KARAKTERXSASI
FENOTIPE
DAN GEMQTIPE
Streptococcus agaiactiae
[SOLAT ASAL INDONESlA
PENYEBAB
MASTITIS
SUBKLINIS PADA
SAP1
PERAH
QLEH
:
AGNESIA ENDANG TRI HASTUTI
WAHYUNI
PROGRAM
PASCASARJANA
AGMESW ENOANG TRI HASTUTt WAHYUHI. KamWsasi F e w dan Genotipe
a g a W h #
lsolat Asal Indomsh Pmy@bab MasW Subkiinis padaSapi Pmh. Dibimbing okh t WAYAM TEGUH WtBAWAN, MASDUKI
PARTADIREDJA (Aim), FACHRIYAM HASMI PASARIRU, BAM8ANG mPITJO
PRIOSOERYANTU dan WlDYA ASMARA.
Sm#mxas
a g a b w m p a k a n =I&satu
pmyelmb uEama masIi#ssubkiinis pada sapi parah
dm
menrpakm parasitMiart
pada ambing, Padamanu& bakM Eni mnpbablran infeksi mmatd seria pasta sari pa& wnita. Kadttwhsi S. a m M a e ini seafa
i c o n w ~ l
d m g ~
matodase-ng.
mdasawn atxls kekradaan antfgm tipe $ w a gemuka~in Sd b a r n *s.
agalwfhe dapat dibagi ka dahm Wmmpa serotip pitu 9antigen
polisdwida dan 3 mtQm protein. Wahupun metode irri swingdigu-n
namun s&duhp metodeini mash mempunyai
kekmahan
d a m mwnbedakm bakM secata M hdiskriminat#/ rinci, apakgi masih banyak S.
qp&&w
y ~ l n g Mum bisa dibngirern ke&!am
serrotipey m g
acla atau kdmpdr nontypabh (P17").W h
h m a iEu pmbkaim baru dmgan matihats e a m
gemtip Weri p&uh.
PenManini W j u m unktk rnqetahui pbedpsean
*ran
#rot@, huhngan fvtnotipedan
senrtfpasewta
mengaabhui e l DNA maupun kekembatan mtw Esolrd Sdmgmanfaat dari peditian ini dihsrapkan dapat mmberi masukm
d a m
pngendalianm a a s %Winis wrta menmican ciri W a r
png
dap& digunakan untukmmgetehui kemungkinan tajadinya
infeksi
a i m antam sapi dmmnusia
yang
hrguna untuit W i apiderniolqis cian zoonosis.PreridtbnifiW kWri ini dilakukan berclasafkm morfoiogi koloni dan
a n y a
%mama
faMw
Cirn'stieW n s
M w h &&mn yang dihasiikm. Smmtara p m t u a n gnrp baMeridilakukan dengan
uji imundikrsil AgarGel
PtWpitafion T& (AGPT) dmgm mmggmakan serum W t k & M a p gntp B.Eemrtifw
dad S.a g a k b e tiitentuh dengan meilhat pula prtumbuhm pada media
cair
T m Hewit Broth (THFI), medmagar
lmkl safiwar, panjang m idm
karaktw prmukaanltridmfobisbs kbwi thngan Safi &#gation Test. Setman serotipe
dad
S.W a W e ditantukan dengan uji serottpe mmggunakan anthwm spesifik brhatbp antgen fipe i d a t nrjukan dengan AEPT. Ganotipa
&ri
Wdwi dilahkan d-n metode rrrsrcrr, mstmfm * . !Bgm811t lempdynmphiism (MFLP) schizotypngdmgan p u W
fiaWg&
eMny,fiamsis
(PFGE)Had pen Jitian mnunjukkan ada prkiam distribusi s e r d p S. -kcthe dari
W r ,
Boptali
dan Makng.Kabanyakan
isalat
S.a
g
-
adalah
NT dsnanwen
protein X adaiah ~Totiptb y m g paling serirtg mum& S. a p myaw
turnbuh jemih pa&
media
cair, bersifat irarnpak p&da nwdi agar tunak, m p u n y a irantai yang panjmg dan twrsifat hidrafabiic. Sebaliknya S.
ag&&ae
ymgl t u Mkemh pada media
a i r ,
kdoni brsifat difus, berantai pen& dan bemifa4 hidtofilik. Ada hut#rngm antam femtlpe dmgm mmtipe S,sga-.
Harsil gmotipe dewanmtde
MFLP/ schhutyping dengan PFGE hmpak S. ag&&m menghasilhnm a n Deowbo N-ic A M (DNA) yang & i t dan dari 21 isoM
arfa
t5pub
profile DNA yam bMda dan ditamukan 3 isalat yang tidak dapat dipom d& endm S m l . Dari pnelitiatl inidapat
disimpulkan hhwa (1)ada
W a a n semtipentar wilayah,
(2)
actia
hubungan
a t a m fenutipe dm -tips#am
(3)a&
ABSTRACT
AGNES& WDANG TRI HASTUTI WAWUNI. Charactlerizatbn
of
Pheraotype andGenotype
of
StmpWwwsagak&ae
Isolated from tndonesia As Causetim Agent uf StMinic=nl Mastitis in DairyCam.
Under 4 k directionof
I WAYAN TEGUHWMWAN, MASDUKI PARTADIREDJA, FACHRIYAN MASMI PASARIBU,
BAMBANG PONTJQ PRIOSOERYANTO and WDYA ASMARA
Strreptocoocus aga- is
one
of
$M meina@mb
responsible POT subdinid mastitisin
dairy M eand
asan
oblig&e parasite in wddw.In
human, this tiacteriaare #e h d q j -use
of
human
pstpwhm ancl nmmW infeckhs.Charackwk&im
of
S.ag#I&b
trsuaiiydana
by
canvantiotral samtypng methods,Up
to
now,
b a aon
the occumceof
surface antigens. S. could bd e w in
to
9 potysaccharitfesnntigem
ptncj 3 protein antigens, Amugh thism e t ) r o d i s a f f e n ~ b u t ~ ~ t y t h e m e t h & M BackdasMddiydiscriminabry
~ ~ a n d t h e e ~ s m w s f ~ M ( N T ) , ~ a n e w q p r o a & t > y genotypbg Mbeused. T # w a i m o f t t r i s ~ ~ s t o t a P a w h o w t h e ~ b u i b n
uf
-type of S. qjaiadiae, ihe relatianstripof
plwnotyp~ mdmoty#m
and
4hap r a f i l e o f D N A a n d t h e g ~ ~ ~ ~ ~ ~ ~ * T I r e h m F i t a f t b s rewar& might give an infomatian in pravmtiva of the sut3dMml mastitis and
to
fmda ~ c m r l r ~ t h a t c a n k u s e d f o r ~ n o f a m s ~ n ~ h u m a n and m e .
F'1-alchtifhtion
of this bacteria was done basedon
lhe presentaf
Christie
Atkins Mum& Petmen piranarnm
and
c o b y morphoioglks. Mik determinationuf
group
8 was dons by usinggraup
8
s p W antisauum inimmunadifusion assay1 Agar Gel Precipitation Test (AGPn. The pbnotyplc of
isdata was determined
by
the growth patternof
S, agai&min
Todd Hwitk 8mth(THB) and soft-agar media, the chain langtfr
of
hctaria and ther
w
W
charactat/hydmpbbidty was determined by using Satt Aggregation Test
of
S. agaiadiae was donaby
using spesific antiserumagainst
S. a p k # a mfemnctastrains
in AGPT. The genotypicof
thebacteria
was d mby
macrr, m4rictianfragment bty#h polymorphism (MFLPY schi;k*ng mthad using PuWfiaid gel eiactrophoresis (PFGE)
.
Resub
of
the pmssnt study strowed $ha4there are deferences
disbibubnof
serotrpa uf S.
ag#I#cfiae
komm r ,
byolali ang Malmg.Mast
S. egaWa@isolates
are
nontypeabk strain(NT)
and lhetype
a&$pn X is the mostfrecluant
serotype. S. ipgalache mi grew as sediment wi4h char supernatant in fluid medium
(THB), formed compact cdwties in &-agar and
have
a lung &stin famation with ahydrophobic surface. In contrast,
bacteria
g m w
krrt>id in fluid medium, showeddiffuse colonies in
--agar
andhave
a sttortchain
fwmafim will haw a hirjmphilisurFace.
