalat pengorok daun dan pengorok ini
menimbulkan kerugian yang besar. Informasi tentang dinamika populasi lalat
pengorok daun maupun parasitoidnya pada bawang daun menjadi sangat penting
untuk diketahui sehingga pengendalian lalat pengorok daun ini dapat dilakukan
dengan efisien dan efektif.
O. chromatomyiae merupakan parasitoid yang diandalkan dapat
mengendalikan lalat pengorok daun di daerah dataran tinggi. Namun dalam
pemanfaatan di lapangan perlu kajian yang lebih dalam seperti bioekologi
parasitoid dan hubungan parasitoid dengan tanaman inang, sehingga pelepasan di
lapangan akan lebih tepat dan bermakna. Kajian bioekologi parasitoid seperti
neraca kehidupan yang meliputi laju reproduksi, laju pertumbuhan instristik,
persebaran umur stabil, dan waktu generasi serta kajian tanggap parasitoid
terhadap kelimpahan inang (tanggap fungsional) menjadi sesuatu yang utama
dalam pemanfaatan parasitoid ini.
Penggunaan insektisida untuk mengendalikan serangga hama merupakan
sesuatu yang lazim. Dalam konsep pengendalian hama terpadu (PHT), insektisida
merupakan salah satu komponen yang keberadaanya kadang kala masih sangat
diperlukan di samping komponen pengendalian yang lain. Namun dalam
insektisida dapat berakibat buruk bagi keberadaan musuh alami lalat pengorok
daun (Johnson et al. 1980). Oleh karena itu, diperlukan penelitian tentang
insektisida yang dapat mengendalikan lalat pengorok daun, tetapi aman terhadap
musuh alami.
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) mengidentifikasi spesies lalat pengorok
daun pada pertanaman sayuran dataran tinggi serta parasitoid yang berasosiasi, (2)
mengetahui tingkat parasitisasi parasitoid khususnya O. chromatomyiae pada
berbagai jenis tanaman inang, (3) mengkaji dinamika populasi lalat pengorok
daun dan parasitoidnya pada tanaman bawang daun (4) mengevaluasi selektifitas
beberapa jenis insektisida terhadap parasitoid khususnya O. chromatomyiae (5)
mengevaluasi potensi keefektifan parasitoid O. chromatomyiae berdasarkan
parameter demografi yang meliputi laju pertambahan intristik (r), reproduksi
bersih (Ro), dan masa generasi (T), proporsi usia stabil (px), dan (6) mengevaluasi
potensi keefektifan parasitoid O. chromatomyiae berdasarkan tanggapnya
terhadap peningkatan kelimpahan inang, serta menetapkan laju pelacakan inang
(a) dan masa penanganan inang (Th).
Manfaat Penelitian
Hasil penelitian ini diharapkan pertanaman sayuran yang terserang lalat
pengorok daun dapat dikendalikan dengan parasitoid O. chromatomyiae sehingga
dapat meningkatkan kesejahteraan petani dan menghasilkan tanaman sayuran
TINJAUAN PUSTAKA
Lalat Pengorok Daun
Menurut Parrella (1987), lalat pengorok daun mempunyai lebih dari 300
spesies yang sebagian besar tersebar di daerah beriklim sedang dan beberapa
spesies ditemukan di daerah tropis. Spencer dan Steyskal (1986) menyebutkan
terdapat tiga spesies lalat pengorok daun yang bersifat polifag yaitu Liriomyza
huidobrensis (Blanchard), L. sativae Blanchard dan L. trifolii (Burgess).
Di Indonesia, lalat pengorok daun diperkirakan sudah ada sejak tahun
1990-an, namun pertama kali ditemukan dan diketahui sebagai hama sekitar bulan
Sepetember 1994 di daerah Cisarua, Bogor (Rauf 1995). Tiga spesies yang telah
dilaporkan banyak menimbulkan kerugian di berbagai daerah di Indonesia adalah
L. huidobrensis, L. sativae dan L. chinensis (Rauf & Shepard 2001).
Liriomyza huidobrensis (Blanchard)
Lalat pengorok daun, L. huidobrensis berasal dari Amerika Selatan. Hama ini tersebar di Eropa, Afrika, dan Asia. Persebaran di Asia meliputi Malaysia,
Indonesia, Filipina, Vietnam, Thailand, Srilangka, India, Pakistan, Laos, China,
dan Banglades (Weintraub & Horowitz 1994; Rauf & Shepard 2001). Di
Indonesia L. huidobrensis dilaporkan telah ditemukan di Jawa, Sumatra dan
Sulawesi Selatan (Rauf 1995).
Imago L. huidobrensis berukuran 1.7-2.3 mm. Imago betina umumnya
berukuran lebih besar dari imago jantan, dan muncul dari puparium yang
berukuran lebih besar (Parrella 1987). Lama hidup imago betina berkisar antara
6-14 hari, sedang jantan 3-9 hari (Supartha 1998).
Betina meletakan telur rata-rata 8-14 butir per hari. Jumlah telur yang
diletakkan selama hidupnya berkisar antara 42-301 butir (Supartha 1998). Telur
diletakkan satu per satu pada permukaan daun (Parrella 1987). Telur bening dan
berukuran 0.28 x 0.5 mm (Parrella 1987). Stadium telur berlangsung selama 2-4
hari (Supartha 1998). Larva terdiri atas tiga instar, dengan waktu perkembangan
untuk setiap instar 2-4 hari (Supartha 1998). Periode puparium berkisar antara
Liriomyza sativae Blanchard
Menurut Spencer (1973), negara asal L. sativae adalah Amerika. Di
Indonesia, L. sativae telah tersebar di dataran rendah Jalur Pantura mulai dari
Karawang sampai Brebes. Selain itu, L. sativae juga tersebar di daerah
Bogor-Jawa Barat dan Banjar-Kalimantan Selatan (Rauf & Shepard 2001; Susilawati
2002).
