ISOLASI DAN SELEKSI BAKTERI SELULOLITIK
TERMOFILIK DARI KAWAH AIR PANAS
GUNUNG PANCAR, BOGOR
HARTANTI
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
1
ABSTRAK
HARTANTI. Isolasi dan Seleksi Bakteri Selulolitik Termofilik dari Kawah Air
Panas Gunung Pancar, Bogor. Dibimbing oleh SURYANI dan I MADE ARTIKA.
Selulase memiliki peran penting dalam industri kertas, tekstil, makanan,
dan detergen. Aplikasi selulase pada industri membutuhkan selulase yang aktif
dan stabil pada suhu tinggi. Eksplorasi bakteri termofilik dari kawah air panas
merupakan salah satu cara untuk mendapatkan selulase termostabil. Penelitian ini
bertujuan mengisolasi bakteri termofilik dari kawah air panas Gunung Pancar
yang menghasilkan selulase termostabil. Aktivitas selulolitik ditentukan oleh
kemampuan bakteri dalam menghidrolisis substrat
carboxymethyl cellulose
ABSTRACT
HARTANTI. Isolation and Selection Thermophilic of Cellulolytic Bacteria from
Crater of Mount Pancar Hot Spring, Bogor. Under the direction of SURYANI and
I MADE ARTIKA.
1
ISOLASI DAN SELEKSI BAKTERI SELULOLITIK
TERMOFILIK DARI KAWAH AIR PANAS
GUNUNG PANCAR, BOGOR
HARTANTI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biokimia
DEPARTEMEN BIOKIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi : Isolasi dan Seleksi Bakteri Selulolitik Termofilik dari Kawah Air
Panas Gunung Pancar, Bogor
Nama
: Hartanti
NIM
: G84050551
Disetujui
Komisi Pembimbing
Dr. Suryani, M.Sc Dr. I Made Artika, M.App.Sc
Ketua Anggota
Diketahui
Dr. I Made Artika, M.App.Sc
Ketua Departemen Biokimia
1
PRAKATA
Alhamdulillah, segala puji bagi Allah SWT atas segala nikmat dan
karuniaNya sehingga penulis dapat menyelesaikan penelitian dan penulisan karya
ilmiah ini. Karya ilmiah yang berjudul Isolasi dan Seleksi Bakteri Selulolitik
Termofilik dari Kawah Air Panas Gunung Pancar, Bogor, ditulis berdasarkan hasil
penelitian di Laboratorium Biokimia selama bulan Maret sampai September 2009
sebagai prasyarat untuk memperoleh gelar Sarjana Sains pada Departemen
Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam.
Ungkapan terimakasih penulis sampaikan kepada Dr. Suryani, M.Sc dan
Dr. I Made Artika, M.App.Sc selaku pembimbing atas bimbingan, saran, kritik,
serta dukungannya selama penelitian dan penulisan karya ilmiah ini. Terimakasih
kepada ayah, ibu, kakak atas segala dorongan dan semangat yang diberikan, serta
kepada Riza, Mela, Fitria, Dini, Ratna, Bemby, dan Puspa atas bantuannya selama
penelitian ini berlangsung.
Semoga karya ilmiah ini bermanfaat bagi perkembangan ilmu pengetahuan
Indonesia. Amin.
Bogor, Februari 2010
Hartanti
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 1 Agustus 1987 dari ayahanda
Mai Wiharja dan ibunda Eti Suriati. Penulis merupakan putri keempat dari empat
bersaudara.
Tahun 2005 penulis lulus dari SMUN 1 LeuwiLiang-Bogor dan pada
tahun yang sama lulus seleksi masuk Institut Pertanian Bogor melalui jalur
Undangan Seleksi Masuk IPB (USMI). Penulis tercatat sebagai mahasiswa
Departemen Biokimia, Fakultas Matematika dan Ilmu Pengetahuan Alam.
1
DAFTAR ISI
Halaman
DAFTAR GAMBAR ... vi
DAFTAR LAMPIRAN ... vi
PENDAHULUAN .. ...1
TINJAUAN PUSTAKA
Bakteri Termofilik ... 1
Bakteri Selulolitik ... 2
Kompleks Selulase ... 3
Pemanfaatan Selulase ...4
Isolasi dan Seleksi Bakteri ...4
BAHAN DAN METODE
Alat dan Bahan ...5
Metode ...5
HASIL DAN PEMBAHASAN
Isolat Bakteri Hasil Peremajaan ... 6
Hasil Isolasi Koloni Tunggal Isolat Kawah Putih dan Kawah Merah ... 7
Hasil Seleksi Bakteri Selulolitik ...8
Pola Pertumbuhan Bakteri Selulolitik ...9
Hasil Uji Aktivitas Selulase Secara Kuantitatif ... 10
SIMPULAN DAN SARAN
Simpulan ... 12
Saran ... 12
UCAPAN TERIMAKASIH ... 12
DAFTAR GAMBAR
Halaman
1 Sisi aktif selulase ...3
2 Tahapan-tahapan hidrolisis selulosa ...4
3 Hasil inkubasi bakteri Kawah Putih, media kontrol, dan bakteri
Kawah Merah ...7
4 Hasil pengenceran isolat Kawah Putih dan Kawah Merah ...8
5 Hasil seleksi aktivitas selulolitik isolat Kawah Putih dan Kawah Merah ...9
6 Pola pertumbuhan bakteri selulolitik isolat KP4 dan KM1 ... 10
DAFTAR LAMPIRAN
Halaman
1 Tahapan penelitian ... .... 16
2 Pembuatan media, bufer, dan pereaksi DNS ... 17
3 Pembuatan kurva standar glukosa ... 18
4 Perhitungan konsentrasi glukosa sampel ... 19
1
PENDAHULUAN
Enzim merupakan produk bioteknologi yang menarik perhatian karena peranannya dalam berbagai bidang, terutama bidang industri. Salah satu enzim yang berpotensi dalam bidang industri adalah selulase. Selulase dapat diaplikasikan dalam industri kertas, tekstil, makanan, dan detergen. Selain itu, enzim ini digunakan untuk meningkatkan kualitas nutrisi pakan ternak dan berperan penting dalam biokonversi selulosa menjadi berbagai komoditas senyawa kimia (Ibrahim & El-diwany 2007).
Industri-industri yang memanfaatkan selulase umumnya beroperasi pada suhu tinggi sehingga dibutuhkan enzim yang memiliki stabilitas dan aktivitas yang tinggi pada kondisi suhu ekstrim. Kegiatan yang saat ini semakin intensif dilakukan adalah eksplorasi mikroba termofilik yang dapat menghasilkan selulase termostabil dari berbagai sumber alam. Pemanfaatan enzim termostabil dalam bidang industri sangat menguntungkan karena laju reaksi lebih cepat, mengurangi resiko kontaminasi, mengurangi biaya pendinginan pada proses fermentasi skala besar, kelarutan substrat yang tinggi, dan menurunkan viskositas media tumbuh (Edwards 1990).
Organisme yang sering digunakan sebagai penghasil enzim termostabil adalah mikroba, seperti bakteri dan fungi. Hal ini karena biodeversitas mikroba tinggi sehingga menyediakan sumber enzim yang dapat dieksplorasi secara terus-menerus. Mikroba dapat memproduksi enzim dengan laju yang sangat cepat dan dengan biaya yang rendah. Hal ini terkait dengan kultivasi mikroba dan pemurnian enzim yang dihasilkan. Laju produksi enzim dapat ditingkatkan melalui seleksi galur, induksi mutan, dan memperbaiki kondisi kultur. Selain itu, dapat dilakukan melalui rekayasa genetik sehingga dapat menyediakan berbagai jenis enzim, bahkan yang berasal dari mikroba yang hidup pada lingkungan yang ekstrim (Ratledge & Kristiansen 2001).
Produksi selulase termostabil dapat dilakukan melalui eksplorasi bakteri termofilik penghasil selulase yang berasal dari lingkungan ekstrim alam Indonesia. Salah satu lingkungan ekstrimnya adalah kawah air panas yang merupakan habitat bagi pertumbuhan bakteri termofilik. Kawah air panas yang dapat ditemukan di daerah Bogor salah satunya terdapat di kawasan wisata air
panas Gunung Pancar. yang terletak di Desa Karang Tengah, Citeureup, Bogor.
Isolat yang digunakan dalam penelitian ini adalah hasil isolasi dari penelitian sebelumnya oleh Harahap (2007), yang belum diketahui potensinya dalam menghasilkan selulase termostabil. Penelitian ini bertujuan mengisolasi bakteri termofilik dari kawah air panas Gunung Pancar yang berpotensi menghasilkan selulase termostabil. Hipotesis penelitian ini adalah isolat bakteri dari kawah air panas Gunung Pancar, Bogor, berpotensi menghasilkan selulase termostabil.