Them
is a relationship btween phenotype and semtyped
S.aga-.
Genotypic usingsdrixamng
meahads by PFGE with Smal restriction enzyme showd that S.agalacthe
pmdumd Deaxyrib Nudaic Acid (DNA) discrete ftagmants.
Thmare
3 isolatesof
S. aga-4
k
4
a n nut be W C f E K j by Snraienzyme.
Q m i c DNA analysis af 21isolates
sirowed 15 & i tDNA
pPafibs. The cclndusionof
ttrese
research are ( 3 )there
is a difbmntof
distibutian ofserotypes, (2) there is a relationship between phenotype and
smtype
and (3) tiweSUUAT PERMYATAAN
Dengan ini saya rnenyatakan bahwa disertasi berjudul :
"KARAKER15ASI FENOTfPE DAM
GENQTIPE
S-usag&bCtJBe
1-TASAL
lN00NESfA PEWEBAB MASTITlS SUBKtJNfSPADA
SAP#P€RAHm
adalah
b r t a r menrpakan h s i i kertya saya sendiri dan k i u m p m h dipubliksikan.Semw sumtwr data cSan infomasi yam tiiunakan b h h dinyatakan secara jdas
dan &pat
dipxiksa kebnarannya.Bogor, Agustus 20I32
Yang menyatakan
KAMKERISASI
FEWOTIPE
DAN
GENOIIPE
S-US
agalectiae
lSOlAT ASAL INDUNESIA
PENYEBAS
MASTlnS
SUSKLINIS
?ADA
SAP!
PERAH
PROGRAM PASCASARJANA
tNSTITUT PERTANIAM
BOGQR
Judu l Disertasi : Karakterisasi Eenotipe
dan
GenotipaStmptacomrs
agdiacfiae lsolat Asal Indonesia Penyebab Mastitis Subklinis pada Sapi Perah. Nama : Agnesia Endang Tri Hastuti WahyuniNRP : 985102
Program Studi : Sains Veteriner
Dr. Drh. I Wayan Twuh Wibawan, MS Ketua
Prof. Drh. Masduki Paftadiredja. MSc, Ph. D tAlrn) Dr. D&. Factlfivan Hasmi
Pasaribu
Anggota Anggata
X
-
O h . W#dya Asmara. SU., Ph.0
Angwb
2 .
Ketua
Program Studi Sains Veteriner ram PascasaFjanaPenulis dilahirkan di Karanganyar Solo pada tanggal 15 Agustus 1962 sebagai anak ketiga dari 6 bersaudara dari pasangan
SoekranBo
(Aim) dan Sami (Almh). Pendidikan Sa jana di tempuh di Fakultas KedoMeran Hewan UniversitasGadjah Mada Yogyakarta, lulus pada tahun 1986
dan
gelar DoMer Hewan diperoleh pada tahun 1988. Pada tahun 1995 penulis diterima di -ram Studi Sains Veteriner Program Pascasa jana IPB, dan gelar Magi* Sam diperdeh tahun 1998. Padatahun
yang sama penulis melanjutkan ke jenjang Program Doktor pada Program Studi yang sama dan pada perguruan tinggi yang sama pula. Beasiswa pendidikan pascasajana baik program Magister maupun program Doktor diperoleh dari Tim Manajemen Program Doktor Direktorat Jendral Pendiiikan TinggiDepartemen Pendidikan Nasional. Penulis bekeja sebagai Staf pengajar di
Fakultas Kedokteran Hewan Universitas Gadjah Mada di LabcmWium Mikrobiologi sejak Januari 1991, bidang penelian yang digeluti adalah bakteriologi. Sebelum menjadi staf pengajar, penulis pemah bekerja pada petemah sapi perah PT. Nandi Amerta Agung di Salatiga Jawa Tengah selama kurang lebih 2 tahun.
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala kasih dan kanmia-Nya penulis berhasil menyelesaikan karya ilmiah yang berjudul
Karakterisasi Fenotipe dan Genotipe Stmptococcus agalactiae lsolat Asal lndonesia
Penyebab Mastitis Subklinis pada Sapi Perah. Karya ilmiah ini merupakan hasil penelitian terhadap salah satu penyebab utama mastitis subklinis pada sapi perah, yang dilakukan sejak tahun 1999 dari 3 daerah yaitu Bogor, Boyolali dan Malang.
Pada kesempatan ini penulis mengucapkan terima kasih kepada Bapak Dr.
Drh. I Wayan Teguh Wibawan, MS; Prof. Drh. Masduki Partadiredja, MSc, Ph.D
(Alm); Dr. Drh. Fachriyan Hasmi Pasaribu; Drh. Bambang Pontjo Priosoeryanto, MS,
Ph.D dan Drh. Wdya Asmara, SU, Ph.0 selaku Tim Komisi Pembimbing. Kepada
Tim Manajemen Program Doktor Direktorat Jendral Pendidikan Tinggi Departemen Pendidikari Nasional penulis juga sampaikan terima kasih. Rasa terima kasih juga penulis sampaikan kepada Bapak Dr. Drh. I Wayan Teguh Wtbawan, MS atas bantuan sebagian dana penelitian dari Penelitian Hibah Bersaing VII Direbrat Jendral Pendidikan tinggi Departemen Pendidikan Nasional. Kepada lndonesia Toray Science Foundation (ITSF) 8 tahun 2001 yang telah rnemberikan dana sehingga penelitian ini be rjalan lancar. Ucapan terima kasih pula penulis sampaikan kepada Kepala Laboratorium Terpadu Fakultas Kedokteran Hewan lnstitut Pertanian Bogor yang telah mengijinkan dan memberi kesempatan sehingga penelitian ini dapat berjalan lancar. Tak lupa kepada Direktur SEAMEO BIOTROP yang telah memberi ijin untuk melaksanakan sebagian penelitian ini di Laboratorium
Bioteknologi dan kepada Bapak Dr. Ir. Antonius Suwanto, MSc yang telah memberi
kesempatan dan memberi masukan penulis ucapkan terima kasih. Terima kasih juga kepada Drh. Budi Tri Akoso, MSc, PhD., Direktur Kesehatan Hewan, Direktorat Jendral Bina Produksi Peternakan, Departemen Pertanian dan Dr.dr. Sri Budiarti Poetwanto, Staf Pengajar Jurusan Biologi FMIPA-IPB yang telah bersedia menjadi
Dosen Penguji Luar Komisi. Untuk rekan-rekan: Endang Endrakasih, Titiek
Djanatun, Mahdi Abrar, Sri Estuningsih, Eva Harlina, Fadrial Kamil, Zinatul Hayati atas saran, dorongan semangat dan kerja samanya selama ini. Untuk : Juwanto, Budi Pumomo, Fitria Khumiawati, Enysa, Diantyna, Endang Sri Pertiwi dan Ira Ramadhani yang telah membantu dalam pelaksanaan penelitian, juga terima kasih
untuk Bapak Drs. Agus Somantri yang te!ah membantu menyediakan media selama
penelitian ini, bapak Drs. Eddy Yusuf dari LIP1 Cibinong yang telah ikut memberi masukan dan bantuannya. Ungkapan terima kasih juga untuk almarhum ayah (Soekranto) dan almathumah ibu (Sami), kakak-kakak (Drs. Joko Supriyanto, MS; Ir. Edhy Sriyarmanto, k4Sc) dan adik-adik (Dra. Elizabeth Titik Kuspangestiningrum; Dra. Yasinta Wadhani Sutera Dewi; Dra. Anastasia Sutjijati Sulistijaningsih) serta keluarga besar bapak mertua Drs. Soetjipta, MSc. Untuk suami Drh. Setio Seputro Wdodo Triono, ke-dua buah hatiku Dhimas Husada dan Setia Bhagawanta, mbak Prantina atas segala bantuan, perhatian, pengertian, doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2002
DAFTAR
IS1
Halaman
...
DAFTAR TABEL vi
...
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN
...
viiiI
.
PENDAHULUAN1 1
. .
Latar Belakang...
11.2. Tujuan Penelitian
...
3 1.3. Hipotesis...
3II . TINJAUAN PUSTAKA
...
