1
KARAKTERISTIK ENZIM SERUPA TRIPSIN
DARI CACING TANAH
RUDY WIJAYA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
Rudy Wijaya. F251030071. Karakteristik Enzim Serupa Tripsin dari Cacing Tanah. Dibawah Bimbingan : Prof. Dr. Ir. Maggy Thenawijaya Suhartono dan Dr. Ir. Dahrul Syah, MSc.
Kasiat cacing tanah dalam pengobatan telah lama diketahui, dimana salah satu komponen aktifnya adalah enzim protease serin serupa tripsin. Enzim ini diketahui mampu untuk mendegradasi fibrin/fibrinogen dalam trombus secara langsung sehingga diharapkan dapat digunakan dalam pengobatan penyakit-penyakit penyumbatan pembuluh darah.
Cacing tanah yang digunakan dalam penelitian ini berasal dari Serpong, Tangerang, Indonesia. Produksi tepung cacing dilakukan dengan menggunakan oven vakum pada tekanan 300 psi, dengan suhu 50oC selama 48 jam. Rendemen tepung cacing tanah yang diperoleh setelah disaring dengan saringan 900 mesh adalah 9,61% (berat basah) untuk bagian halus sedangkan bagian yang kasar sebesar 1,55% (berat basah).
Enzim protease cacing tanah dimurnikan dengan menggunakan teknik kromatografi filtrasi gel dan kromatografi afinitas. Enzim kasar cacing tanah memiliki aktivitas optimum pada pH 8,0 di dalam 0,05 M bufer fosfat dengan suhu 60oC. Adanya ion-ion Na, Co, Cu, Ba, K, Li, Mn, Mg, Ca, Zn dan Fe yaitu dalam bentuk garam dengan ion klorida pada konsentrasi masing-masing sebesar 5 dan 1 mM, deterjen (Tween 20, SDS dan Triton X-100 pada konsentrasi 5% dan 1% b/v), urea (5% dan 1% b/v) dan 2-merkaptoetanol (1% dan 0,5% b/v) serta EDTA (5 dan 1 mM) tidak menunjukkan pengaruh terhadap aktivitas enzim, sebaliknya senyawa PMSF dengan konsentrasi 5 mM sudah mampu menghambat 78% aktivitas enzimatik dari enzim kasar cacing tanah. Enzim kasar cacing tanah dengan aktivitas spesifik 0,28 U/mg juga mampu untuk mendigesti fibrinogen, kasein, fibrin dan gelatin dengan konsentrasi masing-masing substrat sebesar 2% b/v.
Pemurnian enzim dengan matriks Sephadex G-100 dan dilanjutkan dengan kolom Hitrap Benzamidine FF berhasil memurnikan dua fraksi enzim, yaitu fraksi nomor delapan menunjukkan adanya lima fraksi enzim dan fraksi nomor 24 dengan satu fraksi aktif enzim. Eluat Sephadex G-100 mempunyai aktivitas spesifik sebesar 1,37 U/mg pada substrat kasein, sedangkan fraksi nomor delapan dan nomor 24 dari kolom Hitrap Benzamidine FF masing-masing mempunyai aktivitas spesifik sebesar 1,92 U/mg dan 111,36 U/mg.
Hasil analisis zimogram menunjukkan bahwa berat molekul dari fraksi enzim murni nomor delapan adalah 20, 22, 27, 40 dan 44 kD. Sedangkan fraksi enzim murni nomor 24 menunjukkan berat molekul 20 kD yang menyerupai berat molekul enzim tripsin. Pengujian stabilitas pH enzim murni pada substrat fibrin dan fibrinogen menunjukkan bahwa enzim ini stabil pada kisaran pH 2 hingga 12.
3 PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul : “Karakteristik Enzim Serupa Tripsin Dari Cacing Tanah”, adalah benar-benar merupakan karya saya sendiri dengan bimbingan dari komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Semua sumber informasi dan data yang berasal atau yang dikutip dari karya yang telah diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dapat diperiksa kebenarannya.
Bogor, Desember 2005
KARAKTERISTIK ENZIM SERUPA TRIPSIN
DARI CACING TANAH
RUDY WIJAYA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
5 Judul Tesis : Karakteristik Enzim Serupa Tripsin dari Cacing Tanah Nama Mahasiswa : Rudy Wijaya
Nomor Pokok : F 251 03 0071/ILMU PANGAN
Menyetujui, 1. Komisi Pembimbing
Prof. Dr. Ir. Maggy T. Suhartono Dr. Ir. Dahrul Syah, MSc Ketua Anggota
2. Ketua Program Studi Ilmu Pangan 3. Dekan Pascasarjana
Prof. Dr. Ir. Betty Sri Laksmi Jenie, MS Prof. Dr. Ir. Sjafrida Manuwoto, MSc
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 30 Oktober 1975, sebagai anak kesembilan dari sepuluh bersaudara dari pasangan Pohandi Wijaya dan Meimunah.
Penulis menyelesaikan pendidikan Strata 1 dari Jurusan Teknologi Pangan dan Gizi, Fakultas Teknologi Pertanian IPB pada tahun 1998. Pada tahun 2003 penulis melanjutkan studi Strata 2 pada Program Studi Ilmu Pangan, Sekolah Pascasarjana IPB.
7
Hak cipta milik Rudy Wijaya, tahun 2005
Hak cipta dilindungi
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah Bapa atas segala anugrah dan kasih setia-Nya sehingga penulis dapat menyelesaikan pendidikan S2 di Sekolah Pascasarjana IPB.
Tesis ini disusun berdasarkan penelitian yang dilakukan penulis di Laboratorium Mikrobiologi dan Biokimia, Pusat Penelitian Bioteknologi IPB. Pada kesempatan ini penulis menyampaikan rasa terima kasih dan penghargaan yang sebesar-besarnya kepada :
1. Prof. Dr. Ir. Maggy Thenawijaya Suhartono selaku dosen pembimbing I yang telah memberikan bimbingan, kepercayaan dan fasilitas yang sangat membantu penulis untuk menyelesaikan penelitian.
2. Dr. Ir. Dahrul Syah, MSc. selaku dosen pembimbing II yang telah bersedia meluangkan waktu untuk membimbing penulis dan atas kepercayaannya sehingga penulis dapat melanjutkan studi S2.
3. Dr. Ir. Ratih Dewanti Hariyadi, MSc. selaku penguji luar Komisi Pembimbing atas saran dan bantuannya dalam penulisan tesis.
4. Ibu Tami Idiyanti, MSc. atas bantuan dan waktu yang telah diberikan kepada penulis selama pembuatan tepung cacing.
5. Siti, Darta, Bobi, Agnes, Ibu Eni, Ibu Yuyun, Ibu Tati dan teman-teman seperjuangan di Lab. MB.
6. Ir. Ika Malikha atas segala bantuan, kepercayaan dan keceriaan selama penulis melakukan penelitian.
7. Ir. Yanti, MSi. atas segala bantuan, diskusi dan saran-sarannya selama penulis melakukan penelitian
8. Tino, sumber motivasi, inspirasi dan pendorong penulis ketika menghadapi berbagai masalah selama menempuh studi S2.
9. Rekan-rekan seperjuangan di Lab. Biorin, Mashuda dan Firdaus, terima kasih atas dorongan yang diberikan.
9 12. Serta kepada segenap teman dan kerabat yang tidak dapat penulis sebutkan satu
persatu, terima kasih dan penghargaan yang sebesar-besarnya atas perhatian dan bantuannya selama ini.
Penulis menyadari bahwa tesis ini masih terdapat banyak kekurangan yang memerlukan penyempurnaan di masa datang. Penulis sangat menghargai atas kritik dan saran terhadap tesis ini, yang tentunya diharapkan dapat memberikan manfaat yang sebesar-besarnya bagi semua pihak yang memerlukan.
Bogor, Desember 2005
DAFTAR ISI
Halaman
DAFTAR ISI ... viii
DAFTAR GAMBAR ... x
DAFTAR TABEL ... xii
DAFTAR LAMPIRAN ... xiii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 2
TINJAUAN PUSTAKA ... 3
Cacing Tanah (Lumbricus rubellus) ... 3
Protease Cacing Tanah ... 5
Potensi Lumbrokinase sebagai Enzim Trombolitik ... 10
Pemurnian Enzim ... 13
Ekstraksi Enzim ... 13
Pemekatan Enzim ... 14
Fraksinasi Enzim ... 14
Elektroforesis Gel ... 16
SDS PAGE ... 16
Zimografi ... 16
BAHAN DAN METODE ... 18
Bahan dan Alat ... 18
Metode Penelitian ... 18
Pembuatan Tepung Cacing ... 18
Pemurnian Enzim Serupa Tripsin dari CacingTanah ... 18
Pemurnian Enzim Serupa Tripsin Menggunakan Matriks Sephadex G-100 ... 19
Pemurnian Enzim Serupa Tripsin Menggunakan Kolom Hitrap Benzamidine FF ... 19
1
KARAKTERISTIK ENZIM SERUPA TRIPSIN
DARI CACING TANAH
RUDY WIJAYA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
ABSTRAK
Rudy Wijaya. F251030071. Karakteristik Enzim Serupa Tripsin dari Cacing Tanah. Dibawah Bimbingan : Prof. Dr. Ir. Maggy Thenawijaya Suhartono dan Dr. Ir. Dahrul Syah, MSc.
Kasiat cacing tanah dalam pengobatan telah lama diketahui, dimana salah satu komponen aktifnya adalah enzim protease serin serupa tripsin. Enzim ini diketahui mampu untuk mendegradasi fibrin/fibrinogen dalam trombus secara langsung sehingga diharapkan dapat digunakan dalam pengobatan penyakit-penyakit penyumbatan pembuluh darah.
Cacing tanah yang digunakan dalam penelitian ini berasal dari Serpong, Tangerang, Indonesia. Produksi tepung cacing dilakukan dengan menggunakan oven vakum pada tekanan 300 psi, dengan suhu 50oC selama 48 jam. Rendemen tepung cacing tanah yang diperoleh setelah disaring dengan saringan 900 mesh adalah 9,61% (berat basah) untuk bagian halus sedangkan bagian yang kasar sebesar 1,55% (berat basah).
