DWI NINGSIH SUSILOWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi berjudul Analisis Komunitas dan Fungsi Bakteri Rhizosfer Tanaman Padi pada Gradien Salinitas Tanah Pesisir adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2015 Dwi Ningsih Susilowati
Rhizosfer Tanaman Padi pada Gradien Salinitas Tanah Pesisir. Dibimbing oleh ANTONIUS SUWANTO, I MADE SUDIANA, dan NISA RACHMANIA MUBARIK.
Salinitas merupakan salah satu cekaman abiotik yang mempengaruhi pertumbuhan dan produktivitas tanaman padi, terutama pada lahan-lahan sawah di daerah pesisir. Cekaman garam menyebabkan ketidakseimbangan ionik dan osmotik di dalam sel tanaman. Berbagai mikrob diketahui berasosiasi dengan rhizosfer tanaman padi dan beberapa diantaranya bermanfaat bagi tanaman. Penelitian ini dilakukan untuk mengetahui informasi keragaman dan kelimpahan struktur komunitas, serta karakter fungsional bakteri rhizosfer tanaman padi yang ditanam di tanah pesisir dengan tiga taraf salinitas yang berbeda dari taraf sangat rendah hingga taraf moderat. Sampel tanah diambil dari dua jenis tanah yang berbeda dari dua lokasi di Jawa Barat, yaitu tanah marin dari Subang dan fluvio-marin dari Indramayu. Populasi dan struktur komunitas bakteri rhizosfer dianalisis pada saat tanaman padi memasuki fase primordia dan pembungaan dengan metode kultur (culture-dependent) dan metode nonkultur (culture independent). Total populasi bakteri rhizosfer berada di kisaran 104-106 CFU / g tanah, dan secara statistik total populasi pada tanah salin moderat tidak berbeda secara signifikan dibandingkan dengan tanah salin rendah dan sangat rendah. Keragaman jenis bakteri rhizosfer dari tanah salin moderat lebih rendah daripada tanah salin rendah dan sangat rendah. Genus Bacillus (filum Firmicutes) ditemukan pada berbagai taraf salinitas tanah baik moderat, rendah, dan sangat rendah, seperti B. subtilis complex. Sekelompok bakteri dari filum Proteobacteria dan Actinobacteria juga diperoleh selain filum Firmicutes pada metode kultur. Analisis pustaka klon menunjukkan adanya lebih banyak filum (11 filum) yang diperoleh dari beragam bakteri rhizosfer tanaman padi pada tanah pesisir Subang dan Indramayu pada fase primordia antara lain Proteobacteria, Acidobacteria, Chloroflexi, Cyanobacteria, Nitrospirae, Gemmatimonadetes, Bacteriodetes, Aquificae, Actinobacteria, Firmicutes, dan Elusimicrobia dibandingkan metode kultur. Firmicutes mendominasi struktur komunitas bakteri rhizosfer tanaman padi pada metode kultur, sedangkan dengan metode nonkultur diketahui Proteobacteria dan Acidobacteria lebih dominan. Metode nonkultur juga memberikan informasi yang jelas adanya sejumlah bakteri dari laut yang terbawa arus intrusi (bakteri transient) sehingga berada bersama-sama dengan bakteri indigenos pada tanah sawah pesisir. Ada kecenderungan bahwa salinitas tanah memiliki efek negatif pada kelimpahan Betaproteobacteria dan efek positif pada kelimpahan Alphaproteobacteria dan Deltaproteobacteria. Hal ini menyebabkan nilai indeks keragaman spesies yang cukup tinggi pada tanah dengan taraf salinitas moderat, dan tidak berbeda secara nyata antara ketiga taraf salinitas, baik dengan metode kultur dan metode nonkultur. Isolat-isolat bakteri rhizosfer yang diperoleh pada metode kultur diketahui memiliki karakter fungsional sebagai penambat N2, pelarut fosfat, pendegradasi selulosa, dan penghasil IAA. Hal ini
tanaman, jumlah anakan efektif, bobot kering tajuk, dan bobot kering akar) pada fase primordia dan fase pembungaan, demikian juga dengan parameter komponen hasil panen (jumlah malai, panjang malai, biji isi per malai, bobot total biji per tanaman, dan bobot kering bulir padi).
of Rhizosphere Bacteria associated with Rice Plant Grown on Coastal Soil with Different Levels of Salinity. Supervised by ANTONIUS SUWANTO, I MADE SUDIANA, and NISA RACHMANIA MUBARIK.
Salinity is one of the abiotic stresses affect the growth and productivity of rice plant, especially on paddy field in the coastal areas. Salt stress induces ionic and osmotic imbalance inside plant cells. Various microbes associated with the rhizosphere of rice plants and some are beneficial to the plants. This research was conducted to find out information on the diversity and abundance of community structure and functional character of rhizosphere bacteria associated with rice plant grown on coastal soil with three different levels of salinity from very low saline to moderate saline. The soil samples collected from two different types of soil were taken from two locations in West Java, namely marine soils from Subang and fluvio-marine soils from Indramayu. Community structures and population of the rhizosphere bacteria were monitored during primordial and flowering plant stages in greenhouse experiments using both culture-dependent approaches and molecular methods (clonal 16S rRNA gene sequence analysis). 16S rDNA sequences obtained by direct cloning from soil DNA and isolated bacterial populations were analyzed and compared with each other. Total population of the rhizosphere bacteria were in the range of 104-106 CFU/g soil. Population number in moderate saline soil was not significant compared to the low and non saline soil. The diversity of rhizosphere bacteria from the moderate saline soil were lower than from low and non saline soil of the control pot.
Bacillus (phylum Firmicutes) found in a variety of soil salinity level either moderate, low, and very low, such as B. subtiliscomplex. A group of bacteria of the phylum Proteobacteria and Actinobacteria also be obtained in addition to the phylum Firmicutes from the culture dependent method. Clone library analysis revealed that the rhizosphere bacterial community structure of the rice plant on the marine soil from Subang and fluvio-marin from Indramayu in a primordial plant stage consisted of 11 phyla (Proteobacteria, Acidobacteria, Chloroflexi, Cyanobacteria, Nitrospirae, Gemmatimonadetes, Bacteriodetes, aquificae, Actinobacteria, Firmicutes, and Elusimicrobia). Firmicutes dominated the rhizosphere bacterial community structure of rice plants in culture dependent methods, whereas Proteobacteria and Acidobacteria dominated in culture independent methods. There is a tendency that the salinity of the soil has a negative effect on the abundance of Betaproteobacteria and positive effect on the abundance of Alphaproteobacteria and Deltaproteobacteria. Culture independent methods provide accurate information about the origin of a number of marine bacteria coexisting with indigenous bacteria on coastal wetland. This causes the index value of species diversity is high enough on the soil with moderate salinity level, but not significantly different among the three salinity level, either by culture dependent and culture independent methods. Rhizosphere bacterial isolates obtained in culture dependent methods are known to have a functional character like N2 fixing, phosphate solubilizing, cellulose degrading, and IAA
caused a reduction on plant growth parameters (plant height, number of effective tillers, shoot dry weight, root dry weight) at the primordial and yield components of rice (number of panicles, panicle length, percentage seeds per panicle, seed weight per plant, and weight of rice grains).
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
DWI NINGSIH SUSILOWATI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Mikrobiologi (MIK)
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Prof Dr Aris Tri Wahyudi, MSi Dr Ir Happy Widiastuti, MSi
Penguji pada Ujian Terbuka: Prof Dr Aris Tri Wahyudi, MSi Dr Ir Happy Widiastuti, MSi
NIM : G 361100041
Disetujui oleh Komisi Pembimbing
Prof Dr Antonius Suwanto, MSc Ketua
Prof (R) Dr I Made Sudiana, MSc Dr Nisa Rachmania Mubarik, MSi Anggota Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana Mikrobiologi
Prof Dr Anja Meryandini,MS Dr Ir Dahrul Syah, MScAgr
Puji dan syukur penulis panjatkan ke hadirat Allah SWT karena dengan perkenan-Nyalah penulis dapat menyelesaikan karya ilmiah dengan judul: Analisis Komunitas dan Fungsi Bakteri Rhizosfer Tanaman Padi pada Gradien Salinitas Tanah Pesisir. Penelitian ini dilakukan sejak bulan Juni 2012 hingga Juni 2014 di Laboratorium Ekologi dan Fisiologi C, Bidang Mikrobiologi, Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI) dan di Laboratorium Mikrobiologi dan rumah kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB. BIOGEN). Terima kasih penulis sampaikan kepada Prof Dr Antonius Suwanto, MSc., Prof (R) Dr I Made Sudiana, MSc., dan Dr Nisa Rachmania Mubarik, MSi. selaku pembimbing atas saran-saran yang diberikan untuk kesempurnaan substansi dan format disertasi ini. Disampaikan juga terima kasih kepada Dr Ir Iman Rusmana, MSi., Dr Puspita Lisdiyanti, MSc., Dr Ir Suryo Wiyono, MScAgr., Dr Ence Darmo Jaya Supena, MS., Prof Dr Aris Tri Wahyudi, MSi., dan Dr Ir Happy Widiastuti, MSi. atas kritik dan saran yang diberikan selama pelaksanaan ujian kualifikasi, ujian tertutup dan terbuka . Demikian juga terima kasih kepada Bapak Rofiq dan Bapak Wahyu staf Balai Besar Penelitian dan Pengembangan Sumber Daya Lahan Pertanian (BBSDLP) atas ijin akses file data peta tanah dan sebaran tanah sawah di daerah Jawa Barat; Shigeto Otsuka, PhD, Dr Uus Saefulloh, Dr Iman Hidayat, Prof (R) Dr Ika Mariska dan Bapak Deddy Erfandi atas kesempatan diskusinya selama ini, serta Prof Dr Anja Meryandini, MS. dan Dr Gayuh Rahayu, MSc yang senantiasa mengingatkan penulis untuk segera menyelesaikan studinya. Ungkapan terima kasih juga disampaikan kepada suami dan anak-anak tercinta, kedua orang tua dan saudara-saudara atas kesabaran dan dukungannya selama ini, Pak Karmin, Pak Holil, dan Pak Uci (alm) yang banyak membantu selama pengambilan sampel tanah, teman-teman seperjuangan (Mbak Atit, Mbak Santi, Mbak Nani, dan Mbak Ernin) atas kesetiakawanan dan kerjasamanya, serta semua pihak yang secara tidak langsung membantu kelancaran penelitian dan proses penyusunan disertasi.