Isolat bakteri termofilik penghasil selulase yang diperoleh dari penelitian ini diharapkan dapat memiliki potensi untuk diaplikasikan pada proses-proses industri yang melibatkan penggunaan selulase pada suhu tinggi. Selain itu, eksplorasi dan isolasi bakteri termofilik penghasil selulase dapat digunakan sebagai salah satu cara untuk mengenalkan potensi dan keragaman bakteri termofilik dari kawah air panas di wilayah Indonesia. Selain itu, dapat menambah koleksi bakteri termofilik penghasil selulase yang berasal dari isolat lokal.
TINJAUAN PUSTAKA
Bakteri Termofilik
Bakteri termofilik merupakan kelompok bakteri yang memiliki kondisi pertumbuhan optimum pada suhu tinggi (Nam et al. 2004). Bakteri termofilik berbeda dengan sel-sel eukariotik karena kemampuannya untuk beradaptasi dan tumbuh pada suhu tinggi serta kondisi ekstrim, seperti salinitas tinggi (NaCl jenuh), pH ekstrim (<2.0,>10.0), dan tekanan substrat. Berdasarkan kisaran suhu pertumbuhannya, bakteri termofilik terdiri atas tiga golongan, yaitu termofilik (45-65 o
C), ekstrim termofilik (65-85 oC), dan hipertermofilik (85-110 oC) (Andrade et al.
1999).
berhasil diisolasi dari daerah sumber panas bumi, sedimen lautan geotermal, dan kawah air panas. Termofilik halofil dari laut dalam juga telah berhasil diisolasi. Salah satunya adalah spesies Thermus yang dapat tumbuh dengan adanya NaCl 3% atau lebih (Edwards 1990).
Selain kedua lingkungan tersebut, bakteri termofilik juga dapat ditemukan pada tanah, kompos, sampah, dan lumpur sungai. Fikrinda (2000) berhasil mengisolasi bakteri termofilik yang berasal dari tanah ekosistem air hitam, Kalimantan Tengah. Mikroba termofilik yang telah ditemukan dalam kompos, diantaranya adalah Thermonospora, Thermoactinomyces, Thermonospora chromogena, Streptomyces, Bacillus, Actinomycetes, dan Thermus
(Mayende et al. 2006).
Eksplorasi terhadap bakteri termofilik yang berasal dari sumber air panas Gunung Pancar juga sudah dilakukan. Delapan belas isolat termofil aerob yang berhasil diisolasi dari sumber air panas Gunung Pancar memiliki berbagai aktivitas enzim hidrolitik ekstraseluler, yaitu proteolitik, amilolitik, lipolitik, kitinolitik, dan xilanolitik (Dirnawan 1999). Harahap (2007), berhasil melakukan amplifikasi gen 16S-rRNA bakteri yang diisolasi dari Kawah Merah, Putih, Asin, dan Hitam Gunung Pancar. Bakteri yang berasal dari Kawah Hitam juga telah diketahui memiliki potensi sebagai bioflokulan (Susanti 2007). Suyono et al. (2008), mengisolasi bakteri termofilik penghasil asam laktat dari Kawah Putih Gunung Pancar.
Menurut Brock (1986), terdapat tiga faktor yang menyebabkan bakteri termofilik mampu bertahan dan berkembang dalam kondisi suhu tinggi, yaitu kandungan enzim dan protein lebih stabil dan tahan panas dibandingkan dengan mesofil, molekul pensintesis protein (seperti ribosom dan komponen lainnya) stabil terhadap panas, dan membran lipid sel termofil mengandung banyak asam lemak jenuh yang membentuk ikatan hidrofobik yang sangat kuat. Bakteri termofilik mampu mensintesis molekul stabil, seperti enzim yang mampu mengkatalis reaksi-reaksi biokimia pada suhu tinggi dan lebih stabil dibandingkan dengan enzim dari mesofil. Enzim ini tidak hanya stabil terhadap suhu tinggi tetapi juga terhadap protein-protein denaturan, seperti detergen, pelarut organik, serta enzim protease (Andrade et al. 1999).
Sifat-sifat tersebut sangat diperlukan oleh industri-industri berbasis enzim. Oleh karena itu, bakteri termofilik menawarkan
keuntungan dalam bidang industri dan bioteknologi (Mayende 2006). Enzim termostabil dari bakteri termofilik juga cocok digunakan sebagai model untuk menentukan termostabilitas protein dan potensinya sebagai biokatalis dalam bioteknologi modern. Aplikasi lain dari enzim termofilik adalah pengembangan proses baru untuk mengurangi pelepasan bahan kimia berbahaya ke lingkungan dengan mengganti reaksi kimia yang ada dengan proses enzimatik, yang dikenal dengan bioremediasi (Andrade et al.
1999).
Bakteri Selulolitik
Bakteri selulolitik adalah bakteri yang mampu menghidrolisis kompleks selulosa menjadi oligosakarida yang lebih kecil dan akhirnya menjadi glukosa. Glukosa tersebut digunakan sebagai sumber karbon dan sumber nutrisi bagi pertumbuhan organisme ini. Bakteri selulolitik mensintesis seperangkat enzim yang mampu menghidrolisis selulosa. Enzim tersebut adalah kompleks selulase. Enzim ini disintesis oleh mikroba selama tumbuh dalam media selulosa (Ibrahim & El-diwany 2007).
Mikroba yang mampu mendegradasi selulosa kristal dapat mensekresikan kompleks selulase (Shimada et al. 1994). Selulase dihasilkan sebagai respon terhadap adanya selulosa pada lingkungannya. Proses ini berlangsung apabila terjadi kontak langsung antara sel bakteri dan permukaan selulosa (Busto et al. 1995). Kemampuan biosintesis selulase dimiliki oleh banyak mikroba (Raza & shafiq-Ur-Rehman 2008). Mikroba penghasil selulase secara ekstraseluler tersebar pada kapang dan bakteri. Meskipun bakteri selulolitik memiliki sistem metabolisme yang berbeda dengan kapang dan sedikit sekali data tentang bakteri penghasil enzim ini, akan tetapi, umumnya diasumsikan memiliki tingkah laku yang sama (Fikrinda 2000).
3
Selain itu, jumlah dan komponen selulase yang dihasilkan dipengaruhi oleh jenis substrat, konsentrasi substrat, dan suhu (Aguiar 2001).
Beberapa bakteri selulolitik termofilik telah berhasil diisolasi dan diidentifikasi. Ibrahim & El-diwany (2007), telah mengisolasi bakteri selulolitik termofilik yang memiliki aktivitas optimum pada suhu 75 oC dari kawah air panas Egyptian, pantai laut merah. Fikrinda (2000), mengisolasi bakteri selulolitik yang mampu tumbuh pada suhu 60-70 oC dari ekosistem air hitam Kalimantan Tengah.
Secara alami, bakteri dapat menghidrolisis selulosa baik secara aerob maupun anaerob, akan tetapi tidak dapat secara kedua-duanya (Lynd et al. 2002). Bakteri selulolitik anaerob hanya tumbuh pada sumber selulosa dan produk hidrolitiknya. Bakteri ini tidak dapat tumbuh pada monosakarida, oligosakarida, dan polisakarida yang berasal dari gula lain selain glukosa (Lynd et al. 2002). Bakteri selulolitik aerob dapat menggunakan sumber karbon lain di samping glukosa.
Kompleks Selulase
Selulase merupakan kelompok enzim yang dapat mengkatalis hidrolisis ikatan -1.4-glikosidik dalam selulosa, selodekstrin, selobiosa, dan turunan selulosa lainnya (Raza & Shafiq-Ur-Rehman 2008). Molekul tersebut dihidrolisis menjadi unit-unit monomer yang lebih kecil, seperti glukosa. Enzim ini mampu menghidrolisis ikatan -1.4-glikosidik di antara residu glikosil melalui mekanisme hidrolisis asam (Lynd et al. 2002). Struktur selulase terdiri atas satu pusat katalitik, daerah pengikatan selulosa, dan rantai terglikosilasi. Sisi aktif enzim ditunjukkan pada Gambar 1.