Imago L. sativae berukuran kecil yaitu panjang 1.5 mm dengan warna
hitam kecoklatan dan terdapat bintik kuning pada tubuhnya. Imago betina lebih
besar dibandingkan dengan imago jantan. Lama hidup imago berkisar antara
10-20 hari tergantung pada kondisi lingkungan dengan kemampuan menghasilkan
telur 600-700 butir (Mau & Kessing 1991; Capinera 2001).
Telur L. sativae bewarna putih, berbentuk lonjong dengan panjang 0.23
mm dan lebar 0.13 mm. Telur diletakkan satu per satu di dalam jaringan mesofil,
di bawah permukaan daun. Tiga hari kemudian telur akan menetas menjadi larva.
Larva terdiri dari tiga instar dan masing-masing instar memerlukan waktu
pertumbuhan 2-3 hari (Mau & Kessing 1991; Capinera 2001). Larva instar akhir
akan keluar dari daun dan menjatuhkan diri ke tanah untuk membentuk puparium
(Parrella 1987). Periode puparium berlangsung selama 5-12 hari (Mau & Kessing
1991)
Liriomyza chinensis (Kato)
L. chinensis berasal dari daerah Asia bagian Timur (Taiwan dan Jepang)
dan sekarang telah tersebar di Eropa (Spencer 1973; Dempewolf 2004). L.
chinensis diduga masuk ke Indonesia melalui sayuran yang diimpor dari wilayah
Asia bagian Timur (Rauf 2001).
Imago L. chinensis berukuran 2.00-2.39 mm. Imago betina mempunyai
ukuran yang lebih besar dari imago jantan (Nawin 2003). Imago yang baru
muncul bersifat fototaksis positif dan naik ke bagian atas tanaman (Parrella 1987).
Lama hidup imago berkisar antara 8-12 hari dengan kemampuan menghasilkan
telur 17-95 butir (Nawin 2003).
Telur L. chinensis bewarna putih bening yang diletakkan secara tunggal,
tetapi seringkali diletakkan secara bertumpuk. Larva yang baru keluar dari telur
kekuningan yang mengecil pada bagian depan. Larva terdiri atas empat instar
(Parrella 1987). Larva instar akhir akan keluar dari rongga daun untuk
membentuk puparium di dalam tanah. Puparium berwarna kuning hingga coklat
kekuningan (Nawin 2003).
Chromatomyia horticola Goureau
C. horticola tersebar di Eropa, Afrika Utara, Mesir, Libia, Maroko, Afrika
Selatan, Madagaskar, Kamerun, Kenya, Turki, India, Cina, Jepang, Iran, Filipina,
Malaysia, Korea dan Nepal (Spencer 1973; Dempewolf 2004). Hama ini telah
lama dilaporkan keberadaanya di Indonesia (Kalshoven 1981).
Seekor betina C. horticola dapat menghasilkan 100-500 telur, dan setiap
harinya mampu meletakkan 50 butir telur (Dempewolf 2004). Telur diletakkan
oleh betina dalam permukaan atas dan permukaan bawah daun (Tandon 1965
dalam Dempewolf 2004). Telur menetas menjadi larva instar awal 1-4 hari
setelah peletakan. Periode larva berkisar antara 5-6 hari. Larva mengorok daun
secara tidak teratur.
Larva berkembang menjadi puparium dengan periode puparium selama
9-5 hari. Pupa tetap tinggal di dalam jaringan daun. Puparium bewarna abu-abu.
Imago bewarna hitam dengan panjang sayap 2.2 – 2.7 mm (Dempewolf 2004).
Kerusakan dan Kehilangan Hasil Akibat Serangan Lalat Pengorok Kerusakan pada tanaman dapat disebabkan oleh aktifitas peneluran dan
makan imago serta korokan larva. Perilaku peneluran dan makan imago serta korokan larva Liriomyza dapat menyebabkan dua bentuk kerusakan. Bentuk
kerusakan pertama disebabkan oleh tusukan ovipositor imago untuk keperluan
peneluran dan makan, yang merusak sel-sel jaringan daun tanaman. Gejala
kerusakan yang ditimbulkan adalah bintik-bintik putih pada permukaan daun atas
(Spencer 1973). Bentuk kerusakan ke dua disebabkan oleh larva yang memakan
jaringan dengan membuat korokan berliuk-liuk seperti gambar gerakan ular yang
menyebabkan rusaknya sel-sel jaringan pagar dan bunga karang mesofil (Cardova
& Karel 1990). Kedua bentuk kerusakan tersebut dapat mengurangi luas bagian
daun yang berfotosintesis sehingga menurunkan hasil dan kualitas daun sebagai
Gejala serangan lalat pengorok daun dimulai pada daun bagian bawah,
kemudian daun bagian tengah, dan yang lebih parah pada bagian pucuk (Cisneros
& Mujica 2000). Chavez dan Raman (1987) melaporkan bahwa di Amerika
Serikat dan Peru, serangan L. huidobrensis dapat mengakibatkan kehilangan hasil
pada kentang sekitar 35% sedang di Lembang kehilangan hasil sekitar 34%
(Soeriaatmadja & Udiarto 1996). Lebih lanjut dikemukakan oleh Rauf et al.
(2000) bahwa berdasarkan hasil survei di Bandung dan Garut (Jawa Barat),
Banjarnegara dan Wonosobo (Jawa Tengah), Alahan Panjang (Sumatera Barat),
dan Karo (Sumatera Utara), petani setempat melaporkan kehilangan hasil akibat
serangan Liriomyza pada tanaman kentang sekitar 30 – 70%.
Faktor yang Mempengaruhi Perkembangan Populasi Pengorok Daun Naik turunnya populasi organisme di suatu ekosistem dipengaruhi oleh
faktor-faktor lingkungan fisik, inang atau sumber pakan, ruang populasi itu
sendiri, dan spesies atau organisme lain. Faktor-faktor tersebut berperan dalam
menentukan laju natalitas, mortalitas dan migrasi (Dent 1995).
