(
Oryza sativa
L) SPESIFIK LAHAN PASANG SURUT
PARLIN HALOMOAN SINAGA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi „Analisis Genetik Potensi Ratun
Genotipe Padi (Oryza sativa L.) Spesifik Lahan Pasang Surut ’ adalah karya saya sendiri dengan arahan Komisi Pembimbing dan belum diajukan dalam bentuk apapun
kepada Perguruan Tinggi manapun. Sumber informasi yang berasal atau kutipan dari
karya yang diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan
dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, April 2015
Parlin Halomoan Sinaga
Lahan pasang surut di Indonesia terdapat seluas 20.1 juta hektar, 9.53 juta hektar di antaranya potensial untuk lahan pertanian, dan dari 9.53 juta hektar tersebut, 6 juta hektar berpotensi untuk areal tanaman pangan. Lahan pasang surut seluas 6 juta hektar seharusnya dapat menjadi sentra produksi padi utama di Indonesia. Usahatani padi di lahan pasang surut dihadapkan pada masalah lingkungan yang bersifat suboptimal untuk pertumbuhan tanaman, kekurangan tenaga kerja, sosial budaya, kekurangan modal usaha, dan fanatisme petani terhadap varietas lokal.
Produktivitas padi di lahan pasang surut Kalimantan berkisar 3.2 – 4.2 t ha-1. Produktivitas tersebut dapat ditingkatkan tanpa harus menanam dua kali dalam setahun, yaitu dengan memanfaatkan ratun. Ratun adalah tanaman yang tumbuh dari tunas yang terdapat di buku batang padi yang tersisa saat panen.
Kemampuan tanaman padi untuk menghasilkan ratun dapat ditentukan oleh: sifat genetik dan lingkungan. Kemampuan menghasilkan ratun merupakan karakter varietas yang dikendalikan secara genetik dan oleh karena itu berbeda pada berbagai jenis padi.
Potensi ratun pada padi-padi pasang surut perlu dikaji untuk memberi arah pemuliaan padi ratun. Penelitian berjudul “Keragaman dan Analisis Genetik Potensi Ratun Genotipe Padi (Oryza sativa L.) Spesifik Lahan Pasang Surut” bertujuan untuk menjelaskan lingkungan seleksi, keragaman potensi ratun berbagai genotipe padi pasang surut, aspek fisiologis, kendali genetik ratun, memperoleh galur padi tipe baru ratun, dan menyusun paket teknologi budidaya padi tipe baru ratun spesifik lahan pasang surut.
Penelitian dilaksanakan dalam enam (6) kegiatan. Percobaan di lapangan dilaksanakan dengan menggunakan rancangan acak kelompok lengkap yang diulang 3 kali. Bibit berumur 21 hari sejak semai (hss) ditanam 1 bibit per lubang tanam dengan jarak tanam 20 cm x 20 cm. Tanaman dipupuk dengan 150 kg ha-1 Urea, SP36 100 kg ha-1 SP36, dan 75 kg ha-1 KCl. Tanaman utama dipanen setelah 95% malai menguning. Pemotongan batang saat panen dilakukan pada ketinggian 10 cm dari permukaan tanah. Tanaman ratun dipupuk dengan 50 kg ha-1 Urea, 30 kg ha-1 SP36, dan 25 kg ha-1 KCl.
Hasil penelitian menunjukkan bahwa respon genotipe-genotipe padi terhadap ketiga lingkungan seleksi berdasarkan tipologi lahan pasang surut tidak berbeda, yang ditunjukkan oleh jumlah rumpun hidup yang sama antar tipologi lahan. Seleksi untuk kemampuan meratun dapat dilakukan pada tipologi lahan sulfat masam maupun lahan bergambut dengan tipe luapan air B dan C. Genotipe IPB107-F-7-3 mampu menghasilkan rumpun hidup terbanyak sebesar 86.6%.
Keragaman genetik dari kemampuan meratun genotipe padi luas. Seleksi untuk kemampuan meratun dapat dilakukan melalui karakter tanaman utama sebagai karakter sekunder, yaitu: diameter batang, tumbuh cepat dan vigor, daun hijau saat panen (stay green), anakan produktif banyak, dan produktivitas tanaman utama tinggi. Seleksi dengan menggunakan indeks seleksi menghasilkan sembilan genotipe dengan kemampuan meratun baik, yaitu: IPB97-F-13-1-1, IPB107-F-14-4-1, IPB107-F-14-5-1, IPB Batola 6R, Inpago IPB 8G, IPB 4S, IPB Batola 5R, IPB 3S, dan IPB107-F-18.
Hasil ratun genotipe padi dipengaruhi secara nyata oleh interaksi jumlah buku sisa x genotipe x lokasi. Pemotongan tanaman utama dengan menyisakan satu buku menyebabkan komponen hasil dan hasil ratun rendah pada lahan tipe B maupun C. Pemotongan tanaman utama dengan menyisakan tiga buku sisa memberikan hasil terbaik genotipe IPB97-F-13-1-1 dan IPB 3S di lokasi tipe luapan B dan C.
Hasil ratun tidak dipengaruhi oleh interaksi lokasi x waktu pemupukan tanaman utama x waktu panen tanaman utama x genotipe. Hasil panen ratun maupun total hasil (ratun + tanaman utama), berbeda antar tipologi lahan pasang surut. Hasil ratun genotipe IPB107-F-18 lebih tinggi jika tanaman utama dipupuk dan dipanen lebih awal baik di lahan tipe B maupun C. Panen tanaman utama lebih awal lima hari dari umur panen normal di lokasi Kapuas memberikan total hasil tanaman utama dan ratun tertinggi yaitu 5.12 t ha-1 GKG dan terendah 4.34 t ha-1 GKG jika genotipe ditanam di Pulang Pisau. Panen tanaman utama yang lebih lambat dari waktu panen normal di Kapuas memberikan hasil yang rendah.