Enzim protease cacing tanah dimurnikan dengan menggunakan teknik kromatografi filtrasi gel dan kromatografi afinitas. Enzim kasar cacing tanah memiliki aktivitas optimum pada pH 8,0 di dalam 0,05 M bufer fosfat dengan suhu 60oC. Adanya ion-ion Na, Co, Cu, Ba, K, Li, Mn, Mg, Ca, Zn dan Fe yaitu dalam bentuk garam dengan ion klorida pada konsentrasi masing-masing sebesar 5 dan 1 mM, deterjen (Tween 20, SDS dan Triton X-100 pada konsentrasi 5% dan 1% b/v), urea (5% dan 1% b/v) dan 2-merkaptoetanol (1% dan 0,5% b/v) serta EDTA (5 dan 1 mM) tidak menunjukkan pengaruh terhadap aktivitas enzim, sebaliknya senyawa PMSF dengan konsentrasi 5 mM sudah mampu menghambat 78% aktivitas enzimatik dari enzim kasar cacing tanah. Enzim kasar cacing tanah dengan aktivitas spesifik 0,28 U/mg juga mampu untuk mendigesti fibrinogen, kasein, fibrin dan gelatin dengan konsentrasi masing-masing substrat sebesar 2% b/v.
Pemurnian enzim dengan matriks Sephadex G-100 dan dilanjutkan dengan kolom Hitrap Benzamidine FF berhasil memurnikan dua fraksi enzim, yaitu fraksi nomor delapan menunjukkan adanya lima fraksi enzim dan fraksi nomor 24 dengan satu fraksi aktif enzim. Eluat Sephadex G-100 mempunyai aktivitas spesifik sebesar 1,37 U/mg pada substrat kasein, sedangkan fraksi nomor delapan dan nomor 24 dari kolom Hitrap Benzamidine FF masing-masing mempunyai aktivitas spesifik sebesar 1,92 U/mg dan 111,36 U/mg.
Hasil analisis zimogram menunjukkan bahwa berat molekul dari fraksi enzim murni nomor delapan adalah 20, 22, 27, 40 dan 44 kD. Sedangkan fraksi enzim murni nomor 24 menunjukkan berat molekul 20 kD yang menyerupai berat molekul enzim tripsin. Pengujian stabilitas pH enzim murni pada substrat fibrin dan fibrinogen menunjukkan bahwa enzim ini stabil pada kisaran pH 2 hingga 12.
3 PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul : “Karakteristik Enzim Serupa Tripsin Dari Cacing Tanah”, adalah benar-benar merupakan karya saya sendiri dengan bimbingan dari komisi pembimbing dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Semua sumber informasi dan data yang berasal atau yang dikutip dari karya yang telah diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dapat diperiksa kebenarannya.
Bogor, Desember 2005
KARAKTERISTIK ENZIM SERUPA TRIPSIN
DARI CACING TANAH
RUDY WIJAYA
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Pangan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
5 Judul Tesis : Karakteristik Enzim Serupa Tripsin dari Cacing Tanah Nama Mahasiswa : Rudy Wijaya
Nomor Pokok : F 251 03 0071/ILMU PANGAN
Menyetujui, 1. Komisi Pembimbing
Prof. Dr. Ir. Maggy T. Suhartono Dr. Ir. Dahrul Syah, MSc Ketua Anggota
2. Ketua Program Studi Ilmu Pangan 3. Dekan Pascasarjana
Prof. Dr. Ir. Betty Sri Laksmi Jenie, MS Prof. Dr. Ir. Sjafrida Manuwoto, MSc
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 30 Oktober 1975, sebagai anak kesembilan dari sepuluh bersaudara dari pasangan Pohandi Wijaya dan Meimunah.
Penulis menyelesaikan pendidikan Strata 1 dari Jurusan Teknologi Pangan dan Gizi, Fakultas Teknologi Pertanian IPB pada tahun 1998. Pada tahun 2003 penulis melanjutkan studi Strata 2 pada Program Studi Ilmu Pangan, Sekolah Pascasarjana IPB.
7
Hak cipta milik Rudy Wijaya, tahun 2005
Hak cipta dilindungi
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Allah Bapa atas segala anugrah dan kasih setia-Nya sehingga penulis dapat menyelesaikan pendidikan S2 di Sekolah Pascasarjana IPB.
Tesis ini disusun berdasarkan penelitian yang dilakukan penulis di Laboratorium Mikrobiologi dan Biokimia, Pusat Penelitian Bioteknologi IPB. Pada kesempatan ini penulis menyampaikan rasa terima kasih dan penghargaan yang sebesar-besarnya kepada :
1. Prof. Dr. Ir. Maggy Thenawijaya Suhartono selaku dosen pembimbing I yang telah memberikan bimbingan, kepercayaan dan fasilitas yang sangat membantu penulis untuk menyelesaikan penelitian.
2. Dr. Ir. Dahrul Syah, MSc. selaku dosen pembimbing II yang telah bersedia meluangkan waktu untuk membimbing penulis dan atas kepercayaannya sehingga penulis dapat melanjutkan studi S2.
3. Dr. Ir. Ratih Dewanti Hariyadi, MSc. selaku penguji luar Komisi Pembimbing atas saran dan bantuannya dalam penulisan tesis.
4. Ibu Tami Idiyanti, MSc. atas bantuan dan waktu yang telah diberikan kepada penulis selama pembuatan tepung cacing.
5. Siti, Darta, Bobi, Agnes, Ibu Eni, Ibu Yuyun, Ibu Tati dan teman-teman seperjuangan di Lab. MB.
6. Ir. Ika Malikha atas segala bantuan, kepercayaan dan keceriaan selama penulis melakukan penelitian.
7. Ir. Yanti, MSi. atas segala bantuan, diskusi dan saran-sarannya selama penulis melakukan penelitian
8. Tino, sumber motivasi, inspirasi dan pendorong penulis ketika menghadapi berbagai masalah selama menempuh studi S2.
9. Rekan-rekan seperjuangan di Lab. Biorin, Mashuda dan Firdaus, terima kasih atas dorongan yang diberikan.
9 12. Serta kepada segenap teman dan kerabat yang tidak dapat penulis sebutkan satu
persatu, terima kasih dan penghargaan yang sebesar-besarnya atas perhatian dan bantuannya selama ini.
Penulis menyadari bahwa tesis ini masih terdapat banyak kekurangan yang memerlukan penyempurnaan di masa datang. Penulis sangat menghargai atas kritik dan saran terhadap tesis ini, yang tentunya diharapkan dapat memberikan manfaat yang sebesar-besarnya bagi semua pihak yang memerlukan.
Bogor, Desember 2005
DAFTAR ISI
Halaman
DAFTAR ISI ... viii
DAFTAR GAMBAR ... x
DAFTAR TABEL ... xii
DAFTAR LAMPIRAN ... xiii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 2
TINJAUAN PUSTAKA ... 3
Cacing Tanah (Lumbricus rubellus) ... 3
Protease Cacing Tanah ... 5
Potensi Lumbrokinase sebagai Enzim Trombolitik ... 10
Pemurnian Enzim ... 13
Ekstraksi Enzim ... 13
Pemekatan Enzim ... 14
Fraksinasi Enzim ... 14
Elektroforesis Gel ... 16
SDS PAGE ... 16
Zimografi ... 16
BAHAN DAN METODE ... 18
Bahan dan Alat ... 18
Metode Penelitian ... 18
Pembuatan Tepung Cacing ... 18
Pemurnian Enzim Serupa Tripsin dari CacingTanah ... 18
Pemurnian Enzim Serupa Tripsin Menggunakan Matriks Sephadex G-100 ... 19
Pemurnian Enzim Serupa Tripsin Menggunakan Kolom Hitrap Benzamidine FF ... 19
11
Analisis SDS PAGE dan Zimografi ... 22
Penentuan Suhu Optimum ... 22
Penentuan pH Optimum dan Kestabilan pH ... 22
Pengaruh Deterjen, Inhibitor dan Protein Denaturan ... 22
Pengaruh Ion Logam ... 22
Spesifitas Substrat ... 23
HASIL DAN PEMBAHASAN ... 24
Penentuan Rendemen dan Total Protein Cacing Tanah ... 24
Karakterisasi Protease Kasar Cacing Tanah ... 24
Pemurnian Enzim ... 34
Spesifitas Substrat ... 39
Stabilitas pH ... 40
SIMPULAN DAN SARAN ... 42
DAFTAR PUSTAKA ... 43
DAFTAR GAMBAR
Halaman Gambar 1. Perbandingan mekanisme reaksi enzimatis protease
(Creighton 1997) ... 7 Gambar 2. Mekanisme kerja lumbrokinase sebagai obat trombolitik
(Yanti 2003) ... 12 Gambar 3. Ilustrasi beberapa metode fraksinasi enzim ... 15 Gambar 4. Pengaruh SDS terhadap protein ... 16 Gambar 5. Suhu optimum aktivitas enzim kasar cacing tanah ... 25 Gambar 6. Struktur tiga dimensi enzim fibrinolitik komponen A dari
E. fetida (Tang et al. 2002) ... 26 Gambar 7. Pengaruh pH dan larutan bufer terhadap enzim kasar
cacing tanah ... 27 Gambar 8. Pengaruh ion logam terhadap aktivitas enzim kasar ... 28 Gambar 9. Struktur dua dimensi PMSF ... 29 Gambar 10. Ikatan disulfida pada komponen A enzim fibrinolitik
E. fetida ... 30 Gambar 11. Spesifitas substrat dari enzim kasar cacing tanah ... 32 Gambar 12. Metode skrining awal aktivitas fibrinolitik dari ekstrak enzim 32 Gambar 13. Analisis zimogram kasein (A) (10%) dan SDS PAGE
(B) (10%) dari ekstrak kasar cacing tanah ... 33 Gambar 14. Pemurnian enzim protease cacing tanah dengan teknik
kromatografi filtrasi gel ... 35 Gambar 15. Matriks kolom Hitrap Benzamidin FF ... 36 Gambar 16. Interaksi enzim protease serin dengan inhibitor (Creighton
1997) ... 36 Gambar 17. Pemurnian enzim protease cacing tanah dengan teknik
kromatografi afinitas ... 37 Gambar 18. Analisis SDS PAGE (10%) (A) dan zimogram fibrin
13 Gambar 19. Analisis zimogram spesifitas enzim terhadap substrat
kasein (A), gelatin (B), fibrin (C) dan fibrinogen (D) ... 40 Gambar 20. Analisis zimogram stabilitas pH dari enzim murni
DAFTAR TABEL
Halaman Tabel 1. Kandungan zat gizi pada cacing tanah ... 4
Tabel 2. Kebutuhan asam amino esensial (Boutrif 1991) ... 5 Tabel 3. Sekuen N-terminal asam amino dari fraksi-fraksi
Lumbrokinase murni (Cho et al. 2004) ... 8 Tabel 4. Karakteristik protease dari cacing tanah ... 9 Tabel 5. Metode kromatografi untuk fraksinasi protein enzim
(Ersson et al. 1998)... 15 Tabel 6. Prosedur analisis aktivitas protease dengan metode
15 DAFTAR LAMPIRAN
Halaman Lampiran 1. Diagram alir pemurnian enzim serupa tripsin dari
cacing tanah ... 49 Lampiran 2. Komposisi gel pemisah dan penahan untuk SDS PAGE ... 50 Lampiran 3. Komposisi gel pemisah dan penahan untuk zimografi.... ... 50 Lampiran 4. Karakteristik kolom Hitrap Benzamidine FF ... 51 Lampiran 5. Prosedur pembuatan pereaksi kimia ... 52 Lampiran 6. Teknik pengembangan dan pengemasan matriks
Sephadex G-100 ... 55 Lampiran 7. Kurva standar Bradford ... 56 Lampiran 8. Kurva standar zimogram 10% ... 57 Lampiran 9. Kristal struktur komponen A enzim fibrinolitik dari
E. fetida ... 58 Lampiran 10. Metode seleksi awal enzim fibrinolitik pada media
agar fibrin ... 61
PENDAHULUAN
Latar Belakang
Reaksi-reaksi enzimatis dalam makhluk hidup merupakan suatu hal yang sangat penting untuk kelangsungan hidupnya, yaitu dengan berperan dalam sejumlah reaksi biokimia seluler. Salah satu enzim tersebut adalah protease. Protease tidak hanya diperlukan untuk degradasi protein yaitu sebagai sumber nutrien makhluk hidup tetapi juga berperan dalam sejumlah mekanisme patogenisitas, proses koagulasi darah, proses sporulasi, diferensiasi, proses pasca translasi protein dan mekanisme ekspresi protein ekstra seluler (Rao et al. 1998). Sedangkan penggunaannya dalam industri meliputi industri pangan maupun non pangan. Dalam industri pangan protease telah lama digunakan sebagai pengempuk daging, penjernih bir, pembuatan keju, kecap ikan, protein hidrolisat dan komponen aroma serta untuk memperbaiki tekstur roti dan kue. Untuk industri non pangan protease digunakan dalam pembuatan deterjen, untuk pengolahan kulit, penelitian-penelitian dalam bidang bioteknologi modern misalnya dalam teknik ekstraksi DNA serta dalam aplikasi medis yaitu sebagai agen terapeutik (Rao et al. 1998; Suhartono 1992).