Faktor lingkungan fisik antara lain adalah suhu dan kelembaban, baik pada
tajuk tanaman maupun di permukaan tanah. Lama perkembangan Liriomyza lebih
singkat pada suhu yang tinggi. Pada suhu 20 °C, lama perkembangan berkisar
antara 23.7-27.0 hari, sedang pada suhu 25 °C berkisar antara 15.0-20.9 hari
(Parrella 1987). Perkembangan L. trifolii dengan tingkat mortalitas rendah
membutuhkan suhu optimum sekitar 25-30 °C (Leibee 1984). Pada tanaman
krisan, imago yang keluar dari puparium pada suhu 15.6, 21.2, 26.7, 32.2, dan
37.8 °C masing-masing adalah 68, 80, 92.5, 75.5, dan 0 % (Minkenberg & van
Lenteren 1986). Pada suhu 15 °C kematian L. trifolii tinggi terutama pada fase
telur dan larva (Parrella 1987; Minkenberg 1988).
Kelembaban relatif udara
mempengaruhi perilaku makan, jumlah imago yang muncul dan daya bertahan
hidup. Jumlah puparium yang berhasil menjadi imago meningkat bila udara pada
permukaan pupariumnya lebih lembab. Pada tanaman kentang, populasi
Liriomyza meningkat pada musim kering dan menurun bila hujan turun terus
menerus (Debiyanto et al. 1996). Populasi puparium meningkat bila kelembaban
pada kelembaban 76-100% dapat dihasilkan puparium sebanyak 65-88% (Parrella
1987).
Perkembangan populasi Liriomyza spp. dipengaruhi oleh jenis, kuantitas,
dan kualitas tanaman inang sebagai sumber makanan. Distribusi dan kerapatan
trikoma, kandungan fenolat dan nutrisi tanaman mempengaruhi pemilihan
tanaman inang oleh imago Liriomyza spp. (Fagoonee & Toory 1983). Kandungan
N (nitrogen) daun tanaman inang berkorelasi positif dengan kelimpahan populasi
Liriomyza di lapangan (Minkenberg & van Lenteren 1986; Parrella 1987). Pada
kandungan nitrogen daun yang lebih tinggi, aktifitas makan dan keperidian
Liromyza meningkat (Minkenberg & van Lenteren 1986). Ketersediaan berbagai
jenis tanaman inang di lapangan selain membantu pertumbuhan dan
perkembangan serangga, juga membantu pemencarannya. Selain itu, sifat polifag
yang dimiliki Liriomyza spp. memungkinkan memencar lebih cepat (Supartha
1998).
Faktor spesies atau organisme lain meliputi pesaing dan musuh alami.
Musuh alami Liriomyza yang banyak ditemukan adalah predator dan parasitoid
(Minkenberg & van Lenteren 1986).
Liriomyza dapat dimangsa oleh berbagi jenis serangga predator
(Coleoptera, Hemiptera, Diptera, Dermaptera, Hymenoptera) dan laba-laba
(Cisneros & Mujica 2000; Harwanto et al. 2002). Hampir semua predator bersifat
generalis, kecuali Dolichopodidae dan Empididae (Diptera) yang bersifat
spesialis. Kemampuan predator dalam mempengaruhi perkembangan populasi
Liriomyza sulit untuk dievaluasi (Cisneros & Mujica 2000).
Ragam spesies parasitoid Liriomyza spp. tidak selalu sama pada
masing-masing jenis tanaman inang dan daerah geografis yang berbeda (Minkenberg &
van Lenteren 1986). Terdapat 40 spesies parasitoid yang memarasit Liriomyza
spp. pada fase larva dan puparium (Oatman 1959 dalam Supartha 1998),
diantaranya ada empat famili yang berasosiasi dengan L. trifolii dan L. sativae
pada tanaman tomat, yaitu Braconidae, Cynipidae, Eulophidae, dan Pteromalidae
(Schuster & Wharton 1993).
Di Indonesia telah diketahui 18 spesies parasitoid pengorok daun L.
Braconidae (Rauf et al. 2000; Susilawati 2002). Hemiptarsenus varicornis
(Eulophidae) dan Opius chromatomyiae (Braconidae) merupakan spesies yang
dominan ditemukan pada pertanaman sayuran dataran tinggi (Purnomo 2003).
Pengendalian Lalat Pengorok Daun
Teknik pengendalian yang umum dilakukan terhadap lalat pengorok daun
di negeri asalnya adalah cara budidaya, kimiawi, dan pengendalian hayati. Cara
budidaya yang biasa diterapkan adalah sanitasi, penyiangan gulma, pemangkasan
bagian daun yang terserang dan pemasangan penutup tanah dari plastik hitam.
Penggunaan perangkap kuning berperekat di rumah-rumah kaca juga hasilnya
dinilai kurang memuaskan sehingga kurang layak dimasyarakatkan (Minkenberg
& van Lenteren 1986). Walaupun demikian, cara pengendalian yang disebut
terakhir di Peru dinilai efektif mengendalikan L. huidobrensis (Chaves & Raman
1987; Raman 1988; Raman & Radcliffe 1992).
Insektisida yang mengandung zat pengatur tumbuh (ZPT) serangga lebih
efektif dibandingkan dengan insektisida lain yang berspektrum luas yang telah
digunakan dalam usaha pengendalian Liriomyza spp. Dua jenis ZPT tersebut
adalah Tripene ZR-619 dan Kinoprene ZP-777. Kedua ZPT tersebut dilaporkan
sangat berbahaya bagi parasitoid penting seperti Opius dimiatus Ash.
(Minkenberg & van Lenteren 1986).