Tanaman ratun sensitif terhadap pengaruh lingkungan. Genotipe IPB97-F-13-1-1, IPB 4S, dan IPB 3S merupakan genotipe terbaik berdasarkan rata-rata hasil tanaman utama+ratun di tiga lokasi. Pada lingkungan yang sesuai seperti Kapuas, IPB 3S dapat memberikan total hasil 7.09 t ha-1 GKG. Berdasarkan total hasil, genotipe IPB97-F-13-1-1 dan IPB 4S beradaptasi pada lingkungan sub-optimal (bi < 1). Hasil rata-rata ratun IPB97-F-13-1-1, IPB 4S, dan IPB 3S menempati peringkat terbaik sekaligus di 3 lokasi. Dengan menggunakan genotipe ratun berdaya hasil tinggi dan stabil, teknologi ratun mampu meningkatkan secara signifikan hasil padi di lahan pasang surut. Kontribusi ratun terhadap hasil padi berkisar 31.3 % - 61.9 %.
Penelitian ini menghasilkan beberapa informasi, yaitu: 1) karakter meratun genotipe (kemampuan tunas untuk tumbuh 3 hari setelah panen tanaman utama) adalah sama pada lahan tipe luapan B dan C, tetapi hasil ratun dipengaruhi oleh lingkungan yang ditunjukkan oleh interaksi yang nyata genotipe x lingkungan; 2) metode seleksi padi ratun menggunakan kriteria seleksi batang tanaman utama yang besar; 3) ideotipe padi ratun tipe baru; 4) teknik budidaya ratun spesifik lahan pasang surut; 5) genotipe padi ratun yang stabil; 6) hubungan karakter N batang terhadap hasil ratun; dan 7) informasi aksi gen dalam pemuliaan ratun.
Penelitian lebih lanjut perlu untuk mengetahui penyebab kehilangan rumpun, potensi memperoleh genotipe genjah yang mengandung N dan karbohidrat tinggi, dan respon petani dan nilai ekonomi padi ratun.
Tidal land in Indonesia has an area of 20.1 million hectares, 9.53 million ha of which the potential for agricultural, and from the 9.53 million ha mentioned, 6 million ha are potential for food crop (especially rice). With an area 6 million ha, tidal land should be able to become a major rice production centers in Indonesia. However, efforts to increase the production within the area of tidal land faced with the problem suboptimal environment for plant growth, less of labor, social, cultural, poverty, and the fanaticism of farmers to local varieties.
Rice production in tidal land is still low because the dominant varieties grown by farmers is low yielding local varieties and planting season only once a year. Rice productivity in Central Kalimantan 3.2 t ha-1 and in South Kalimantan 4.2 t ha-1.
Rice production in tidal land can actually be improved without the twice planting season a year, that is by utilizing ratoon. Ratoon are plants that grow from buds contained in the node remaining rice straw at harvest. Research of tidal rice ratoon in Indonesia is still very limited and the reports of breeding ratoon tidal rice has not been found.
The ratooning ability of the rice plant can determined by genetic and environmental characteristics, such as sunlight, temperature, water availability, soil fertility, and pests and plant diseases. Moreover, it can also be influenced by cutting height, fertilizing, water management, and harvesting time. The ratooning ability is a characters of variety. The expression of character of ratoon is controlled genetically and therefore different in different types of rice. According to Turner and Fund (1993), the factors that influence the yield of ratoon quite clear yet known.
It is necessary to assess the ratooning ability of tidal land rice to give the direction of ratoon rice breeding. Research of ratooning ability and the performance of cultivation technology of tidal rice ratoon and the opportunities of a new type of rice ratoon assembly specific for tidal land is supposed to give information the rice that have character ratoon, cultivation techniques that expressed ratoon character, and character that should be corrected so that the yield of the ratoon are not different sharply of the main crop. The research will initiate the discovery of a high yielding new type rice ratoon and adapted well in the tidal land. It required information of sources ratoon character, the character most associated with ratoon that can be used as selection criteria, heritability, gene control, inheritance, stability, and cultivation techniques..
The results showed that the rice ratooning ability can be seen from the ability of bud in node to grow a few days after main crop harvested. Response of genotypes shown by the number of ratoon living clumps did not differ between the typology of acid sulfate soil or peat land with the overflow type B or C. The different of soil environment does not cause different in ratooning ability of rice. Selection can be done on acid sulfate soil and peatlands with overflow type B and C. Genotype IPB107-F-7-3 was able to produce the higest living clumps of 86.6%.
develop rice ratoon need to consider the character stay green and genotypes that are responsive to fertilizer N. The ratooning ability of tidal swamp rice is controlled by two major genes with epistatic gene action. The agronomic characters of ratoon is controlled by many genes with additive gene action and there is the influence of both the duplicate and complementary epistasis.
Interaction of location x number of node left x genotype was highly significant (P <0.01) for ratoon yield. The treatment one node left at harvest showed lowest yield and yield components of ratoon on land that is not waterlogged or flooded land. Stagnant water in overflow type B causes more ratoon buds dead at lower cutting. Genotype IPB97-F-13-1-1 and IPB 3S gave the highest yield on 3 node left in all locations. The potential rice ratoon genotypes grown in a suitable environment and cut at three node could give a total yield of 5.85 t ha-1 dry milled grain. Contributions of ratoon yield was 46-66% of the main crops.
Interaction effect of location x fertilizer application time of main crop x harvest time of main crop x genotype was not significant on ratoon yields. The ratoon yield was higher if the main crop fertilized on 23 daf in overflow type C and 23-30 daf in overflow type B, harvested 25-30 daf, and using genotype IPB107-F-18. The combination of Kapuas with main crop harvest 25 daf provides the highest total yield 5.12 t ha-1 dmg and the lowest if harvested 30-35 daf. Late harvest time is not recommended in the dry land with overflow type C.
Ratoons were sensitive to environmental change. Genotype IPB97-F-13-1-1, IPB 4S, and IPB 3S gave main crop+ratoon yield 5.26 t/ha, 5.14 t/ha, and 5.64 t/ha dry milling grain (DMG) respectively. Based on yield of main crop+ratoon, IPB97-F-13-1-1 and IPB 4S were adaptive to suboptimum condition (bi<IPB97-F-13-1-1). Ratoon yield of genotype IPB97-F-13-1-1, IPB 4S, and IPB 3S were the highest in three locations. The ratoon contribution to rice yield was from 31.3% to 61.9% of main crop.