Enzim protease merupakan enzim yang memegang 60% dari total penjualan enzim dunia yang mencapai dua miliar US $ dengan peningkatan nilai jual mencapai enam hingga tujuh persen per tahun, sedangkan dari kuantitasnya meningkat sebesar 10 hingga 15% per tahun (Suhartono 2000). Impor enzim Indonesia juga terus meningkat yaitu dari senilai 124,1 juta US $ pada tahun 2000 (BPS 2000) menjadi 127,4 juta US $ (BPS 2001).
Kasiat cacing tanah dalam pengobatan telah lama diketahui, terutama oleh masyarakat Cina, Jepang, Korea dan Taiwan. Spesies cacing tanah yang digunakan antara lain Lumbricus kwangtungensis, L. natives, L. rubellus, Eisenia foetida dan E. rosea. Walaupun berbeda spesiesnya, tepung cacing biasanya dijual dengan nama dagang tepung Lumbrikus.
17 Pada saat ini enzim fibrinolitik yang telah digunakan sebagai agen terapeutik pengobatan penyakit penyumbatan pembuluh darah adalah urokinase dan tissue plasminogen activator (t-PA). Tetapi dalam aplikasinya, enzim-enzim ini segera didegradasi dalam sirkulasi darah sebelum mencapai efek terapeutiknya, selain itu beberapa efek samping seperti anaphylaxis dan reaksi imunologi juga telah dilaporkan (Yokoigawa et al. 1991; Nakajima et al. 1993). Kelebihan lumbrokinase antara lain dapat dikonsumsi secara oral, aman, non toksik dan tidak menimbulkan efek samping terhadap fungsi hati, jantung, ginjal, sistem respirasi dan sistem saraf.
Tujuan Penelitian
TINJAUAN PUSTAKA
Cacing Tanah (Lumbricus rubellus)
Cacing tanah L. rubellus berasal dari Eropa, sehingga dikenal juga dengan sebutan cacing Eropa, cacing introduksi atau cacing Jayagiri (Rukmana 1999). Jenis cacing ini mampu hidup pada media rendah nutrisi, daya reproduksinya tinggi (106 kokon/tahun) serta mampu menghasilkan kompos dari bahan organik dalam jumlah besar.
Pengklasifikasian cacing tanah L. rubellus (Hegner dan Engemann 1968) adalah sebagai berikut :
Dunia : Animalia Divisi : Vermes Filum : Annelida Kelas : Oligochaeta Ordo : Opisthopora Famili : Lumbricidae Genus : Lumbricus Spesies : rubellus
19 Cacing L. rubellus telah dimanfaatkan secara luas antara lain untuk penghasil pupuk organik, bahan pakan ternak dan ikan, umpan pancing, bahan baku obat dan kosmetik serta bahan baku makanan-minuman.
Palungkun (1999) menyatakan bahwa kandungan protein cacing tanah jauh lebih tinggi dibandingkan kandungan lemaknya, dan tersusun atas sembilan asam amino esensial dan empat asam amino non esensial. Cacing tanah juga mengandung fosfor, kalsium dan serat kasar. Kandungan berbagai zat gizi dalam cacing tanah disajikan pada Tabel 1.
Tabel 1 Kandungan zat gizi pada cacing tanah
Zat Gizi Komposisi (%)
Protein 64 - 76
Asam amino esensial - arginin - histidin - isoleusin - leusin - lisin - metionin - fenilalanin - treonin - valin 4,13 1,56 2,58 4,84 4,33 2,18 2,25 2,95 3,01 Asam amino non esensial
- sistein - glisin - serin - tirosin 2,29 2,92 2,88 1,36
Lemak 7 - 10
Serat kasar 1,08
Fosfor (P) 1,00
Kalsium (Ca) 0,55
Sumber : Palungkun (1999)
Tabel 2 Kebutuhan asam amino esensial (Boutrif 1991)
Asam amino Anak-anak (2-5 tahun) Tikus percobaan (mg/g protein)
Arginin - 50
Histidin 19 25
Isoleusin 28 42
Leusin 66 62
Lisin 58 58
Metionin dan sistin 25 50
Fenilalanin dan tirosin 63 66
Treonin 34 42
Triptofan 11 12,5
Valin 35 50
Cacing tanah diketahui mampu untuk mengakumulasi logam-logam berat dalam jaringannya, yaitu dalam bentuk tidak larut sehingga tidak mengganggu reaksi-reaksi biokimia dalam sitoplasma. Mekanisme toleransi ini diduga disebabkan oleh adanya gen yang menyandikan protein berbobot molekul rendah yang kaya akan sistein (metalotionein) (Sturzenbaum et al. 1998).
Selain kemampuannya untuk bertahan terhadap adanya kontaminasi logam berat, cacing tanah diketahui juga menghasilkan peptida dengan sifat antimikroba. Cho et al. (1998) telah berhasil mengisolasi dan mengkarakterisasi peptida tersebut dari L. rubellus serta diberi nama lumbrisin I. Peptida ini kaya akan prolin dari total 62 asam amino (15% prolin dalam rasio molar dengan berat molekul 7.231 Da). Peptida tersebut menunjukkan aktivitas antimikroba in vitro pada sejumlah besar mikroorganisme tanpa aktivitas hemolitik. Sedangkan dari E. foetida juga telah berhasil diisolasi peptida dengan aktivitas antimikroba yaitu fetidin (Milochau et al.
1997; Lassegues et al. 1997) dan OEP3121 dengan berat molekul 510,8 Da serta komposisi asam aminonya adalah ACSAG (Yan et al. 2004). Fetidin menunjukkan aktivitas antimikroba terhadap Bacillus megaterium sedangkan OEP3121 menunjukkan aktivitas antimikroba terhadap bakteri uji Eschericia coli,
Staphylococcus aureus dan Pseudomonas aeruginosa.
Protease Cacing Tanah
21 sederhana. Protease dapat dihasilkan secara ekstraseluler dan intraseluler dari tanaman, hewan dan mikroba (Ward 1983).
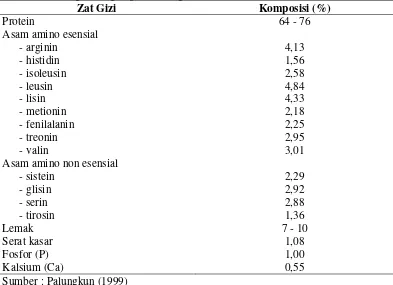
Creighton (1997) membagi protease menjadi empat kelas utama berdasarkan gugus fungsional pada sisi aktifnya, yaitu serin (E.C. 3.4.21), tiol atau sulfidril (E.C. 3.4.22), karboksil atau asam (E.C. 3.4.24) dan metal (logam) (E.C. 3.4.23). Protease tiol atau sulfidril dihambat oleh adanya senyawa oksidator, alkilator dan logam berat yaitu dengan mengikat gugus tiolnya. Enzim-enzim yang termasuk ke dalam golongan protease tiol adalah papain, fisin dan bromelin. Protease metal (logam) merupakan jenis enzim protease yang dapat diaktifkan oleh adanya ion-ion logam seperti Mg, Zn, Co, Fe, Cd dan Cu. Golongan enzim ini dihambat oleh adanya senyawa pengkelat logam seperti EDTA. Contoh protease yang termasuk ke dalam golongan protease metal antara lain karboksi peptidase A, beberapa amino peptidase dan beberapa protease bakteri. Protease asam merupakan golongan enzim protease yang keaktifannya disebabkan oleh adanya gugus karboksil pada sisi aktifnya. Golongan enzim protease ini dihambat oleh adanya bromofenolsil bromida atau pelarut diazo. Enzim-enzim yang termasuk ke dalam golongan ini antara lain pepsin, renin dan protease lain yang aktif pada kisaran pH rendah (antara 2 hingga 4).
Protease serin merupakan suatu endopeptidase misalnya tripsin, kimotripsin, elastase dan subtilisin. Golongan enzim ini umumnya aktif pada kondisi pH netral dan alkali (7-11). Protease serin dihambat oleh senyawa diisopropilflorofosfat (DFP), fenilmetilsulfonilflorida (PMSF), N-p-torsil-L-lisin-klorometilketon (TLCK), soy bean trypsin inhibitor (SBTI) dan lima bean trypsin inhibitor atau aprotinin (LBTI). Perbandingan mekanisme reaksi enzimatis dari beberapa golongan protease disajikan pada Gambar 1.