Sumber dana penelitian ini diperoleh dari proyek SATREP LIPI-JST dan Dana DIPA APBN TA. 2013 dan 2014 (PJ. Ir Yadi Suryadi, MSc). Penulis sangat berterima kasih telah dilibatkan dalam proyek SATREP LIPI-JST, sehingga penelitian ini dapat diselesaikan. Demikian juga kepada Bapak Yadi, penulis sampaikan terima kasih atas perhatian dan kerjasamanya dalam pengadaan sejumlah bahan-bahan kimia yang diperlukan untuk penelitian ini.
Semoga hasil penelitian ini dapat bermanfaat bagi pengembangan ilmu pengetahuan dan dapat digunakan sebagai informasi awal untuk menangani permasalahan lahan-lahan pertanian di daerah pesisir Indonesia dari sudut pandang mikrobiologi.
Bogor, Juli 2015
DAFTAR TABEL iii
DAFTAR GAMBAR. iii
DAFTAR LAMPIRAN iv
1 PENDAHULUAN Latar Belakang 1
Perumusan Masalah 2 Tujuan Penelitian 2 Hipotesis Penelitian 3 Manfaat Penelitian 3 Kebaruan Penelitian (Novelty) 3
Ruang Lingkup Penelitian 4 2 TINJAUAN PUSTAKA Salinitas Tanah 5 Dampak Salinitas Tanah terhadap Tanaman Padi 6 Komunitas dan Aktivitas Mikrob Rhizosfer 6
Dampak Salinitas Tanah terhadap Mikrob Rhizosfer 8
Metode Identifikasi Komunitas Mikrob 9
3 METODE Tempat dan Waktu Penelitian 10
Bahan dan Alat Penelitian 10
Prosedur Analisis 11
4 HASIL DAN PEMBAHASAN Sifat-sifat Fisik, Kimia, dan Mikrobiologi Tanah Sawah Pesisir Subang dan Indramayu 17
Analisis Komunitas dan Fungsi Bakteri Rhizosfer Tanaman Padi pada Gradien Salinitas Tanah Pesisir dengan Metode Kultur 22
Analisis Komunitas Bakteri Rhizosfer Tanaman Padi pada Fase Primordia pada Gradien Salinitas Tanah Pesisir dengan Metode Nonkultur 36
Perbandingan Metode Kultur dan Nonkultur untuk Analisis Komunitas Bakteri Rhizosfer Tanaman Padi pada Fase Primordia pada Gradien Salinitas Tanah Pesisir 44
Hubungan antara Pertumbuhan dan Produktivitas Tanaman Padi dengan Struktur Komunitas Bakteri Rhizosfer pada Gradien Salinitas Tanah Tanah Pesisir 48
5 SIMPULAN DAN SARAN Simpulan 56
Saran 56
DAFTAR PUSTAKA 58
LAMPIRAN 70
tanah pesisir di bak percobaan 23 2 Indeks keragaman spesies bakteri rhizosfer tanaman padi pada tanah pesisir Subang dan Indramayu pada fase primordia dan pembungaan dengan metode kultur 27 3 Keragaman fungsional bakteri rhizosfer tanaman padi dari tanah
pesisir Subang dan Indramayu 31 4 Indeks keragaman bakteri rhizosfer tanaman padi pada tanah pesisir Subang dan Indramayu pada fase primordia dengan metode
nonkultur 43
5 Pengaruh taraf salinitas tanah terhadap parameter pertumbuhan tanaman padi pada fase primordia (35 hari setelah pindah tanam)
di bak percobaan 51 6 Analisis korelasi Pearson antara salinitas dan parameter pertumbuhan tanaman padi pada fase primordia 51 7 Pengaruh taraf salinitas tanah terhadap parameter pertumbuhan tanaman padi pada fase pembungaan (70 hari setelah pindah tanam) di bak
percobaan 51
8 Analisis korelasi Pearson antara salinitas dan parameter pertumbuhan tanaman padi pada fase pembungaan 52 9 Pengaruh taraf salinitas tanah terhadap parameter komponen hasil tanaman padi pada 105 hari setelah pindah tanam di bak percobaan 52 10 Analisis korelasi Pearson antara salinitas dan parameter komponen
hasil tanaman padi 53
DAFTAR GAMBAR
1 Bagan alur penelitian 4
2 Populasi bakteri tanah (log CFU/gram tanah) dari dua tanah sawah pesisir Subang and Indramayu, dengan taraf salinitas moderat,
rendah, dan sangat rendah 19 3 Amplifikasi 16S rRNA genom bakteri tanah dari daerah pesisir
Indramayu 20
4 Jenis-jenis spesies bakteri yang dominan pada tanah sawah pesisir Subang (A) dan Indramayu (B) 21 5 Populasi bakteri rhizosfer tanaman padi varietas Mekongga
(log CFU/gram tanah) pada fase primordia dan fase pembungaan dari tanah marin-Subang dan fluviomarin-Indramayu pada taraf
salinitas moderat, rendah, dan sangat rendah 22 6 Jenis-jenis spesies bakteri rhizosfer yang dominan pada tanaman padi pada fase primordia (35 hari setelah pindah tanam) pada tanah
sawah pesisir Subang (A) dan Indramayu (B) 24 7 Jenis-jenis spesies bakteri rhizosfer yang dominan pada tanaman padi pada fase pembungaan (70 hari setelah pindah tanam) pada tanah
sejumlah bakteri rhizosfer tanaman padi varietas Mekongga pada
tanah sawah pesisir Subang dan Indramayu 37 10 Hubungan filogenetik bakteri rhizosfer tanaman padi dari pustaka
klon 16S rRNA dari tanah sawah pesisir Subang dengan salinitas moderat yang memiliki kesamaan homologi dengan sekuen-sekuen yang diisolasi dari lingkungan salin atau alkali 40 11 Hubungan filogenetik bakteri rhizosfer tanaman padi dari pustaka
klon 16S rRNA dari tanah sawah pesisir Indramayu dengan salinitas moderat yang memiliki kesamaan homologi dengan sekuen-sekuen yang diisolasi dari lingkungan salin atau alkali 41 12 Komposisi relatif klon-klon pada taraf filum dari pustaka klon 16S
rRNA tanah pesisir Subang 46 13 Komposisi relatif klon-klon pada taraf filum dari pustaka klon 16S
rRNA dari tanah pesisir Indramayu 47 14 Parameter pertumbuhan tanaman padi varietas Mekongga, (A) tinggi
tanaman dan (B) jumlah anakan, pada fase primordia dan pembungaan pada tanah marin-Subang dan fluviomarin-Indramayu, dari tiga taraf salinitas moderat, rendah dan sangat rendah 49 15 Parameter pertumbuhan tanaman padi varietas Mekongga, (A) bobot
kering tajuk dan (B) bobot kering akar, pada fase primordia dan pembungaan pada tanah marin-Subang dan fluviomarin-Indramayu, dari tiga taraf salinitas moderat, rendah, dan sangat rendah 50 16 Parameter komponen hasil tanaman padi varietas Mekongga pada
umur 105 hari setelah tanam, (A) jumlah malai, (B) panjang malai, dan (C) persentase biji isi per malai, dari tanah marin-Subang dan fluviomarin-Indramayu, dari tiga taraf salinitas moderat, rendah,
dan sangat rendah 54 17 Parameter komponen hasil tanaman padi varietas Mekongga pada
umur 105 hari setelah pindah tanam, (A) bobot total biji pertanaman (g/tanaman) dan (B) bobot kering bulir padi (g/tanaman), dari tanah marin-Subang dan fluviomarin-Indramayu, dari tiga taraf salinitas moderat, rendah, dan sangat rendah 55
DAFTAR LAMPIRAN
1 Peta lokasi sebaran sawah daerah Jawa Barat (A Subang dan
B Indramayu) 70
2 Karakteristik fisikokimia tanah sawah pesisir Desa Patimban, Kecamatan Pusaka Negara, Kabupaten Subang, dan Desa Eretan
Kulon, Kecamatan Kandanghaur, Kabupaten Indramayu, Jawa Barat 71 3 Deskripsi padi varietas Mekongga 72 4 Jenis-jenis bakteri tanah dari lahan sawah daerah pesisir Subang
6 Jenis-jenis bakteri rhizosfer tanaman padi pada gradien salinitas tanah pesisir Subang dan Indramayu yang diisolasi pada fase
pembungaan 77
7 Dinamika keberadaan bakteri rhizosfer tanaman padi pada fase primordia dan pembungaan pada gradien salinitas tanah pesisir
Subang dan Indramayu 79 8 Jenis-jenis bakteri rhizosfer hasil pustaka klon 16S rRNA 81 9 Afiliasi filogenetik dan kelimpahan bakteri rhizosfer tanah pesisir
Subang 93
10 Afiliasi filogenetik dan kelimpahan bakteri rhizosfer tanah pesisir
Latar Belakang
Salinitas adalah fitur alami dari ekosistem di daerah kering dan semi kering dengan tingkat evapotranspirasi yang tinggi, sehingga berdampak pada terjadinya akumulasi garam pada permukaan tanah. Lebih dari 800 juta hektar lahan di seluruh dunia dipengaruhi oleh garam (FAO 2008). Salinitas tanah menunjukkan variasi temporal dan spasial yang tinggi bergantung pada iklim, elevasi, drainase, tekstur tanah, dan faktor-faktor terkait lainnya (Tripathi et al. 2006, Tripathi et al.