2.1
.
Streptococcus agalactiae2.1.1. Mikrobiologi S
.
agalactiae...
2.1.2. ldentifikasi S
.
agalactiae...
2.1.3. lnfeksi oleh S
.
agalacfiae pada Hewan dan Manusia...
2.2.Fenotipe
S.
agalactiae...
...
2.3. Sem?ipe S.
agalactiae...
2.3.1
.
Antigen Polisakarida...
2.3.2. Antigen Pmt3in
2.4. Faktor Virulen S
.
agahcfiae...
...
2.4.1. Faktor Virulen Struktural
2.4.2. Faktor Virulen Nonstnrktural
...
2.5. Mastitis Subklinis oleh S.
agalactiae...
...
2.5.1. Mastitis Subklinis
...
2.5.2. Deteksi Mastitis Subklinis
2.5.3. S . agalactiae Sebagai Penyebab Mastitis Subklinis
...
Ill
.
BAHAN DAN METODE3.1. Bahan
...
.
3.1 1. lsolat Bakteri 30
...
3.1.2. Hewan Percobaan 30
...
3.1.3. Bahan Kimia, Media dan Alat 30
Metode Penelitian
3.2.1. Penapisan Mastitis Subklinis
...
31 3.2.2. Preidentifikasi S.
agalactiae...
31...
3.2.2.1. Morfologi Koloni Streptokokus 31
3.2.2.2. Uji CAMP
...
31...
3.2.3. ldentifikasi S
.
agaladrae 32...
3.2.3.1. Penentuan Grup (Serogruping) 32
...
3.2.3.1 .a. Pembuatan Ekstrak Antigen untuk Serogruping 32
3.2.3.1
.
b. Uji lmunodifusi dengan Agar Gel Prwiphtion (AGP)33
...
3.2.4. Fenotipe S
.
agalactiae 34...
3.2.4.1. Pertumhhan pada Media Cair (THB) 34
3.2.4.2. Pertumbuhan pada Media Agar Lunak
...
34...
3.2.4.3. Panjang Rantai 35
3.2.4.4. Uji Hidrofobisitasl Karakter Permukaan
...
35 3.2.5. Penentuan Serotipe Secara Serologi I Semfyping...
35...
3.2.5.1. Pernbuatan Antigen Permukaan lsdat Rujukan 35
3.2.5.2. Pembuatan dan Preparasi Antiserum lsdat Rujukan
..
36
...
3.2.5.3. Uji Imunodiisi Antiserum yang Belum Diabsorbsi 37
...
3.2.5.4. Pembuatan Antiserum Monospesifik 38
3.2.5.5. Serotyping S
.
agaladiae lsolat Lapang dengan Antiserum Monospesifik...
38 3.2.6. Genotipe S.
agalactiae...
3.2.6.1. Penyiapan Suspensi Bakteri 39
...
3.2.6.2. Penyiapan DNA Genom Utuh dalam Blok Agarose 39
....
3.2.6.3. Pemotongan DNA Genom dengan Enzim Restriksi
40
3.2.6.4. Pemisahan Molekul DNA dengan PFGE
...
40...
3.2.6.5. Analisis Data 41
IV . HASlL DAN PEMBAHASAN
4.1. Penapisan Mastitis Subklinis dengan Menggunakan Pereaksi IPB-1
..
42...
4.2. Preidentifikasi S
.
agalactiae 434.3. ldentifikasi S
.
agaladiae...
45 4.4. Fenotipe S.
agalactiae pada Media Cair. Agar Lunak. PanjangRantai dan Hidrofobisitas Bakteri
...
50...
4.6 Genotipe S
.
a g a k h e dariBogor.
Boyolalidan Malang
denganPFGE
...
64V
.
KESIMPULAN DAN SARANDAFTAR
TABEL
Halaman
...
1 Jadwal penyuntikan dan pemanenan serum kelinci
2 Hasil penapisan sampel susu dari Bogor, Boyolali dan Malang terhadap mastitis subklinis dengan rnenggunakan pereaksi IPB-1
...
3 Hasil identifikasi S. agalactiae isolat dari Bogor, Boyolali cian Malang dengan serogruping menggunakan uji AGP
...
4 Hasil uji CAMP positif yang bukan S. agaladiae dari Bogor, Boyolali dan Malang
...
5 Hasil isolasi S. agaladiae dari sampel susu mastitis subklinis dariBogor, Boyolali dan Malang..
...
:.
...
i...
6 Fenotipe S. agaladiae dari Bogor, Boyolali dan Malang pa& berbagai
...
media
7a Data spesifisitas antiserum sebelum absorbsi antara antiserum isolat
...
rujukan dengan antigen permukaan isolat rujukan
7b Data spesifisitas antiserum sesudah absorbsi antar antiserum isolat rujukan dengan antigen permukaan isolat rujukan
...
8 Hasil uji sebaran semtipe S. agaladiae dari Bogor, Boyolali dan
...
Malang
9 Hubungan antara serotipe dan fenotipe S. agaladiae dari Bogor,
Boyolali dan Malang
...
10 P i DNA S. agakctiae dengan enzim resbiksi Smal dari Bogor,
DAFTAR GAMBAR
Halaman
1 Struktur natif antigen polisakarida tipe la dan Ib
...
13...
2 Stnrtur natif antigen polisakarida tipe 11 13
...
3 Struktur natif antigen polisakarida tipe 111 14
...
4 Struktur natif antigen polisakarida tipe IV
...
.
.
155 Struktur natii antigen polisakarida tipe V
...
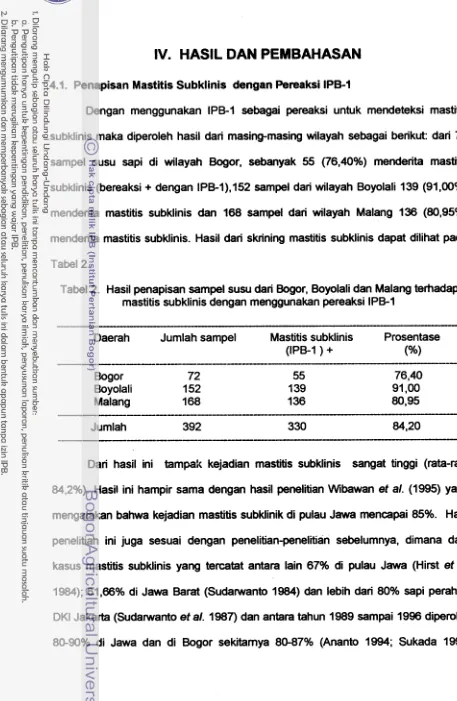
156 Morfologi koloni S
.
agalactiae yang diisolasi dari susu sapi perahmastitis subklinis pada media agar darah
...
447
Hasil uji CAMP antara P-hemolitik S . sums strain Pernth dengankoloni streptokokus
...
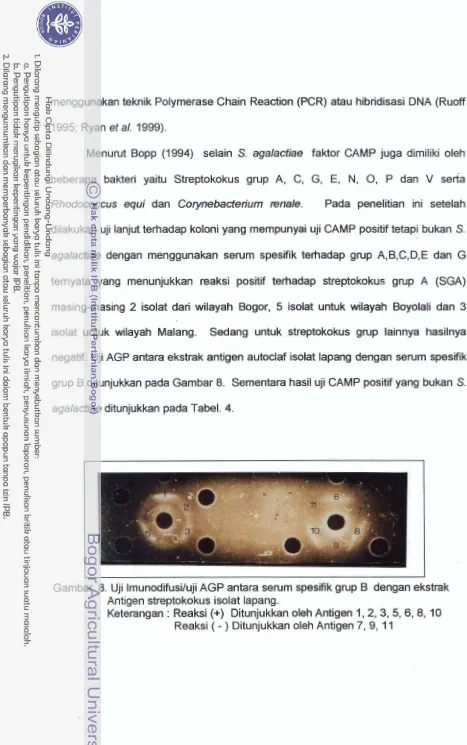
458 Uji imunodifusi/ uji AGP antara serum spesifik grup B dengan ekstrak
antigen stt-eptokokus isolat lapang
...
479 Fenotipe S
.
agalactiae pada media cair (THB)....
53...