Protease cacing tanah dihasilkan secara intraseluler, dan pada tahun 1991 Mihara et al. berhasil mengekstrak dan memurnikan enam fraksi protease dari cacing
mengkatalisis hidrolisis ester terutama etil asetat dan bioplastik poli (R)3-hidroksibutirat. substrat O H + C X O R
Im ImH+
O X C R O -O C O R Im + HX
Produk tetrahedral intermediat enzim asil Asilasi
Deasilasi A. Protease serin
substrat H + C X O R ImH+ X C R O -C O R Im + HX
Produk tetrahedral intermediat enzim asil Asilasi
Deasilasi S
-B. Protease tiol
S S
ImH+
C. Protease karboksil
Asp 32 C O O -H O C O Asp 215 O H H O C NHR' R Asp 32 C O O H O C O Asp 215 OH O C NHR' R -Asp 32 C O O -H O C O Asp 215 OH C NHR' R OH Asp 32 C O O H O C O Asp 215 OH O C
NH2R' R
-D. Protease Metal
C O Zn2+ O -O H H +C O NH R' R C O Zn2+ OH C O -O H NH R' R C O Zn2+ O -O C OH RH2O
[image:32.612.162.495.145.674.2]23 Berdasarkan pemecahan ikatan peptida substrat, lumbrokinase termasuk ke dalam endopeptidase karena menguraikan ikatan peptida pada rantai dalam protein secara acak sehingga didapatkan produk peptida dan polipeptida (Nakajima et al.
1996). Sedangkan dari segi sisi aktif enzim, lumbrokinase termasuk ke dalam protease serin (E.C. 3.4.21.99). Penelitan Mihara et al. (1991) menunjukkan bahwa lumbrokinase dari L. rubellus dihambat secara spesifik oleh DFP, LBTI dan SBTI. Hasil ini didukung oleh Cho et al. (2004) dan Yanti (2003) dengan menggunakan inhibitor PMSF. Penelitian lebih lanjut menggunakan aprotinin, TLCK, N torsil-L-fenilalanin klorometil keton (TPCK), SBTI, LBTI dan leupeptin menunjukkan penghambatan yang signifikan sedangkan EDTA tidak menunjukkan adanya penghambatan sehingga disimpulkan bahwa fraksi-fraksi enzim lumbrokinase termasuk ke dalam protease serin serupa tripsin (Cho et al. 2004; Park et al. 1998). Dari uji sekuen N-teminal asam amino pada protease cacing menunjukkan bahwa enzim memiliki similaritas lokal dengan protease serin lainnya seperti plasmin, tripsin, kimotripsin, elastase dan faktor koagulan IX (Nakajima et al. 1993; 1996). Pada Tabel 3 disajikan sekuen N-terminal asam amino dari fraksi-fraksi enzim Lumbrokinase murni.
Tabel 3 Sekuen N-terminal asam amino dari fraksi-fraksi Lumbrokinase murni (Cho
et al. 2004)
Fraksi Sekuen N-terminal asam amino
Tabel 4 Karakteristik protease dari cacing tanah
Mihara
et al.
(1991)
Nakajima
et al.
(1993)
Park
et al
.
(1998)
Yanti (2003)
Cho
et al.
(2004)
Wang
et al.
(2003)
Fraksi
6
6
2
6
6
7
Bobot
molekul
(kD)
23,5 – 34,2
24,0 – 43,0
34,0 dan
34,2
20 – 41
24,6 – 33,0
23,028 – 29,690
pI
3,52 - 4,12
3,40 – 4,85
-
-
-
3,46 – 3,94
pH optimum
7,4 – 9,0
9 – 11
-
8,0
4 – 12
-
Suhu
optimum
(
oC)
37
60
-
60
50
-
Stabilitas
(pH, suhu)
3 – 10
37 – 60
oC
1 – 11
60
oC
2 – 11
65
oC
2 – 12
55
oC
4 – 12
50 – 55
oC
-
Inhibitor
DFP,SBTI, LBTI
DFP, SBTI,
aprotinin
TLCK
PMSF
PMSF, aprotinin,
TLCK,TPCK, SBTI,
LBTI, leupeptin
-
Jenis enzim
Protease serin
Protease tripsin
Protease
tripsin
Protease serin
Protease tripsin
Protease serin
Spesifitas
substrat
-
Kasein, elastin,
kolagen, albumin,
keratin, hemoglobin
-
Kasein, albumin,
protein susu,
gelatin,
fibrinogen, fibrin
-
S4760, M4765,
S7388, I6886, B7632,
T6140, V6258,
Chromozyme t-PA
Teknik
Pemurnian
Sephadex G-100 dan
N
α
-(
ε
-aminokaproil)-DL-homoarginin
heksilester Sepharose
-
-
Streamline DEAE
dan Superdex 75
DEAE toyopearl 650,
fenil toyopearl,
benzamidine sepharose
6B, Sephacryl S-200
Hiprep DEAE,
Resource Q,
Resource ISO, Mono
Q, Resource PHE
Keterangan : S4760 (substrat elastase), M4765 (substrat untuk leukosit elastase manusia), S3788 (substrat kimotripsin), I6886
(D-25 Struktur protease cacing L. rubellus berupa satu rantai polipeptida tunggal yang tersusun atas 282 asam amino dengan ukuran molekul 30 kD (Choi et al. 1996). Hasil analisis SDS-PAGE menunjukkan protease murni L. rubellus terdiri dari enam fraksi enzim dengan ukuran 23,5; 27,4; 27; 28,5; 34 dan 34,2 kD (Mihara et al. 1991), 24; 27; 36; 38; 40 dan 43 kD (Nakajima et al. 1993), 34 dan 34,2 kD (Park et al.
1998), 24,6; 26,8; 28,2; 25,4; 33,1; 33,0 kD (Cho et al. 2004), 20; 23; 26; 31; 36; 41 kD (Yanti 2003). Sedangkan spesies cacing tanah lain yaitu E. foetida menunjukkan adanya tujuh fraksi enzim dengan berat molekul 24,663; 29,515; 29,690; 29,595; 24,201; 24,170; 23,028 kD (Wang et al. 2003). Lumbrokinase kaya asam amino asparagin dan asam aspartat tetapi miskin akan lisin dan prolin (Mihara et al. 1991; Nakajima et al. 1993). Selain itu lumbrokinase tidak mengandung gula (Nakajima et al. 1993). Pada Tabel 4 disajikan perbandingan karakteristik protease hasil pemurnian dari cacing tanah.
Potensi Lumbrokinase Sebagai Enzim Trombolitik
Obat trombolitik berperan untuk melarutkan trombus dengan cara mengubah plasminogen menjadi plasmin, suatu protease serin yang menghidrolisis fibrin dan melarutkan bekuan darah (Mycek et al. 2001). Fibrin merupakan zat pengikat dari gumpalan darah (trombin).
Obat-obat trombolitik dapat dibedakan menjadi dua yaitu fibrinolisin (elase) dan Zat Aktivator Plasminogen/ZAP (Tjay dan Rahardja 2002). Elase merupakan enzim fibrinolitik yang langsung merombak jaringan fibrin dari trombus dan protein plasma lainnya, seperti fibrinogen, faktor beku V dan VIII. Sedangkan ZAP bekerja secara tidak langsung dengan menstimulasi perubahan plasminogen menjadi plasmin yang selanjutnya melarutkan fibrin dalam trombus melalui sistem fibrinolisis. Umumnya ZAP merupakan enzim berdaya fibrinolitik seperti streptokinase, urokinase, stafilokinase, anistreplase, aktivator plasminogen jaringan (t-PA), alteplase dan reteplase.
Streptokinase merupakan isolat protein ekstraseluler dari filtrat kultur
banyak dengan harga lebih murah, tetapi selama proses pemurnian mudah terkontaminasi oleh protein lain sehingga dapat menimbulkan efek antigenik.
Urokinase adalah enzim yang disintesis oleh ginjal dan bekerja secara langsung mengubah plasminogen menjadi plasmin aktif serta menguraikan fibrin dan fibrinogen (Mycek et al. 2001, Katzung 2002). Awalnya urokinase diisolasi dari urin manusia, tetapi sekarang dapat diproduksi dari kultur sel jaringan ginjal manusia sehingga harganya mahal dan jumlahnya terbatas. Keunggulan urokinase antara lain toksisitasnya rendah, tidak menyebabkan reaksi alergi, tidak menginduksi pembentukan antibodi, tidak menimbulkan efek samping dan afinitasnya lebih selektif pada fase pembentukan trombus (Suhartono 1992).
Anistreplase (anisoylated plasminogen streptokinase activator complex; ASPAC) merupakan kompleks plasminogen murni manusia dengan streptokinase yang telah diasilasi untuk melindungi situs aktif enzim (Katzung 2002). Kelompok asil akan dihidrolisis pada saat digunakan sehingga kompleks streptokinase-plasminogen aktif akan terbebaskan. Plasmin yang tebentuk akan menguraikan jaringan fibrin. Kelebihan dari anistreplase adalah selektivitasnya lebih tinggi dan aktivitas trombolitiknya lebih kuat dan lama.
Secara endogenus plasminogen dapat diaktifkan oleh aktivator plasminogen jaringan (t-PA), yaitu suatu protease serin yang berasal dari sel melanoma manusia. Enzim ini akan mengaktifkan plasminogen yang terikat pada fibrin sehingga fibrinolisis yang terjadi hanya terbatas pada trombus yang terbentuk saja (Mycek et al. 2001). Contoh t-PA yang diperoleh dari teknik DNA rekombinan adalah alteplase dan reteplase. Alteplase merupakan t-PA manusia yang belum dimodifikasi sedangkan reteplase adalah t-PA manusia yang telah dihilangkan beberapa sekuen asam aminonya (Katzung 2002).