This research resulted in some information, namely: 1) the ratooning ability genotype (the ability to grow shoots 3 days after the main crop harvest) is similar on some tidal land tipology with overflowing B and C, but the yield of ratoon are influenced by environmental indicated by the significant interaction genotype x environment; 2) selection method using the large main plant culm as selection criteria; 3) ideotype of new plant type rice ratoon; 4) cultivation techniques of ratoon specific in tidal land; 5) The stable rice ratoon genotypes; 6) The relation of N content in the culm to the yield of ratoon; and 7) the action of genes in breeding of ratoon.
Some research will be conducted to determine the relationship of carbohydrate with a node position in order to clarify many researchers who expressed the opinion that carbohydrate is important for ratooning ability of rice. Further research is necessary to determine the cause of the loss clumps, to find the early maturing genotypes containing high N and carbohydrate, and the response of farmers and rice ratoon economic value.
© Hak Cipta Milik Institut Pertanian Bogor, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
PARLIN HALOMOAN SINAGA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup Disertasi:
1. Prof. Dr. Ir. Munif Ghulamahdi, MS
(Staf Pengajar Pada Departemen Agronomi dan Hortikultura IPB Bogor)
2. Dr. Desta Wirnas, SP. MSi
(Staf Pengajar Pada Departemen Agronomi dan Hortikultura IPB Bogor)
Penguji Luar Komisi pada Ujian Terbuka Disertasi:
1. Prof. Dr. Ir. Bambang Sapta Purwoko, MSc.
(Staf Pengajar Pada Departemen Agronomi dan Hortikultura IPB Bogor)
2. Prof. Dr. Ir. Masganti, MS.
Nama : Parlin Halomoan Sinaga
NIM : A263100061
Disetujui oleh
Komisi Pembimbing
Dr.Ir Hajrial Aswidinnoor, MSc Ketua
Prof. Dr. Ir. Didy Sopandie, M.Agr Dr. Ir. Trikoesoemaningtyas, MSc.
Anggota Anggota
Diketahui oleh
Ketua Program Studi Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Yudiwanti Wahyu EK, MS Dr. Ir. Dahrul Syah, M.Sc.Agr
Sang Penyelamat yang selalu memberkahi sehingga penelitian berjudul “Analisis Genetik Potensi Ratun Genotipe Padi (Oryza sativa L) Spesifik Lahan Pasang Surut” dapat selesai dengan baik.
Penelitian dan pengembangan padi ratun merupakan salah satu upaya menyelesaikan masalah stagnasi produksi padi lahan pasang surut akibat kesulitan meningkatkan intensitas pertanaman. Penemuan genotipe padi yang memiliki karakter ratun baik akan berpengaruh positif terhdadap produksi padi di lahan pasang surut. Disertasi ini adalah salah satu bentuk kepedulian terhadap perbaikan sistem usahatani, perekonomian petani, dan ketahanan pangan nasional.
Penulis menyampaikan penghargaan dan ucapan terima kasih kepada:
1. Dr. Ir. Hajrial Aswidinnoor, MSc., Prof. Dr. Ir. Didy Sopandie, MAgr. dan Dr. Ir. Trikoesoemaningtyas, MSc. selaku komisi pembimbing atas bimbingan, motivasi, dan masukan ilmu selama penyusunan proposal, pelaksanaan penelitian, dan penulisan disertasi.
2. Prof. Dr. Ir. Munif Ghulamahdi, MS. dan Dr. Desta Wirnas, SP. MP. Sebagai penguji luar komisi pada Pra Kualifikasi dan Ujian Tertutup.
3. Prof. Dr. Ir. Bambang Sapta Purwoko, MSc. dan Prof. Dr. Ir. Masganti, MS selaku penguji luar komisi pada Ujian Terbuka.
4. Seluruh staf pengajar di Program Studi Pemuliaan dan Bioteknologi Tanaman IPB yang telah membekali penulis dengan berbagai ilmu yang berguna.
5. Kepala Badan Penelitian dan Pengembangan Pertanian yang telah memberikan penugasan kepada penulis untuk melanjutkan pendidikan.
6. Dr. Ir. Ali Jamil, MP. yang telah mengajukan penulis sebagai petugas belajar
7. Prof. Dr. Ir. Masganti, MS. dan Dr.Ir. Kasdi Subagyono, MSc. yang telah membantu dan meringankan beban penulis selama melaksanakan penelitian.
8. Dr. Ir. Hajrial Aswidinnoor, MSc. dan Supartopo yang telah menyediakan materi genetik untuk diuji.
9. Pelaksana teknis Bapak Ajum Ali Akbar, Bapak Tumiran, dan Ibu Lila yang telah membantu pelaksanaan penelitian di lapangan.
10.Ibu Ika Malikhah yang telah membantu menganalisis karbohidrat dan tim laboratorium Balai Penelitian tanah yang telah membantu menganalisis tanah dan nitrogen batang padi.
11.Ibu Dorlan Sipahutar, Emisari Ritonga, Marsid Jahari, dan Usman yang sudah membantu selama penulis melaksanakan studi.
penulis melaksanakan pendidikan.
Akhirnya, semoga informasi dari penelitian ini dapat membuka wacana bagi pemuliaan padi ratun dan menjadi bahan pertimbangan bagi peningkatan produksi padi di lahan pasang surut. Semoga Tuhan yang memiliki segala ilmu yang baik melimpahkan berkahNya.