10 Fenotipe S
.
agalactiae pada media agar lunak 5411 Morfologi bakteri S
.
agaladiae dengan pengecatan Gram...
5612 Hasil uji Imunodisi/ uji AGP antara serum monospesifik t e h d a p
antigen tipe dengan antigen perrnukaan S
.
agalactiae isolat lapang ... 6213 Pita DNA genom isolat S
.
agalactiae berasal dari Bogor. Boyolali dan...
Malang dengan enzim restriksi Smal dengan PFGE 65
14 Dendrogran yang menunjukkan hubungan kekerabatan antar isolat S
.
Htxh
man...
..
1
SW-sifat
dan hasit uji S.
aga- dari Bugor ,..,.,...
042 Siat-sifat dan hasil uji S
.
agabctipe4 dari Ek, yubli...
85
4 Data Bier berda-n ada-tidaknya pobngan DMA S
.
agalacfh
dariI.
PENDAHULUAN
1
.I.
Latar Belakang
Penyakit radang ambing atau yang dikenal sebagai mastitis merupakan masalah
utama
&lam dunia petemakan sapi perah, karena dapat menyebabkan kenrgian yang besar akibat adanya penunman produksi susu, penurunan kualitas susu, penyingkiran susu, biiya perawatan dan pengobatan yang tinggi sertapengafkiran temak lebih awal. Kejadian mastitis sekitar 97-98% merupakan rnastitis subklinis, sedangkan 23% merupakan hsus mastitis klinis yang terdeteksi (Sudarwanto 1999). Menurut Wibawan
et a/.
(1995) kejadian mastitis subklinis di Indonesia sangat tinggi (85%). Mastitis subklinis merupakan peradangan jaringan interna kelenjar ambing tanpa ditemukan adanya gejala klinis baik pa& susu maupun ambingnya, namun terjadi peningkatan jumlah sel radang, ditemukanmikroorganisme patogen dan te rjadi perubahan kirnia susu (Sudarwanto 1999).
Dari hasil penelitian Wibawan et al. (1995) terisolasi Stn?ptococcus agalactiae pada kejadian mastitis subklinis sebesar 83% di wilayah Bogor, 82% untuk wilayah Boyolali dan sebesar 80% untuk wilayah Malang. Sedang Benda et
al. (1997) mengatakan bahwa dua bakteri
patogen
penyebab mastitis yang sering ditemukan yaitu S. agalacfiae (92%) dan Staphfloaxcus aums (67%). S.agalactiae merupakan parasit oMigat pada ambmg sapi perah (Kennedy 1970). Selain menyebabkan mastitis subklinis, S. agalactiae atau dikenal sebagai
Streptokokus Grup B (SGB) pada kedokteran manusia dapat rnenyebabkan pneumonia, septisemia maupun meningitis pa& neonatal dan saluran kelamin wanita sebagai reservoimya (Baker 1980; Limansky et al. 1998). Angka prevalensi
dilahirkan oleh ibu dengan kolonisasi SGB akan mendapatkan kolonisasi yang sama
melalui transmisi vertikal yaitu melalui saluran kelamin ibu saat melahirkan (Edward
& Baker 1995).
Karakterisasi bakteri ini secara konvensional dilakukan dengan metode
serotyping. Berdasarkan spesifisitas antigen permukaan S. agalactiae ada beberapa
serotipe yaitu yang memiliki antigen polisakarida yang sampai saat ini ada 9 serotipe
yaitu la, Ib, 11, Ill, IV, V, VI, VII, Vlll dan yang memiliki antigen protein yaitu c, R, X
(Gravekam et a/. 1999; Bushman 1998; Kogan et all 1996). Menurut Wibawan &
Laemmler (1990a) isolat S. agalactiae dapat memiliki serotipe dengan antigen
polisakarida baik berdiri sendiri maupun dalam bentuk kombinasi dengan antigen
protein, misalnya ldc, IIIX. Namun demikian ada isolat yang belum bisa
diklasifikasikan kedalam serotipe yang ada dan disebut sebagai nonjrpeable (NT).
Hasil sebaran serotipe ini penting untuk diketahui karena dapat dipakai untuk
menduga faktor viwlen yang paling sering munculldominan dalam proses infeksi.
Disamping itu juga merupakan inforrnasi dasar dalam mempelajari epidemiologi
penyakit maupun untuk menemukan cara pengendalian penyakit tersebut. Meski
metode ini sering dipakai, namun sebetulnya mempunyai kelemahan dalam
membedakan baktefi secara lebih diskriminatiflrinci, apalagi masih banyak isolat S.
agalactiae dari sapi yang termasuk kelompok NT (Blumberg et a/. 1992; Fasola et a/.
1993; Limansky et a/. 1998). Akhir-akhir ini pendekatan secara molekuler bertujuan
untuk mengetahui perbedaan genom diantara organisme yang sangat dekat telah
digunakan untuk analisa S. agalactiae yaitu dengan Restriction Enzyme Fragment
Length Polymorphism (RLFP), -typing, Multi locus enzyme electrophoresis dan
1992; Denning et a/. 1989; Fasola et a/. 1993; Bentley & Leigh 1995; Gordillo et a/. 1 993; Quentin et a/. 1995)
Meski kejadian mastitis subklinis pada sapi perah yang disebabkan oleh S.
agalactiae di Indonesia tinggi, namun penelitian mengenai S. agalactiae sebagai
penyebab masih sedikit dilakukan. Bahkan sampai saat ini belum atau masih sedikit
diketahui bagaimana sebaran serotipe, ekspresi fenotipe apalagi profil DNA dari bakteri tersebut. Sampai saat ini faktor virulen S. agalactiae yang paling dominan
berperan dalam penampilan mastitis subklinis masih sangat terbatas. Kajian
epidemiologi dengan mempehitungkan sebaran serotipe belum banyak dilakukan.
Oleh karena itu penelitian tentang karakterisasi fenotipe dan genotipe S. agalactiae
isolat asal Indonesia sebagai penyebab mastitis subklinis pada sapi perah perlu
dilakukan.
1.2.
Tujuan Penelitian
Tujuan dari penelitian ini adalah : (1) melihat sebaran serotipe S. agalacfiae
dari Bogor, Boyolali dan Malang, (2) melihat hubungan antara fenotipe dengan
serotipe dari isolat yang diperoleh (3) melihat profil DNA dan kekerabatan isolat dari masing-masing serotipe maupun masing-masing daerah.
1.3.
Hipotesis
:1. Ada pola sebaran serotipe S. agalactiae yang berbeda-beda antar daerah
2. Ada hubungan antara fenotipe dengan serotipe
3. Ada perbedaan profil DNA dan kekerabatan dari masing-masing serotipe dan
II.
TINJAUAN PUSTAKA
2.1. Streptococcus agalactiae 2.1 .I. Mikrobiologi S. agalactiae
Streptokokus berasal dari bahasa Yunani yaitu
Sfepbs
yang berarti flexibel atau mudah disesuaikan dan coccus yang artinya bijilbibi. Sehingga menurut Fosteret a/. (1957) bisa diirtikan seperti untaian biji yang panjang menympai bentuk kalung. Sedang kata agalacfiae berarti ingin air susu yang menunjukkan habitat dari
bakteri tersebut (Sneath et al. 1986). Bakteri ini pertama kali diisolasi oleh Nocard dan Mollereau pada tahun 1887 dari kasus mastitis. Oleh karena itu mikroorganisme ini diberi nama S. nocadi (Jelinkova 1977).
S. agalacfiae adalah bakteri gram positif, tidak bergerak, tidak berspora, berbentuk bulat/kokus, tersusun &lam rantai bervariasi antara 20 sampai 40 sel &n dapat membentuk kapsul (mikrokapsul). Bakteri ini dikelompokkan dalam streptokokus yang bersifat pyogen yang beftanggung jawab temadap infeksi
bernanah, bebntuk bulat atau ovoid dengan diameter 0,6
-
1,2 pm dapat dikenal dari sifat-sifat biokimia yang dimilikinya (Rotta 1986; Joklik 1992; Gotoff 1992; Edwards & Baker 1995). Berdasarkan hernolise yang dihasilkan pada agar darah domba 5%, bakteri tersebut rnemiliki hemolise alfa, beta dan garna. Hemolise alfa ditunjukkan dengan adanya zona kehijauan disekitar koloni, zona terang dan luas disekitar koloni apabila bakteri mempunyai hemolise betadan
tidak terbentuknyazona disekitar koloni menunjukkan sifat hemolisis gama (Kelser & Scoening 1948).