Selulase diklasifikasikan menjadi tiga kelompok, yaitu endo-1,4- -D-glukanase (EC 3.2.1.4), exo-1,4- -D-glukanase (EC 3.2.1.92), dan -D-glukosidase (EC 3.2.1.21) (Sinegani & Emtiazi 2006). Ketiga komponen enzim tersebut bekerjasama dalam menghidrolisis selulosa yang tidak dapat larut menjadi glukosa sehingga aktivitas gabungan dari ketiga enzim tersebut dapat diukur dengan mengidentifikasi jumlah glukosa yang dihasilkannya (Fikrinda 2000).
Endoglukanase merupakan komponen selulase yang menghidrolisis daerah amorf selulosa secara acak. Hidrolisis oleh enzim ini membentuk oligosakarida dengan panjang rantai yang berbeda-beda dan membentuk
ujung rantai non-pereduksi baru (Sinegani & Emtiazi 2008). Endoglukanase selalu ditemukan dalam mikroba selulolitik, baik fungi maupun bakteri. Endoglukanase menghidrolisis selodekstrin dan turunan selulosa lain, seperti carboxymethylcellulose
(CMC). Enzim ini memiliki aktivitas yang tinggi terhadap substrat CMC sehingga disebut pula dengan CMCase.
Eksoselobiohidrolase (eksoglukanase) adalah komponen enzim yang produk hidrolisis utamanya adalah selobiosa. Enzim ini memecah selulosa dengan cara menghilangkan ujung akhir gugus selobiosa pada rantai selulosa (Raza & Shafiq-Ur-Rehman 2008). Eksoglukanase memiliki aktitivitas tinggi dalam menghidrolisis selulosa kristal tetapi sangat rendah pada selulosa amorf (Sinegani & Emtiazi 2006). Selobiohidrolase juga memiliki kemampuan dalam menghidrolisis selotriosa maupun selotetraosa yang merupakan hasil kerja endoglukanase. Kerja sama endoglukanase dan eksoglukanase dapat menghasilkan hidrolisis selulosa yang optimum (Fikrinda 2000).
-glukosidase merupakan komponen selulase yang memutuskan unit glukosa secara spesifik dari ujung nonpereduksi dari selooligosakarida (Sinegani & Emtiazi 2006). Enzim ini tidak menghidrolisis CMC atau selulosa tetapi menghidrolisis selooligosakarida, pNPG, dan selobiosa menjadi glukosa. Komponen enzim ketiga dari selulase ini, bertugas dalam mengatur seluruh proses selulolitik dan merupakan enzim terpenting meskipun tidak beraksi secara langsung dalam menghidrolisis selulosa. Hal tersebut dikarenakan kemampuan enzim ini dalam menghidrolisis selobiosa yang merupakan penghambat bagi aktivitas endoglukanase dan eksoglukanase dalam menghidrolisis selulosa (Fikrinda 2000).
Efektivitas biokonversi selulosa tergantung dari sumber selulase, jenis substrat selulosa, dan kondisi optimum untuk produksi dan aktivitas enzim (Alam et al. 2004). Aktivitas total selulase ditentukan dari aktivitas campuran enzim yang menghidrolisis bahan yang mengandung selulosa dan menghasilkan produk akhir berupa glukosa. Aktivitas ini menggambarkan pengaruh dari kerja ketiga enzim yang berbeda dan pengaruh hambatan dari produk akhir. Secara umum, hidrolisis selulosa oleh kompleks selulase terbagi ke dalam tiga tahapan utama (Gambar 2).
Tahap pertama dilakukan oleh enzim endoglukanase. Enzim ini menyerang daerah amorf dari selulosa secara acak dan membentuk ujung-ujung nonpereduksi yang akan memudahkan kerja eksoglukanase. Tahapan selanjutnya dilakukan oleh eksoglukanase yang menghidrolisis daerah kristal dari selulosa dengan melepaskan selobiosa. Selobiosa akan dihidrolisis oleh -glukosidase menghasilkan glukosa (Raza & Shafiq-Ur-Rehman 2008).
Gambar 2 Tahapan-tahapan hidrolisis selulosa.
Pemanfaatan Selulase
Selulase dalam industri kertas digunakan sebagai biopulping dan biobleaching. Selulase
dapat memperhalus bubur kertas, membantu proses pemutihan (bleaching) sehingga kertas lebih putih, dan memperkuat serat kertas sehingga permukaannya menjadi lebih halus. Sebelumnya, proses tersebut dilakukan secara konvensional menggunakan bahan kimia, seperti klorin dan klorin dioksida sebagai agen pemutih. Proses tersebut dapat menghasilkan sekitar 95 % limbah cair yang dapat mencemari lingkungan (Andrade et al 1999).
Selulase berperan dalam proses deinking, yaitu penghilangan tinta agar memperoleh serat kertas yang baik pada pembuatan kertas daur ulang. Proses ini secara konvensional dilakukan dengan menggunakan natrium hidroksida, hidrogen peroksida, agen pengkelat, dan natrium silikat. Penggunaan bahan-bahan kimia tersebut dapat mencemari lingkungan karena mempengaruhi nilai BOD dan COD air (Risjimana et al. 2002).
Penggunaan selulase pada industri tekstil berfungsi menjaga warna kain agar tetap cemerlang (Ibrahim & El-diwany 2007). Selulase juga berperan sebagai pembersih komponen tumbuhan pada proses akhir pembuatan wol. Selulase dapat berikatan secara kuat dan selektif pada molekul selulosa sehingga dimanfaatkan sebagai agen pembawa bahan pelembut (softener) menuju serat kain pada industri detergen. Industri minuman (jus buah-buahan) menggunakan selulase untuk meningkatkan perolehan jus lebih banyak karena selulase dapat menguraikan sari-sari buah yang masih terperangkap dalam buah (Heaton 2004).
Salah satu peran selulase yang terpenting adalah biokonversi selulosa menjadi berbagai senyawa kimia yang bermanfaat. Selulosa memegang peranan yang sangat potensial sebagai sumber serat, energi, dan makanan (Baig et al. 2004). Senyawa ini baru dapat dimanfaatkan setelah mengalami pengolahan yang optimal dengan memanfaatkan selulase. Selulase mampu menghidrolisis selulosa menjadi glukosa. Glukosa dapat ditransformasi menjadi etanol, butanol, aseton, protein sel tunggal, dan metan (Fikrinda 2000). Selain itu, hidrolisis selulosa dan hemiselulosa dengan selulase dapat meningkatkan efisiensi limbah pertanian sebagai bahan pakan dan dapat mengurangi dampak negatif dari polusi limbah terhadap lingkungan (Sari 2008).
Isolasi dan Seleksi Bakteri
Bakteri di habitat alaminya terdapat dalam populasi campuran. Spesies yang akan
5
diidentifikasi harus dipisahkan dari populasi campuran tersebut sehingga diperoleh biakan murni. Teknik isolasi digunakan untuk memperoleh biakan murni yang terdiri atas satu jenis bakteri yang diinginkan. Tahap pertama yang harus dilakukan dalam proses isolasi adalah pengambilan contoh mikroba dari habitat aslinya. Teknik pengambilan contoh harus sesuai dengan ekologi organisme tersebut dalam lingkungan alaminya. Menurut Labeda (1990), untuk mencari mikroba baru, maka harus memperhatikan berbagai kombinasi faktor ekologi yang ada.
Faktor penting yang harus diperhatikan dalam mengisolasi bakteri adalah komponen media yang digunakan untuk menumbuhkan bakteri tersebut. Media harus berisi zat hara untuk pertumbuhan bakteri. Selain nutrisi, hal lain yang perlu diperhatikan adalah faktor lingkungan, seperti keasaman dan suhu media. Kondisi pH ekstrim dapat merusak struktur dinding sel bakteri sehingga merusak sistem metabolismenya (Pelczar & Chan 2008).
Media yang digunakan untuk isolasi dapat berupa media diferensial, media selektif, dan media penyubur (Pelczar & Chan 2008). Media diferensial merupakan media penunjang kehidupan berbagai kelompok bekteri. Media selektif-diferensial digunakan untuk tujuan identifikasi. Media penyuburan sering digunakan untuk memacu pertumbuhan organisme yang diharapkan sehingga dapat diisolasi (Hidayat et al. 2006). Isolat yang diperoleh dimurnikan dan ditumbuhkan dalam media padat.
Setelah diisolasi, bakteri yang diperoleh dapat diseleksi. Tahapan ini bertujuan menguji kemampuan isolat menghasilkan produk yang diharapkan. Seleksi biasanya menggunakan uji sederhana atau menggunakan media spesifik dengan substrat tertentu (Hidayat et al. 2006).