Bogor, April 2015
Hal
DAFTAR TABEL xvii
DAFTAR GAMBAR xix
DAFTAR LAMPIRAN xxi
PENDAHULUAN
Latar Belakang 1
Tujuan Penelitian 4
Manfaat Penelitian 5
Kebaruan Penelitian 5
Ruang Lingkup Penelitian 5
RESPON GENOTIPE PADI TERHADAP LINGKUNGAN SELEKSI DI LAHAN PASANG SURUT
Pendahuluan 8
Bahan dan Metode 10
Hasil 11
Pembahasan 14
Simpulan 15
KERAGAMAN GENETIK DALAM MENGHASILKAN RATUN 68 GENOTIPE PADI
Pendahuluan 17
Bahan dan Metode 18
Hasil 20
Pembahasan 33
Simpulan 37
DASAR FISIOLOGI DAN AKSI GEN KARAKTER POTENSI MERATUN GENOTIPE PADI
Pendahuluan 39
Bahan dan Metode 40
Hasil 44
Pembahasan 71
Simpulan 75
PENGARUH GENOTIPE DAN JUMLAH BUKU SISA PADA PEMOTONGAN TANAMAN UTAMA TERHADAP KARAKTER RATUN DI LAHAN PASANG SURUT
Pendahuluan 77
Bahan dan Metode 78
Hasil 79
Pembahasan 84
RESPON RATUN GENOTIPE PADI TERHADAP WAKTU PANEN TANAMAN UTAMA DAN WAKTU PEMBERIAN PUPUK DI LAHAN PASANG SURUT
Pendahuluan 86
Bahan dan Metode 88
Hasil 89
Pembahasan 96
Simpulan 97
KERAGAAN RATUN GALUR-GALUR HARAPAN PADI PASANG SURUT DI TIGA LINGKUNGAN TUMBUH
Pendahuluan 99
Bahan dan Metode 100
Hasil 102
Pembahasan 108
Simpulan 109
PEMBAHASAN UMUM 109
SIMPULAN
SARAN
113
114
DAFTAR PUSTAKA 115
LAMPIRAN 125
DAFTAR TABEL
Hal
2.1 Pengaruh genotipe padi terhadap jumlah anakan produktif tanaman utama
12
2.2 Pengaruh genotipe padi dan lingkungan terhadap hasil panen
tanaman utama 12
2.3 Pengaruh genotipe padi terhadap jumlah rumpun hidup ratun pada
umur 3-7 hari setelah tanaman utama dipanen 13
3.1 Ragam fenotipe, genetik, dan heritabilitas karakter 68 genotipe padi. 20 3.2
3.3
Keragaan tanaman utama 68 genotipe padi pada tahap penyaringan di Dramaga-Bogor, 2012.
Koefisien keragaman genetik karakter ratun dari 68 genotipe padi
22 26
3.4 Keragaan ratun 68 genotipe padi di Bogor tahun 2012. 27
3.5 Indeks seleksi 9 genotipe ratun berdasarkan karakter tanaman utama dan pengaruh langsung karakter tanaman utama terhadap
rumpun hidup dan hasil panen sembilan genotipe padi 44 4.2 Hubungan karbohidrat, nitrogen, dan diameter batang tanaman utama
dengan jumlah rumpun hidup dan hasil ratun tanaman ratun padi. 45 4.3 Keragaan tanaman utama genotipe padi Siam Narsis, IPB97-F-13-1-1 46 4.4 Keragaan genotipe padi Inpara 2, IPB97-F-13-1-1, IPB107-F-14-4-1,
dan Inpago IPB 8G 46
4.5 Keragaan ratun padi Siam Narsis dan IPB97-F-13-1-1 47
4.6
4.7
4.8
Keragaan ratun padi Inpara 2, IPB97-F-13-1-1, IPB107-F-14-4-1, dan Inpago IPB 8G
Pola pewarisan dan aksi gen karakter meratun pada padi generasi F2 dari empat pasang persilangan antara padi ratun dengan non ratun Resume hasil analisis skewness dan kurtosis berbagai karakter dari empat pasang persilangan padi
47
48
71 5.1 Pengaruh kombinasi dua faktor perlakuan terhadap tanaman utama
dan ratun padi 81
5.2
6.1
Pengaruh kombinasi lokasi, jumlah buku sisa, dan genotipe terhadap tanaman utama dan ratun padi
Interaksi waktu panen tanaman utama padi dengan lokasi