Pembagian kelompok Streptokokus pertama kali oleh Sherman (1937) menjadi : (1) Streptokokus Piogenik, biasanya P-hemolisis, tidak tahan panas dan
yang
kuat Tennasuk diilamnya ~ t o c o t m s agalactiae dan Stleptouxws pyugenes. (2) Streptokokus Viridans, tumbuh pada suhu 45% tidak tumbuh pada suhu 1 0 ' ~ ~ termasuk spesies Streptococcus themrophilus, Stteptoaxxus bovis danStreptococcus mutans. (3) Streptokokus laktik, tumbuh pada suhu 1 0 ' ~ tidak tumbuh pada 4 5 ' ~ ~ spesies yang termasuk antara lain Stteptomccus lactis dan
Streptococcus cmmoris. (4) Enterokoki, yang memiliki kemampuan hemolisis bewariasi, tumbuh pada suhu antara 10
-
45%. Beberapa spesies yang termasuk di dalamnya adalah Stmptococcus bcalis, Streptococlcus lquifaciens, Stmptococcuszyogenes dan Streptococcus dumns.
Lancefield pada tahun 1933 mengamati suatu komponen polisakarida (substansi C) yang terdapat pada dinding sel bakteri streptokokus. Reaksi dengan
serum yang mengandung antibodi spesifik terhadap bakteri tersebut menyebabkan presipitasi, dari sinilah streptokokus dibedakan berdasarkan spesifikasi grup dengan
A, B, C, dan sebagainya (Wilson & Miles 1975). Sampai saat ini telah d i u k a n 20
grup streptokokus yang terdiri dari grup A sampai V kecuali grup I dan J.
Streptocaxs agalactiae tennasuk grup B, memiliki komposisi karbohidrat g w p spesifik tertentu yang terdiri dari D-glukosarnin, D-galaktosa, glusitol dan L-
rhamnosa (Joklik 1992; Edwards & Baker 1995j
2.1.2. ldentifikasi S. agalactiae
Untuk identitifikasi S. agalactiae dilakukan terlebih dahulu preidentifikasi
bakteri dengan melihat morfologi koloni dan uji Christie Atkins Muend, Petersen
(CAMP). Morfologi koloni dari streptokokus dilihat pada pertumbuhan di media agar darah domba 5%. Koloni akan berbentuk bulat kecil, dengan permukaan sedikit
hemolik 11439% strain. Namun demikian 5.15% adabh non hemolitik, sedangkan
untuk a-hemolitik jarang ditemukan (Joklik 1992; Edwards & Baker 1995).
Preidentifikasi dengan uji CAMP didasarkan atas kejasama yang sinergis
antara Staphy/ococcus aumus p-hemolitik strain Pertsch dengan S. agalactiae pada
agar darah sehingga membentuk zona kepala panah (amhead) atau setengah
bulan dengan lisis sempuma pada P-hemolitik (Brown et a/. 1981). Menurut Lay &
Hastowo (1992) S. aureus P-hemolitik mempunyai sifat hemolisis tidak sempuma
(partial hamolytic) akan berubah menjadi hemolisis sempuma sehingga terlihat zona
yang lebih terang di sekitar pertumbuhan S. agalactiae. Semua strain S. agalactiae
(Lancefield grup B) membentuk bahan-bahan yang dapat berdifusi untuk melengkapi
lisisnya eritrosit domba oleh hemolisin-P S. aumus sehingga memberikan reaksi
CAMP positif (Christie et a/. 1944). Hal yang sama telah dilakukan oleh W~bawan et
a/. (1995) dan Wahyuni (1998) dengan menggunakan S. sums (3-hemolitik strain
Pertsh.
ldentifikasi definitif untuk S. agalactiae (serogmuping), dapat dideteksi
berdasarkan jsnis kahhidrat yang terdapat pada dinding sel spesifik grup yang
selanjutnya dikenal dengan ca&ohydmte specific group. Pada S. agalactiae,
sejumlah metode diagnosis baik untuk menentukan serogrup maupun serotipe yang
dapat digunakan antara lain: imunodifusi, countecumnt immunoeledmphoresis,
enzyme-linked immunoso&ent assay (ELISA), imunofloresen tidak langsung,
koaglutinasi dengan stafilokokus dan aglutinasi dengan lateks (Towers et at. 1990;
Wibawan & Pasaribu 1993; Edwards & Baker 1995; Ruoff 1995). Salah satu
metode yang sering dipakai adalah metode imunodifusi (Ouchterlony). Metode ini
pembacaannya yang relatif lama (18-24 jam) dan memerlukan bahan yang relatif
mahal (VVibawan & Pasaribu 1993).
2.1.3. lnfeksi oleh S. agalacfiae pada Hewan dan Manusia
Selain menyebabkan mastitis subklinis pada sapi perah, bakteri ini juga
dilaporkan menyerang kerbau, domba, kambing dan babi. Pada bidang veteriner
selain menyebabkan mastitis, bakteri ini dapat pula menyebabkan abortus dan
cervicitis pada kuda dan babi , abses pada gajah, anjing, merpati, cicak, kadal dan
ikan (Pasaribu et al. 1993; Butter & Moor 1967). lnfeksi bakteri S. agalactiae
pada babi dan dari nutria (hewan pengerat), temyata hanya memperlihatkan sedikit
gejala klinis (Wibawan et al, 1993a). lnfeksi oleh bakteri ini dilaporkan dapat
menyebabkan kematian neonatal pada anjing, namun tingkat morbiditasnya lebih
rendah dibandingkan pada bayi manusia meski secara klinis ada kemiripan
(Komblatt et al. 1983). Sedang menurut Spitznagel et al. (1983) bakteri ini
menyebabkan arthritis pada tikus setelah 6-8 hari, tidak ada lesi atau luka pada kulit,
..
ekor maupun telinga.
Selain menyebabkan infeksi pada hewan, S. agalactiae atau yang lebih
dikenal sebagai Streptokokus Grup B (SGB) pada manusia temyata dapat
menyebabkan meningitis dan septisemia pada neonatal (Baker 1980; Limansky et al.
1998). Bakteri ini pertama kali dilaporkan sebagai patogen pada manusia yang
menyebabkan infeksi pascasalin pada tahun 1935. Manifestasi klinis infeksi
pasmsalin yang disebabkan oleh SGB ini antara lain endometritis, endomiometritis
ataupun endomioparametritis. Pasien-pasien yang menjalani seksiosesaria
oleh bakteri ini (aWmnri 1998). Nampaknya kasus pada manusia meningkat terns
setiap tahunnya demikian juga pada hewan (Schwartz et a/. 1994).
Manifestasi klinis infeksi neonatal dibedakan menjadi dua bentuk yaitu infeksi
dengan onset dini (early onset) dan infeksi dengan onset lambat (late onset). lnfeksi
early onset dapat tejadi ketika bayi masih dalam uterus (Katz 1993) atau dalam
minggu pertama kelahiran (biasanya dalam 20 jam setelah lahir) (Edwards & Baker
1995). Manifestasi yang paling dominan adalah pneumonia (Tamura et a/; 1994).
Pada keadaan yang lebih berat dapat terjadi sepsis yang disertai kegagalan
pemafasan. Pneumonia ini terjadi karena adanya apirasi cairan amnion yang
terinfeksi dalam uterus atau melalui vagina pada saat dilahirkan (Rubens 1992).
Sedang infeksi late onset biasanya tejadi minggu pertama kelahiran hingga
mencapai usia 3 minggu, dengan infeksi yang paling dominan adalah meningitis
dengan mortalitas 1530% (Katz 1993).