Proses isolasi bakteri termofilik sama dengan teknik isolasi pada umumnya. Beberapa tahapan yang dapat dilakukan untuk mengisolasi dan seleksi bakteri termofilik adalah pemilihan substrat yang sesuai dengan bakteri yang akan diisolasi. Sebagian sampel yang diperoleh dapat ditumbuhkan langsung dalam agar (dicawankan) atau disuspensikan dan diencerkan dalam media cair sebelum dicawankan. Tahapan selanjutnya adalah prosedur penyuburan dengan cara menambahkan nutrisi tertentu agar meningkatkan populasi bakteri yang ingin
diisolasi. Setelah itu, dilakukan pemurnian, dan isolasi bakteri yang diinginkan (Labeda 1990).
BAHAN DAN METODE
Alat dan Bahan
Alat-alat yang digunakan adalah autoklaf,
laminar air flow cabinet, jarum ose, inkubator, sentrifus, spektrofotometer, termometer, pH meter, freezer, tabung Eppendorf, neraca analitik, magnetic stirrer, cawan Petri, dan peralatan gelas yang umum digunakan di laboratorium.
Bahan-bahan yang digunakan adalah isolat bakteri termofilik yang berasal dari kawah merah dan putih Gunung Pancar, Bogor, yang telah diisolasi sebelumnya oleh Harahap (2007). Bahan lain yang digunakan adalah larutan amonium sulfat (NH)4SO4, magnesium sulfat (MgSO4.7H2O), kalsium klorida (CaCl2.2H2O), kalium dihidrogen fosfat (KH2PO4), natrium klorida (NaCl), asam sitrat, dinatrium hidrogen fosfat (Na2HPO4), asam klorida (HCl), natrium hidroksida (NaOH), ekstrak khamir, pepton, tripton, bakto agar, natrium azida, standar glukosa, pereaksi asam-3,5-dinitrosalisilat (DNS), carboxymethyl cellulose (CMC) 1%, merah kongo (congo red) 0,1 %, etanol 70%, dan akuades.
Metode Penelitian
Penelitian ini dilakukan dalam lima tahap yang meliputi: peremajaan bakteri, isolasi bakteri penghasil selulase, seleksi bakteri selulolitik, pembuatan kurva pertumbuhan, dan pengujian aktivitas selulase secara kuantitatif.
Peremajaan Bakteri
Kultur bakteri yang berada dalam stok gliserol disegarkan dalam media Thermus cair (Lampiran 2). Sebanyak dua lup ose kultur bakteri dimasukkan ke dalam 10 mL media, kemudian diinkubasi selama 16 jam pada suhu 55 oC.
Isolasi Koloni Tunggal Bakteri
Thermus yang mengandung substrat CMC 1%. Setelah itu, diinkubasi selama 48 jam pada 55°C.
Seleksi Bakteri Selulolitik
Seleksi dilakukan berdasarkan zona bening yang terbentuk dari setiap koloni. Setiap koloni yang terbentuk dari hasil isolasi dipindahkan ke dalam dua cawan, yaitu cawan replika dan master yang berisi media agar Thermus yang mengandung CMC 1%. Setelah itu, diinkubasi selama 48 jam pada suhu 55 oC. Setelah masa inkubasi berakhir, dilakukan pewarnaan merah kongo 0,1% pada cawan replika untuk memperjelas zona bening yang terbentuk, setelah 15 menit warna dicuci dengan NaCl 1 M (Teather & Wood 1981). Isolat yang dapat memproduksi selulase, yaitu isolat yang membentuk zona bening di sekitar koloni dipilih, kemudian ditumbuhkan dalam media Thermus cair yang mengandung substrat CMC 1%.
Pembuatan Kurva Pertumbuhan
Isolat bakteri selulolitik yang telah ditumbuhkan dalam media Thermus cair diinkubasi pada suhu 55 oC. Setiap 2 jam, dilakukan pengukuran optical density (OD) pada panjang gelombang 600 nm. Pengukuran OD dilakukan sampai terbentuk kurva pertumbuhan.
Uji Aktivitas Selulase Secara Kuantitatif (Miller 1959)
Kultur bakteri yang telah ditumbuhkan dalam media cair yang mengandung substrat CMC 1% disentrifugasi pada kecepatan 10 000 g selama 15 menit. Bagian supernatan digunakan untuk pengujian aktivitas enzim ekstraseluler. Sebanyak 1 mL supernatan dicampur dengan 1 mL CMC 1% pada bufer Mc Ilvaine pH 7,2 (dengan komposisi bufer seperti pada Lampiran 2). Setelah itu, campuran diinkubasi pada 55 °C selama 60 menit. Reaksi tersebut dihentikan dengan penambahan 3 mL pereaksi DNS. Kontrol negatif merupakan enzim yang langsung diinaktifasi dengan DNS.
Campuran divorteks, kemudian di- didihkan selama 15 menit dalam penangas air mendidih, lalu didinginkan terlebih dahulu. Setelah itu, dilakukan pengukuran serapan dengan spektrofotometer pada panjang gelombang 540 nm. Sebagai standar digunakan larutan glukosa dengan konsentrasi 0.01-0.1 mg/mL. Aktivitas enzim dihitung dengan persamaan kurva standar dari larutan enzim yang menghasilkan gula pereduksi.
HASIL DAN PEMBAHASAN
Isolat Bakteri Hasil Peremajaan
Tahapan pertama yang dilakukan sebelum mengisolasi bakteri penghasil selulase adalah peremajaan biakan bakteri yang berada dalam stok gliserol. Peremajaan kultur bakteri ini bertujuan mengaktivasi dan mempersiapkan sel pada fase eksponensial. Hal ini dapat mempercepat fase adaptasi pada saat proses isolasi. Bakteri yang berada dalam fase eskponensial atau tahap propagasi ini mensintesis enzim dan mengatur aktivitasnya sehingga mampu tumbuh lebih efisien dalam kondisi baru. Peremajaan juga memberikan nutrisi baru bagi bakteri sehingga sel-selnya dapat tumbuh sehat.
Peremajaan dilakukan dalam media
Thermus cair. Media Thermus merupakan media spesifik yang digunakan untuk menumbuhkan bakteri termofilik. Media ini mengandung ekstrak khamir, pepton, dan garam-garam mineral berupa amonium sulfat, magnesium sulfat, kalsium klorida, kalium fosfat, dan natrium klorida. Ekstrak khamir merupakan sumber nitrogen, gula, nutrisi organik maupun anorganik sedangkan pepton berfungsi sebagai sumber nitrogen yang menyediakan asam amino dan peptida pendek. Garam-garam mineral berfungsi menyediakan nutrisi anorganik untuk menopang pertumbuhan bakteri.
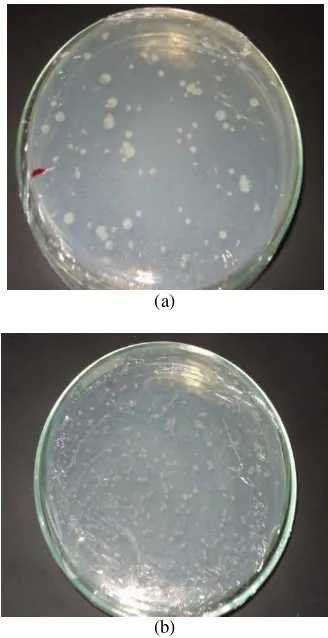
Suhu dan pH yang digunakan untuk menumbuhkan bakteri Kawah Putih dan Kawah Merah adalah 55 oC dan 7. Suhu dan pH ini sesuai dengan kondisi saat bakteri pertama kali diisolasi dari air kawah Gunung Pancar (Harahap 2007). Peremajaan dilakukan selama 16 jam, bakteri yang tumbuh dapat dilihat dari perubahan warna media, yaitu dari bening menjadi keruh. Pertumbuhan bakteri dapat menyebabkan kekeruhan pada medianya.
7
Gambar 3 Hasil inkubasi bakteri Kawah Putih (a), media kontrol (b), dan bakteri Kawah Merah (c) pada media Thermus cair.
Hasil Isolasi Koloni Tunggal Isolat Kawah Putih dan Kawah Merah
Proses isolasi bakteri termofilik tidak jauh berbeda dengan isolasi bakteri lainnya, kecuali kebutuhan terhadap suhu tinggi. Media isolasi harus dijaga agar tidak kering akibat suhu tinggi. Tahapan pertama yang dilakukan untuk mengisolasi bakteri termofilik adalah pemilihan media dan substrat yang sesuai dengan bakteri yang akan diisolasi (Labeda 1990). Tahapan isolasi ini bertujuan mendapatkan bakteri dalam koloni tunggal dari populasi campuran bakteri yang berasal dari Kawah Merah dan Kawah Putih.