82 90 6.2
6.3
6.4
Pengaruh tunggal faktor lokasi, waktu pemupukan, waktu panen tanaman utama, dan genotipe terhadap tinggi, jumlah anakan produktif, jumlah rumpun hidup, diameter batang, hasil ratun padi Pengaruh waktu pemupukan tanaman utama terhadap hasil ratun padi di Kapuas dan Pulang Pisau
Efek genotipe dan interaksi lokasi x waktu panen terhadap total hasil panen padi
92
93
94 6.5 Hasil tanaman utama dan ratun tiga genotipe padi yang diperlakukan
dengan waktu pemupukan dan waktu panen tanaman utama 95
7.1 Karakteristik fisika dan kimia tanah di lokasi penelitian 103
7.2 Uji beda nyata hasil tanaman utama dan ratun 10 genotipe padi di
tiga lingkungan tumbuh berdasarkan uji Tuckey 105
7.3 Nilai parameter-parameter stabilitas hasil tanaman utama 10 genotipe
padi yang ditanam di tiga tipologi lahan pasang surut. 106
7.4 Nilai parameter-parameter stabilitas hasil tanaman utama + ratun 10
genotipe padi. 106
DAFTAR GAMBAR
Hal
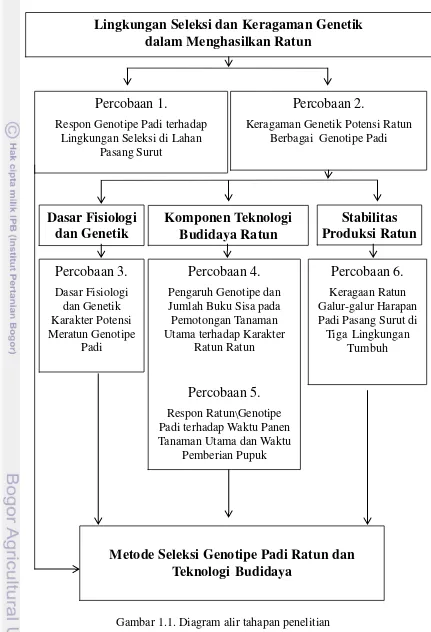
1.1 Diagram alir tahapan penelitian 6
2.1 Penampilan rumpun hidup genotipe padi 3 – 7 hari setelah panen
3.2 3.3
Keragaan ratun padi yang tumbuh dari nodal rentan terhadap rebah Keragaan genotipe padi yang tidak dapat menghasilkan ratun
26 26 3.4 Ideotipe ratun padi tipe baru yang diharapkan adalah masak
serentak dan tinggi seragam 29
3.5 Pengaruh langsung dan tidak langsung tujuh karakter tanaman
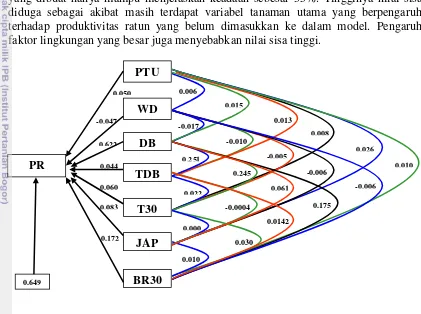
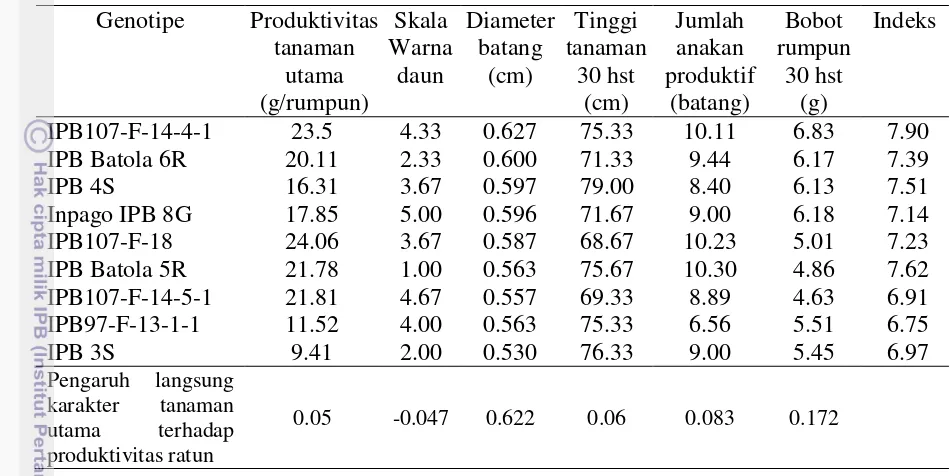
utama terhadap produktivitas ratun padi. 30
3.6 Karakter daun stay green dan non stay green genotipe padi. 31 3.7 Keragaan genotipe padi ratun yang ditanam pada kondisi terkontrol
dapat menghasilkan ratun hingga dua kali, berbeda dengan padi non
ratun tidak menghasilkan ratun. 33
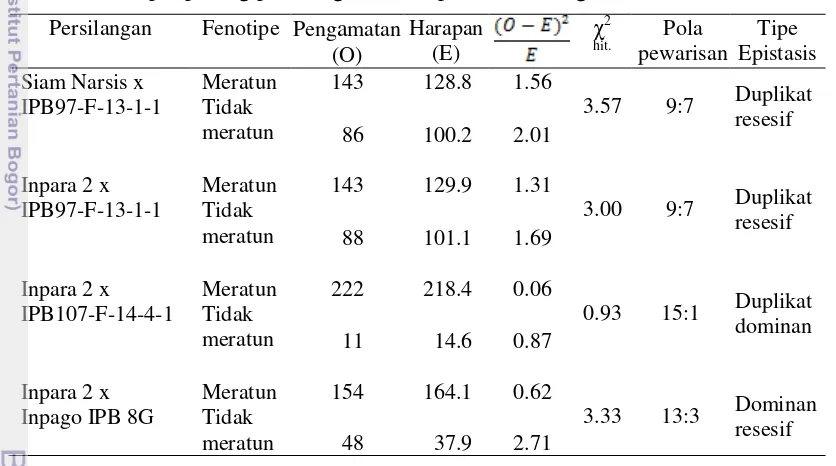
4.1 Sebaran tinggi tanaman ratun padi populasi F2 hasil persilangan
Siam Narsis x IPB97-F-13-1-1 50
4.2 Sebaran tinggi tanaman ratun padi populasi F2 hasil persilangan
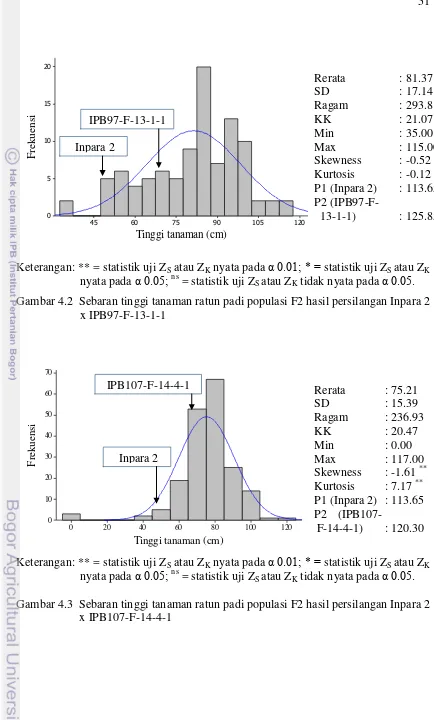
Inpara 2 x IPB97-F-13-1-1 51
4.3 Sebaran tinggi tanaman ratun padi populasi F2 hasil persilangan
Inpara 2 x IPB107-F-14-4-1 51
4.4
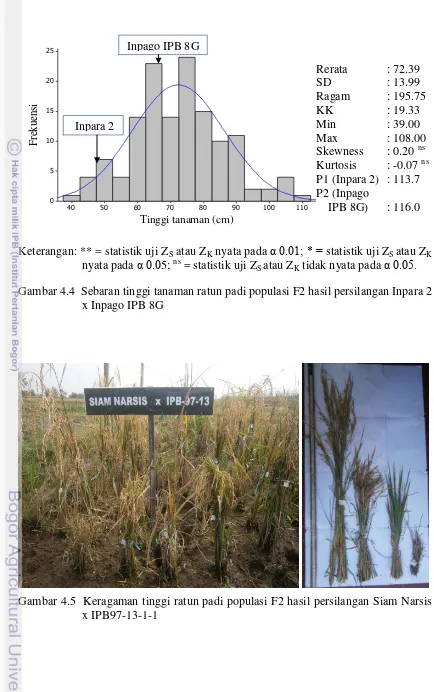
4.5
Sebaran tinggi tanaman ratun padi populasi F2 hasil persilangan Inpara 2 x Inpago IPB 8G
Keragaan tinggi ratun padi populasi F2 hasil persilangan Siam Narsis x IPB97-13-1-1
52
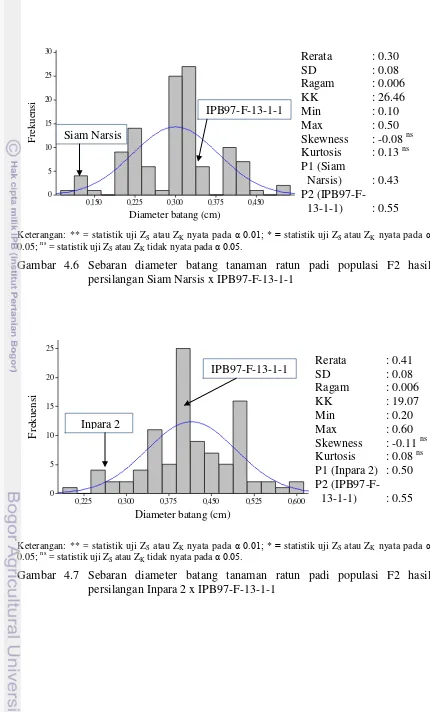
52 4.6 Sebaran diameter batang tanaman ratun padi populasi F2 hasil
persilangan Siam Narsis x IPB97-F-13-1-1 54
4.7 Sebaran diameter batang tanaman ratun padi populasi F2 hasil
persilangan Inpara 2 x IPB97-F-13-1-1 54
4.8 Sebaran diameter batang tanaman ratun padi populasi F2 hasil
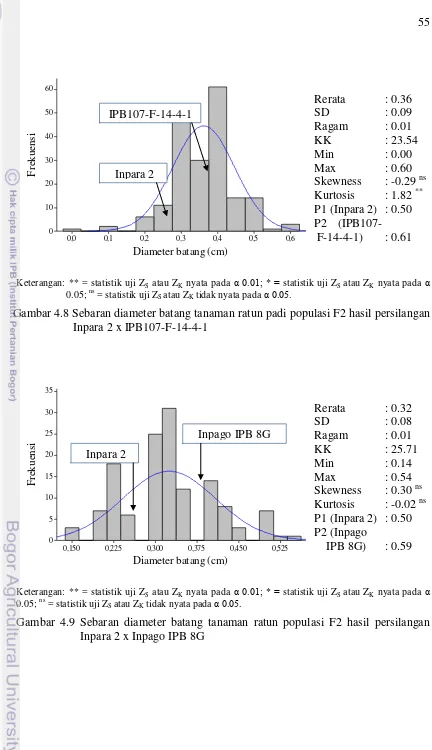
persilangan Inpara 2 x IPB107-F-14-4-1 55
4.9 Sebaran diameter batang tanaman ratun padi populasi F2 hasil
persilangan Inpara 2 x Inpago IPB 8G 55