2.2. Fenotipe S. agalactiae
Patogenesis streptokokus berhubungan dengan fenotipe pada permukaan
set bakteri, kemampuan opsonik dan sifat protektif dari antigen polisakarida maupun
antigen proteinnya (Kling 1997). Fenotipe bakteri S. agalactiae sangat berhubungan
dengan keberadaan antigen permukaan. Pada umumnya bakteri yang memiliki
antigen protein X dan R dan bakteri yang belum dapat digolongkan pada salah satu
tipe yang ada (NT) menunjukkan pertumbuhan yang jemih pada supematan dengan
endapan pada media cair dan membentuk koloni kompak di media agar lunak (soft-
agarj, serta membentuk rantai yang panjang dan bersifat hidrofobik. Sebaliknya
bakteri yang hanya memiliki antigen polisakarida atau dalam bentuk gabungan
cair, koloni difus pada media agar lunak serta membentuk rantai yang pendek
(Wibawan et a/. 1996).
Selain antigen protein, fenotipe bakteri ditentukan juga oleh adanya kapsul.
Asam sialat adalah merupakan komponen utama penyusun kapsuV polisakarida
bakteri tersebut bahkan keberadaan asam sialat akan menentukan pola
pertumbuhan pada media cair, agar lunak, panjang rantai dan hidrofobisitas bakteri
serta efek hambat dalam proses fagositosis (Wibawan & Laemmler 1991 a). Residu
asam sialat terdapat dalam kapsul dari semua tipe ini dan terdapat dalam jumlah
yang besar pada tipe la, II dan Ill (Wibawan & Laemmler & Pasaribu 1992). Lokasi
residu asam sialat pada tipe II adalah unik dibanding yang lain karena asam sialat
tersebut tidak sensiti terhadap neuraminidase karena terikat langsung pada tulang
punggungnya (Jennings et a/. 1983). Kapsul polisakarida dinyatakan sebagai faktor
yang bertanggung jamb terhadap sifat virulensi mikroorganisme tersebut, dan
ketebalan dari kapsul menrpakan faktor utama (Baker & Kasper 1976). Secara
fungsional asam sialat diketahui dapat mencegah aktivasi lain dari komplemen.
Peran asam sialat sebagai faktor vinrlen bisa secara langsung meningkatkan
pabgenesitas dari bakteri tersebut atau juga bisa beriindak sebagai antiagositosis
(Shigeoka et a/. 1983). Sifat fisikokimia kapsul mencerminkan sifat fisikokimia
karbohidrat non stnrktural seperti menyerupai lendir (amorf), bermuatan negatif dan
sangat hidrofilik (Styrer 1988). Selain itu komponen karbohidrat kapsul seperti asam
sialat pada SGB (Shigeoka et a/. 1983, Wibawan & Laemmler 1991b) dan asam
hyaluronat pada Streptokokus gnrp C (SGC) (Durack 1989) umumnya dimiliki oleh
1982) ditambah dengan kemiripan struktur kapsul dengan komponen jaringan tubuh,
sehingga sifat non antigeniknya juga muncul secara in vivo (Wessels et a/. 1989).
Hidrofobisitas merupakan suatu sifat yang diekspresikan oleh suatu bakteri
berdasar atas komponen yang terdapat pada perrnukaannya. Menurut Pasaribu et
al. (1994) S. agalactiae yang memiliki antigen protein X dan c secara umum bersifat
hidrofobik dan yang hanya memiliki antigen polisakarida saja umumnya cenderung
bersifat hidrofilik. Pada S. agalactiae sifat hidmfobisitas berkaitan dengan morfologi
koloni pada agar lunak. Bakteri dengan derajat hidmfobik yang tinggi memiliki koloni
berbentuk kompak pada agar lunak dan permukaan koloni relatif kasar jika
ditumbuhkan pada media agar darah. Sebaliknya bakteri yang bersifat hidmfilik
memiliki koloni diffus pada agar lunak dan permukaan mukoid pada agar darah
(Kane et a/. 1975; W~bawan & Laemmler 1990b). Bakteri ini umumnya tersusun
dengan rantai pendek (diplokoki), namun panjang rantainya dapat dipengaruhi oleh
faktor lingkungan. Pada medium cair biasanya tumbuh dengan rantai yang lebih
panjang (Joklik et al. 1992). Menurut W~bawan et al. (1995), panjang rantai
dipengaruhi juga olen sifat hidrofobisitas dari komponen permukaan bakteri. Bakteri
yartg bersifat hidrofilik tersusun dengan rantai pendek, sedang bakteri yang bersifat
hidrofobik tersusun oleh rantai panjang. S. agalactiae yang diisolasi dari sapi perah
penderita mastitis subklinis biasanya mempunyai rantai yang panjang, sedangkan
SGB dari isolat manusia biasanya mempunyai rantai pendek.
Selain perbedaan ekspresi fenotipe S. agalactiae antara sapi dan manusia
diatas, perlu diketahui juga perbedaan sifat biokimiawi antara keduanya. Hal ini
penting untuk diketahui karena seperti diketahui bahwa di Indonesia pemerahan
sering berhubungan dengan sapinya dan keadaan ini memungkinkan te rjadinya
infeksi silang antara sapi dan manusia. Perbedaan sifat biokimiawi antara S.
agalacfiae dari manusia dan sapi adalah isolat asal sapi cenderung tidak mampu
membentuk pigmen apabila dibiakkan pada media lslam agar (Islam 1977), sedang
isolat asal manusia sebagian besar mampu membentuk pigmen. S. agalactiae asal
sapi sensitif terhadap basitrasin sedang SGB isolat manusia resisten terhadap
basitrasin (10 U) dan sebagian besar S. agalactiae asal sapi mampu memecah
laktosa sedang SGB asal manusia sebasian besar tidak mampu memecah laktosa.
Dari hasil penelitian Pasaribu, W~bawan dan Warsa (1993) dikatakan 98% S.
agalactiae isolat asal sapi mampu memecah laktosa sedang 89% SGB isolat asal
manusia tidak memecah laktosa. Namun dari hasil penelitian W~bawan et a/.
(1993a) isolat S. agalactiae dari babi dan nutrias memperlihatkan sifat menghidrolisa
Natrium hipurat dan salisin, maltose dan sakarose tetapi tidak dengan esculin,
manitol atau inulin. Sebanyak 53% S. agalactiae dari babi memfermentasi laktosa
sedang tidak ada yang memfermentasi laktosa pada nutrias. Sebagian besar bakteri
sensiti terhadap penisilin dan basitrasin 10 U, namun sebagian besar isolat dari babi
resisten terhadap tetrasiklin. Ditambahkan juga bahwa S. agalactiae dari babi maupun nutrias tidak berperan dalam infeksi silang antara hewan maupun antsra
manusia dan hewan.
2.3. Serotipe S. agalactiae
Berdasarkan atas spesifisitas antigen perrnukaan S. agalactiae dapat dibagi
dalam beberapa serotipe yaitu yang memiliki antigen polisakarida yang sejauh ini
ada sembilan serotipe yaitu la. Ib, 11, Ill, IV, V, VI, VII, Vlll dan yang memiliki antigen
al. 1996). Penyebaran serotipe diantara strain dari S. agalactiae berbeda di
beberapa negara, tergantung dari mana isolat tersebut didapat dan waktu
pengambilannya (Anthony & Okada 1977),
2.3.1. Antigen Polisakarida
Komposisi antigen polisakarida tip-spesifik terletak pada kandungan
galaktosa, glukosa, N-asetil glukosamin dan asam N-asetilneuraminik (sialic acid :
asam sialat). Perbedaan untuk masing-masing serotipe terletak pada rantai tulang
punggung dan ikatan rantai antar cabang gugus polisakarida serta ketebalan
kandungan asam sialat (Anthony 1992; Joklik 1992; Tissi 1998).
Tipe la dan Ib memiliki kesamaan struktur tulang punggung dan rantai
samping namun mempunyai perbedaan hanya pada ikatan cabang galaktosa ke
gugus glukosamin. Antigen polisakarida ini terdiri dari galaktose, glukose, N-
asetilglukosamin dan asam sialat dengan perbandingan 2 : 1 : 1 : 1 (Cumming et a/.