Isolasi koloni tunggal bakteri ini dilakukan dalam media Thermus yang mengandung carboxymethyl cellulose (CMC) 1% sebagai substrat. Substrat CMC berfungsi sebagai sumber karbon yang menginduksi sintesis dan sekresi selulase oleh bakteri. Enzim yang memecah makromolekul pada umumnya bersifat ekstraseluler, yaitu setelah diproduksi di dalam sel kemudian dikeluarkan dari sel ke substrat di lingkungannya. Enzim-enzim ekstraseluler umumnya bersifat terinduksi, yaitu produksinya akan meningkat apabila ada substrat yang sesuai. Oleh karena itu, diperlukan substrat yang dapat menginduksi produksi selulase secara optimal.
CMC merupakan substrat terbaik untuk menginduksi sintesis enzim selulolitik ekstraseluler (Alam et al. 2004). Laju degradasi CMC oleh bakteri selulolitik lebih
tinggi dibandingkan dengan substrat lain, yaitu avisel, selobiosa, dan xilan (Ibrahim & El-diwany 2007). Aktivitas komponen selulase yang lain, seperti CMCase dan -glukosidase juga meningkat pada kultur filtrat Aspergillus. niger yang tumbuh pada media yang mengandung CMC (Narasimha et al. 2005).
Carboxymethyl cellulose (CMC)
merupakan substrat sintetik yang berperan sebagai senyawa model selulosa. CMC memiliki banyak daerah amorf sehingga larut dalam air. Selulosa amorf, seperti CMC, dapat dihidrolisis dengan mudah oleh sistem selulase. Menurut Muthuvelayudham & Viruthagiri (2006), selulase mencapai kadar maksimum pada sumber karbon selulosa sintetik, seperti CMC. Konsentrasi CMC yang digunakan adalah 1% karena berdasarkan Narasimha et al. (2005), konsentrasi selulosa 1% merupakan konsentrasi yang optimum untuk produksi selulase.
Isolasi dilakukan dalam media padat dengan teknik cawan tuang. Prinsip dari teknik isolasi ini adalah mengencerkan organisme sehingga individu spesies dapat dipisahkan satu dengan yang lainnya. Setiap koloni yang tampak setelah diinkubasi berasal dari satu sel tunggal. Penumbuhan bakteri hasil pengenceran ini dilakukan melalui proses yang berulang-ulang karena bakteri sulit tumbuh dan beradaptasi dalam kondisi laboratorium. Penumbuhan bakteri ini memerlukan waktu inkubasi yang lebih lama, yaitu 48 jam.
Isolat bakteri kawah putih tampak sebagai koloni-koloni tunggal pada pengenceran 1000x sedangkan isolat kawah merah tampak pada pengenceran 100x. Kedua isolat bakteri tersebut belum terpisah dengan jelas pada pengenceran dibawah 100x. Bakteri isolat kawah merah tidak tumbuh pada pengenceran di atas 100x. Hasil pengenceran kedua isolat ini tampak pada Gambar 4. Seluruh koloni tunggal pada masing-masing kawah memiliki bentuk dan warna yang sama.
Jumlah koloni hasil pengenceran ini tidak harus memenuhi syarat statistik (30-300 koloni) seperti pada teknik kuantitasi mikroba, karena pada tahapan ini yang diinginkan adalah bakteri dalam koloni tunggal, agar sel-selnya berasal dari satu sel induk. Oleh karena itu, koloni bakteri Kawah Putih dan Kawah Merah yang terbentuk hasil pengenceran tersebut dapat digunakan untuk tahap seleksi. Koloni hasil isolasi ditumbuhkan kembali dalam media cair yang mengandung substrat CMC 1%.
(a)
(b)
Gambar 4 Hasil pengenceran isolat Kawah Putih (a) dan Kawah Merah (b).
Hasil Seleksi Bakteri Selulolitik
Tahapan seleksi bertujuan mengetahui bakteri yang memiliki aktivitas selulolitik. Aktivitas tersebut ditunjukkan oleh kemampuan bakteri dalam menghidrolisis substrat CMC. Bakteri yang mampu menghidrolisis CMC akan membentuk zona bening di sekitar koloni. Seleksi ini menggunakan koloni-koloni bakteri yang telah diisolasi.
Setiap koloni dipindahkan ke dalam cawan master dan replika yang berisi media agar Thermus dengan substrat CMC 1%. Koloni bakteri yang dipindahkan ke dalam cawan master langsung dipindahkan ke dalam cawan replika. Dengan demikian, koloni dalam cawan master dan replika merupakan koloni yang sama. Pewarnaan merah kongo dilakukan pada koloni dalam cawan replika untuk memperjelas zona bening yang dihasilkan. Koloni yang berada dalam cawan master digunakan sebagai stok bakteri yang menghasilkan zona bening.
Teknik pewarnaan dilakukan menggunakan pewarna merah kongo 0.1%
sesuai dengan Teather & Wood (1981). Merah kongo berinteraksi kuat dengan ikatan -1,4-glikosidik dalam CMC. Interaksi ini berlangsung secara nonkovalen. Merah kongo dijadikan indikator terjadinya degradasi -D-glukan dalam media agar. Metode ini dipilih karena proses seleksi dapat berlangsung cepat, mudah, dan sensitif. Pewarnaan dengan merah kongo dapat menentukan mikroba selulolitik dalam konsentrasi substrat yang rendah dan dapat mempersingkat waktu inkubasi untuk mendeteksi aktivitas selulase dalam konsentrasi rendah (Teather & Wood 1981). Zona bening yang terbentuk dapat dilihat dengan pencucian menggunakan NaCl 1 M. Merah kongo merupakan garam natrium dari benzidinediazo-bis-1-naphthylamine-4 asam sulfonat (C32H22N6Na2O6S2) sehingga pewarna ini akan larut dan tercuci oleh garam natrium lain, seperti NaCl. Dengan demikian, zona bening yang terbentuk akan tampak jelas.
Seleksi terhadap isolat bakteri kawah putih menunjukkan bahwa dari 60 koloni yang diseleksi terdapat 8 koloni (KP 1, KP 2, KP 3, KP 4, KP 5, KP 6, KP 7, dan KP 10) yang membentuk zona bening sedangkan hasil seleksi terhadap isolat kawah merah hanya 1 koloni (KM 1) yang membentuk zona bening. Zona bening yang terbentuk dari kedua isolat tampak pada Gambar 5. Zona bening menunjukkan zona tempat terputusnya ikatan -1,4-glikosidik yang menghubungkan monomer D-glukosa pada CMC. Berdasarkan hasil tersebut dapat diketahui bahwa isolat Kawah Putih dan Kawah Merah memiliki potensi menghasilkan selulase untuk menghidrolisis ikatan -1,4-glikosidik pada CMC. Selulase yang digunakan untuk proses hidrolisis ini bersifat ekstraseluler. Sifat enzim ekstraseluler terlihat dari zona bening yang terbentuk di sekitar koloni (Lee & Blackburn 1974).
Kemampuan bakteri Kawah Putih dan Kawah Merah membentuk zona bening pada selulosa amorf CMC, menunjukkan adanya komponen selulase, yaitu endo- -1,4-glukanase atau CMCase yang menghidrolisis CMC menjadi selodekstrin, selobiosa, dan glukosa. Enzim ini diproduksi dan disekresikan oleh bakteri karena CMC digunakan sebagai substrat penginduksi. Jenis substrat menentukan jumlah dan komponen selulase yang dihasilkan (Aguiar 2001). Endoglukanase memiliki afinitas yang tinggi terhadap CMC.
diketahui bahwa kawah jumlah bakteri selulolitik dibandingkan dengan kawah & El-diwany (2007), meny hasil seleksi hanya mendapat yang menghasilkan zona menunjukkan bahwa menghasilkan selulase sedik disebabkan oleh rendahnya k organik dalam sampel yang sebagai sumber karbon selulolitik. Oleh karena tahapan penyuburan unt bakteri tersebut.