Penggunaan insektisida kimiawi seperti organoklorin, organfosfat dan
karbamat yang berspektrum luas sudah lama direkomendasikan sebagai cara
pengendalian hama, walaupun cara tersebut dinilai terlalu berbahaya. Cardova
dan Karel (1990) melaporkan bahwa penggunaan insektisida telah banyak
memacu ledakan hama tersebut. Menurut Schreiner et al. (1986), Liriomyza
menjadi resisten terhadap sejumlah insektisida piretroid yang digunakan seperti
permetrin, fenvalerat, dan sipermetrin. Masalah resistensi tersebut telah
dilaporkan sekitar 20 tahun yang lalu oleh Chaves dan Raman (1987). Oleh
karena itu, kalangan peneliti mencari alternatif insektisida lain yang lebih selektif
dan efektif (Spencer 1973). Untuk kasus resistensi tersebut Keil dan Parrella
(1983 dalam Minkenberg & van Lenteren 1986) menyarankan agar melakukan
pergiliran penggunaan jenis insektisida. Selain itu, penggunaan insektisida harus
ekonominya berdasarkan keadaan populasi lalat dan musuh alaminya di lapangan
(Trumble 1985).
Pengendalian hayati Liriomyza spp. dengan menggunakan parasitoid telah
banyak dilakukan. Semenjak tahun1980, pengendalian hayati itu telah dilakukan
di Belanda pada areal seluas lebih kurang 30 ha per tahun. Dalam tahun 1985
dengan pelepasan sebanyak 10 000-20 000 per ha dengan perbandingan kira-kira
satu parasitoid per empat tanaman, telah menunjukkan keberhasilan pengendalian
Liriomyza spp. selama musim tanam. Dacnusa sibirica Telenga (Hymenoptera:
Braconidae) diketahui efektif mengendalian L. trifolii di Belanda, sedangkan di
Inggris dan Swedia, pengendalian dengan D. sibirica tidak berhasil (Minkenberg
1990). Hal ini dapat disebabkan oleh beberapa faktor, di antaranya adalah
kemungkinan terjadinya imigrasi lalat pengorok, waktu pelepasan parasitoid yang
tidak tepat, jumlah parasitoid yang dilepas tidak mencukupi dan mungkin kualitas
parasitoid tidak bagus (Minkenberg 1990).
Faktor yang Mempengaruhi Keberadaan Musuh Alami
Agroekosistem tanaman semusim merupakan ekosistem ephemeral, yang
berarti usianya singkat dan sering mengalami gangguan. Jenis gangguan yang
sering terjadi pada agroekosistem adalah praktek budidaya misalnya pengolahan
tanah, penyiangan, pemakaian insektisida dan pemanenan. Praktek budidaya
tersebut secara ekologi sering tidak mendukung kehidupan musuh alami. Aplikasi
insektisida menyebabkan musuh alami terbunuh, dan praktek budidaya bersih
menyebabkan tanaman inang dan sumber daya tambahan berkurang
ketersediannya (DeBach 1973). Keadaan tersebut pada akhirnya berdampak
terhadap agroekosistem (Altieri & Whitcomb 1979).
Menurunnya keseimbangan dan keanekaragaman dalam agroekosistem,
terutama keanekaragaman musuh alami dapat mendorong terjadinya ledakan
hama. Upaya pengelolaan agroekoisistem yang dapat meningkatkan
keanekaragaman musuh alami dan sekaligus menurunkan kerapatan populasi
hama di antaranya adalah diversifikasi habitat seperti mempertahankan vegetasi
liar di lahan pinggir, tumpang sari, rotasi, tanaman penutup tanah, bahan organik,
tanam tidak serempak dan pengolahan tanah minimum (Altieri & Whitcomb
Pemanfaatan parasitoid sebagai agens pengendalian hayati hama dapat
diterapkan melalui metode konservasi dan augmentasi inundatif. Metode
konservasi diarahkan untuk mempertahankan pelepasan musuh alami agar tetap
lestari, sedangkan augmentasi inundatif merupakan pelepasan musuh alami dalam
jumlah yang relatif besar dengan harapan segera dapat mengendalikan populasi
hama dalam waktu yang singkat (Sosromarsono 1999). Sebelum metode tersebut
dilaksanakan, terlebih dahulu harus diketahui karakter biologi, perilaku, serta
kondisi ekologis yang diperlukan agar musuh alami dapat bekerja dengan baik di
lapangan.
Pemilihan Inang oleh Parasitoid
Sebagian besar parasitoid Hymenoptera dapat memarasit beberapa jenis
inang dan hanya sedikit spesies yang spesifik memarasit satu spesies inang.
Parasitoid yang spesifik tersebut, pada kondisi laboratorium bahkan juga sering
dapat dipelihara pada inang lain yang secara alamiah bukan merupakan inang
karena adanya hambatan waktu dan ruang yang memisahkannya. Kenyataan
bahwa parasitoid dapat dibiakkan di laboratorium dengan serangga bukan inang
alamiah menjadi penting dalam pembiakkan masal parasitoid (Doutt 1959).
Menurut Doutt (1959), terdapat empat tahapan yang harus dilewati agar
parasitoid berhasil memarasit inangnya, yaitu 1) penemuan habitat inang, 2)
penemuan inang, 3) penerimaan inang, dan 4) kesesuaian inang. Selanjutnya
Vinson (1981) menambahkan pengaturan inang sebagai tahap yang kelima karena
keberhasilan parasitisme juga ditentukan oleh kemampuan parasitoid dalam
mengatur fisiologi inangnya.
Dalam penemuan habitat inang, parasitoid terutama dipandu oleh
rangsangan kimia yang berasal dari senyawa-senyawa volatil tanaman.