1981). Antigen polisakarida la mengandung jumlah asam sialat yang lebih banyak
dalam unit struktur kimianya yang diketahui dengan metode DEAE-Sephacel
Chromatography (Decueninck et a/. 1983). Tipe antigen Ib dengan menggunakan
Crossed ImmunoeIectmpho~sis (CIE) dari trichloroacetic acid (TCA)-ekstrak, selain
komponen polisakarida yang sama ditemukan pula rhamnose, dengan penambahan
jumlah molar pada glukosamin dan rhamnose (Cumming et a/. 1981). Struktur dari
Gambar 1. S t ~ k t ~ r nati antigen polisakarida la (A) dan Ib (B)( Wibawan 1993). Keterangan : asam sialat : a-D-NeupNac; Galaktose : p-D-Galp
N-Asetil-Glukosamine : p-D-GlepNac ; Glukosa : p-D-Glep
Tipe II mempunyai dua monosakarida rantai samping yaitu galaktosa dan
asam sialat yang secara langsung memanjang dari pengulangan tulang
punggungnya (Champbel et a/. 1992). Menurut Jenning et a/. (1983) antigen
polisakarida nati yang diisolasi dari tipe II S. agalactiae mengandung D-galaktosa,
D-glukcse, 2-acetamido-2deoxy-D-glukose dan asam sialat dengan ratio molar 3 : 2 : 1 : 1. Clntuk lebih jelasnya komposisi natif antigen polisakarida II ini dapat dilihat
pada Gambar 2.
[image:138.612.53.521.33.798.2]Antigen polisakarida tipe Ill terdiri dari suatu polimer yang mengandung
pengulangan unit-unit tulang punggung trisakarida glukosa
-
N-asetilglukosamin-
galaktosa dan rantai samping asam sialat
-
galaktosa yang terikat ke gugus N-asetilglukosamin (Wessel 1998). Menurut Baker
et
a/. (1976) antigen tipe Ill ini terdiri dariasam sialat (24%), galaktose (25%), heptose (21%). glukose (13%), glukosamin
(1090) dan manose
(7%),
yang diperoleh dari ekstraksi dengan menggunakan bakteriutuh yang diwci dengan larutan netral atau penyangga. Struktur natif antigen
polisakarida tipe Ill dapat dilihat pada Gambar 3.
Gambar 3. Struktur natif antigen polisakarida tipe Ill (Wibawan 1993).
Antigen polisakarida tipe IV mempunyai susunan kimia yang terdiri dari
galaktose, glukose, N-asetiglukosamin dan asam sialat dengan ratio molar 2 : 2 :
1
:1. Sedangkan untuk antigen polisakarida tipe V memiliki komposisi kim~a terdiri dari
glukose, galaktose, 2-acetoamido-2-deoxyglukosa dan asam sialat dengan ratio
molar 3 : 2 : 1 : 1 (Wibawan 1993). Gambar 4. dan 5. menunjukan Struktur natif
[image:139.612.48.516.34.781.2]Gambar 4. Struktur natif antigen polisakarida tipe IV ( Wibawan 1993).
Gambar 5. Struktur natif antigen polisakarida tipe V ( Wibawan 1993).
mempunyai kandungan asam sialat yang tinggi diketahui dapat menghambat jalur
komplemen altematif, sehingga lebih tahan terhadap fagositosis (Anthony 1992).
Sifat fisikokimia kapsul mencerminkan sifat karbohidrat nonstrukural seperti lendir,
bemuatan negatip dan sangat hidrofilik. Selain itu komponen karbohidrat kapsul
seperti asam sialat pada SGB (Wibawan & Laemmler 1991a) dan asam hialuranat
pada SGC (Durack 1989) umumnya dimiliki oleh jaringan tubuh.
2.3.2. Antigen protein
Selain antigen polisakarida, S. agalactiae memiliki antigen protein yang terdiri
dari antigen protein c, R dan protein X. Antigen protein c sebelumnya dikenal
sebagai antigen protein Ibc (Wilkinson & Moody 1969 diacu dalam Wibawan et a/.
1993a). Perubahan penamaan protein antigen Ibc menjadi antigen c diusulkan oleh
Henrichsen et a/. (1984). Antigen protein c merupakan bagian dari antigen
polisakarida Ib dan antigen protein c sendiri (Wibawan & Laemmler 1990b). Hal
senada juga terjadi pada tipe antigen polisakarida Ib, yaitu merupakan antigen
protein c dan antigen polisakarida Ib sendiri. Antigen protein c dibagi menjadi ca
yang merupakan komponpn resisten tripsin dan protease dan tidak mengikat
reseptor imunoglobulin A dan cfl yang sensiti tripsin, protease dan mengikat Fc dari
reseptor imunoglobulin A msnusia ( Michel et al. 1991), bahkan sekarang telah
diremukan c~ dan c6 namun belum dikarakterisasi sifat biokimianya (Wibawan et a/.
1991c; Maeland 1997; Brady et al. 1988). Protein ini telah diekstraksi dari
permukaan sel bakteri dengan pemanasan HCI (Bevanger & lversen 1981),
deterjen non ionik (Russel-Jones et a/, 1984) dan mutanolisin (Madoff et a/. 1991).
sering ditemukan dengan antigen polisakarida la, Ib, II namun jarang dengan Ill
(Bevanger 1983; Johnson & Ferrieri 1984).
Antigen protein R dibagi R1 hingga R4, antigen ini biasanya ditemukan dari
isolat manusia dan jarang ditemukan pada- isolat asal sapi dan mempunyai berat
molekul 116 kDa. Antigen ini terutama R4 dan R1 biasanya bersama dengan
antigen polisakarida tipe II dan Ill dan sedikit dengan polisakarida yang lain (Flores &
Fenieri 1985). Antigen protein ini pertama kali dijelas~an pada streptokokus grup A
tipe 28 (Lancefield et a/. 1952) dan diberi nama R untuk sifat resisten terhadap
tripsin. Antigen protein R ini juga ditemukan pada streptokokus grup C, F, G, dan L
(Lancefiel et a/. 1957, diacu dalam W~bawan & Laemmler 1991c).
Antigen protein X biasanya ditemukan pada isolat dari sapi dan tidak
ditemukan pada isolat asal manusia, mempunyai berat molekul 180 kDa dan
bersifat imunogenik w ~ b a w a n et a/. 1996 & Rainard 1991). Antigen X ini merupakan
antigen permukaan yang bersifat protein labil terhadap tripsin (Jelinkova 1977).
Antigen permukaan ini identik dengan streptokokus patogen lainnya yaihr grup G
dan L (Laemmler et a/. 1987). lsolat S. agalactiae dapat memiliki serotipe dengan
antigen polisakarida baik berdiri sendiri maupun bentuk kombinasi dengan antigen
protein , misalnya la/c, IIIX (Wibawan & Laemmler , 1990a).
Dari hasil penelitian sebelumnya diketahui bahwa S. agalactiae yang diisolasi
dari sapi dan manusia masing-masing memiliki pebedaan yang khusus serta
mempunyai pola penyebaran tipe antigen yang berbeda. Hal ini penting untuk
mempelajari kemungkinan tejadinya infeksi silang dari manusia ke hewan atau
sebaliknya. lsolat S. agalactiae asal sapi tipe antigen yang sering ditemukan adalah
dilaporkan terjadi pada isolat asal manusia sehingga antigen protein X ini dikatakan
sebagai ciri khas markerhallmark isolat S. agalactiae asal sapi (Laemmler et a/.
1993). Pada isolat SGB asal manusia yang sering ditemukan adalah antigen la/c, II,
Ill baik berdiri sendiri maupun kombinasi dengan antigen protein R. Namun menurut
W~bawan et a/. (1995) temyata masih banyak S. agalactiae di Indonesia yang belum
dapat diklasifikasikan pada serotipe yang ada dan disebut sebagai nontypeable
(NT). Strain S. agalactiae pada babi banyak memiliki serotipe Ill dan banyak isolat
yang berasal dari nutrias, memiliki serotipe laic, dimana protein c terdapat dalam
bentuk
cp
2.4. Faktor Virulen S.
agalacfiae
Salah satu faktor penentu keberhasilan infeksi suatu bakteri adalah adanya
faktor virulen yang dimiliki oleh bakteri tersebut. Sifat faktor virulen ini sangat penting
untuk dikaji guna diketahui perannnya dalam mekanisme infeksi. Faktor virulensi S.
agalactiae dapat dibagi menjadi faktor virulensi struktural dan non strukturall
komponen ekstraseluler.