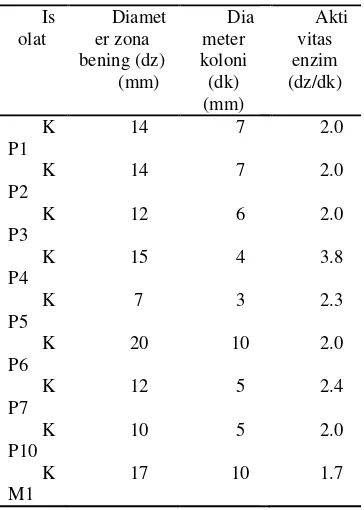
Berdasarkan hasil sel diketahui nilai aktivitas kualitatif. Aktivitas se dinyatakan sebagai nisbah bening dengan diameter kol hasil uji kualitatif (Tabel 1) isolat KP4 memiliki aktivita (3.8) sedangkan isolat aktivitas terendah (1.7). Tuju kawah putih (KP1, KP2, K KP7, dan KP10) memiliki yang hampir sama, yaitu
(a)
(b) : zona bening
Gambar 5 Hasil seleksi ak isolat Kawah Kawah Merah (
KP7 KP6 KP5 KP4 KP10 KM1
putih memiliki tik lebih banyak ah merah. Ibrahim nyebutkan apabila atkan sedikit isolat bening, hal ini bakteri yang ikit. Hal ini dapat kandungan materi ng dapat dijadikan n oleh bakteri itu, dibutuhkan ntuk mengisolasi
seleksi ini dapat s enzim secara selulolitik dapat ah diameter zona oloni. Berdasarkan 1) diketahui bahwa itas enzim tertinggi KM1 memiliki ujuh isolat lain dari KP3, KP5, KP6, ki aktivitas enzim
-2.4.
ing
aktivitas selulolitik h Putih (a) dan h (b).
Bakteri selulolitik isolat Kaw Kawah Merah diuji aktivitas enzi kuantitatif. Uji aktivitas enzim kawah putih dilakukan terhadap ini dipilih karena berdasarkan kualitatif KP4 memiliki aktiv terbesar sedangkan tujuh isolat la aktivitas yang hampir sama. U untuk isolat kawah merah m KM1. Sebelum uji ativitas en kuantitatif, kedua isolat bakteri in pola pertumbuhannya.
Tabel 1 Aktivitas enzim secara ku Is olat Diamet er zona bening (dz) (mm) Dia meter koloni (dk) (mm) K P1
14 7
K P2
14 7
K P3
12 6
K P4
15 4
K P5
7 3
K P6
20 10
K P7
12 5
K P10
10 5
K M1
17 10
Pola Pertumbuhan Bakteri S
Pengamatan pola pertumbu selulolitik dilakukan berdasark turbidimetrik. Bakteri yang tu menghasilkan pertambahan j sehingga dapat diukur berdasarka sel dalam media. Penen pertumbuhan bakteri selulolitik in untuk menentukan waktu akhir fa yang akan digunakan sebagai wa bakteri pada saat pengujian akti selulase secara kuantitatif. Produ mencapai maksimum saat akhir f (Ibrahim & El-diwany 2007).
Perubahan kepekatan sel d diukur setiap 2 jam dengan spek pada panjang gelombang 600 pengukuran setiap 2 jam dipilih se pertumbuhan bakteri yang bertam
KP1
KP2
KP3
9
wah Putih dan zimnya secara untuk isolat ap KP4. Isolat an hasil uji tivitas enzim t lain memiliki Uji aktivitas menggunakan enzim secara ini ditentukan kualitatif Akti vitas enzim (dz/dk) 2.0 2.0 2.0 3.8 2.3 2.0 2.4 2.0 1.7 i Selulolitik
buhan bakteri arkan metode tumbuh akan jumlah sel kan kepekatan entuan pola ini digunakan fase stasioner aktu inkubasi ktivitas enzim duksi selulase r fase stasioner
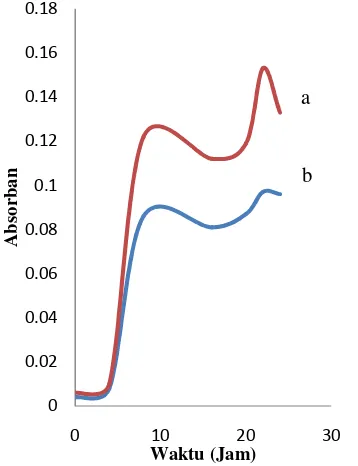
selnya setiap dua jam. Berdasarkan pembuatan kurva pertumbuhan (Gambar 6), bakteri selulolitik isolat KP4 dan KM1 memiliki pola pertumbuhan yang terdiri atas fase adaptasi, pertumbuhan, stasioner, dan fase kematian yang kesemuanya membentuk suatu pola yang sigmoid.
Berdasarkan kurva pertumbuhan dapat diketahui bahwa isolat KP4 dan KM1 memiliki pola pertumbuhan yang sama. Fase adaptasi (lag phase) terjadi pada jam ke-0 hingga jam ke-4. Selama waktu tersebut bakteri baru menyesuaikan diri dengan lingkungan yang baru sehingga sel belum membelah diri. Fase eksponensial (log phase) berada pada kisaran jam 4 hingga jam ke-8. Sel-sel bakteri pada kisaran jam tersebut sangat aktif membelah. Metabolisme sel dan sintesis bahan sel sangat cepat pada fase ini.
Selulase mulai diproduksi saat awal fase eksponensial dan terus meningkat seiring dengan pertumbuhan bakteri (Muthuvelayudham & Viruthagiri 2006; Ibrahim & El-diwany 2007; Maranatha 2008). Enzim ini akan menghidrolisis substrat CMC menjadi molekul yang lebih kecil, seperti selodekstrin, selobiosa, dan glukosa. Komponen tersebut akan dijadikan sebagai sumber karbon untuk pertumbuhannya.
Pertumbuhan bakteri mulai lambat ketika memasuki fase stasioner yaitu dimulai pada jam ke-8. Bakteri pada fase ini pertumbuhannya mulai menurun. Penurunan ini disebabkan adanya bakteri yang mati. Kematian sel disebabkan oleh berkurangnya nutrisi dalam media. Jumlah sel yang mati semakin meningkat sampai terjadi jumlah sel hidup hasil pembelahan sama dengan jumlah sel yang mati. Pertumbuhan bakteri kembali meningkat pada jam ke-22, tetapi peningkatan ini sangat kecil.
Setelah jam ke-22 pertumbuhan bakteri kembali turun. Pengamatan pola pertumbuhan dihentikan pada jam ke-24, yaitu setelah pertumbuhan bakteri menurun kembali, pada jam ini bakteri dianggap telah memasuki fase kematian. Penurunan fase kematian terjadi ketika substrat menurun jumlahnya di bawah konsentrasi yang dibutuhkan untuk menjaga ketahanan sel sehingga sel lisis dan mati (Muthuvelayudham & Viruthagiri 2006).
Fase kematian bakteri tidak dapat diamati sepenuhnya dengan metode turbidimetrik. Metode ini mengukur pertumbuhan berdasarkan kepekatan media dan tidak dapat membedakan bakteri yang hidup dan yang mati. Oleh karena itu, absorbansi sel (kekeruhan media) hanya akan naik dan
stabil. Penurunan sel bakteri setelah fase stasioner dianggap sebagai permulaan fase kematian.
Berdasarkan pola pertumbuhan yang diperoleh dapat diketahui bahwa pertumbuhan bakteri selulolitik isolat KP 4 dan KM 1 dikategorikan memiliki pertumbuhan cepat. Hal ini dapat diamati pada jam ke-24 pola pertumbuhan bakteri sudah memasuki awal fase kematian.
Gambar 6 Pola pertumbuhan bakteri selulolitik isolat KP 4 (a) dan isolat KM1 (b).
Hasil Uji Aktivitas Selulase secara Kuantitatif
Berdasarkan hasil seleksi telah diketahui bahwa isolat Kawah Putih dan Kawah Merah memiliki komponen selulase, yaitu enzim CMCase. Komponen selulase tersebut diuji aktivitasnya secara kuantitatif. Isolat yang diuji aktivitas enzimnya adalah KP 4 dan KM 1. Uji aktivitas enzim dilakukan dengan metode asam dinitrosalisilat atau DNS.
Metode DNS dipilih dalam pengujian ini karena merupakan metode yang umum digunakan untuk pengukuran aktivitas selulase dengan mengukur jumlah gula pereduksi yang terbentuk. Metode ini praktis dan mudah dilakukan untuk pengukuran sampel dalam jumlah banyak. Pengukuran kadar glukosa dengan metode DNS didasarkan pada reaksi asam-3,5- dinitrosalisilat yang direduksi menjadi 3-amino-5-asam nitrosalisilat dalam keadaan
0 0.02 0.04 0.06 0.08 0.1 0.12 0.14 0.16 0.18
0 10 20 30
11
basa. Gugus aldehid akan dioksidasi menjadi gugus karboksil (Miller 1959).