2007). Ada tiga klasifikasi tanah yang digunakan untuk menggambarkan akumulasi garam di dalam tanah. Tanah salin mengandung jumlah garam terlarut yang tinggi, terutama Ca2+, Mg2+, K+, dan Na+, garam Cl-, NO3-, SO4-2, dan CO3-3;
tanah sodik didominasi oleh garam Na+. Tanah salin-sodik memiliki garam Ca2+, Mg2+, dan K+ serta Na+ yang tinggi
Salinitas tanah juga dihadapi di beberapa lahan sawah daerah pesisir di Indonesia. Sekitar 40.3% (3.25 juta ha) dari luas lahan sawah di Indonesia (8.1 juta ha) berada di Pulau Jawa dan tersebar di daerah pesisir Pantai Utara (Pantura). Beberapa daerah sentra produksi padi di wilayah pesisir Jawa Barat seperti kabupaten: Bekasi, Karawang, Subang, Indramayu, dan Cirebon memiliki kontribusi paling tinggi terhadap produksi beras nasional (BPS 2011). Salah satu permasalahan lahan sawah di wilayah tersebut adalah penciutan lahan akibat genangan sebagai dampak peningkatan permukaan air laut dan degradasi serta penurunan produktivitas lahan akibat salinitas. Tanaman padi tergolong ke dalam tanaman yang toleran salin pada kisaran salinitas sebesar 0.9 – 1.6 dS/m dan sangat peka terhadap salinitas pada awal fase bibit. Dengan demikian salinitas merupakan ancaman serius terutama pada lahan sawah daerah pesisir bagi keberlanjutan agroekosistem.
Di Pantura Jawa Barat, dampak kenaikan muka air laut terhadap penurunan produksi padi akibat salinitas terlihat secara nyata di Indramayu. Hasil penelitian menunjukkan tingkat salinitas di Indramayu, Jawa Barat berstatus sedang sampai tinggi masing-masing pada kedalaman 0-30 cm dan 30-70 cm. Kabupaten Karawang dan Subang juga mengalami penurunan produksi beras sekitar 300.000 ton akibat genangan (Boer et al. 2009, Boer et al. 2011). Menurut Suriyan dan Chalermpol (2009), perluasan intrusi air laut mengakibatkan salinisasi tanah, sehingga bersifat toksik bagi tanaman, dan dapat menghambat pertumbuhan dan produksinya. Apabila salinitas meningkat di atas 2 dS/m maka hasil akan menurun sekitar 10% untuk setiap kenaikan 1 dS/m (Grattan et al.
2002).
Namun, sifat mikrobiologi pada rhizosfer tanaman padi pada fase primordia pada lahan sawah di lingkungan salin dan toleransi tanaman terhadap cekaman salinitas yang didapatkan dari bakteri rhizosfer masih jarang dipelajari. Penelitian dengan basis adaptasi mengenai struktur komunitas mikrob rhizosfer tanaman padi pada lahan-lahan sawah di daerah pesisir terkait dengan permasalahan salinitas tanah belum pernah dilaporkan di Indonesia. Keberadaan mikrob rhizosfer banyak terlibat dalam berbagai siklus biogeokimia seperti mineralisasi, siklus hara, dan agregasi tanah, sehingga keragaman mikrob merupakan komponen penting untuk menopang fungsi ekosistem.
Beberapa peneliti melakukan studi komunitas mikrob dengan berbagai pendekatan, baik dengan metode kultur maupun nonkultur. Nelson dan Mele (2007) telah mengamati adanya perubahan struktur komunitas mikrob rhizosfer tanaman gandum sebagai respon terhadap peningkatan konsentrasi boron dan NaCl menggunakan BIOLOG dan analisis Terminal Restriction Fragment Length Polymorphism (T-RFLP). Perubahan terjadi secara nyata pada struktur komunitas bakteri penambat N2, namun tidak demikian pada bakteri pengoksidasi ammonia.
Selanjutnya Ibekwe et al. (2010) mengamati struktur komunitas bakteri rhizosfer tanaman ketimun sebagai efek terhadap boron, pH, dan NaCl tanah menggunakan metode kultur dan nonkultur dengan Denaturing Gradien Gel Electrophoresis
(DGGE). Metode kultur kurang memadai untuk menganalisis komunitas mikrob pada lingkungan alaminya, sehubungan dengan adanya sejumlah besar bakteri yang belum dapat dikulturkan (Amann et al. 1995). Analisis molekuler seperti kloning dan sekuensing gen 16S rRNA juga dapat digunakan untuk memonitor struktur komunitas mikrob tanpa kultivasi (Feiske et al. 1998, Dunbar et al. 1999, Ramirez-Saad et al. 2000).
Perumusan Masalah
Adanya intrusi air laut ke lahan sawah di wilayah pesisir dapat menghambat pertumbuhan tanaman padi dan mempengaruhi penurunan produktivitas lahan tersebut. Agroekosistem di daerah pesisir ini merupakan ekosistem yang dinamis dan memiliki kekayaan mikrob yang potensial bagi pertumbuhan tanaman dan kesuburan tanah. Namun, apakah salinitas tanah akan memiliki kecenderungan menurunkan populasi, diversitas, dan kelimpahan pada struktur komunitas bakteri rhizosfer tanaman padi. Apabila terjadi hal yang demikian, kecenderungan seperti apa yang akan terjadi pada struktur komunitas mikrob rhizosfer tanaman padi? Dan bagaimana pengaruh perubahan tersebut pada parameter pertumbuhan tanaman padi?
Tujuan Penelitian
salinitas tanah pesisir dengan metode kultur dan nonkultur, 2) membandingkan analisis komunitas bakteri rhizosfer tanaman padi pada salah satu fase pertumbuhan tanaman padi yang ditanam pada gradien salinitas tanah pesisir dengan metode kultur dan nonkultur, dan 3) mempelajari pengaruh gradien salinitas lahan sawah daerah pesisir pada struktur komunitas bakteri rhizosfer dan dihubungkan dengan parameter pertumbuhan dan komponen hasil tanaman padi varietas Mekongga
Hipotesis Penelitian
Hipotesis pada penelitian ini ialah (1) semakin rendah salinitas tanah pesisir, maka semakin tinggi keragaman struktur komunitas bakteri rhizosfer tanaman padi; (2) adanya perbedaan gradien salinitas tanah pesisir mempengaruhi kelimpahan jenis bakteri rhizosfer tanaman padi; dan (3) semakin rendah salinitas tanah dan semakin beragam struktur komunitas bakteri rhizosfer tanaman padi akan semakin baik kualitas pertumbuhan tanaman padi.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan informasi dasar mengenai keragaman dan kelimpahan struktur komunitas bakteri rhizosfer tanaman padi pada gradien salinitas tanah pesisir. Data yang diperoleh dapat menjadi basis data untuk pengembangan strategi dan teknologi adaptasi untuk penyesuaian kegiatan dan teknologi dengan kondisi lingkungan yang disebabkan oleh adanya perluasan intrusi air laut. Upaya adaptasi dipandang sebagai langkah penyelamatan agar ketahanan pangan dan sasaran pembangunan pertanian dapat dicapai. Upaya adaptasi dilakukan melalui pengembangan pertanian yang toleran terhadap permasalahan lingkungan saat ini dan di masa yang akan datang.
Kebaruan Penelitian (Novelty)
Ruang Lingkup Penelitian
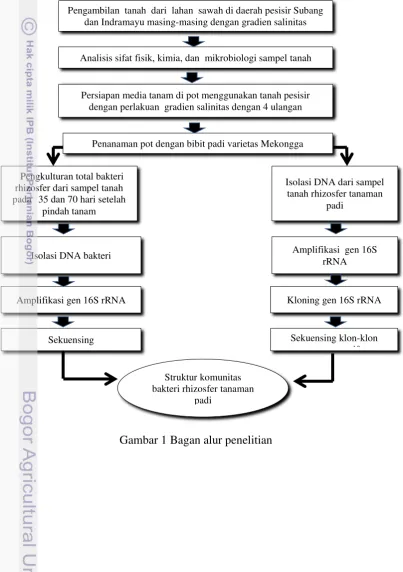
Penelitian ini mengungkap keragaman dan kelimpahan struktur komunitas bakteri rhizosfer tanaman padi pada gradien salinitas tanah pesisir dan dikaitkan dengan pertumbuhan dan produktivitas tanaman padi dengan metode kultur dan nonkultur seperti ditunjukkan pada alur penelitian berikut ini (Gambar 1).