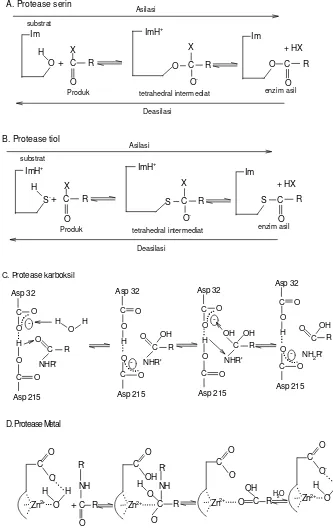
27 Mekanisme kerja lumbrokinase sebagai obat trombolitik secara umum dapat dibedakan menjadi tiga tahapan yaitu menstimulasi plasminogen menjadi plasmin, menghidrolisis fibrin hingga larut dan mendegradasi fibrinogen (Gambar 2). Lumbrokinase memiliki aktivitas fibrinolitik dengan aktivitas serupa tripsin, yaitu dengan pemotongan rantai β-fibrinogen. Sedangkan terhadap rantai γ-fibrinogen laju pemotongannya lebih lambat (Hwang et al. 2002).
Plasminogen
Plasmin
Produk degradasi Fibrinogen Trombin Fibrin
lumbrokinase,
urokinase, streptokinase
lumbrokinase lumbrokinase,
urokinase, streptokinase
[image:37.612.136.505.217.379.2]Produk degradasi ZAP: streptokinase, stapilokinase, urokinase, anistreplase, alteplase, reteplase, lumbrokinase
Gambar 2 Mekanisme kerja lumbrokinase sebagai obat trombolitik (Yanti 2003)
Pengujian in vitro aktivitas fibrinolitik lumbrokinase cair dan amobil menunjukkan bahwa enzim tersebut mampu untuk mendigesti fibrin dengan atau tanpa plasminogen sehingga dapat berpotensi sebagai ZAP maupun digesti langsung (Mihara et al. 1991; Ryu et al. 1994, 1995; Park et al. 1995). Sedangkan dari pengujian in vivo pada hewan tikus dan kelinci menunjukkan bahwa enzim lumbrokinase menghambat pembekuan darah dengan cara memacu plasminogen dalam sel-sel darah sehingga aktivitas penguraian darah meningkat (Kim et al. 1993, 1998; Fan et al. 2001). Pengujian in vivo dari tepung cacing tanah (L. rubellus) dan pengendapan protein enzim menggunakan garam amonium sulfat terhadap monyet
Penelitian Jin et al. (2000) menunjukkan bahwa dari uji in vivo lumbrokinase ternyata efektif untuk penderita aterosklerosis yang pembuluh darahnya mengalami pengapuran. Parameter yang diamati adalah kadar trombosit pasien aterosklerosis yang meningkat sehingga menyulitkan kerja plasminogen dalam menguraikan bekuan darah. Selain aterosklerosis terapi enzim lumbrokinase juga telah diterapkan pada penderita penyakit infark jantung, serebrovaskular dan emboli paru.
Pemurnian Enzim
Tujuan utama pemurnian enzim adalah mengisolasi enzim dengan kemurnian maksimal pada tingkat rendemen yang maksimal dengan biaya yang efektif (Harris dan Angal 1989). Secara umum pemurnian enzim dapat dibagi menjadi ekstraksi, pemekatan dan fraksinasi.
Ekstraksi enzim
Proses ekstraksi bergantung pada sumber dan lokasi enzim. Protease cacing tanah merupakan enzim intraseluler sehingga memerlukan suatu perlakuan khusus untuk memecah dinding sel jaringan untuk mengeluarkan enzim. Proses ekstraksi pada sel hewan dilakukan setelah hewan mati dan dibersihkan, lalu dibekukan untuk meminimalkan reaksi autolisis. Selanjutnya bahan non protein (deposit lemak dan jaringan koneksi) dihilangkan dengan cara memecah membran sel sesuai dengan lokasi enzim.
Ekstraksi protease cacing dilakukan dengan cara pengeringan vakum agar diperoleh produk berupa tepung cacing. Kelebihan tepung cacing adalah lebih tahan lama, karakteristik sifat tetap dan seimbang, tidak mempengaruhi penampakan fisik (warna dan bau), kandungan bakteri kontaminan minim, dan tidak mengalami degradasi selama proses. Jika disimpan pada suhu ruang, protease tepung cacing L. rubellus dapat bertahan hingga lima tahun (Nakajima et al. 2000).
29 lisozim, pembekuan cacing pada suhu -60 hingga -10oC dan pengeringan dengan liofilisasi, oven vakum dan pengeringan semprot.
Pemekatan enzim
Pemekatan enzim dilakukan untuk memisahkan konsentrat protein dari komponen biomolekul lainnya (karbohidrat, lipida dan asam nukleat). Beberapa metode pemekatan yang lazim digunakan dalam pemurnian enzim adalah presipitasi dengan garam, pelarut organik, polimer, dialisis, ultrafiltrasi dan liofilisasi.
Presipitasi dengan garam (amonium sulfat, natrium sulfat) lebih disukai daripada presipitasi dengan pelarut organik (etanol, aseton) dengan alasan pelarut organik cenderung mendenaturasi protein pada suhu agak tinggi, harga mahal dan mudah terbakar. Selain itu presipitasi menggunakan pelarut organik dipengaruhi oleh konsentrasi pelarut organik, konsentrasi protein, kekuatan ionik, pH dan suhu. Garam amonium sulfat dipilih karena kelarutannya tinggi, harganya murah, tidak beracun dan tidak mempengaruhi struktur protein. Sementara presipitasi menggunakan polimer (polietilen glikol) sama halnya dengan presipitasi menggunakan pelarut organik. Hanya polietilen glikol lebih mudah ditangani daripada pelarut organik karena tidak mudah terbakar, tidak beracun, tidak bermuatan dan murah (Suhartono 1989).
Sisa garam dari proses presipitasi enzim dihilangkan dengan cara dialisis menggunakan kantong selofan dan ultrafiltrasi. Sehingga diperoleh konsentrat enzim bebas garam untuk dimurnikan lebih lanjut melalui teknik fraksinasi enzim.
Fraksinasi enzim
[image:39.612.125.521.628.710.2]Tahap akhir dari pemurnian enzim adalah teknik fraksinasi, yang dilakukan dengan tujuan untuk memisahkan protein enzim dari protein non enzim lainnya. Metode fraksinasi yang umum dilakukan adalah kromatografi kolom dan elektroforesis. Pemilihan metode kromatografi kolom bergantung pada sifat protein enzim yang ingin dimurnikan (Tabel 4).
Tabel 5 Metode kromatografi untuk fraksinasi protein enzim (Ersson et al. 1998)
Sifat protein Jenis Kromatografi
Ukuran dan bentuk Filtrasi gel
Muatan neto dan distribusi gugus bermuatan Penukar ion
Titik isolistrik kromatofokusing
Hidrofobisitas Interaksi hidrofobik dan fasa
Pengikatan logam Afinitas ion logam teramobilisasi
Afinitas biospesifik terhadap ligan, inhibitor, reseptor dan antibodi
Afinitas
[image:40.612.131.449.244.457.2]Sehingga perlu dilakukan pengumpulan informasi tentang karakteristik enzim yang akan dimurnikan seperti perkiraan bobot molekul, pI, derajat hidrofobisitas dan adanya gugus sulfidril. Beberapa ilustrasi dari teknik kromatografi kolom dapat dilihat pada Gambar 3.
Gambar 3 Ilustrasi beberapa metode fraksinasi enzim
Elektroforesis Gel
Beberapa teknik elekroforesis untuk memisahkan protein dapat dilakukan dengan mengeksploitasi beberapa sifat utama protein yaitu ukuran, muatan bersih dan hidrofobisitas relatif (Dunn 1989). Beberapa matriks yang dapat digunakan adalah pati, agarosa, selulosa asetat, walaupun kapasitas resolusi yang tinggi dari poliakrilamida menyebabkan metode menggunakan matriks ini yang paling sering digunakan.
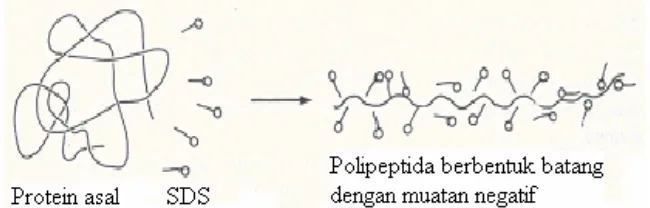
31 amonium persulfat (Dunn 1989). Prinsip analisis SDS-PAGE adalah pemisahan protein berdasarkan berat molekul. Penambahan deterjen anionik (SDS), β -merkaptoetanol dan pemanasan akan merusak struktur tiga dimensi protein. Beta-merkaptoetanol akan memecah ikatan disulfida dan mereduksinya menjadi gugus sulfidril, sedangkan SDS akan bereaksi dengan protein membentuk kompleks bermuatan negatif sehingga protein bergerak dalam medan listrik hanya berdasarkan pada ukuran molekul. Protein berukuran kecil akan bergerak lebih cepat dibandingkan protein berukuran lebih besar (Copeland 1994).
Gambar 4 Pengaruh SDS terhadap protein
Zimografi
Zimografi merupakan suatu teknik untuk menganalisis aktivitas proteolitik (Liota dan Stetler-Stevenson 1990). Mekanisme kerjanya menyerupai SDS-PAGE hanya pada gel pemisah dikopolimerasi substrat protein enzim yang akan diuji. Setelah pemisahan elektroforesis, substrat tersebut akan didegradasi oleh protein enzim yang telah direnaturasi, pada kondisi reaksi (suhu dan bufer optimum enzim) selama waktu tertentu (Kim et al. 1998).
BAHAN DAN METODE
Bahan dan Alat
Bahan baku yang digunakan adalah cacing tanah (L. rubellus) yang diperoleh dari budidaya lokal di Serpong, Tangerang. Bahan kimia penting yang digunakan adalah standar pereaksi Bradford; Folin Ciocalteau (Merck), Bovine Serum Albumine/BSA Fraction V (Merck); kolagen (Sigma), fibrin (Sigma), fibrinogen (Sigma), amonium sulfat teknis; kolom Hitrap Benzamidine FF (Amersham Biosciences); porcine dan bovine tripsin (EC 3.4.21.4) dari Sigma, dan pereaksi elektroforesis SDS-PAGE serta zimogram (Bio-Rad).
Alat-alat utama yang digunakan dalam penelitian adalah spektrofotometer UV/Vis double beam (Hitachi), High Speed Centrifuge CR21G (Hitachi), sentrifugasi mikro (Beckman), inkubator, pH meter, pengering vakum, deep-freezer, kantong dialisis cut-off 12 kD (Sigma), fraction collector dan perangkat elektroforesis Mini-PROTEAN III (Bio-Rad).