4.10 Sebaran jumlah anakan produktif tanaman ratun padi populasi F2
hasil persilangan Siam Narsis x IPB97-F-13-1-1. 57
4.11 Sebaran jumlah anakan produktif tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x IPB97-F-13-1-1 57
4.12 Sebaran jumlah anakan produktif tanaman ratun padi populasi F2
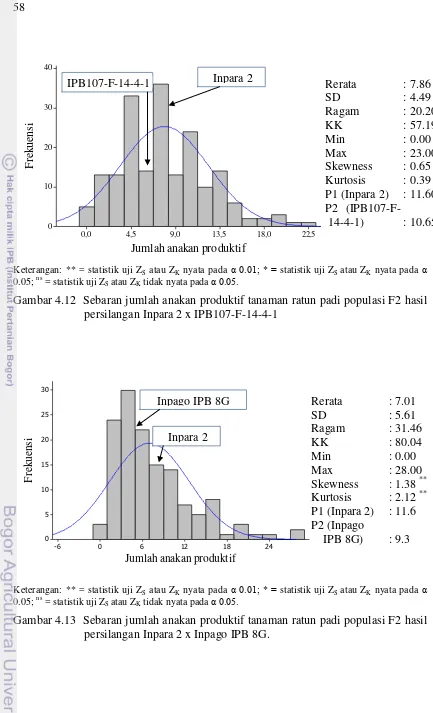
hasil persilangan Inpara 2 x IPB107-F-14-4-1 58
4.13 Sebaran jumlah anakan produktif tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x Inpago IPB 8G 58
4.14 Sebaran bobot jerami per rumpun tanaman ratun padi populasi F2
hasil persilangan Siam Narsis x IPB97-F-13-1-1 60
4.15 Sebaran bobot jerami per rumpun tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x IPB97-F-13-1-1 60
4.16 Sebaran bobot jerami per rumpun tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x IPB107-F-14-4-1 61
4.17 Sebaran bobot jerami per rumpun tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x Inpago IPB 8G 61
4.18 Sebaran bobot gabah per rumpun tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x IPB107-F-14-4-1 63 4.21 Sebaran bobot gabah per rumpun tanaman ratun padi populasi F2
hasil persilangan Inpara 2 x Inpago IPB 8G 64
4.22
4.23
Sebaran bobot jerami+gabah/rumpun tanaman ratun padi populasi F2 hasil persilangan Siam Narsis x IPB97-F-13-1-1
Keragaman kemampuan menghasilkan biji pada tanaman ratun padi F2 hasil persilangan Siam Narsis x IPB97-F-13-1-1
64
66 4.24 Sebaran bobot jerami+gabah/rumpun tanaman ratun padi populasi
F2 hasil persilangan Inpara 2 x IPB97-F-13-1-1 66
4.25 Sebaran bobot jerami+gabah/rumpun tanaman ratun padi populasi
F2 hasil persilangan Inpara 2 x IPB107-F-14-4-1 67
4.26 Sebaran bobot jerami+gabah/rumpun tanaman ratun padi populasi
F2 hasil persilangan Inpara 2 x Inpago IPB 8G 67
4.27 Sebaran indeks panen tanaman ratun padi populasi F2 hasil
persilangan Siam Narsis x IPB97-F-13-1-1 69
4.28 Sebaran indeks panen tanaman ratun padi populasi F2 hasil
persilangan Inpara 2 x IPB97-F-13-1-1 69
4.29 Sebaran indeks panen tanaman ratun padi populasi F2 hasil
persilangan Inpara 2 x IPB107-F-14-4-1 70
4.30 Sebaran indeks panen tanaman ratun padi populasi F2 hasil
persilangan Inpara 2 x Inpago IPB 8G 70
5.1 Respon genotipe padi terhadap jumlah buku yang disisakan saat panen pada dua tipologi lahan pasang surut yang diamati pada
jumlah anakan produktif 80
5.2 Keragaman tinggi buku kedua genotipe padi dalam satu rumpun. 83
5.3
5.4
Pengaruh jumlah buku sisa terhadap jumlah rumpun hidup, hasil ratun, dan total hasil padi di Pelalawan dan Pulang Pisau
Perbandingan rumpun hidup ratun padi pada perlakuan 1 buku dan 3 buku pada lahan tipe luapan B di Kabupaten Pulang Pisau
83
84 7.1 Peringkat 4 genotipe padi di tiga lokasi berdasarkan hasil tanaman
utama dan ratun. 104
7.2 Biplot AMMI 2 hasil tanaman utama, ratun , dan tanaman utama + ratun padi dengan tingkat kesesuaian masing-masing 78,9%, 93%,
Hal
1 Keragaan tanaman utama 68 genotip padi pada tahap penyaringan