Rangsangan tersebut dapat berupa bau yang berasal dari makanan atau tanaman
yang terluka atau yang rusak, organisme yang berasosiasi dengan inang atau inang
itu sendiri. Tanaman merupakan isyarat utama karena tanaman mempunyai peran
yang dominan dalam mendukung suatu habitat yang khas. Akibatnya, suatu
parasitoid terkadang tertarik pada tanaman tertentu meskipun di situ tidak terdapat
inang. Parasitoid juga memarasit inang yang terdapat pada jenis tanaman tertentu
Penemuan inang oleh parasitoid dipandu oleh rangsangan fisik dan kimia
yang dikeluarkan oleh inang. Rangsangan fisik yang berperan terutama suara dan
gerakan. Rangsangan kimia dapat dibagi menjadi dua kelompok. Pertama,
rangsangan kimia yang dapat diterima dari jarak jauh misalnya bau inang.
Rangsangan yang diterima memungkinkan parasitoid untuk melokalisasi areal
pencarian inang. Kedua, rangsangan kimia yang dapat dideteksi hanya dari jarak
dekat, yaitu setelah terjadi kontak fisik. Rangsangan ini biasanya berasal dari
senyawa-senyawa padat atau cair misalnya kotoran inang, sekresi dari kelenjar
labium inang, produk inang lain dan bekas parasitoid lain. Adanya rangsangan ini
memungkinkan terjadinya kontak antara parasitoid dengan inangnya yang
dicirikan oleh perilaku pengujian oleh parasitoid berupa pergerakan memutar
dengan cepat dan perubahan kecepatan pergerakan. Faktor lain yang ikut
menentukan penemuan inang adalah pengalaman dan perilaku orientasi parasitoid
(Weseloh 1981).
Penerimaan inang atau pengenalan inang adalah proses diterima atau
ditolaknya inang untuk peletakkan telur setelah terjadi kontak (Arthur 1981).
Proses tersebut dibagi dalam empat fase yaitu: 1) kontak dan pemeriksaan, 2)
penusukan dengan ovipositor, 3) pemasukan ovipositor dan 4) peletakan telur.
Keempat fase tersebut harus lengkap dan berurutan sehingga bila terjadi hambatan
pada salah satu fase, proses dimulai lagi dari awal.
Seperti halnya tahap sebelumnya, penerimaan inang juga dipandu terutama
oleh rangsangan fisik dan kimia. Selain itu, pengalaman parasitoid sebelumnya,
termasuk tempat perkembangan parasitoid, juga akan berpengaruh pada proses
penerimaan inang. Rangsangan fisik yang berperan adalah kondisi fisik inangnya
seperti ukuran, bentuk, tekstur atau bentuk permukaan, warna dan kandungan air.
Rangsangan lainnya adalah pergerakan inang misalnya kegiatan makan inang dan
perkembangan embrio dalam telur. Rangsangan kimia dapat berasal dari
senyawa-senyawa yang terdapat di luar dan di dalam tubuh inang yang dapat
dideteksi dengan antena, tarsi atau ovipositor. Senyawa-senyawa tersebut dapat
disekresikan melalui kutikula, disekresikan bersama-sama kotoran atau terdapat
Kesesuaian inang yang menentukan keberhasilan perkembangan parasitoid
sampai menjadi imago tergantung pada beberapa faktor, yaitu: 1) kemampuan
parasitoid dalam menghindari atau melawan sistim pertahanan inang, 2) kompetisi
dengan parasitoid lain, 3) adanya toksin yang mengganggu atau merusak telur
atau larva parasitoid, dan 4) kesesuaian makanan parasitoid. Faktor lain yang
berpengaruh adalah infeksi patogen, kerentanan inang, faktor lingkungan dan
pengaruh hormon-hormon pengendali serangga (Vinson & Iwantsch 1980).
Biologi Opius sp.
Parasitoid Opius sp. merupakan endoparasit larva-pupa. Studi biologi dan
morfologi O. dissitus telah diteliti pada inang L. trifolii oleh Bordat et al. (1995),
O. melleus Gahan pada larva lalat buah Famili Tephritidae (Lathrop & Newton
1933) dan Opius sp. pada inang L. huidobrensis (Rustam 2002).
Telur O. dissitus berbentuk lonjong dengan warna putih dan tembus
pandang. Ukuran telur rata-rata 0.22 mm dan stadium telur 1-3 hari (Bordat et al.
1995), sedangkan stadium telur O. melleus lebih lama yakni 3-6 hari (Lathrop &
Newton 1933). Pada inang L. huidobrensis, stadium telur Opius sp. adalah 2 hari
dengan ukuran telur lebih besar dibandingkan dengan O. dissitus yakni 0.26 mm
(Rustam 2002). Setelah telur diletakkan, telur mengalami pertumbuhan dan
perkembangan di dalam tubuh inang.
Larva O. dissitus terdiri atas dua instar, dengan ukuran masing-masing
0.47 mm dan 0.99 mm, sedangkan O. melleus, Diachasma tryoni (Hymenoptera:
Braconidae), larva terdiri atas empat instar (Pemberton & Willard 1918; Lathrop
& Newton 1933; Bordat et al. 1995). Larva O. chromatomyiae terdiri atas dua
instar dengan ukuran yang lebih besar dibandingkan dengan larva O. dissitus dan
O. melleus (Rustam 2002). Larva instar satu mempunyai bagian kapsul kepala
yang kokoh dengan dua pengait yang runcing, tubuhnya ramping dan pada
ujungnya terlihat kasar. Larva instar dua berbentuk bulat, dimana pengaitnya
telah hilang dan berada dalam puparium inang yang panjang dengan warna putih
krem (Bordat et al. 1995; Rustam 2002) dan bersifat tidak aktif (Lathrop &
Newton 1933).
Pembentukan pupa ditandai dengan terbentuknya tonjolan bakal tungkai
kepala, toraks dan abdomen dapat terlihat jelas. Pada awalnya pupa berwarna
kuning pucat dan lama kelamaan berwarna gelap. Tubuh pupa yang berumur
lanjut berwarna hitam dengan ukuran 1.62 mm (Bordat et al. 1995; Rustam 2002),
pembentukan pupa terjadi di dalam puparium inang (Pemberton & Willard 1918)
dan masa perkembangan pupa berlangsung selama 5.93 hari (Rustam, 2002).