Pengukuran aktivitas enzim dimulai dengan menyiapkan ekstrak enzim kasar. Kultur bakteri diinkubasi selama 22 jam. Berdasarkan kurva pertumbuhan, pada jam ke-22, bakteri memasuki akhir fase stasioner. Produksi selulase mencapai maksimum pada akhir fase stasioner (Altatit 2000; Ibrahim & El-diwany 2007). Setelah masa inkubasi berakhir, kultur bakteri disentrifugasi pada kecepatan 10 000 g selama 15 menit. Sentrifiugasi ini akan memisahkan selulase ekstraseluler (CMCase) yang telah dilepaskan ke dalam media dari komponen sel dan komponen media lainnya. CMCase berada dalam fase supernatan. Supernatan mengandung berbagai macam enzim ekstraseluler yang ikut dilepaskan oleh bakteri. Akan tetapi, dengan induksi menggunakan substrat CMC akan meningkatkan sekresi CMCase untuk menghidrolisis substrat.
Supenatan diuji aktivitas enzimnya dengan metode DNS. Pengujian ini menggunakan larutan CMC 1% sebagai substrat. Nilai pH enzim selama pengukuran dipertahankan dengan menambahkan bufer McIlvaine pH 7.2. Campuran tersebut diinkubasi selama 60 menit pada suhu 55 oC. Masa inkubasi ini diperlukan agar CMC dapat dihidrolisis oleh enzim. Hidrolisis CMC terjadi selama 60-70 menit waktu inkubasi (Chang et al. 2009). Hidrolisis CMC menghasilkan produk berupa glukosa yang diukur jumlahnya dengan metode DNS.
Konsentrasi glukosa ditentukan berdasarkan kurva standar glukosa. Kurva standar dibuat dari larutan standar glukosa dengan konsentrasi 0.01-0.1 mg/mL. Persamaan kurva standar yang diperoleh adalah y = 9.0036x – 0.082 dengan R2= 0.9911 (Lampiran 3).
Satu unit aktivitas enzim dinyatakan sebagai jumlah mikromol glukosa yang dihasilkan oleh satu mL enzim setiap menit (µmol/mL/menit atau U/ mL). Berdasarkan hasil uji aktivitas enzim secara kuantitatif (Tabel 2), diketahui bahwa bakteri selulolitik isolat kawah putih (KP 4) memiliki aktivitas enzim lebih besar (6.11 x 10-3 U/mL) dibandingkan dengan isolat kawah merah (KM1) (1.17 10-3 U/mL). Hal ini sesuai dengan nilai aktivitas enzim secara kualitatif, melalui nisbah diameter zona bening dengan diameter koloni (nisbah diameter KP 4 lebih besar dibandingkan dengan isolat KM 1). Uji aktivitas enzim ini juga dilakukan terhadap
contoh koloni yang tidak membentuk zona bening saat seleksi isolat kawah merah. Hasilnya terlihat bahwa koloni kontrol ini tidak memiliki aktivitas enzim (aktivitas nol). Tabel 2 Hasil uji aktivitas enzim secara
kuantitatif
Isolat Aktivitas enzim (U/mL)
KP 4 6.11x 10-3 KM 1 1.17 x 10-3 Kontrol -
Aktivitas enzim yang dimiliki oleh kedua isolat tersebut masih sangat kecil. Hal ini disebabkan karena isolat bakteri tersebut masih merupakan tipe liar (wild type). Narasimha et al. (2005), menyebutkan bahwa produksi selulase oleh tipe liar Bacillus pumilus, Cellulomonas biazotea, dan
Trichoderma aureoviride dalam media cair tidak akan melebihi 1.5 U/mL. Bacillus spp. yang diisolasi dari kompos Brassica memiliki aktivitas maksimum selulolitiknya sebesar 1.90-2.33 U/mL (Chang et al. 2009).
Faktor lain yang menyebabkan rendahnya aktivitas enzim dalam penelitian ini adalah belum dilakukan optimasi kondisi tumbuh bakteri. Suhu dan pH yang digunakan, yaitu 55 oC dan pH 7, merupakan nilai yang sesuai dengan kondisi saat bakteri pertama kali diisolasi. Kondisi fisik yang digunakan ini bukan kondisi optimum bakteri dalam menghasilkan selulase. Suhu dan pH sangat mempengaruhi aktivitas enzim sehingga bakteri harus ditumbuhkan dalam kondisi pH dan suhu yang optimum untuk memperoleh aktivitas selulase yang optimum.
Aktivitas enzim dipengaruhi oleh pH. Perubahan pH akan menyebabkan daerah katalitik dan konformasi enzim berubah. Perubahan pH juga menyebabkan denaturasi enzim dan menyebabkan hilangnya aktivitas enzim. Suhu juga sangat mempengaruhi dalam reaksi enzimatik. Kecepatan reaksi enzim dan energi kinetik bertambah pada suhu optimum sehingga memperbesar peluang enzim dan substrat bereaksi. Suhu yang lebih tinggi dari suhu optimum menyebabkan perubahan konformasi dan enzim terdenaturasi (Girindra 1993). Substrat dapat berubah konformasi pada suhu yang tidak sesuai sehingga tidak dapat masuk ke dalam sisi aktif enzim.
suhu, periode inkubasi, sumber karbon, dan sumber nitrogen merupakan faktor-faktor penting yang mempengaruhi produksi maksimum selulase dan aktivitas enzimnya. Peningkatan konsentrasi urea dari 2-6 g/L dan pengurangan ekstrak khamir dari 6-4 g/L dapat meningkatkan produksi enzim endoselulase pada bakteri Clostridium thermocopriae (Narasimha et al. 2005).
Berdasarkan hasil penelitian diketahui bahwa isolat Kawah Putih dan Kawah Merah memiliki potensi menghasilkan selulase. Usaha-usaha untuk meningkatkan aktivitas selulase pada isolat Kawah Putih dan Kawah Merah perlu dilakukan sehingga kedua bakteri tersebut dapat diaplikasikan secara luas pada industri berbasis enzim. Usaha-usaha yang dapat dilakukan adalah optimasi kondisi fisik tumbuh bakteri hingga rekayasa genetik dengan manipulasi gen (mutasi genetik).
SIMPULAN DAN SARAN
Simpulan
Isolat bakteri yang berasal dari kawah putih (KP1, KP2, KP3, KP4, KP5, KP6, KP7, dan KP 10) dan kawah merah (KM1) Gunung Pancar berpotensi menghasilkan selulase dan memiliki pola pertumbuhan yang sama. Isolat KP 4 memiliki nisbah diameter zona bening dan diameter koloni yang paling tinggi (3.8) sedangkan KM 1 memiliki nisbah yang terkecil (1.7). Hasil uji aktivitas enzim menunjukkan bahwa isolat KP4 memiliki aktivitas enzim yang lebih besar, yaitu 6.11 x 10-3 U/mL, dibandingkan dengan isolat KM1 yang hanya menghasilkan aktivitas 1.17 10-3 U/mL.
Saran
Perlu dilakukan karakterisasi bakteri selulolitik dan selulase yang dihasilkan oleh isolat KP 4 dan KM 1. Karakterisasi yang dilakukan meliputi optimasi (pH, suhu, sumber karbon, dan sumber nitrogen) yang menunjang pertumbuhan optimum bakteri dan aktivitas selulasenya, serta perlu dilakukan identifikasi isolat.
UCAPAN TERIMAKASIH
Ucapan terimakasih disampaikan kepada DIKTI yang telah mendanai penelitian ini
melalui Program Kreativitas Mahasiswa (PKM) serta kepada Efi Safintri Harahap yang telah menyediakan sumber isolat untuk penelitian ini.
DAFTAR PUSTAKA
Aguiar CL. 2001. Biodegradation of the cellulose from sugarcane bagasse by fungal cellulase. Sci Technol Aliment
3:117-121.
Alam MZ, Manchur MA, Anwar MN. 2004. Isolation, purification, characterization of cellulolytic enzymes produced by Streptomyces omiyaensin. J Biol Sci 10:1647-1653.
Andrade CMMC. Jr pereira N, Antranikian G. 1999. Extremely thermophilic microorganisms and their polymer hidrolytic enzymes. Rev de Microbiol 30:287-298.
Baig MMV, Big MLB, Yasmaen M. 2004. Saccharification of banana agro-waste by cellulolytic enzymes. Afr J Biotechnol 3:447-450.
Brock TD. 1986. Thermophiles: General,
Molecular, and Applied
Microbiology. New York: John Wiley & Son.
Busto MD, Ortega N, Perez-Mateos M. 1995. Induction of –glukosidase in fungal and soil bacterial cultures. Soil Biol Biochem 27:949-954.