Gambar 1 Bagan alur penelitian
Pengambilan tanah dari lahan sawah di daerah pesisir Subang dan Indramayu masing-masing dengan gradien salinitas
Analisis sifat fisik, kimia, dan mikrobiologi sampel tanah
Penanaman pot dengan bibit padi varietas Mekongga
PPengkulturan total bakteri rhizosfer dari sampel tanah pada
35 dan 70 hari setelah
pindah tanam
Isolasi DNA dari sampel tanah rhizosfer tanaman
padi
Isolasi DNA bakteri Amplifikasi gen 16S
rRNA
Amplifikasi gen 16S rRNA Kloning gen 16S rRNA
Sekuensing klon-klon representatif Sekuensing
Struktur komunitas bakteri rhizosfer tanaman
padi tanaman padi
2 TINJAUAN PUSTAKA
Salinitas Tanah
Salinitas menunjukkan kadar garam yang terlarut di dalam air atau larutan tanah yang dinyatakan dalam satuan miligram per liter (mg/L) atau disebut total garam terlarut (Total Dissolved Salts/TDS). Pengukuran TDS secara tidak langsung biasanya dilakukan dengan pendekatan electrical conductivity (EC). Satuan Internasional (SI) untuk salinitas adalah siemens per metre (S/m). Namun penggunaan satuan decisiemens per metre (dS/m) lebih sering digunakan. Sebagai patokan umum, nilai salinitas air biasa (tap water) ialah 0.3 dS/m dan air salin sekitar 30 dS/m (Water 2007).
Proses akumulasi garam pada permukaan tanah terjadi karena gerakan garam dari profil tanah bawah (sub-soil) ke bagian atas (top-soil). Pada bagian atas terjadi penguapan yang intensif, sehingga menyebabkan larutan garam bergerak secara kapilaritas ke atas, menguap, dan meninggalkan endapan garam di permukaan tanah. Apabila proses ini berlangsung terus menerus sepanjang tahun, maka terbentuk tanah garam. Di Indonesia proses ini tidak berlangsung sepanjang tahun, hanya terdapat di daerah panas dan kering. Pada musim kemarau terjadi salinisasi, sebaliknya pada musim hujan terjadi desalinisasi. Pengurangan kadar garam di permukaan tanah terjadi karena curah hujan yang turun kemudian melindi ke bawah. Proses salinisasi hanya terjadi pada tanah yang mempunyai tekstur halus sampai sangat halus (Rosmarkam dan Yuwono 2002). Van Asten et al. (2004) menyatakan bahwa air mampu melarutkan molekul garam dan mengangkutnya sebagai aliran permukaan (run off) maupun pencucian (leaching), sehingga kadar garam dapat berkurang. Pada skala bentang lahan, salinitas tanah dapat berkurang akibat pasokan air hujan dan air irigasi pada volume dan intensitas yang cukup.
Dampak kenaikan muka air laut ialah meningkatnya salinitas air tanah dan tergenangnya sebagian lahan di wilayah pantai. Peningkatan salinitas tanah menjadi 3.9 dan 6.5 dS/m akan menurunkan hasil padi sebesar 25% dan 55% (Zeng dan Shannon 2000). Penurunan produksi bersifat linier sebesar 10% akibat peningkatan salinitas 1-2 dS/m. Luas lahan sawah yang terancam akibat salinitas maupun tenggelam di Pantura Jawa diperkirakan 292-400 ribu ha (3.7%). Ancaman paling serius akibat penciutan lahan terjadi di Jawa Barat dengan penurunan produksi hingga 16.000 ton per tahun (Boer et al. 2011).
Dampak Salinitas Tanah terhadap Tanaman Padi
Tanaman padi tergolong tanaman yang moderat sensitif terhadap salinitas, yaitu tanaman yang masih toleran pada taraf EC berkisar antara 0.9 – 1.6 dS/m. Cekaman salinitas akan terlihat terutama pada masa pembibitan dan awal pertumbuhan yang berdampak pada penurunan hasil. Penurunan produksi padi sawah akibat cekaman salinitas sudah banyak dilaporkan di antaranya sebagai dampak salinitas dari air irigasi. Nilai EC air irigasi < 0.7 dS/m tidak menimbulkan masalah salinitas, nilai EC antara 0.7 – 1.7 dS/m mulai memperlihatkan masalah salinitas ringan sampai sedang, dan nilai EC > 2.0 dS/m terjadi penurunan produksi secara signifikan (Scardaci et al. 1999). Tiap peningkatan 1 unit nilai EC air genangan dengan nilai EC > 2.0 dS/m menurunkan hasil padi sampai 1 t/ha (Asch dan Wopereis 2001).
Rendemen padi paling sensitif terhadap kondisi salin ditentukan selama tahap inisiasi malai. Kerentanan padi terhadap kondisi salin bervariasi selama tahap pertumbuhannya. Padi relatif toleran kondisi salin pada saat perkecambahan, bahkan pada beberapa kasus hingga 16.3 dS/m tidak terpengaruh secara signifikan (Heenan et al. 1988, Khan et al. 1997). Padi menjadi sangat sensitif pada tahap bibit muda yang berdampak pada kemampuannya bertahan pada lahan salin (Lutts et al. 1995) dan relatif kurang sensitif selama tahap akhir reproduksi (pematangan biji-bijian). Salinitas secara signifikan dapat mengurangi jumlah anakan per tanaman, jumlah gabah per malai, kesuburan, panjang malai, dan cabang primer per malai (Heenan et al. 1988, Cui et al. 1995, Khatun et al.
1995, Zeng et al. 2002). Penurunan jumlah anakan per tanaman dan jumlah gabah per malai merupakan penyebab utama kehilangan hasil pada satu kultivar padi pada kondisi adanya cekaman salinitas (Zeng dan Shannon 2000). Jumlah gabah per malai menjadi komponen hasil yang paling sensitif. Komponen ini ditentukan pada tahap reproduksi awal, sekitar inisiasi malai (Hoshikawa 1989, Counce et al. 2000).
Komunitas dan Aktivitas Mikrob Rhizosfer
terhadap lingkungannya, sehingga kestabilan struktur populasi mikrob menjadi ukuran kualitas habitat tersebut (McChang et al. 1999).
Menurut Sorensen (1997), rhizosfer ialah volume tanah yang berada di sekitar perakaran dan dipengaruhi oleh perakaran tanaman. Bagian ini dikenal penting untuk kesehatan tanaman dan kesuburan tanah. Beberapa studi menunjukkan bahwa keragaman struktur dan fungsi dari populasi rhizosfer dipengaruhi oleh spesies tanaman sehubungan dengan perbedaan dalam eksudasi akar dan rhizodeposisi pada daerah perakaran. Faktor-faktor yang mempengarui komposisi komunitas mikrob rhizosfer antara lain jenis tanah, tahap pertumbuhan, praktek pertanaman, sejarah penggunaan lahan, dan faktor lingkungan lainnya (Grayston et al. 1998, Lupwayi et al. 1998, Doi 2011). Tilak et al. (2005) menyatakan bahwa komunitas bakteri rhizosfer tanaman memiliki sistem yang efisien untuk mengambil dan melakukan katabolisme senyawa-senyawa organik yang berada di dalam eksudat akar.
Mikrob rhizosfer memiliki pengaruh yang baik pada pertumbuhan dan kesehatan tanaman dengan cara melarutkan unsur hara, menambat N2, atau
menghasilkan zat pemacu tumbuh (Hoflich et al. 1994, Patten dan Glick 1996). Bakker et al. (1991) menyatakan bahwa beberapa mikrob rhizosfer memiliki aktivitas biokontrol, menurunkan penyakit tanaman dengan menekan patogen tanah melalui kompetisi nutrisi, siderofor, atau antibiosis. Adanya interaksi mikrob dengan tanaman inang dapat mempengaruhi berbagai proses biogeokimia yang selanjutnya mempengaruhi siklus unsur-unsur penting di dalam tanah.
Tilak et al. (2005) mengemukakan bahwa sejumlah spesies bakteri rhizosfer tanaman tergolong ke dalam genus Azospirillum, Alcaligenes, Arthrobacter,
Bacillus, Burkholderia, Enterobacter, Erwinia, Flavobacterium, Pseudomonas,
Rhizobium, dan Serratia yang mampu memberikan efek positif bagi pertumbuhan tanaman. Rangarajan et al. (2002) melakukan pengamatan terhadap keragaman populasi Pseudomonas pada 5 lahan sawah dengan gradien salinitas di sepanjang pesisir Tamil Nadu di India bagian Selatan. Hasil yang diperoleh ialah bahwa peningkatan salinitas rhizosfer tanaman padi berpengaruh terhadap penurunan keragaman bakteri Pseudomonas dengan kecenderungan Pseudomonas fluorescens mendominasi lahan tidak salin; sedangkan P. alcaligenes dan P. pseudoalcaligenes mendominasi lahan salin. Keragaman Pseudomonas pada lahan salin tampak lebih tinggi pada lahan sawah organik yang menunjukkan adanya efek positif pertanian organik pada keragaman Pseudomas pada kondisi salin. Lebih lanjut Tripathi et al. (2002) juga telah mengamati keragaman bakteri tahan salin pada daerah rhizosfer tanaman padi. Setelah diperoleh 14 galur bakteri yang diisolasi menggunakan medium semi padat Nitrogen Free bromthymol blue (NFb) yang telah diperkaya dan menunjukkan toleransinya pada konsentrasi NaCl sebesar 3%, kemudian galur-galur tersebut dianalisis dengan teknik Amplified Ribosomal DNA Restriction Analysis (ARDRA) menggunakan enzim restriksi
Sau3AI, AluI dan RsaI serta teknik Random Amplified Polymorphic DNA
(RAPD). Hasil yang diperoleh menunjukkan bahwa terdapat 4 grup yang berkerabat dekat dengan Alcaligenes xylosoxidans, Ochrobactrum anthropi,
Bakteri baru tersebut memiliki kemampuan menambat nitrogen, melarutkan fosfat dengan adanya NaCl. Analisis filogenetik berdasarkan sekuen gen 16S rRNA menunjukkan bahwa galur-galur ini dekat dengan genus Acidomonas, Asaia,
Acetobacter, Gluconacetobacter, Gluconobacter, dan Kozakia.