di Dramaga-Bogor tahun 2012 (lanjutan Tabel 3.2). 125
2 3
4 5
6
7
8
9
Keragaan ratun 68 genotipe padi di Bogor tahun 2012.
Koefisien korelasi karakter-karakter tanaman utama dan hasil ratun padi
Penyaringan genotipe berpotensi ratun di Bogor tahun 2012 Lokasi penelitian di Desa Danda Jaya, Kecamatan Rantau Badauh, Kabupaten Barito Kuala, Provinsi Kalimantan Selatan
Lokasi penelitian di Desa Petak Batuah, Kecamatan Kapuas Murung, Kabupaten Kapuas, Provinsi Kalimantan Tengah Keragaan ratun galur-galur padi tipe baru di Desa Petak Batuah, Kecamatan Kapuas Murung, Kabupaten Kapuas, Provinsi Kalimantan Tengah
Keragaan ratun di Desa Mentaren, Kecamatan Kahayan Hilir, Kabupaten Pulang Pisau, Kalimantan Tengah
Lokasi penelitian di Desa Sungai Solok, Kecamatan Kuala Kampar, Kabupaten Pelalawan, Provinsi Riau
127
129 130
130
131
132
133
Penulis lahir di Parapat, Sumatra Utara pada 03 Maret 1973 dari Bapak J. Sinaga dan Ibu H. Sirait. Pendidikan sarjana diselesaikan di Jurusan Agronomi Fakultas Pertanian Universitas Riau pada tahun 1997. Pada tahun 1999 - 2000 memperoleh kesempatan mengikuti program pendidikan penyetaraan S1 non pemuliaan tanaman menjadi S1 pemuliaan tanaman di Universitas Padjadjaran Bandung. Pada tahun 2003 menyelesaikan pendidikan Magister Pertanian Bidang Kajian Pemuliaan Tanaman di Universitas Padjadjaran Bandung.
1
PENDAHULUAN
Latar Belakang
Produktivitas padi di Indonesia sudah lebih tinggi jika dibandingkan dengan produktivitas padi di beberapa negara penghasil padi di Asia (Wurjandari dan Syam 2007), tetapi masih berpeluang ditingkatkan agar dapat memenuhi tingkat konsumsi dalam negeri, ekspor, dan mempertahankan ketahanan pangan jangka panjang. Menurut BPS (2014), produktivitas dan produksi padi nasional tahun 2013 masing-masing adalah 5.15 t ha-1 dan 71 291 494 t gabah kering giling (GKG) atau setara beras 42 774 896 t. Tahun 2014 produktivitas diramalkan hanya 5.13 t ha-1 (GKG) dan produksi sebesar 70 607 231 t GKG (setara beras 42 364 339 t) (BPS 2015a).
Jumlah penduduk Indonesia tahun 2010 sebanyak 237 641 326 jiwa dan laju pertambahan penduduk 1.49 % per tahun (BPS 2015b). Pada tahun 2014 penduduk Indonesia sebesar 252 juta jiwa sehingga kebutuhan akan beras dengan jumlah konsumsi 139 kg per kapita per tahun adalah 35 028 000 ton. Dengan demikian Indonesia surplus beras 7 336 339 t tahun 2014.
Kondisi tersebut kemungkinan tidak dapat dipertahankan karena sistem usahatani padi di Indonesia sangat rentan terhadap perilaku musim dan alih fungsi lahan yang menyebabkan pemerintah harus mengimpor beras saat tertentu. Menurut Sopandie (2006), ketergantungan impor komoditas pangan termasuk beras yang relatif tinggi dan melambatnya laju peningkatan produktivitas menunjukkan bahwa persoalan yang menghadang di depan adalah tantangan meningkatkan kapasitas produksi pangan nasional secara berkelanjutan.