Chang CC, Ng CC, Wang CY, Shyu YT. 2009. Activity of celullase from
Thermoactinomycetes and Bacillus
spp. isolated from Brassica waste compost. Sci Agric 66:304-308.
Dianawati A. 2000. Isolasi dan seleksi selulolitik dari lumpur aktif unit pengolahan limbah [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
13
Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Edwards C. 1990. Microbiology of Extreme Environments. New York: Mc Graw-Hill Publishing Company
Fikrinda. 2000. Isolasi dan karakterisasi bakteri penghasil selulase ekstermofilik dari ekosistem air hitam [tesis]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Girindra A. 1993. Biokimia I. Jakarta: Gramedia Pustaka.
Harahap ES. 2007. Amplifikasi gen 16S-rRNA bakteri termofilik dari kawah air panas, Gunung Pancar Bogor [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Heaton K. 2004. Industrial uses of thermophilic cellulase [terhubung berkala]. http:// theguardians.com/Microbiology/gm_ mbm04.htm [ 23 September 2008].
Hidayat N, Padaga MC, Suhartini S. 2006.
Mikrobiologi Industri. Yogyakarta: ANDI.
Ibrahim ASS, El-diwany AI. 2007. Isolation and identification of new cellulases producing thermophilic bacteria from an Egyptian hot spring and some properties of the crude enzyme.
J Appl Sci 1:473-478.
Labeda DP. 1990. Isolation of Biotechnological Organisms from Nature. New York: Mc Graww-Hill.
Lee BH, Blackburn TH. 1974. Cellulase production by a thermophilic
Clostridium spesies. Appl Microbiol
30:346-353.
Lynd LR, Weimer PJ, Zyl WH, Pretorius I. 2002. Microbiol cellulose utilization: fundamentals and biotechnology.
Microbiol Mol Biol Rev 66:506-793.
Maranatha B. 2008. Aktivitas enzim selulase isolat asal Indonesia pada berbagai
substrat limbah pertanian [skripsi]. Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Mayende L, Wilhelmi BS, Pletschke BI. 2006. Cellulase (CMCase) and polyphenol oxidase from thermophilic Bacillus spp. isolated from compost. Soil Biol Biochem
38:2963-2966.
Miller GL. 1959. Use of dinitrosalycilic acid reagent for determination of reducing sugar. Anal Chem 31:426-428.
Muthuvelayudham R, Viruthagiri T. 2006. Fermentative production and kinetics of cellulase protein on Trichoderma reesei using sugarcane bagasse and rice straw. Afric J Biotechnol
5:1873-1881.
Narasimha G et al. 2005. Nutrient effects on productions of cellulolytic enzymes
by Aspergillus niger. J Biotechnol 5: 472-476.
Nam et al. 2004. -Galaktosidase gene of
Thermus thermophilus KNOUC 112 isolated from hot springs of a volcano area in New Zealand: identification of bacteria, cloning, and expression of the gene in
Escherichia coli. J Anim Sci
17:1591-1598.
Pelczar MJ, Chan ECS. 2008. Dasar-Dasar Mikrobiologi. Jilid 1. Hadioetomo RS, Imas T, Tjitrosomo SS, Angka SL, Penerjemah; Jakarta: UI Pr. Terjemahan dari: Elements Of Microbiology.
Ratledge C, Kristiansen B. 2001. Basic Biotechnology. Second Edition. Cambridge: University Pr.
Raza AM, Shafiq-Ur-Rehman. 2008. Production and characterization of endo- -1,4-glukanase from thermofilic fungus. J Biotechnol 8: 3297-3302.
Sari WW. 2008. Karakterisasi selulase bakteri asal tanah pertanian Jawa Tengah dan Jawa Barat [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Shimada K, Karita S, Sakka K, Ohmiya K. 1994. Cellulase, Xylanase, and Their Genes from Bacteria. New York: Marcel Dekker Inc.
Sinegani AAS, Emtiazi G. 2006. The relative effects of some elements on DNS method in cellulase assay. J Appl Sci Environ 10: 93-96.
Susanti HE. 2007. Isolasi dan optimasi flokulasi bakteri penghasil bioflokulan dari sumber perairan di daerah Bogor [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Suyono et al. 2008. Isolasi bakteri termofilik asam laktat dari kawah air panas Gunung Pancar Bogor [laporan akhir PKM]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Teather RM, Wood PJ. 1981.Use of congo red-polysaccharide interactions in enumeration and characterization of cellulolytic bacteria from the bovine rumen. Appl Environ Microbiol
16
Lampiran 1 Tahapan penelitian
Isolasi koloni tunggal bakteri
Seleksi bakteri selulolitik dengan
metode
screening plate
(zona bening)
Peremajaan bakteri
Pembuatan kurva pertumbuhan
bakteri selulolitik
Uji aktivitas CMCase
dengan metode DNS
18
Lampiran 2 Pembuatan media, bufer, dan pereaksi DNS
1.
Media cair
Thermus
Sebanyak 0.1 gram (NH)4SO4, 0.25 gram MgSO4.7H2O, 0.125 gram
CaCl2.2H2O, 0.3 gram KH2PO4, 1 gram NaCl, 2 gram ekstrak khamir, dan
4 gram pepton dilarutkan dalam 1 L akuades, lalu dihomogenkan dengan
memanaskannya. Nilai pH media dijadikan 7. Sebanyak 10 mL larutan
tersebut dipipet ke dalam labu Erlenmeyer 250 mL, lalu ditutup dengan
kapas dan kertas alumunium. Media disterilisasi dengan autoklaf pada 121
oC, 1 atm, selama 15 menit.
2.
Pembuatan buffer Mc llvaine
Sebanyak 13.8 mL asam sitrat 0.1 M dicampur dengan 86.2 mL
natrium fosfat 0.2 M dan 0.2% natrium asida. Nilai pH buffer dijadikan
7.2. Larutan disimpan pada suhu 4
oC.
3.
Pembuatan pereaksi DNS
Lampiran 3 Pembuatan kurva standar glukosa
Larutan
Volume
standar (µL)
Volume
buffer (µL)
Konsentra
si
(mg/mL)
Absorbans
Blanko
0
1000
0.0000
0.000
1
10
990
0.0100
0.018
2
20
980
0.0200
0.048
3
30
970
0.0300
0.161
4
40
960
0.0400
0.262
5
50
950
0.0500
0.319
6
60
940
0.0600
0.463
7
70
930
0.0700
0.529
8
80
920
0.0800
0.638
9
90
910
0.0900
0.781
10
100
900
0.1000
0.829
Contoh perhitungan (standar 1):
Konsentrasi standar awal = 1mg/mL
M1 x V1 = M2 x V2
1 (mg/mL) x V1= 0.0100 (mg/mL) x 1 mL
V1 = 0.01 mL = 10 µL
20
Lampiran 4 Perhitungan konsentrasi glukosa sampel
Sampel
Absorabans
[Glukosa]
(mg/mL)
KP4
0.154
0.0262
KP4 (-)
0.055
0.0152
KM1
0.039
0.0134
KM1 (-)
0.020
0.0113
Kontrol (-)
0.000
0.0000
Keterangan: KP4 (-) dan KM1 (-) = Kontrol negatif isolat KP4 dan KM1
Kontrol (-) = Koloni yang tidak membentuk zona bening
Contoh perhitungan (sampel KP4):
Dari persamaan garis kurva standar glukosa
y= 9.0036 x – 0.082
0.154 = 9.0036 x – 0.082
0.154 + 0.082 = 9.0036x
0.236 = 9.0036 x
x = 0.0262
x = [glukosa] = 0.0262 mg/mL
y = 9.0036x - 0.082 R² = 0.9911
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9
0 0.02 0.04 0.06 0.08 0.1 0.12
A
b
sor
b
a
n
s
Lampiran 5 Perhitungan aktivitas enzim CMCase
Unit enzim= µmol produk/mL enzim/menit
= U/mL
Contoh : aktivitas enzim CMCase KP4
Aktivitas enzim=
xFP
xBMglukosa
menit
T
to
Glukosa
topt
Glukosa
)
(
]
[
]
[
−
X
10
3µmol
Keterangan : [Glukosa]topt : konsentrasi glukosa inkubasi 1 jam
[Glukosa]to : konsentrasi glukosa inkubasi 0 jam
FP
: faktor pengenceran
T (menit)
: waktu inkubasi
FP = volume total / volume enzim
= 3 mL / 0.5 mL
= 6
AE =
6
180
60
0152
.
0
0262
.
0
x
×
−
x 10
3µmol