Dampak Salinitas Tanah terhadap Mikrob Rhizosfer
Lingkungan salin merupakan habitat yang ekstrim untuk pertumbuhan mikrob. Kelimpahan, komposisi, keragaman, dan fungsi metabolik komunitas mikrob lebih rendah di lingkungan salin dan hipersalin (Jiang et al. 2007). Tipe lingkungan seperti ini disukai oleh sejumlah bakteri halotoleran yang tidak memerlukan NaCl untuk tumbuh dan halofilik yang memerlukan NaCl untuk pertumbuhannya. Ada 3 kelompok bakteri halofil berdasarkan respon terhadap NaCl, yaitu agak halofil (tumbuh pada 2-5% NaCl), halofil moderat (tumbuh pada 5-20% NaCl), dan halofil ekstrim (tumbuh pada 20-30% NaCl).
Salinitas pernah dipelajari terutama sebagai parameter yang menurunkan proliferasi mikrob dan aktivitasnya (Lu et al. 2001, Caton et al. 2004). Nelson dan Mele (2007) telah mengamati adanya perubahan struktur komunitas mikrob rhizosfer tanaman gandum sebagai respon terhadap peningkatan konsentrasi boron dan NaCl menggunakan BIOLOG dan analisis Terminal Restriction Fragment Length Polymorphism (T-RFLP). Perubahan terjadi secara nyata pada struktur komunitas mikrob rhizosfer tanaman gandum. Adanya NaCl sangat berpengaruh nyata pada perubahan ini, terutama pada bakteri rhizosfer penambat N2, namun
tidak demikian pada bakteri pengoksidasi ammonia. Penjelasan mengenai hasil ini diduga terkait dengan adanya perubahan fisik di daerah rhizosfer sehubungan dengan tekanan osmotik dan gangguan fungsi tanaman. Adanya gangguan pada tanah dengan kandungan NaCl tinggi berdampak pada air yang diaplikasikan tidak dapat dipergunakan dengan baik, sehingga tanah menjadi lembab. Peningkatan kelembaban akan menurunkan kandungan O2 dan meningkatkan akumulasi CO2,
sehingga terjadi seleksi jenis mikrob dan aktivitasnya. Perubahan struktur komunitas mikrob rhizosfer juga terkait dengan kualitas dan kuantitas eksudat akar tanaman gandum pada habitat tersebut.
Ibekwe et al. (2010) mengamati struktur komunitas bakteri rhizosfer tanaman ketimun sebagai efek terhadap salinitas tanah (3 dan 8 dS/m), boron (0.7, 5, dan 8 mg/L), dan pH (6.5 dan 8) dengan metode kultur dan nonkultur dengan
Metode Identifikasi Komunitas Mikrob
Metode yang umum digunakan untuk keperluan identifikasi komunitas mikrob ada dua, yaitu: (1) metode yang mengandalkan penggunaan teknik molekuler untuk analisis DNA; dan (2) metode yang hanya mengidentifikasi mikrob yang dapat dikulturkan. Beberapa metode molekuler yang melibatkan ekstraksi dan analisis DNA dari seluruh komunitas mikrob untuk identifikasi sidik jari genetiknya meliputi Terminal Restriction Fragment Length Polymorphism (T-RFLP), Denaturing Gradient Gel Electrophoresis (DGGE), Thermal Gradient Gel Electrophoresis (TGGE), Single Strand Conformation Polymorphism (SSCP),
Automated Ribosomal Intergenic Spacer Analysis (ARISA), amplifikasi fragmen-fragmen asam nukleat dengan teknik Polymerase Chain Reaction (PCR), dan
Fluorescent In Situ Hybridization (FISH). Struktur dari profil-profil yang didapatkan selanjutnya dianalisis melalui Principal Component Analysis (PCA) atau analisis komputasi (misalnya analisis kluster dan dendrogram). Walaupun sidik jari DNA tadi bersifat culture-independent, namun dapat menaksir secara cepat kompleks komunitas mikrob dari berbagai lingkungan (Dunbar et al. 2000, Ranjard et al. 2001, Smalla et al. 2001), yang hasilnya kemungkinan dipengaruhi oleh metode sekuensing berbasis PCR yang mungkin bias dengan hasil analisis berbasis molekuler dari struktur komunitas mikrob (Jones dan Thies 2007). Metode identifikasi mikrob yang dapat dikulturkan dilakukan dengan mengkultivasi berbagai tipe mikrob di dalam sampel pada nutrisi selektif untuk memacu pertumbuhannya. Struktur komunitas ditaksir dengan mengidentifikasi isolat-isolat dari koloni yang dominan. Prosedur ini tidak efisien dari segi biaya dan tenaga, karena setiap isolat harus dipelajari lebih lanjut dengan mengamati sifat fisiologi, taksonomi, dan reaktivitasnya terhadap pewarna. Basis data yang lengkap yang terdiri atas sejumlah isolat telah dikembangkan untuk membantu studi mengenai komunitas mikrob dengan pendekatan culture-dependent
(Kampfer et al. 1996). Menurut Hattori (1988), populasi bakteri yang diambil dari sampel alam mungkin tidak terwakili secara akurat pada saat ditumbuhkan pada media agar-agar. Teknik pengkulturan kurang memadai untuk menganalisis komunitas mikrob pada lingkungan alaminya, sehubungan dengan adanya sejumlah besar bakteri yang belum dapat dikulturkan (Amann et al. 1995). Pendekatan culture-dependent memiliki kelemahan dalam hal selektivitas yang rendah dan tidak efisien karena periode pengkulturan yang lama, dan teknik ini juga kurang akurat menggambarkan komunitas bakteri yang disebabkan oleh perubahan kondisi lingkungan (Liu et al. 1997)
3 METODE
Tempat dan Waktu Penelitian
Penelitian ini dilakukan di Laboratorium Mikrobiologi dan Rumah Kaca Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB. Biogen) dan Laboratorium Fisiologi C, Pusat Penelitian Biologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI). Penelitian ini dilakukan sejak bulan Juni 2012 sampai dengan bulan Juni 2014.
Bahan dan Alat Penelitian
Sampel tanah
Tanah untuk media tanam di pot pada penelitian ini diambil dari 2 lokasi lahan sawah di daerah pesisir Jawa Barat, yaitu dari Desa Patimban, Kecamatan Pusaka Negara, Kabupaten Subang dan Desa Eretan Kulon, Kecamatan Kandanghaur, Kabupaten Indramayu. Setiap lokasi lahan sawah di daerah tersebut diambil 3 jenis sampel tanah (salinitas sedang dengan EC 2 - 4 dS/m, salinitas rendah dengan EC < 2 dS/m, dan salinitas sangat rendah sebagai kontrol dengan EC mendekati 0 dS/m) pada kedalaman hingga 20 cm (lapisan tanah olah). Peta lokasi dan data-data pendukung seperti karakteristik fisikokimia tanah, letak koordinat, jarak pengambilan dari batas pantai ditunjukkan pada Lampiran 1 dan Lampiran 2.
Bahan
Bahan-bahan yang digunakan pada penelitian ini meliputi: bahan kimia untuk pembuatan media dan reagen, antibiotik, akuades, kit Wizard DNA genomic extraction kit (Promega), IsoilTM DNA Kit (Nippon Gene), kit GeneJET PCR Purification (Thermo Scientific), TOPO TA Kloning kit (Invitrogen), Qiaprep® Spin Miniprep kit (Qiagen), bufer Tris-Acetic acid-EDTA (TAE) (50x, 1x, dan 0.5x), bufer Tris-EDTA (TE), kloroform, ethanol 70%, natrium asetat 3 M, GoTaq® Green Master Mix, loading dye (Promega, Madison), nuclease free water, marker DNA ladder 100 bp dan 1 kb, primer-primer 16S rRNA, agarosa, etidium bromide.
Alat
Prosedur Analisis
Persiapan sampel dan analisis sampel tanah
Sifat fisik dan kimia tanah dari daerah pesisir Subang dan Indramayu dianalisis terlebih dahulu meliputi tekstur tanah, total C, total N, P2O5, K2O, pH,
salinitas (EC dan TDS), Kapasitas Tukar Kation (KTK), dan Kejenuhan Basa (KB) dilakukan di Balai Penelitian Tanah, sedangkan analisis total dan jenis bakteri tanah dilakukan di BB. Biogen Bogor. Analisis total bakteri tanah dilakukan dengan metode plate count agar, sebanyak 10 gram sampel tanah dimasukkan ke dalam 90 ml larutan fisiologis (8.5 g NaCl per liter akuades) selanjutnya dihomogenkan selama 30 menit dan diikuti dengan pengenceran serial dari 10-1 sampai 10-5. Sebanyak 100 µl dari hasil pengenceran ditumbuhkan pada media ekstrak tanah instan (Himedia) yang terdiri atas glukosa 0.1%, dikalium fosfat 0.05%, dan ekstrak tanah 1.775%. Inkubasi dilakukan selama 1-2 minggu pada suhu ruang. Koloni yang tumbuh dihitung dan diambil koloni-koloni yang dominan untuk diidentifikasi secara molekuler.
Sampel-sampel tanah bagian rhizosfer dipersiapkan dengan memasukkan tanah-tanah dari kedua daerah pesisir tersebut pada bak-bak percobaan yang terbuat dari kayu dengan ukuran 75 cm x 70 cm x 30 cm dan diisi dengan 80 Kg tanah/bak. Bibit padi varietas Mekongga dipersiapkan dengan cara merendam benih padi selama 24 jam di dalam air yang telah ditambahkan NaCl 30 gram/L, kemudian benih yang tenggelam diambil dan dicuci, selanjutnya diperam selama 24 jam menggunakan kain saring dan disebar di atas bak plastik untuk persemaian hingga berumur 25 hari, dan setiap bak-bak percobaan ditanami dengan 4 bibit padi dengan jarak tanam 25 cm x 25 cm. Pemupukan tanaman dengan takaran 300 Kg/ha Urea, 100 Kg/ha SP36, dan 100 Kg/ha KCl. Pupuk Urea disebar 3 kali, yaitu sebelum tanam, 21, dan 42 hari setelah pindah tanam. Pupuk SP36 diberikan seluruhnya sebelum tanam, sedangkan pupuk KCl disebar 2 kali, yaitu sebelum tanam dan 21 hari setelah pindah tanam.