Metode Penelitian Pembuatan Tepung Cacing (Modifikasi Yanti 2003)
Satu kilogram cacing L. rubellus dibersihkan dan dicuci dengan air mengalir. Lalu cacing tersebut direndam dalam 2,5 L akuades pH 5,8 (dengan penambahan asam sitrat dan 1,5 g KH2PO4) selama 2,5 jam untuk menghilangkan kotorannya. Selanjutnya cacing dibekukan menggunakan freezer, suhu -17oC lalu diblender dan dibekukan kembali. Pengeringan cacing dilakukan dengan menggunakan pengering vakum, suhu 50oC dan tekanan 300 psi selama 48 jam. Cacing kering diblender dan disaring (± 900 mesh) sehingga diperoleh tepung cacing.
Pemurnian Enzim Serupa Tripsin Dari Cacing Tanah (Modifikasi Mihara 1991; Yanti 2003)
33 dalam 50 mM bufer fosfat pH 8,0 lalu disentrifugasi pada kecepatan 2.000 g, 4oC selama 5 menit. Supernatan didialisis dalam 20 mM bufer fosfat pH 8,0 dengan menggunakan kantung dialisis (cut-off 12 kD).
Dialisat disaring dengan kertas saring 0,45 µm lalu dipekatkan dengan menggunakan teknik liofilisasi. Hasil pemekatan dimurnikan dengan menggunakan filtrasi gel (matriks Sephadex G-100).
Pemurnian Enzim Serupa Tripsin Menggunakan Matriks Sephadex G-100 Dialisat yang telah dipekatkan dimurnikan lebih lanjut menggunakan kolom Sephadex G100, dengan bufer elusi 50 mM bufer fosfat pH 8,0. Setiap fraksi diatur agar diperoleh 100 tetes (sekitar 3 ml).
Pemurnian Enzim Serupa Tripsin Menggunakan Kromatografi Afinitas
Sebelum dimurnikan dengan menggunakan kolom Hitrap Benzamidine FF sampel enzim dengan aktivitas tertinggi pada pemurnian dengan teknik filtrasi gel dipekatkan lalu disaring dengan kertas saring 0,45 µm. Hasil pemekatan ini diencerkan dengan bufer 50 mM Tris HCl, 0,5 M NaCl, pH 7,4. Bufer pengikat dan pencuci yang digunakan adalah 0,05 M Tris-HCl, 0,5 M NaCl pH 7,4. Bufer elusi adalah 0,05 M glisin pH 3,0. Setiap tabung pengumpul fraksi diisi dengan 0,4 ml bufer 1 M Tris-HCl pH 9,0 dan digunakan menampung 2 ml eluat.
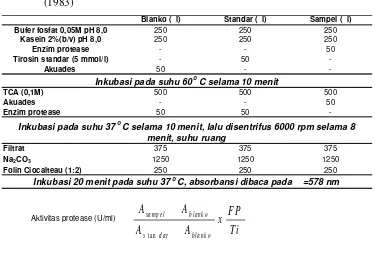
Analisis Aktivitas Protease
Tabel 6 Prosedur analisis aktivitas protease dengan metode Bergmeyer dan Grassl (1983)
Blanko ( l) Standar ( l) Sampel ( l) Bufer fosfat 0,05M pH 8,0 250 250 250
Kasein 2%(b/v) pH 8,0 250 250 250
Enzim protease - - 50
Tirosin standar (5 mmol/l) - 50
-Akuades 50 -
-TCA (0,1M) 500 500 500
Akuades - - 50
Enzim protease 50 50
-Filtrat 375 375 375
Na2CO3 1250 1250 1250
Folin Ciocalteau (1:2) 250 250 250
Aktivitas protease (U/ml)
Inkubasi pada suhu 60oC selama 10 menit
Inkubasi pada suhu 37oC selama 10 menit, lalu disentrifus 6000 rpm selama 8 menit, suhu ruang
Inkubasi 20 menit pada suhu 37oC, absorbansi dibaca pada =578 nm
T i
F P
x
A
A
A
A
b la n k o d a r
s
b la n k o sa mp el
tan
Analisis Kadar Protein
Kadar protein ditentukan dengan metode Bradford (1976). Sebanyak 100 µl larutan enzim ditambahkan kedalam tabung berisi 2,5 ml akuades dan 2,5 ml pereaksi kerja Bradford. Perlakuan pada blangko, larutan enzim diganti dengan akuades. Selanjutnya larutan tersebut divorteks dan didiamkan selama 5 menit pada suhu ruang. Absorbansi larutan diukur pada panjang gelombang 595 nm.
Standar protein yang digunakan adalah BSA Fraction V. Pada kurva standar protein, larutan enzim digantikan dengan BSA Fraction V dengan kisaran konsentrasi 0 – 1000 µg/ml. Konsentrasi protein larutan enzim ditentukan berdasarkan persamaan garis linear hubungan antara konsentrasi standar protein dengan absorbansi.
Analisis SDS-PAGE Dan Zimografi
35 Preparasi gel pemisah dan gel penahan
Pembuatan gel pemisah akrilamida 10% dan gel penahan 4% untuk SDS-PAGE dan zimografi dilakukan dengan komposisi yang tercantum pada Lampiran 2 Preparasi sampel dan loading
Untuk SDS-PAGE, 20 µl sampel ditambahkan dengan 5 µl bufer sampel. Tiap sampel diloading ke dalam sumur gel dengan kisaran volume 10-20 ml, dengan penggunaan marker Low Molecular Weight/LMW (Pharmacia) sekitar 5 µl.
Kondisi running, pewarnaan dan pelunturan warna
Gel dilarikan pada tegangan 100 V selama 1,5 jam dalam bufer elektroforesis. Untuk SDS-PAGE, setelah elektroforesis gel langsung diwarnai dengan larutan pewarna Coomasie Brilliant Blue R-250 selama 15 menit. Pelunturan warna dilakukan dengan larutan peluntur sehingga didapatkan pita protein biru dengan latar gel bening. Teknik pewarnaan lain yang dilakukan adalah pewarnaan dengan perak nitrat. Setelah elektroforesis gel digoyang dalam larutan fiksasi selama satu jam, dilanjutkan dalam larutan etanol 50% (v/v) selama 20 menit. Kemudian dalam larutan etanol 30% (v/v) selama 20 menit ( dua kali). Setelah itu digoyang dalam larutan enhancer selama satu menit lalu dicuci dengan akuabidestilata selama 20 detik (tiga kali). Gel lalu digoyang dalam larutan perak nitrat selama 30 menit dan dibilas dengan akuabidestilata selama 20 detik (dua kali). Gel lalu direndam dalam larutan Na2CO3-formaldehid sehingga pita protein terlihat dan reaksi segera dihentikan dengan penambahan larutan fiksasi.
Untuk zimografi, setelah elektroforesis gel direnaturasi dalam larutan Triton X-100 2,5% v/v sambil digoyang selama satu jam. Kemudian gel didigesti dalam 50 mM bufer fosfat pH 8, suhu 60oC selama 20 menit. Gel diwarnai dengan larutan pewarna Coomasie Brilliant Blue R-250 selama 15 menit. Pelunturan warna gel dilakukan dengan larutan peluntur berulangkali sehingga didapatkan pita enzim proteolitik putih dengan latar gel biru.
Penentuan Suhu Optimum (Yanti 2003)
Penentuan pH Optimum dan Kestabilan pH(Yanti 2003)
Penentuan pH optimum untuk aktivitas enzim serupa tripsin dari L. rubellus
dilakukan pada beberapa sistem bufer yaitu bufer glisin-HCl (pH 2,0-3,5), bufer asetat 0,05 M (pH 3,5-5,5), bufer fosfat 0,05 M (pH 5,5-8,5), dan bufer glisin-NaOH 0,05 M (pH 8,5 hingga 12,0). Aktivitas enzim pada pH optimum ditentukan dengan mengukur aktivitasnya pada suhu optimum menggunakan metode Bergmeyer (1983). Penentuan stabilitas pH enzim dilakukan dengan cara menginkubasikan 50 µl enzim dengan 200 µl larutan bufer 0,05 M glisin HCl pH 2,0; 0,05 M bufer fosfat pH 7,0 dan 0,05 M bufer glisin NaOH pH 12,0 selama satu jam. Kestabilan pH ditentukan secara kualitatif dengan cara zimogram.
Pengaruh Deterjen, Inhibitor dan Urea (Yanti 2003)
Pengaruh inhibitor (EDTA 5 dan 1 mM serta PMSF dengan konsentrasi 1 dan 5 mM), deterjen (SDS 5% dan 1% b/v; Triton X-100 5% dan 1% b/v; Tween 20 5% dan 1% b/v) serta urea (5% dan 1% b/v) terhadap aktivitas enzim dilakukan dengan cara menginkubasikan 100 µl enzim dan 100 µl larutan senyawa selama satu jam pada suhu ruang, lalu dianalisis aktivitasnya.
Pengaruh Ion Logam (Yanti 2003)
37 Spesifitas substrat (Yanti 2003)
Uji aktivitas protease pada suhu dan pH optimum dilakukan pada berbagai substrat protein dengan konsentrasi 2% b/v. Substrat yang digunakan adalah kasein, fibrin, fibrinogen, BSA dan kolagen. Substrat tersebut dilarutkan dalam 50 mM bufer fosfat pH 8,0.
HASIL DAN PEMBAHASAN
Penentuan Rendemen dan Total Protein Cacing Tanah
Tepung cacing yang diperoleh dari pengeringan vakum cacing tanah segar dapat dibedakan menjadi dua bagian yaitu bagian kasar dan halus. Untuk bagian halus diperoleh rendemen sebesar 9,61% (berat basah), sedangkan untuk bagian kasar diperoleh rendemen sebesar 1,55%.
Dari penentuan protein terlarut (diasumsikan sebagai protein enzim) diperoleh kandungan protein sebesar 6,71 mg/ml dengan aktivitas spesifik sebesar 0,28 U/mg menggunakan substrat kasein.
Karakterisasi Protease Kasar Cacing Tanah
Enzim protease yang akan dimurnikan dari cacing tanah terdapat bersama-sama dengan berbagai macam protein lainnya sehingga untuk menentukan teknik pemurnian yang sesuai perlu dilakukan karakterisasi enzim kasar terlebih dahulu. Informasi awal tentang karakteristik enzim seperti perkiraan berat molekul, pH dan suhu optimum enzim, inhibitor spesifik untuk menentukan golongan enzim, pengaruh kation, deterjen dan senyawa denaturan, akan sangat menentukan dalam penentuan metode pemurnian yang tepat sehingga kondisi yang dilakukan dapat memurnikan enzim sasaran yang diinginkan.