Imago O. chromatomyiae berwarna coklat kehitaman dengan ukuran yang
hampir sama antara jantan dan betina, yaitu rata-rata 1.72 mm dan 1.80 mm.
Jantan dan betina sulit dibedakan dengan mata biasa, namun dengan bantuan kaca
pembesar terlihat betina mempunyai ovipositor sebagai alat untuk peletakan telur
(Rustam 2002). Antena imago O. dissitus panjang, hitam, dan panjangnya hampir
sama dengan tubuhnya (Bordat et al. 1995). Pada O. chromatomyiae jantan dan
betina sudah bisa dibedakan berdasarkan jumlah ruas antenanya dimana antena
jantan terdiri atas 23 ruas sedangkan betina terdiri atas 25 ruas (Rustam 2002).
Pada O. melleus ovipositor relatif panjang dan merupakan ciri khasnya (Lathrop
& Newton 1933).
Imago parasitoid keluar dengan cara merobek puparium inang. Opius
betina yang baru muncul dapat melakukan parasitisasi dengan menusukkan
ovipositornya ke dalam rongga tubuh larva inang tanpa mematikan atau
melumpuhkannya. Larva inang terparasit tetap hidup dan mengorok daun hingga
menjadi pupa. Parasitoid tetap berkembang di dalam tubuh inang sampai keluar
menjadi imago (Bordat et al. 1995; Rustam 2002). Clausen (1940) melaporkan
bahwa imago betina O. fulvicornis dapat langsung meletakkan telur setelah keluar
dari puparium inang.
Menurut Rustam (2002), umumnya imago jantan muncul lebih awal
dibandingkan imago betina. Sebanyak 85.71% dari imago yang muncul pada hari
pertama adalah jantan. Lebih lanjut Rustam (2002) menjelaskan bahwa seekor
imago betina mempunyai keperidian berkisar antara 49-187 butir telur dengan
nisbah kelamin keturunan yang bias betina yakni sebesar 73.47 %.
Parameter Pertumbuhan Populasi
Neraca kehidupan (life table) merupakan teknik menghitung angka
kelahiran dan angka kematian suatu populasi. Dari data yang dihasilkan dapat
(1997), menyatakan bahwa dari angka kelahiran dan angka kematian dapat
disusun suatu neraca hidup yang menjelaskan perubahan–perubahan kuantitatif
dari suatu populasi selama satu generasi.
Ada dua tipe neraca kehidupan yaitu yang bersifat spesifik umur (age
spesific) atau tabel kehidupan horizontal, serta yang bersifat spesifik waktu (time
spesific) atau tabel kehidupan vertikal. Neraca kehidupan yang berspesifik waktu
menganalisis data yang diambil pada suatu kejadian tunggal, ketika diasumsikan
bahwa semua generasinya sudah saling lingkup dengan sempurna, oleh karena itu
kelas umur secara simultan sama. Neraca kehidupan yang bersifat spesifik umur
mencakup perhitungan yang berulang terhadap suatu kelompok (cohort) tunggal
yang terdiri dari individu yang sama umurnya sepanjang waktu. Tabel ini sering
digunakan dalam entomologi (Bellow & van Driesche 1992).
Menurut Poole (1974), perumusan neraca kehidupan merupakan langkah
pertama dalam menghitung laju pertumbuhan intrinsik (r). Dua data utama yang
dibutuhkan dalam perhitungan tersebut adalah lx, peluang bertahan pada umur x;
dan mx, rataan jumlah keturunan betina yang dihasilkan dalam satu unit waktu
oleh seekor betina berumur x (Birch 1948). Disamping nilai r, dari data tersebut
dapat juga dihitung laju reproduksi kotor (GRR), laju reproduksi bersih (Ro), dan
waktu generasi (T) (Birch 1948; Price 1997).
Neraca kehidupan merupakan riwayat perkembangan cohort yang bersifat
dinamis, mulai dari umur 0 sampai umur dimana semua individu dalam populasi
mati. Dari neraca kehidupan dapat diketahui laju reproduksi bersih (Ro), lama
waktu generasi (T) dan laju pertambahan instrinsik (r) (Tarumingkeng 1992).
Keefektifan musuh alami dalam mengendalikan hama sasaran biasanya
diukur berdasarkan beberapa ciri biologinya, di antaranya adalah (a) kemampuan
mencari inang yang tinggi, terutama pada saat kelimpahan inang rendah, (b)
kekhususan terhadap inang tertentu, (c) potensi reproduksi yang tinggi, (d) kisaran
toleransi terhadap lingkungan yang lebar serta kemampuan memarasit berbagai
instar inang (DeBach 1973).
Salah satu parameter yang paling handal untuk mengukur potensi musuh
alami adalah laju pertambahan intrinsik (r), karena telah mempertimbangkan
kelamin (Carey 1993). Nilai r merupakan salah satu kriteria yang penting untuk
mengevaluasi keefektifan atau potensi dari agens pengendalian hayati (Lee & Ahn
2000), serta dapat digunakan untuk menduga potensi pertumbuhan populasi
musuh alami (Lysyk 2000).
Respon Pemangsaan
Preferensi makanan, selera parasitoid terhadap inangnya, kerapatan inang,
kualitas makanan dan adanya makanan alternatif merupakan faktor lingkungan
yang berperan dalam menentukan laju pemangsaan oleh suatu predator terhadap
mangsanya atau parasitoid terhadap inangnya. Solomon (1949 dalam
Tarumingkeng 1992) mengemukakan teori mengenai respon pemangsaan terhadap
kerapatan inang dalam dua tipe, yaitu tanggap fungsional dan tanggap numerikal.