Rancangan percobaan pada penelitian ini ialah faktorial rancangan acak lengkap (RAL) dengan 2 perlakuan berupa (1) jenis tanah, yaitu bahan induk marin (tanah yang diambil dari Desa Patimban, Kabupaten Subang) dan bahan induk fluvio-marin (tanah yang diambil dari Desa Eretan Kulon, Kabupaten Indramayu) dan (2) gradien salinitas tanah pesisir, yaitu: (1) salinitas sedang (EC 2 - 4 dS/m); (2) salinitas rendah (EC < 2 dS/m); (3) salinitas sangat rendah sebagai kontrol (EC mendekati 0 dS/m) dengan 4 ulangan.
Tiga hari menjelang tanam, tanah dalam pot diairi dan dilumpurkan. Untuk menentukan tingkat salinitas digunakan alat Electrical Conductivity meter (EC meter) dengan cara sampel tanah dalam pot perlakuan diambil pada bagian atas pada kedalaman 0-15 cm dan bagian bawah pada kedalaman 15-30 cm (Erfandi dan Rachman 2011). Selama percobaan berlangsung taraf salinitas tetap dipertahankan dengan jalan mengukurnya setiap seminggu sekali sampai umur 70 HST (akhir fase vegetatif). Tanah dibiarkan tergenang dengan ketinggian 5 cm hingga tanaman berumur 70 HST.
tanaman, meliputi parameter tinggi tanaman (cm), bobot kering (BK) tajuk (g), BK akar (g), dan jumlah anakan produktif diukur pada fase primordia dan fase pembungaan. Tinggi tanaman diukur dari permukaan tanah sampai ujung malai paling panjang. Jumlah anakan produktif merupakan jumlah anakan yang mampu menghasilkan malai per rumpun (Suprihatno et al. 2010). Bobot kering tajuk diperoleh dari bobot tanaman padi bagian atas (tanaman padi yang telah dipotong 4 cm di atas permukaan tanah) yang telah dicuci dan dikeringkan pada suhu 60 oC selama 48 jam hingga nilai bobot kering yang konstan. Parameter komponen hasil yang meliputi jumlah malai, pajang malai, persentase biji isi/malai (%), bobot total biji/rumpun (g), dan BK bulir padi (g) diukur pada waktu panen (105 hari setelah pindah tanaman).
Hasil pengukuran parameter pertumbuhan dan komponen hasil tanaman padi dianalisis dengan manggunakan analysis of variance (ANOVA) untuk rancangan faktorial RAL. Perbedaan rata-rata pada setiap perlakuan (taraf salinitas tanah) ditentukan dengan menggunakan uji selang berganda Duncan pada taraf uji 5%. Analisis korelasi Pearson dilakukan pada parameter pertumbuhan dan komponen hasil tanaman padi dengan salinitas untuk mengetahui hubungan parameter tanaman dengan salinitas. Kedua analisis tersebut dilakukan dengan menggunakan program SPSS versi 17.0.
Penelitian Tahap I: Analisis komunitas dan fungsi bakteri rhizosfer tanaman padi dengan metode kultur
Selama pertumbuhan tanaman dilakukan analisis komunitas bakteri rhizosfer tanaman pada fase primordia dan fase pembungaan dengan metode kultur dan nonkultur. Analisis komunitas dan fungsi bakteri rhizosfer tanaman padi dengan metode kultur dilakukan dengan tahapan kegiatan meliputi isolasi bakteri rhizosfer tanaman padi, ekstraksi DNA dan amplifikasi gen 16S rRNA bakteri tersebut, selanjutnya identifikasi. Isolat-isolat yang diperoleh dan sudah diidentifikasi selanjutnya dilakukan karakterisasi fungsional.
Isolasi bakteri rhizosfer tanaman padi
Sampel perakaran tanaman padi dengan tanah yang menempel pada perakaran diambil dari setiap pot perlakuan dan dimasukkan ke dalam plastik, selanjutnya disaring untuk memisahkan bagian akar dengan tanah bagian rhizosfer (Siciliano dan Germida 1999). Sebanyak 10 g sampel tanah dimasukkan ke dalam 90 ml larutan fisiologis, dikocok selama 30 menit, sehingga diperoleh pengenceran 10-1 dan selanjutnya dibuat seri pengenceran sampai 10-5. Selanjutnya sebanyak 100 μl hasil pengenceran 10-3 - 10-5 ditumbuhkan pada cawan petri yang berisi media ekstrak tanah instan. Inkubasi dilakukan selama 1 – 2 minggu pada suhu ruang. Koloni yang tumbuh dimurnikan dan disimpan pada media agar-agar ekstrak tanah untuk keperluan analisis lebih lanjut.
Ekstraksi DNA dan amplifikasi gen 16S rRNA bakteri rhizosfer tanaman padi
DNA genom dari isolat bakteri rhizosfer tanaman padi diekstraksi menggunakan kit (Wizard DNA genomic extraction kit, Promega). Amplifikasi gen 16S rRNA dilakukan dengan primer 27F (5’AGA GTT TGA TCC TGG CTC
cetakan, masing-masing 1.0 µl primer, 12.5 µl Go Taq Green Master Mix 2x dan
nuclease free water hingga volume 25 μl. Amplifikasi DNA dilakukan dengan kondisi: denaturasi awal 94 oC selama 5 menit, diikuti 30 siklus yang terdiri atas denaturasi pada suhu 94 oC selama 1 menit 30 detik, penempelan primer pada suhu 55 oC selama 45 detik, polimerasi selama 1 menit pada suhu 72 oC dan pada siklus terakhir dilakukan perpanjangan waktu polimerasi selama 1 menit. Penghentian reaksi dilakukan dengan penurunan suhu ke 15 oC. Pemurnian produk PCR dilakukan dengan metode pengendapan etanol (Sambrook dan Russell 2001).
Identifikasi isolat bakteri rhizosfer tanaman padi
Produk PCR gen 16S rRNA dari isolat bakteri rhizosfer tanaman padi diidentifikasi secara molekuler dengan melakukan sekuensing. Sekuensing DNA dilakukan di Macrogen (Korea). Hasil sekuen dianalisis dengan menggunakan program BIOEDIT dan sebelumnya dilakukan pemeriksaan terlebih dahulu basa-basa nukleotidanya dengan program CHROMASPRO. Sekuen yang telah diedit selanjutnya disejajarkan untuk mencari kesamaannya menggunakan CLASSIFIER (http://rdp.cme.msu.edu) dari database RDP-II, Michigan State University, MI, USA menggunakan Seqmatch version 3.
Karakterisasi fungsional bakteri rhizosfer tanaman padi
Karakterisasi fungsional bakteri rhizosfer tanaman padi yang dilakukan meliputi penapisan bakteri pelarut fosfat, penambat N2, pendekomposisi selulosa,
dan penghasil IAA.
Penapisan bakteri pelarut fosfat. Bakteri rhizosfer tanaman padi diseleksi kemampuannya dalam melarutkan fosfat anorganik (trikalsium fosfat) menggunakan media agar-agar yang terdiri atas 1% glukosa, 0.05% (NH4)2SO4,
0.01% MgSO4.7H2O, 0.05% ekstrak ragi, 0.02% KCl, 0.02% NaCl, 0.0002%
FeSO4.7H2O, 0.0002% MnSO4.H2O, 0.5% Ca3(PO4)2. Inkubasi dilakukan pada
suhu 28 oC selama 7 hari. Terbentuknya zona bening di sekitar koloni bakteri mengindikasikan kemampuan pelarutan fosfat (Gupta et al. 1994). Percobaan dilakukan dengan 3 ulangan untuk setiap jenis bakteri.
Penapisan bakteri penambat N2. Penapisan awal bakteri pemfiksasi N2
dilakukan pada media semi padat Nitrogen free bromthymol blue (NFB) yang terdiri atas 0.5% asam DL-malat, 0.4% KOH, 0.05% K2HPO4, 0.01%
MgSO4.7H2O, 0.005% MnSO4.H2O, 0.002 % NaCl, 0.001% CaCl2 0,005%
FeSO4.7H2O, 0,0002% Na2MoO4.2H2O dan 0,175% agar bakto, 2 ml 0,5%
bromtimol biru (BTB). Lima hari setelah inkubasi pada suhu 28oC, bakteri yang menunjukkan adanya pembentukan pelikel di bawah permukaan media diduga awal sebagai bakteri yang positif menambat N2 (Dobereiner 1995).