Penentuan Suhu Optimum
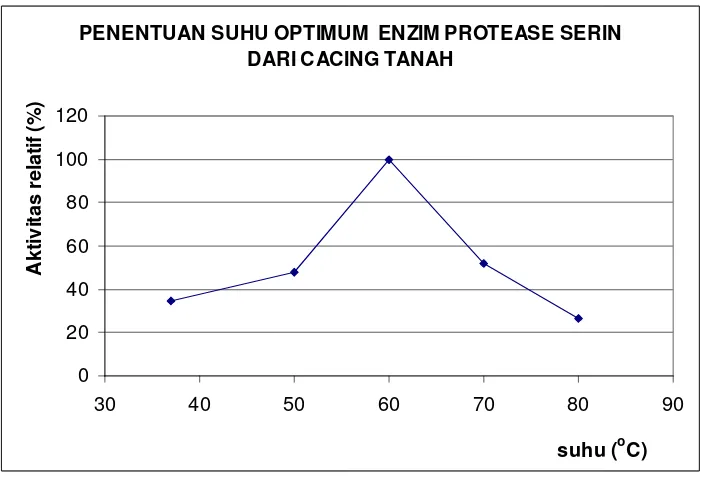
Dengan menggunakan 0,05 M bufer fosfat pH 7,0 diperoleh aktivitas optimum protease kasar cacing tanah pada suhu 60oC, dengan aktivitas sebesar 3,20 U/ml (Gambar 5). Peningkatan suhu inkubasi menjadi 70oC menyebabkan penurunan aktivitas relatif menjadi 52,1% dan jika suhu inkubasi ditingkatkan menjadi 80oC aktivitas relatif yang terukur menjadi 26,4%.
39
PENENTUAN SUHU OPTIMUM ENZIM PROTEASE SERIN DARI CACING TANAH
0 20 40 60 80 100 120
30 40 50 60 70 80 90
suhu (oC)
A
k
ti
v
it
as r
e
lat
if (%
[image:49.612.145.497.80.319.2])
Gambar 5 Suhu optimum aktivitas enzim kasar cacing tanah
Dalam reaksi enzimatis, suhu berperan dalam meningkatkan interaksi antara substrat dengan enzim sehingga dari Gambar 5 terlihat bahwa peningkatan suhu dari 37 oC hingga 60 oC menyebabkan aktivitas enzim meningkat. Tetapi enzim juga merupakan protein globular sehingga pada suhu yang terlalu tinggi dapat menyebabkan enzim terdenaturasi dan menyebabkan terjadinya penurunan aktivitas bahkan dapat menyebabkan hilangnya akvitas enzim. Denaturasi enzim disebabkan oleh rusaknya interaksi-interaksi non kovalen (ikatan hidrogen, interaksi Van der Waals, interaksi hidrofobik dan interaksi elektrostatik) yang berperan dalam menjaga keutuhan struktur tiga dimensi dari enzim (Hames dan Hooper 2000). Pada Gambar 6 ditampilkan struktur tiga dimensi enzim fibrinolitik komponen A dari salah satu spesies cacing tanah yaitu E. fetida. Dari gambar tersebut diperlihatkan bahwa enzim fibrinolitik cacing tanah tersusun atas tiga rantai polipeptida, sedangkan sekuen asam amino dari enzim fibrinolitik tersebut disajikan pada Lampiran 9.
Hasil penelitian Yanti (2003) menunjukkan bahwa enzim protease L. rubellus
Gambar 6 Struktur tiga dimensi enzim fibrinolitik komponen A dari E. fetida (Tang
et al. 2002)
Penentuan pH optimum
41
Penentuan pH optimum pada berbagai pH dan jenis bufer
0 20 40 60 80 100 120
1 2 3 4 5 6 7 8 9 10 11 12 13
pH A k ti v it as r el at if (% )
[image:51.612.147.493.75.264.2]bufer glisin HCl bufer asetat bufer fosfat bufer glisin NaOH
Gambar 7 Pengaruh pH dan larutan bufer terhadap enzim kasar cacing tanah Derajat keasaman dari lingkungan protein enzim akan mempengaruhi kondisi ionisasi gugus-gugus fungsionalnya yang berperan dalam keaktifan enzim. Gugus ionik berperan penting dalam menjaga konformasi sisi aktif enzim untuk mengikat dan mengubah substrat menjadi produk. Pemilihan pH bufer yang tidak tepat dapat menyebabkan terganggunya interaksi substrat dengan enzim. Sebagai contoh protease serin, misalnya tripsin mempunyai tiga gugus asam amino yang berperan penting dalam reaksi enzimatis yaitu Asp 102, Ser 195 dan His 57. Pada kondisi asam, Asp 102 dan His 57 akan terprotonasi sehingga Ser 195 tidak mampu untuk memutus ikatan peptida pada substrat. Perubahan pH yang ekstrim akan menyebabkan enzim mengalami denaturasi karena terganggunya interaksi-interaksi non kovalen yang menjaga kestabilan struktur tiga dimensi enzim (Hames dan Hooper 2000).
Pengaruh Ion Logam
Pengaruh ion logam terhadap aktivitas enzim kasar cacing tanah
0 20 40 60 80 100 120 140
Kontrol Na Co Cu Ba K Li Mn Mg Ca Zn Fe
Ion logam
A
k
ti
v
it
as r
el
at
if (%
)
[image:52.612.136.504.77.313.2]5 mM 1 mM
Gambar 8 Pengaruh ion logam terhadap aktivitas enzim kasar
Pengaruh Inhibitor, deterjen dan urea
Pengujian enzim kasar cacing tanah terhadap adanya EDTA, suatu senyawa pengkelat logam menunjukkan bahwa senyawa ini tidak berpengaruh terhadap aktivitas enzim sehingga dapat disimpulkan bahwa enzim protease cacing tanah tidak termasuk ke dalam golongan protease logam (Tabel 6). Protease logam memiliki atom logam pada sisi aktifnya sehingga bila atom logam ini dikelat oleh EDTA akan menyebabkan turunnya aktivitas enzim (Suhartono 1992).
43
F
S O
O
[image:53.612.290.347.86.194.2]CH2
Gambar 9 Struktur dua dimensi PMSF
Hasil ini didukung oleh beberapa publikasi yang juga menyatakan bahwa ekstrak enzim cacing tanah (L. rubellus) dihambat secara spesifik oleh inhibitor spesifik protease serin (Cho et al. 2004, Park et al. 1998, Nakajima et al. 1993, Mihara et al. 1991). Hasil serupa diperoleh Wang et al. (2003) pada cacing tanah E. foetida.
Tabel 7 Pengaruh inhibitor, deterjen dan urea
Bahan Konsentrasi
Aktivitas relatif (%)
kontrol 100
Tween 20 5% 109
1% 97
Triton X100 5% 111
1% 105
SDS 5% 102
1% 99
Urea 5 % 119
1 % 96
2-merkaptoetanol 1% 139
0,5% 133
EDTA 5 mM 83
1 mM 109
DMSO 50% 104
PMSF 5 mM 22
1 mM 66
[image:53.612.199.444.354.569.2]menyebabkan denaturasi protein dan bila dihilangkan mungkin dapat menyebabkan renaturasi protein enzim (Bolag dan Edestein 1991, Choi et al. 2001).
[image:54.612.153.491.285.498.2]Protein denaturan (urea) dan 2-merkaptoetanol (senyawa pereduksi) juga tidak mempengaruhi aktivitas enzim cacing tanah. Senyawa 2-merkaptoetanol merupakan senyawa pereduksi ikatan disulfida pada protein. Ikatan disulfida merupakan ikatan kovalen yang terbentuk dari dua residu sistein yang berperan penting dalam menjaga konformasi tiga dimensi protein. Hasil yang diperoleh berbeda dengan penelitian Yanti (2003) yang menyatakan bahwa ekstrak kasar enzim dihambat aktivitasnya, dan bahkan setelah dimurnikan akan dihambat total aktivitas proteolitiknya oleh 2-merkaptoetanol.
Gambar 10 Ikatan disulfida pada komponen A enzim fibrinolitik E. fetida
45 Sedangkan urea merupakan senyawa yang digunakan terhadap protein yang kelarutannya terbatas atau cenderung mengendap. Urea akan terurai dan membentuk ion-ion sianat yang bereaksi dengan gugus amino protein membentuk turunan-turunan karbamil yang stabil melalui ikatan ionik (Dunn 1989).
Spesifitas substrat
Enzim protease cacing tanah mampu mendigesti berbagai macam substrat seperti kasein, gelatin, albumin, keratin, fibrin, fibrinogen, keratin, elastin dan hemoglobin (Yanti 2003, Nakajima 1993). Kasein merupakan fosfoprotein yang terdiri atas unit struktur α-kasein (122 kD), β-kasein (24,1 kD) dan χ- kasein. Kasein merupakan protein utama pada susu, yaitu sekitar 80% dari total protein susu.
Gelatin merupakan hasil hidrolisis parsial dari kolagen dalam air mendidih. Polipeptida ini kaya akan residu prolin dan hidroksiprolin serta bersifat larut dalam air. Sedangkan kolagen bersifat tidak larut dan termasuk ke dalam protein serat.
Fibrin merupakan protein tidak larut yang berasal dari degradasi fibrinogen oleh trombin selama proses pembekuan darah. Degradasi fibrin dalam trombus dikatalisis oleh enzim plasmin atau fibrinolisin sehingga diperoleh peptida-peptida berukuran lebih kecil dan larut (Sadikin 2001).
Albumin merupakan protein larut air dan termasuk ke dalam protein globular. Albumin kaya akan asam amino hidrofobik, akan tetapi keberadaannya terletak sedemikian rupa sehingga berada pada bagian dalam. Sedangkan gugus hidrofilik berada di bagian luar sehingga protein ini larut air.
SPESIFITAS SUBSTRAT DARI PROTEASE CACING TANAH KASAR
0 10 20 30 40 50 60 70 80 90 100 110
kasein fibrinogen fibrin BSA kolagen gelatin
substrat
A
k
ti
v
it
as r
e
lat
if (
%
[image:56.612.163.478.81.287.2])
Gambar 11 Spesifitas substrat dari enzim kasar cacing tanah
Sebagai metode cepat untuk seleksi awal enzim fibrinolitik mungkin dapat dilakukan dengan cara mengkopolimerisasi substrat fibrin pada media agar, lalu dibuat lubang sumur sebagai tempat untuk ekstrak enzim yang akan diuji (Lampiran 10). Aktivitas fibrinolitik enzim ditunjukkan setelah pewarnaan menggunakan larutan staining, dimana terbentuknya halo disekeliling sumur menunjukkan adanya aktivitas fibrinolitik (Gambar 12).