Tanggap fungsional merupakan salah satu ukuran untuk menentukan
keefektifan suatu predator atau parasitoid sebagai agens dalam pengendalian
hayati (Doutt 1973). Menurut Solomon (1949 dalam Hassel 2000) tanggap
fungsional merupakan perubahan jumlah mangsa yang diserang oleh individu
predator/parasitoid akibat perubahan kepadatan populasi mangsa per satuan
waktu.
Holling (1959 dalam Price 1997) mengkategorikan tanggap fungsional
menjadi tiga tipe: 1) Tipe respon fungsional linear (Tipe I), laju pemangsaan oleh
predator meningkat atau menurun sebanding dengan kerapatan mangsa dan
mencapai tahap kejenuhan. Tipe I ini biasanya ditemukan pada predator yang
bersifat pasif seperti laba-laba, 2) Tipe respon fungsional hiperbolik (Tipe II), laju
pemangsaan secara progresif semakin menurun. Respon ini sangat umum di antara
serangga-serangga predator dan parasitoid dan 3) Tipe respon fungsional sigmoid
(Tipe III), perubahan pemangsaan berlangsung lambat, diikuti dengan
peningkatan kemudian mendatar (asimptotik).
Menurut Murdoch dan Oaten (1975 dalam Wang dan Ferro 1998), pada
dasarnya tanggap fungsional merupakan komponen yang sangat essensial dari
dinamika interaksi antara parasitoid dengan inang dan juga sangat penting untuk
determinasi stabilitas dari sistem yang dikelola. Kajian mengenai tanggap
banyak dilakukan seperti yang diteliti oleh Wang dan Ferro (1998); Buchori
(2003); Hidrayani (2003).
Respon pemangsaan yang kedua adalah tanggap numerik, yaitu
pemangsaan menyebabkan perubahan kerapatan predator atau parasitoid pada
suatu luasan pemangsaan tertentu. Ada dua mekanisme yang mendorong
terjadinya tanggap numerik yaitu 1) peningkatan laju reproduksi predator atau
parasitoid bila mangsa berlimpah dan 2) keterpikatan predator pada tempat
begerombolnya mangsa, dikenal juga dengan tanggap agregasi.
Hubungan antara Inang, Parasitoid, dan Tanaman
Perilaku parasitoid terhadap inangnya tidak hanya dipengaruhi oleh
senyawa-senyawa kimia yang dihasilkan oleh serangga inang, tetapi juga oleh
makanan inang itu sendiri (Vinson 1984). Menurut Takabayashi et al. (1998),
parasitoid merupakan musuh alami yang agak khusus yang menyerang sejumlah
spesies herbivora tertentu. Kekhususan itu tidak hanya tergantung pada
herbivoranya, tetapi juga tanaman inangnya. Parasitoid mungkin hanya dapat
menemukan inang yang makan pada satu atau beberapa spesies tanaman saja,
sedangkan diketahui bahwa serangga tersebut mempunyai kisaran inang yang
luas.
Tingkat parasitisasi dan distribusi beberapa parasitoid pengorok daun tidak
merata pada berbagai tanaman inang, dan berbeda-beda menurut jenis tanaman
dan lokasi ditemukan. Selain itu parasitoid Liriomyza spp. dapat dikelompokkan
sesuai dengan spesies serangga inang, tanaman inang, dan daerah geografis
tempat tanaman inang tumbuh (Johnson & Hara 1987; Rauf et al. 2000).
Di Indonesia, parasitoid yang dominan memarasit L. huidobrensis adalah
H. varicornis dan O. chromatomyaie (Hidrayani 2003; Purnomo 2003), sedangkan
spesies parasitoid yang dominan memarasit L. sativae adalah Acecodes deluchii
(Rauf et al. 2000; Susilawati 2002; Tapahillah 2002). Di California, spesies
parasitoid yang dominan menyerang L. sativae adalah Chrysocharis parksi (
Zehnder & Trumble 1984).
Menurut Purnomo (2003), H. varicornis merupakan spesies parasitoid
yang dominan ditemukan pada tanaman kentang, sedangkan Tran dan Takagi
ditemukan pada tanaman bawang daun. Distribusi parasitoid Liriomyza spp. pada
berbagai tanaman inang tidak merata. Pengendalian biologi yang efektif
memerlukan adanya kesesuaian antara spesies parasitoid dengan serangga inang
dan tanaman inangnya.
Aplikasi Insektisda dalam Pengendalian Liriomyza
Penggunaan insektisida sintetik sangat beresiko karena dapat
menimbulkan dampak negatif, khususnya terhadap parasitoid sebagai serangga
musuh alami lalat pengorok daun. Menurut Purnomo (2003), jika insektisida
terpaksa digunakan, hendaknya insektisida tersebut bersifat selektif dan tidak
berdampak negatif bagi aktifitas musuh alami.
Penggunaan insektisida kimiawi seperti organoklorin, organofosfat, dan
karbamat yang berspektrum luas sudah sejak lama direkomendasikan sebagai cara
pengendalian hama, walaupun cara ini dinilai terlalu berbahaya. Penggunaan
insektisida telah banyak memacu ledakan hama sekunder, karena Liriomyza
menjadi resisten terhadap sejumlah insektisida yang digunakan. Oleh sebab itu
penggunaan insektisida harus atas hasil pengamatan agroekosistem untuk
mengetahui ambang ekonominya berdasarkan keadaan populasi lalat dan musuh
alaminya (Cardova & Karel 1990).
Penggunaan bioinsektisida merupakan salah satu alternatif yang tepat
untuk mengurangi dampak negatif yang ditimbulkan oleh insektisida sintetik.