Penapisan bakteri pendekomposisi selulosa. Penapisan isolat selulolitik dilakukan pada media agar Carboxy Methyl Cellulose (CMC) yang mengandung 0,5%, carboxymethyl selulosa; 0.1% NaNO3, 0.1% K2HPO4, 0.1% KCl, 0.05%
MgSO4.7H2O, 0.05% ekstrak ragi, 1.5% agar-agar bakto, pH 8,0. Inkubasi
Penapisan bakteri penghasil IAA. Bakteri rhizosfer tanaman padi ditumbuhkan pada suhu 28oC selama 24 jam pada medium NB. Sebanyak 100 µl kultur tersebut ditambahkan ke dalam 10 ml media minimal yang terdiri atas 0.136% KH2PO4, 0.213% Na2HPO4, 0.02% MgSO4.7H2O dan 10 ml trace
element (700 mg CaCl2.2H2O, 300 mg FeSO4.7H20, 20 mg MnSO4.H20, 40 mg
CuSO4.5H2O, 20 mg ZnSO4.7H2O, 3 mg H3BO3, 7 mg CoCl2.6H2O, 4 mg
Na2MoO4.H2O, dan 1 ml H2SO4 untuk total volume 1 liter) yang telah ditambah
dengan 1 ml L-triptofan (10% glukosa, 1% L-triptofan, dan 0.1% ekstrak ragi disaring menggunakan filter milipore 0,2 m). Selanjutnya kultur bakteri rhizosfer diinkubasi pada suhu 28oC selama 48 jam. Setelah itu dilakukan sentrifugasi kultur pada kecepatan 6000 xg, pada suhu 4oC selama 10 menit untuk memisahkan sel bakteri dengan bagian supernatannya. Sebanyak 1 ml bagian supernatan dari kultur tersebut diambil dan ditambah dengan 2 ml reagen (1 ml 8.12% FeCl3.6H2O, 50 ml HClO4 35% di dalam botol gelap), kemudian
diinkubasi pada suhu 28oC selama 25 menit. Pembacaan absorbansi dilakukan pada 530 nm menggunakan spektrofotometer. Konsentrasi IAA pada setiap sampel ditentukan dengan dibandingkan dengan kurva standar IAA. Pembacaan dilakukan dengan 3 ulangan untuk masing-masing jenis bakteri (Gupta et al.
2012).
Penelitian Tahap II: Analisis komunitas bakteri rhizosfer tanaman padi dengan metode nonkultur
Analisis komunitas bakteri rhizosfer tanaman padi dengan metode nonkultur dilakukan dengan tahapan kegiatan ekstraksi DNA dari sampel tanah rhizosfer tanaman padi dan amplifikasi gen 16S rRNA, kloning dan sekuensing gen 16S rRNA, serta analisis data sekuen.
Ekstraksi DNA tanah dan amplifikasi gen 16S rRNA
Ekstraksi dan pemurnian DNA dilakukan langsung dari sampel tanah rhizosfer tanaman padi yang diambil dari setiap pot perlakuan (duplo untuk setiap pot perlakuan) menggunakan IsoilTM DNA Kit (Meis dan Chen 2003). Sebanyak 0.5 g sampel tanah ditempatkan pada tabung beat beads 2 ml kemudian ke dalam tabung ditambahkan 950 μl Larutan Pelisis BB (pH 8,6) yang mengandung 1% SDS, 100 mM Tris HCl, 200 mM EDTA, dan 200 mM Na2HPO4, dan 50 μl Larutan Pelisis 20S yang mengandung 20% SDS.
Polyethylene glycol (PEG) dan 1.5 M Tris-HCl, kemudian dihomogenisasi perlahan-lahan. Selanjutnya dilakukan sentrifugasi selama 20 menit dengan kecepatan 16000 g (rcf) pada suhu 4 °C. Supernatan dibuang perlahan, kemudian ke dalam tabung ditambahkan larutan pencuci sebanyak 1 ml dan disentrifugasi kembali pada kecepatan 16000 g (rcf) pada suhu 4 °C selama 15 menit, kemudian supernatan dibuang dengan hati-hati. Langkah terakhir ialah menambahkan etanol
70% dingin sebanyak 1 ml dan 100 μl Natrium asetat, kemudian larutan disentrifugasi pada kecepatan 16000 g (rcf) pada suhu 4 °C selama 5 menit. Pelet DNA dikeringkan dengan membalik tabung di atas tisu kering selama 15 menit. Ke dalam tabung yang berisi pelet DNA dilarutkan dengan 20-50 μl bufer TE (10 mM Tris base, 1 mM EDTA; pH 8). Setelah didapatkan DNA hasil isolasi total komunitas bakteri, dilanjutkan dengan pengecekan hasil isolasi DNA menggunakan nanodrop dan juga dilakukan elektroforesis.
Amplifikasi gen 16S rRNA dilakukan dengan primer 63f (5’ CAG GCC
TAA CAC ATG CAA GTC 3’) dan 1387r (5’ GGG CGG WGT GTA CAA GGC 3’) (Marchesi et al. 1998). Proses amplifikasi PCR dibuat dengan mencampurkan secara homogen komponen-komponen berikut ini: 8.5 μl ddH2O (nuclease free water), 12.5 μl Go Taq Green Master Mix 2x, masing-masing 1.0 μl primer untuk
gen 16S rRNA dan 2.0 μl DNA cetakan, sehingga total volume 25 µl. Kondisi PCR sama dengan kondisi PCR di atas dengan suhu penempelan primer 55 °C. Kualitas produk PCR diperiksa menggunakan nanodrop dan dielektroforesis pada gel agarosa.
Elektroforesis dilakukan untuk mendeteksi hasil isolasi DNA dan amplifikasi PCR. Pertama, dilakukan pembuatan gel agarosa dengan 1% gel agarosa (0.4 g agarosa dalam 40 ml bufer TAE 1x) dipanaskan sampai larut. Gel yang sudah larut ditunggu sampai hangat kemudian dituang ke dalam cetakan yang telah disiapkan sebelumnya. Gel dibiarkan memadat selama 30 menit. Apabila gel sudah siap, sisir dilepaskan dari cetakan kemudian gel diletakkan ke dalam tangki elektroforesis yang berisi bufer TAE 1x hingga terendam setinggi 1-2 mm. Pengecekan hasil isolasi DNA dilakukan dengan mencampurkan 3 μl DNA hasil ekstraksi dengan 1μl loading buffer di atas plastik parafilm, kemudian diaduk perlahan dengan pipet dan dimasukkan ke dalam sumur yang terdapat pada gel. DNA hasil amplifikasi PCR dideteksi dengan memasukkan 3 μl hasil PCR dan 2 μl gen penanda DNA ke dalam sumuran pada gel. Selanjutnya alat elektroforesis disambungkan ke sumber tegangan. Proses elektroforesis berlangsung selama 25 menit pada tegangan 100 volt. Gel direndam di dalam larutan etidium bromida selama 15 menit kemudian dibilas dalam bufer TAE 1x. Gel diletakkan di atas UV transluminator dan didokumentasikan.
Kloning dan sekuensing gen 16S rRNA
Produk PCR dari setiap perlakuan (2 produk PCR per perlakuan) dikumpulkan dan dimurnikan dengan GeneJET PCR Purification Kit (Thermo Scientific). Selanjutnya gen hasil amplifikasi diligasikan ke dalam vektor pCR TOPO 2.1 (Invitrogen) dan ditransformasi ke Escherichia coli kompeten TOP10 (Invitrogen) sesuai dengan metode dari kit (TOPO TA Kloning Kit). Rekombinan positif yang diindikasikan oleh koloni-koloni putih di cawan petri berisi media LA yang telah ditambah dengan Ampisilin 50 µg/ ml dan 40 µl
40 klon dari setiap perlakuan. Verifikasi plasmid rekombinan dilakukan dengan mengamplifikasi gen 16S rRNA yang sudah diligasikan ke dalam vector pCR TOPO 2.1 menggunakan primer M13f (5’ TGT AAA ACG ACG GCC AGT 3’) dan M13r (5’ AGG AAA CAG CTA TGA CCA T 3’). DNA plasmid selanjutnya diisolasi menggunakan Qiagen Mini Prep. (Qiagen) dan disekuen dengan dikirimkan ke Macrogen (Korea). Klon-klon yang disekuensing merupakan klon-klon yang representatif setelah dilakukan pemotongan dengan enzim restriksi RsaI dan MspI.
Analisis data sekuen
4 HASIL DAN PEMBAHASAN
Sifat-sifat Fisik, Kimia, dan Mikrobiologi Tanah Sawah Pesisir Subang dan Indramayu
Karakteristik tanah sawah di pesisir Subang dan Indramayu hampir sama, yaitu untuk parameter C tergolong rendah karena berada pada kisaran 1-2%, demikian juga N yang berada pada kisaran 0.1-0.2% (Lampiran 2). Rendahnya kadar bahan organik tanah sawah banyak dipicu oleh peningkatan penggunaan pupuk anorganik terutama nitrogen tanpa diikuti penggunaan pupuk organik yang memadai. Secara umum lahan sawah yang digunakan secara intensif, sebagian telah terdegradasi yang dicirikan oleh penurunan produktivitas tanah, kandungan C organik dan unsur-unsur hara tanah makro. Hal ini dapat berakibat hilangnya fungsi penting bahan organik dalam memelihara produktivitas tanah yang berujung pada kerusakan fisik, kimia, dan biologi tanah. Hasil penelitian dari 8.1 juta ha lahan sawah di Indonesia, sekitar 65% tanah sawah mempunyai kandungan C organik rendah sampai sangat rendah (Kasno et al. 2000). Parameter P2O5 dan
K2O tergolong sangat tinggi di atas 60%, dengan pH tanah masam hingga agak
masam. Nilai pH yang baik untuk pertumbuhan tanaman padi berkisar antara 5.5-6.5 (Supparattanapan et al. 2009). Salinitas di tanah pesisir Subang dan Indramayu biasanya diinduksi oleh adanya penggenangan sebagai akibat naiknya air laut. Salinitas seperti ini sering berasosiasi dengan pH tanah yang rendah. Tanah pesisir pada kedua wilayah tersebut tergolong ke dalam kelas tekstur liat atau klei (Lampiran 2). Tingginya persentase klei perlu diperhatikan kaitannya dengan salinitas, karena lapisan klei dan juga debu bersifat relatif impermeable
atau tidak dapat ditembus. Hal ini berakibat lambatnya proses filtrasi atau
leaching garam, sehingga proses desalinisasi terjadi sangat lambat (FAO 2005). Kapasitas tukar kation (KTK) menunjukkan jumlah kation yang dapat dijerap oleh tanah per satuan berat tanah dan sangat erat hubungannya dengan kesuburan tanah. Tanah dengan KTK tinggi mampu menjerap dan menyediakan unsur hara lebih baik daripada tanah dengan KTK rendah (Sarwono 2010). Nilai KTK tanah pesisir Subang dan Indramayu masih tergolong sedang sampai tinggi dengan dominansi kation basa berupa Na, Ca, Mg, dan K (Lampiran 2).