Gambar 12 Metode skrining awal aktivitas fibrinolitik dari ekstrak enzim
Penentuan Berat Molekul
[image:56.612.248.393.471.579.2]47 51 kD (Gambar 13). Mihara et al. (1991) dengan teknik SDS PAGE melaporkan bahwa protease fibrinolitik dari cacing tanah L. rubellus terdiri dari enam fraksi dengan berat molekul 23,5; 27; 27,4; 28,5; 34 dan 34,2 kD. Publikasi Nakajima et al.
(1993) juga melaporkan bahwa terdapat enam fraksi enzim dari cacing tanah, dengan berat molekul 24, 27, 36, 38, 40 dan 43 kD. Cho et al. (2004) dengan teknik pengendapan menggunakan amonium sulfat dan kromatografi kolom berhasil mengisolasi enam fraksi fibrinolitik dengan ukuran 24,6; 26,8; 28,2; 25,4; 33,1 dan 33 kD. Spesies cacing tanah lain juga dilaporkan memiliki aktivitas fibrinolitik, dari cacing Eisenia foetida berhasil diisolasi tujuh fraksi enzim dengan berat molekul 24,66; 29,52; 29,69; 24,20; 24,17; 23, 03 dan 29,60 kD (Wang et al. 2003).
[image:57.612.130.494.301.529.2]
Gambar 13 Analisis zimogram kasein (A) (10%) dan SDS PAGE (B) (10%) dari ekstrak kasar cacing tanah. Baris M : marker LMW (α-laktalbumin 14,4 kD; soybean trypsin inhibitor 20,1 kD; karbonik anhidrase 30 kD; ovalbumin 45 kD; bovine serum albumin 66 kD dan fosforilase b 97 kD)
Selain dari cacing tanah, ternyata enzim fibrinolitik juga dapat dihasilkan oleh bakteri Bacillus amyloliquefaciens DJ-4 yang diisolasi dari makanan tradisional Korea, natto. Enzim fibrinolitik ini memilki berat molekul 38, 53 dan 80 kD (Choi dan Kim 2001). Publikasi Seo dan Lee (2004) dengan menggunakan Bacillus firmus
40 34 27 14,4
97 66 45 30 20,1
20 22 43
M S M S
A
51
NA-1 yang diisolasi dari natto juga berhasil menunjukkan bahwa strain bakteri tersebut mampu menghasilkan enzim fibrinolitik.
Pemurnian enzim
Protein enzim yang diinginkan dari suatu organisme terdapat dalam bentuk campuran dengan berbagai senyawa lain seperti lipida, protein/peptida maupun debris sel sehingga perlu dilakukan tahap pemurnian. Tahapan pemurnian enzim yang umum dilakukan meliputi pembuatan ekstrak, pengendapan baik dengan garam ataupun pelarut organik, dialisis dan kromatografi.
Pada penelitian ini setelah tepung cacing diekstraksi dalam 0,05 M bufer fosfat pH 8,0 selama 24 jam dan disentrifus dilakukan penambahan garam amonium sulfat teknis sebanyak 65% b/v (Subandrio 2004). Prinsip pengendapan protein enzim dengan garam amonium sulfat didasarkan pada efek salting in dan salting out. Pada konsentrasi rendah adanya garam akan meningkatkan kelarutan protein enzim, tetapi pada konsentrasi yang lebih tinggi keberadaan garam akan menyebabkan terjadinya pengendapan protein enzim. Jika dibandingkan dengan ekstrak kasar pengendapan amonium sulfat berhasil meningkatkan kemurnian enzim menjadi 1,13 kali dengan aktivitas spesifik sebesar 0,31 U/mg.
Garam amonium sulfat pada presipitat protein enzim dihilangkan dengan menggunakan kantung dialisis nitroselulosa asetat (cut off 12 kD). Penggunaan kantung dialisis dengan cut off 12 kD selain menghilangkan garam amonium sulfat juga akan menyebabkan molekul-molekul kecil berukuran kurang dari 12 kD keluar dari kantung dialisis ke dalam bufer perendam. Proses dialisis selain menghilangkan garam amonium sulfat dan molekul-molekul kecil lainnya juga menyebabkan terjadinya pengenceran terhadap presipitat protein enzim. Sehingga sebelum dimurnikan lebih lanjut dengan teknik filtrasi gel, dialisat dipekatkan terlebih dulu dengan teknik pengeringan beku. Kemurnian enzim berhasil ditingkatkan menjadi 1,33 kali dari ekstrak kasar dengan aktivitas spesifik sebesar 0,37 U/mg.
49 tanah dimurnikan dengan teknik kromatografi afinitas terlebih dulu dimurnikan dengan teknik filtrasi gel. Prinsip dari teknik kromatografi filtrasi gel adalah pemisahan molekul berdasarkan perbedaan ukurannya. Matriks yang digunakan adalah Sephadex G-100 dengan kisaran pemisahan 4 hingga 150 kD (Scopes 1986). Dari Gambar 14 terlihat bahwa pemurnian menggunakan Sephadex G-100 berhasil mendapatkan dua puncak protein, sedangkan berdasarkan aktivitasnya diperoleh satu puncak dengan aktivitas sebesar 1,62 U/ml.
Matriks Sephadex G-100 tersusun atas ikatan silang dekstran dengan epiklorhidrin. Porositas dari matriks merupakan penentu batasan dari molekul yang dapat dipisahkan (Pingoud et al. 2002). Fraksi dengan aktivitas tertinggi dipekatkan terlebih dulu dengan teknik pengeringan beku sebelum dilakukan pemurnian lebih lanjut menggunakan kolom kromatografi afinitas Hitrap Benzamidine FF.
P E M U R N IAN D IAL IS AT C AC IN G T AN AH D E N G AN T E K N IK K R O M AT O GR AG I F IL T R AS I GE L
0 0.75 1.5 2.25 3 3.75 4.5 5.25 6 6.75 7.5 8.25 9
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30
[image:59.612.130.508.327.540.2]Fra ksi U V ( 280 n m ) -0.20 0.40 0.60 0.80 1.00 1.20 1.40 1.60 1.80 2.00 U /m l absorbansi aktivitas (U/ml)
Gambar 14 Pemurnian enzim protease cacing tanah dengan teknik kromatografi filtrasi gel
sehingga diduga pengikatan antara enzim dengan ligan terjadi pada sisi aktif serin, yaitu pada gugus OH dari asam amino serin dengan gugus NH dari p-aminobenzamidin (Gambar 16).
Gambar 15 Matriks kolom Hitrap Benzamidin FF
Setelah enzim target berikatan dengan ligan maka terdapat beberapa cara untuk melepaskan enzim dari ligan yaitu elusi menggunakan bufer dengan pH rendah, elusi menggunakan bufer yang ditambahkan ligan kompetitor dan menggunakan denaturan seperti urea 8 M atau guanidin hidroklorida 6 M. Pada penelitian ini digunakan elusi menggunakan bufer dengan pH rendah yaitu 0,05 M bufer glisin pH 3,0.
R C
O
N H R '
+ E O H R C
O
-H N
R '
O E
H+ H
N N
H
Inhib ito r E nzim
Gambar 16 Interaksi enzim protease serin dengan inhibitor (Creighton 1997)
[image:60.612.192.448.381.504.2]51
PEMURNIAN MENGGUNAKAN KOLOM HITRAP BENZAMIDINE FF
-0.05 0.10 0.15 0.20 0.25 0.30 0.35 0.40 0.45 0.50 0.55 0.60 0.65 0.70 0.75
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 2122 23 24 25 26 27 28 2930 3132 33 3435 36
[image:61.612.129.518.79.256.2]Fraksi -10.00 20.00 30.00 40.00 50.00 60.00 70.00 80.00 90.00 100.00 110.00 120.00 [protein](mg/ml) aktivitas (U/ml) aktivitas spesifik (U/mg)
Gambar 17 Pemurnian enzim protease cacing tanah dengan teknik kromatografi afinitas
Eluat fraksi nomor 8 dan 24 diuji lebih lanjut dengan menggunakan teknik SDS PAGE dan zimografi untuk mengetahui estimasi berat molekul dan kemampuan digesti fibrin dan fibrinogen serta kestabilannya terhadap pH. Ringkasan tahap pemurnian protease dari cacing tanah disajikan pada Tabel 8.
Hasil pemurnian menggunakan kolom Hitrap Benzamidin FF memperoleh isolat enzim murni dengan nilai aktivitas spesik yang tinggi 1,92 U/mg (fraksi 8) dan 111,36 U/mg (fraksi 24) atau dapat diperoleh tingkat kemurnian sebesar 6,97 dan 404,20 kali dari ekstrak awal enzim cacing tanah. Sedangkan hasil pemurnian menggunakan kolom Streamline-DEAE dan Superdex 75 prep grade berhasil memperoleh tiga fraksi dengan aktivitas spesifik 6,136; 6,945 dan 1,527 U/mg atau kemurnian isolat enzim yang diperoleh adalah 12,73; 14,41 dan 3,17 kali dari ekstrak awal cacing tanah (Yanti 2003).
Tabel 8 Ringkasan tahap pemurnian protease cacing tanah Tahap Vol
[image:61.612.129.521.636.692.2]Presipitat 65% 50 2,91 145,35 9,35 467 0,31 61,63 1,13 Dialisat 57 2,27 129,29 6,18 352 0,37 54,82 1,33 Eluat Sephadex 22 1,62 35,73 1,19 26 1,37 15,15 4,96
Eluat Hitrap
H8 H24
2,4 2,4
0,54 0,04
1,31 0,09
0,28 0,0003
0,68 0,0008
1,92 111,36
0,55 0,06
6,97 404,20
Profil pita protein hasil pemurnian menggunakan kolom kromatografi afinitas disajikan pada Gambar 18. Konsentrasi protein sampel fraksi delapan yang digunakan adalah 0,224 mg/ml sedangkan dari fraksi nomor 24 adalah 0,00024 mg/ml. Konsentrasi protein sampel ini yang digunakan dalam analisis-analisis selanjutnya menggunakan teknik SDS PAGE dan zimografi.
[image:62.612.124.520.77.205.2] [image:62.612.135.494.330.506.2]
Gambar 18 Analisis SDS PAGE (10%) (A) dan zimogram fibrin (10%) (B) dari enzim murni cacing tanah. Baris M : marker LMW (α-laktalbumin 14,4 kD; soybean trypsin inhibito