2.4.1. Faktor Virulen Struktural
Faktor virulensi struktural dibentuk oleh komponen-komponen penyusun sel
baik komponen perrnukaan maupun komponen penyusun dinding sel bakteri. Faktor
virulensi tersebut antara lain adalah antigen polisakarida, antigen protein dan asam
lipoteikoat serta antigen protein lainnya (Wibawan & Lemmler 1991 b; Anthony 1992).
Antigen polisakariddkapsul umumnya terdiri dari asam hialuronat, sedangkan
dinding sel terdiri dari peptidoglikan. Kebanyakan kapsul berupa
asam uronat dan gula asam organik seperti asam tartrat dan asam asetat
(Kleinsmith et
a/.
1995).Antigen protein merupakan faktor virulensi yang imunogenik (Gravekamp et
a/. 1997; Rainard 1991), sedang antigen polisakarida sebagai pembentuk kapsul
bakteri merupakan faktor virulen yang tidak imunogenik (Kling et a/. 1997) dan
berperan sebagai anti fagositosis (Wibawan & Laemmlor 1991b). Pada S. agalactiae
diketahui bahwa kapsul berperan dalam menekan aktivitas komplemen, sehingga
proses eliminasi bakteri oleh makrofag dan PMN terhambat (Salasia et. a/. 1994).
Antigen polisakarida dapat berdiri sendiri namun dapat juga bergabung dengan
antigen protein (Wibawan & Laemmler 1992; Laemmler 1995; Gravekamp et a/.
1997) . Peran tipe antigen sebagai faktor virulen yang berpengaruh pada ekspresi fenotipe termasuk karakter permukaan ditentukan oleh keberadaan tipe antigen dan
kapsul di permukaan bakteri. Kedua ha1 ini mendasari patogenesitas bakteri ini
pada kasus mastitis dan kejadian sepsis maupun meningitis (Baker 1980). Protein
permukaan akan ikut menentukan sifat antigenik bakteri tergantung dari komposisi
asam amino yang menyusunnya. Oleh karena itu peran komponen-komponen
protein permukaan maupun antigen polisakarida penting untuk dipelajari sebagai
faktor virulen untuk memilih kandidat vaksin.
Kapsul merupakan lapisan teriuar bakteri yang memilikinya, sedang kapsul
pada S. agalactiae tersusun oleh asam sialat dan senyawa karbohidrat lainnya yang
membentuk struktur oligosakarida spesifik untuk tiap serotipe (Utarna et a/. 1997).
Antigen polisakarida sebagian besar tersusun oleh asam sialat, bahkan asam sialat
merupakan komponen utama penyusun kapsul bakteri tersebut (Wibawan et a/.
Komponen antigen permukaan lain yang dimiliki oleh bakteri S. agalactiae
adalah hemaglutinin, adhesin dan protein hidrofobik wbawan et al. 1993b; Kurl et
a/. 1989). Adhesi adalah menempelnya bakteri pada permukaan mukosa. Proses
adhesi dapat dibedakan dalam dua bentuk yaitu bersifat irreversible dan spesifik
serta ~versible dan non spesifik. Penempelan ini diperantarai oleh adhesin sebagai
komplek polimer permukaan bakteri yang mengandung protein, polisakarida, asam
sialat atau konjugatnya (Roth 1988). Adhesin adalah struktur dari bakteri yang
memperantarai adhesi, sedangkan adhesin yang menghemaglutinasi eritrosit adalah
hemaglutinin (Isaacon 1985).
Pada S. agalactiae hemaglutinin berperan dalam adhesi pada sel epitel
ambing sapi perah. Hal ini telah dibuktikan oleh Wahyuni (1998) yang melihat peran
hemaglutinin tersebut dengan menggunakan kerokan sel epitel ambing sapi perah.
Hemaglutinin merupakan faktor virulen struktural yang dimiliki oleh S. agalactiae asal
sapi. Hemaglutinin ini mampu mengaglutinasi beberapa eritrosit hewan tetapi tidak
dengan eritrosit manusia. Antibodi spesifik hemaglutinin mampu menghambat
adhesi S. agalacfiae pada sel epitel ambing. Hal yang sama juga dilakukan oleh
Wibawan et a/. (1993b) dengan menggunakan sel-sel heLa. Adhesi oleh
hemaglutinin ini diduga bersifat spesifik dan imversible . Menurut Ruup et a1 .
(1995) perlekatan awal bakteri pada suatu permukaan dianggap menjadi sebuah
proses remrsible (dapat balik) yang dapat dipengaruhi oleh faktor-faktor non
spesifik. Sedang menurut Allison dan Sutherland (1987) diacu dalam Gilbert (1991)
proses ini diikuti dengan suatu ikatan yang irreversible dan spesifik baik melalui
perlekatan permukaan yang spesifik atau memproduksi polimer ekstraseluler.
memfasilitasi dan mengakibatkan adhesi. Reseptor hemaglutinin pada sel epitel
ambing sapi perah ini telah diisolasi dan dikarakterisasi oleh Estuningsih (1998) dan
temyata mampu menghambat aktivitas adhesi.
Pada S. agalactiae selain protein hemaglutinin, protein hidrofobik berperan
juga dalam mekanisme adhesi ini. Hal ini telah dibuktikan oleh Wahyuni 1998 dan
Wibawan et a/. (1992) yang telah menunjukkan bahwa S. agalactiae yang
mempunyai protein hidrofobik mempunyal kemampuan menempel jauh lebih besar
dibandingkan ysng tidak. Adhesi dari S. agalactiae pada sel epitel buccal sapi
dihambat oleh adanya protein hidrofobik yang berhasil diisolasi maupun dari
antibodi spesifik terhadap protein hidrofobik tersebut.
2.4.2. Faktor Virulen Nonstruktural
Faktor virulensi nonstruktural (metabolit) yang merupakan produk
ekstraseluler dari bakteri yang memiliki komponen antigenik berupa enzim atau
protein yang dilepaskan ke dalam medium perkembangbiakkannya dan berguna
dalam membantu proses infeksi. Untuk S. agalactiae produk ekstra sel dari bakteri
ini antara lain hemolisin, faktor CAMP, hipurikase, nuklease, protease,
neuraminidase, hiilorunidase dan C5a-ase (Takashi et a/. 1999; Timoney 1993;
Gotoff 1992). Faktor virulensi ini umumnya berperan dalam proses invasi.
Hyaluronidase dikenal sebagai spreading factor yang berperan untuk merusak
struktur jaringan. Menurut Davies et a/. (1973) hyaluronidase bersifat antigenik yang
2.5. Mastitis Subklinis oleh S. agalactiae
2.5.1. Mastitis Subklinis
Mastitis adalah peradangan pada jaringan ambing yang disertai perubahan
fisik, kimia, mikrobiologik dan adanya kenaikan jumlah sel radang terutama leukosit
dalam susu dan dapat disertai dengan perubahan patologi pada jaringan ambing
(Faull, 1987 diaw dalam Ananto, 1995). Mastitis menurut gejalanya dibagi mastitis
subklinis dan mastitis klinis. Mastitis klinis senantiasa diikuti oleh tandc-tanda klinis
baik berupa pembengkakan , pengerasan ambing, rasa sakit, panas, kemerahan dan
penurunan fungsi ambing (Sudawanto 1993). Sedang mastitis subklinis adalah
peradangan jaringan intema kelenjar ambing tanpa ditemukan adanya gejala klinis
baik pada susu maupun pada ambingnya, namun tejadi peningkatan jumlah sel
radang, ditemukan mikroorganisme patogen dan tejadi perubahan kimia susu
(Sudanvanto 1999). Pada umumnya mastitis subklinis akan berlanjut menjadi
mastitis kronis yang kadang-kadang didahului oleh munculnya mastitis akut ataupun
sub-akut dan mastitis seperti inilah biasanya sering menimbulkan terbentuknya
jaringan ikat pada ambing sapi.
Kejadian mastitis sekitar 97-98% merupakan mastitis subklinis sedang 2-3%
merupakan kasus masititis klinik yang terdeteksi, sehingga kasus mastitis itu seperti
'fenomena gunung es'. Dan hasil penelitian Wibawan et a/. (1995) kejadian rnastitis
subklinik di pulau Jawa mencapai 85%. Jumlah kasus mastitis di Indonesia terus
meningkat dari tahun ke tahun. Data kasus mastitis subklinis yang tercatat antara
lain