Kejenuhan basa menunjukkan perbandingan antara jumlah kation-kation basa dan jumlah semua kation yang terdapat dalam kompleks jerapan tanah. Kation-kation basa umumnya merupakan unsur hara yang diperlukan tanaman, namun bersifat mudah tercuci (Sarwono 2010). Tanah pesisir Subang dan Indramayu berdasarkan kriteria penilaian sifat kimia tanah (Puslittanah 1983) berada >70%, yang berarti memiliki kejenuhan basa sangat tinggi dan belum banyak mengalami pencucian (Lampiran 2).
untuk menambah informasi dari aspek kesehatan tanah, karena selain mikrob yang dapat dikultur juga memberikan informasi mikrob yang tidak dapat dikulturkan. Adanya informasi sifat-sifat fisik, kimia, dan mikrobiologi tanah dapat digunakan untuk menduga kondisi tanah-tanah sawah di wilayah pesisir setelah adanya pengaruh intrusi air laut.
Sebelum dilakukan pengkondisian terhadap tanah dari lahan sawah daerah pesisir Subang dan Indramayu, terlebih dahulu dilakukan tahapan isolasi bakteri dominan yang berada pada kedua jenis tanah tersebut untuk mengetahui informasi sifat mikrobiologi. Penelitian ini hanya difokuskan pada bakteri, meskipun masih banyak jenis mikrob lainnya yang berperan di dalam tanah seperti archaea, khamir, dan kapang. Media isolasi yang digunakan untuk pertumbuhan bakteri ini ialah media agar-agar ekstrak tanah, yang memiliki komposisi berupa glukosa, dikalium fosfat, ekstrak tanah, dan agar-agar (pH 6.8+0.2). Ekstrak tanah menyediakan semua nutrien penting yang diperlukan untuk pertumbuhan mikrob tanah. Glukosa bertindak sebagai sumber karbon dan dikalium fosfat sebagai penyangga medium. Sejumlah peneliti menggunakan media agar-agar ekstrak tanah, R-2A agar dan 1% nutrien agar untuk mempelajari keragaman bakteri tanah dengan metode kultur. Isolat-isolat yang tumbuh pada media agar-agar ekstrak tanah ternyata memiliki nilai indeks keragaman Shannon-Weaver and indeks
Simpson’s (kekayaan spesies) paling tinggi dibandingkan dengan media R-2A dan 1% nutrient agar. Filum-filum yang ditemukan pada ketiga media tersebut ialah Actinobacteria, Bacteroidetes, Firmicutes, dan Proteobacteria, dari genus Bacillus,
Flavobacterium, Pseudomonas, Rhizobium, and Streptomyces (Lutton et al.
2013). Hal ini sejalan dengan hasil penelitian lain bahwa pengkulturan menggunakan media agar-agar ekstrak tanah menghasilkan keragaman spesies bakteri lebih tinggi daripada media nutrien agar (Taylor 1951). Sekelompok bakteri tertentu memiliki kecepatan tumbuh lebih tinggi pada media ini, karena media agar-agar ekstrak tanah sangat sesuai dengan nutrisi di dalam tanah, bahkan spesies-spesies berpotensi baru seperti Acrocarpospora dan Streptosporangium
ditemukan pada media ini (Hamaki et al. 2005).
populasi minimum yang diduga di dalam tanah dan masih di bawah perkiraan, karena masih banyak bakteri yang belum dapat dikulturkan pada media buatan disebabkan oleh faktor nutrien, temperatur, pH, fisik, dan interaksi antar mikrob.
Gambar 2 Populasi bakteri tanah (log CFU/gram tanah) dari dua tanah sawah pesisir Subang and Indramayu, dengan taraf salinitas moderat ( ), rendah ( ), dan sangat rendah ( )
Hasil analisis sekuen terhadap isolat-isolat dominan yang tumbuh pada media agar-agar ekstrak tanah dari sampel tanah pesisir Subang dan Indramayu menunjukkan ragam dari isolat yang ada. Isolat-isolat tersebut telah diisolasi DNA-nya dan dilakukan amplifikasi gen 16S rRNA menggunakan primer universal 27F dan 1492R, sehingga diperoleh produk PCR berukuran 1465 pasang basa (Gambar 3). Analisis sekuen terhadap produk PCR 16S rRNA dan analisis bioinformatika hasil sekuen menunjukkan adanya keragaman jenis bakteri tanah dari lahan sawah daerah pesisir Subang dan Indramayu. Hasil sekuen menunjukkan ragam bakteri tanah pada taraf salinitas sangat rendah relatif lebih tinggi daripada taraf salinitas moderat (Gambar 4). Galur-galur yang tergolong ke dalam genus Bacillus, Klebsiella, dan Pseudomonas paling sering ditemukan di antara semua isolat. Genus Bacillus ditemukan pada semua taraf salinitas tanah, terutama Bacillus cereus complex, B. subtilis complex, dan B.megaterium.
Bacillus cereus complex merupakan nama kolektif untuk B. cereus dan kerabat dekatnya seperti B. anthracis, B. thuringiensis, dan sebagainya (Jensen et al. 2003). Bacillus cereus dan B. anthracis bersifatpatogen pada manusia dan hewan ternak, sedangkan B. thuringiensis bersifat patogen pada serangga. Bacillus subtilis complex juga mencakup banyak spesies yang berkerabat dekat satu dengan yang lain seperti B. amyloliquefaciens, B. atrophaeus, B. vallismortis
Gambar 3 Amplifikasi 16S rRNA genom bakteri tanah dari daerah pesisir Indramayu (M: 1 kb ladder, 1: Er C2, 2: Er C4, 3: Er C6, 4: Er A3, 5: Er A14)
Karakter fenotipik dan biokimia yang dapat membedakan spesies-spesies yang berkerabat dekat di dalam B. cereus dan B. subtilis sangat sedikit. Habitat B. subtilis complex biasanya di tanah, sumber air, atau berasosiasi dengan tanaman.
Bacillus sp. memiliki kemampuan heterotrofik yang kuat untuk mendegradasi beragam sumber karbon. Genus Klebsiella yang ditemukan di daerah Indramayu ialah Klebsiella oxytoca dan K. variicola. Klebsiella spp. sering ditemukan pada berbagai lingkungan seperti tanah dan air, memberikan kontribusi pada proses-proses biogeokimia, dan merupakan komponen utama mikroflora dari beberapa tipe lingkungan (Niemela dan Vaatanen 1982). Klebsiella oxytoca dan K. variicola dikenal sebagai bakteri yang mampu menambat N2 atau dikenal sebagai
bakteri diazotrof (Ladha et al. 1983). Genus lain seperti Vibrio merupakan salah satu jenis bakteri yang tergolong ke dalam kelompok marine bacteria yang secara umum habitat alaminya di laut. Hasil ini menunjukkan adanya variasi kondisi mikrob awal dari struktur populasi bakteri tanah dari dua lokasi tanah pesisir.
Gambar 4 Jenis-jenis spesies bakteri yang dominan pada tanah sawah pesisir
Subang (A) dan Indramayu (B) ( Bacillus subtilis complex,
Bacillus cereus complex, Bacillus megaterium, Vibrio furnissii,
Lysinibacillus boronitolerans, Staphylococcus pasteuri,
Klebsiella variicola, Klebsiella oxytoca, Pseudomonas
plecoglossicida, Pseudomonas knackmussii, Pseudomonas
Analisis Komunitas dan Fungsi Bakteri Rhizosfer Tanaman Padi pada Gradien Salinitas Tanah Pesisir dengan Metode Kultur
Analisis komunitas dan fungsi bakteri rhizosfer tanaman padi
Analisis komunitas juga dilakukan pada rhizosfer tanaman padi yang memasuki fase primordia (35 hari setelah pindah tanam) dan fase pembungaan (70 hari setelah pindah tanam) yang ditanam pada gradien salinitas tanah pesisir Subang dan Indramayu. Sampel diambil dari daerah rhizosfer tanaman padi dari setiap taraf salinitas, dan dianalisis dengan metode kultur. Sejumlah sampel tanah yang menempel pada perakaran tanaman padi dari setiap perlakuan dikulturkan pada media agar-agar ekstrak tanah untuk mengetahui populasi dan ragam bakteri yang dominan pada masing-masing habitat. Populasi bakteri rhizosfer tanaman padi yang dinyatakan dalam log CFU per gram tanah menunjukkan bahwa baik pada fase primordia maupun fase pembungaan populasi bakteri rhizosfer pada tanah dengan taraf salinitas sangat rendah cenderung lebih tinggi daripada tanah dengan taraf salinitas rendah dan moderat. Semakin rendah taraf salinitas maka semakin tinggi populasi bakteri rhizosfer tanaman padi (Gambar 5).
Gambar 5 Populasi bakteri rhizosfer tanaman padi varietas Mekongga (log CFU/gram tanah) pada fase primordia dan fase pembungaan dari tanah marin-Subang dan fluviomarin-Indramayu pada taraf salinitas moderat ( ), rendah ( ), dan sangat rendah ( )