Menurut Copping dan Menn (2000), istilah bioinsektisda mencakup berbagai
bahan pengendalian hama seperti: mikroorganisme (virus, bakteri, cendawan),
nematoda entomopatogen, insektisida botani, metabolit sekunder yang dihasilkan
mikroorganisme, dan feromon serangga. Penelitian mengenai pemanfaatan
bahan-bahan alami untuk perlindungan tanaman semakin gencar dilakukan selama
tiga dekade terakhir. Hingga saat ini terdapat tiga sumber insektisida alami yang
penting dan memiliki prospek yang baik untuk dikembangkan lebih lanjut, yaitu
tumbuhan, mikroorganisme tanah, dan organisme laut (Prijono 1999).
Insektisida berupa ekstrak daun dan biji nimba (Azadirachta indica A.
Juss) (Meliaceae), dengan bahan aktif azadiraktin dilaporkan sebagai insektisida
selektif, memiliki aktifitas biologi insektisida, mengurangi keperidian imago
perkembangan larva, mengurangi terbentuknya puparium, menyebabkan
pergantian kulit abnormal pada Musca autumnalis (Diptera: Muscidae)
(Schmutterer 1995).
Abamektin merupakan salah satu bioinsektisida yang berasal dari
mikroorgnisme yang merupakan hasil fermentasi dari bakteri Streptomyces
avermitilis. Abamektin telah memperlihatkan hasil yang sangat efektif dalam
pengendalian serangga hama (Pienkowski & Mehring 1983 dalam Leibee 1988).
Hasil penelitian yang dilakukan oleh Leibee (1988) memperlihatkan bahwa
penggunaan abamektin untuk mengendalikan L. trifolii dapat membunuh larva
pengorok sampai 99.5%. Sivapragasan dan Syed (1999) menginformasikan
bahwa abamektin merupakan salah satu bioinsektisida yang populer digunakan di
Malaysia dalam mengendalikan Liriomyza spp. Siromazin merupakan kelompok
zat pengatur tumbuh serangga yang efektif digunakan untuk mengendalikan
Liriomyza spp. yang telah terdaftar dan diizinkan oleh Menteri Pertanian dengan
1. LALAT PENGOROK DAUN DAN PARASITOIDNYA PADA
PERTANAMAN SAYURAN DATARAN TINGGI DI
KABUPATEN CIANJUR-BOGOR, JAWA BARAT
Abstrak
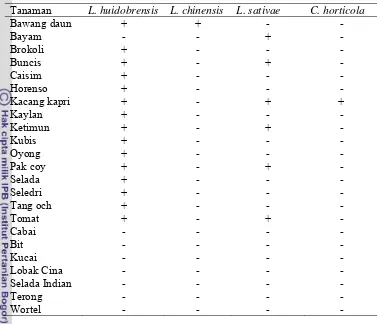
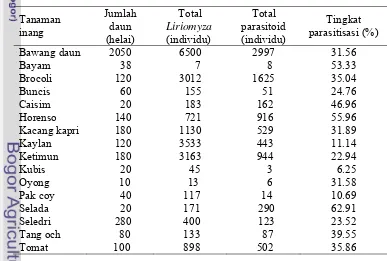
Lalat pengorok daun Liriomyza spp. adalah hama yang banyak menimbulkan kerusakan pada berbagai pertanaman sayuran dataran tinggi di Indonesia. Penelitian ini bertujuan untuk mengidentifikasi spesies Liriomyza yang menyerang pertanaman sayuran dataran tinggi, parasitoid yang berasosiasi dengan pengorok dan tingkat parasitisasi Opius chromatomyiae pada pertanaman sayuran dataran tinggi. Survei dilakukan dengan mengambil 20-40 helai daun tanaman sayuran yang mempelihatkan gejala korokan lalat pengorok daun. Imago lalat pengorok daun dan parasitoid yang muncul dari daun contoh diidentifikasi. Parasitisasi dihitung dengan membandingkan jumlah parasitoid dan imago lalat pengorok daun. Terdapat empat spesies lalat pengorok daun yang menginfestasi pertanaman sayuran dataran tinggi, yakni Liriomyza huidobrensis, L. chinensis, L. sativae, dan Chromatomyia horticola. Diketahui delapan spesies parasitoid yang berasosiasi dengan lalat pengorok daun, enam spesies dari famili Eulophidae, yaitu Hemiptarsenus varicornis, Asecodes deluchii, Neochrysocharis okazakii, N. formosa, Neochrysocharis sp., Quadrastichus liriomyzae dan masing-masing satu spesies dari famili Braconidae dan Eucoilidae yaitu Opius chromatomyiae dan
Gronotoma micromorpha. Parasitoid O. chromatomyiae merupakan parasitoid yang dominan pada skala ketinggian 1001-1300 m dan 1301-1600 m dari permukaan laut dengan tingkat parasitisasi berkisar antara1.84 – 62.26%. Dari 16 spesies tanaman yang diinfestasi oleh lalat pengorok daun, O. chromatomyiae
dapat memarasit lalat pengorok daun pada 13 tanaman.
Kata kunci: lalat pengorok daun, Liriomyza spp., parasitoid, sayuran
Abstract
Leafminer, Liriomyza spp. is the most important pest of highland vegetables in Indonesia. The objectives of the research were to (1) identify
Liriomyza species attacking higland vegetables and its parasitoids and, (2) investigate paratism rate of Opius chromatomyiae. Surveys on leafminer and its parasitoids were conducted by collecting 20-40 damage leaves. Adults of leafminer and its parasitoids were then identified. Parasitism was counted by dividing the number of parasitoid by total individuals emerging. As a result, there were four leafminers species were found associating with various kinds of vegetables. They were Liriomyza huidobrensis, L. chinensis, L. sativae, and
Chromatomyia horticola. Moreover, this study also revealed that there were eight species of parasitoid associated with the leafminers. Parasitoids Hemiptarsenus varicornis, Asecodes deluchii, Neochrysocharis okazakii, N. formosa,
Neocrhysocharis sp., and Quadrastichus liriomyzae belong to family of Eulophidae, Opius chromatomyiae belongs to familiy of Braconidae, and