Penurunan luas lahan sawah produktif di Pulau Jawa turut mempengaruhi produksi beras nasional setiap tahun. Posisi Pulau Jawa sebagai lumbung pangan akan bergeser dalam jangka panjang. Hasil penelitian Irawan dan Friyatno (2005) menunjukkan bahwa dalam kurun waktu 18 tahun (1981-1998) di Jawa telah terjadi pengurangan lahan sawah seluas 1 juta hektar atau rata-rata sekitar 55 ribu hektar per tahun tanpa memperhitungkan penambahan luas sawah akibat pencetakan sawah baru atau berkurang sekitar 27 ribu hektar per tahun karena adanya pencetakan sawah baru. Alih fungsi lahan yang paling intensif di Pulau Jawa terjadi di dataran rendah (Verburg et al. 1999) dimana sawah berada dalam porsi yang luas.
Kalimantan seluas 5.939 juta ha, Papua seluas 6.415 juta ha, Sulawesi seluas 371 ribu ha, dan di Maluku serta Nusa Tenggara seluas 237 ribu ha.
Peningkatan produksi dan produktivitas padi di wilayah pasang surut akan berdampak besar terhadap ketahanan pangan dan perekonomian nasional, tetapi peningkatan produksi masih sulit dilakukan karena terhambat oleh: 1) kondisi alam yang suboptimal, 2) sosial budaya petani, 3) kemiskinan, 4) kekurangan tenaga kerja, dan 5) fanatisme petani terhadap kultivar lokal. Banjir pasca musim panen besar di lahan rawa (Adigbo et al. 2012) atau kekeringan menyebabkan tanah tidak dapat ditanami. Lima masalah tersebut menyebabkan usaha peningkatan intensitas pertanaman (IP) dari satu kali setahun menjadi lebih dari satu kali cukup sulit dilakukan. Menurut Aswidinnoor et al. (2008), kondisi tenaga kerja dan modal yang terbatas dengan lahan garapan yang luas per rumah tangga, serta adanya perasaan puas akan hasil yang diperoleh menyebabkan petani tidak dapat menanam padi dua kali setahun. Petani transmigran yang didatangkan dari Pulau Jawa dan wilayah lainnya yang terbiasa dengan usahatani intensif menggunakan varietas unggul, akhir-akhir ini justeru banyak yang beralih menanam padi lokal seperti Siam yang hanya ditanam sekali setahun. Petani sangat sedikit yang tetap bertahan dengan pola tanam 2 kali setahun menggunakan varietas unggul yang dilepas pemerintah dengan alasan hasil yang diperoleh sudah melebihi konsumsi rumah tangga dan jika dijual harganya sangat rendah dibandingkan harga jual padi lokal, sehingga tidak cukup untuk mengembalikan modal usahatani.
Salah satu cara untuk meningkatkan produksi beras di wilayah-wilayah yang sulit mengadopsi peningkatan IP dan varietas unggul baru (VUB) adalah menerapkan sistem budidaya ratun dan merakit varietas unggul padi tipe baru (PTB) yang memiliki kemiripan dengan kultivar lokal yang biasa ditanam petani. Pada umumnya petani padi lokal di lahan rawa pasang surut Kalimantan sudah mengenal istilah ratun atau singgang (Jawa) / turiang (Sunda) yaitu tanaman padi yang tumbuh dari batang sisa panen (Harrell et al. 2009) tetapi tidak banyak yang memanfaatkannya sebagai sumber penghasilan karena hasil ratun biasanya sedikit. Hasil ratun yang rendah ini berhubungan dengan kultivar yang dominan ditanam petani yaitu kultivar lokal berdaya hasil rendah. Dengan pengenalan PTB yang memiliki potensi ratun baik dan sifat berasnya mirip dengan kultivar lokal, diharapkan usaha peningkatan produksi akan lebih mudah.
Beberapa keuntungan dari penerapan ratun antaralain: 1) biaya produksi lebih rendah karena tidak perlu pengolahan tanah dan penanaman ulang, 2) pupuk yang dibutuhkan lebih rendah (½ dari dosis yang diberikan pada tanaman pertama), 3) umur panen lebih pendek, 4) hasil yang diperoleh dapat mencapai 66 persen dari tanaman utama (Flinn dan Mercado 1988), dan 5) tanaman pada sistem ratun menggunakan air 60% lebih sedikit dari tanaman utama (Oad et al. 2002).
Kemampuan tanaman padi untuk menghasilkan ratun dapat ditentukan oleh: sifat genetik dan lingkungan, seperti sinar matahari, suhu, ketersediaan air, tingkat kesuburan tanah, dan keadaan hama dan penyakit tanaman. Selain itu dapat juga dipengaruhi tinggi pemotongan, pemupukan, pengelolaan air (Ichii 1984, de Datta dan Bernasor 1988; Mahadevappa 1988), dan umur panen (Bahar dan de Datta 1977). Kemampuan menghasilkan ratun merupakan karakter varietas. Ekspresi karakter ratun dikendalikan secara genetik dan oleh karena itu berbeda pada berbagai jenis padi (Bahar dan de Datta 1977). Menurut Turner dan Fund (1993), faktor-faktor yang mempengaruhi hasil ratun belum cukup jelas diketahui.
Susilawati et al. (2010) telah melaporkan bahwa dari 18 genotipe padi yang diuji, sembilan genotipe menghasilkan ratun yang baik dengan jumlah anakan 6 – 25 batang per rumpun. Genotipe yang memiliki batang besar, kokoh dan hijau, daun yang lebat, serta tunggul yang tetap vigor dan hijau setelah panen tanaman utama menghasilkan ratun yang baik. Susilawati et al. (2012) membuktikan bahwa pupuk N, P dan K berperan penting dalam meningkatkan komponen hasil dan hasil ratun padi.Genotipe-genotipe memberikan respon yang berbeda terhadap perlakukan pemupukan. Genotipe IPB106-7-47-DJ-1, IPB106-F-8-1 dan varietas hibrida Rokan menghasilkan ratun tertinggi dengan dosis pemupukan (kg ha-1) 45 N + 27 P2O5 sedangkan genotipe Cimelati dan Hipa-5 menghasilkan ratun tertinggi pada dosis pemupukan (kg ha-1) 45 N + 30 K2O.
Ratun sudah cukup lama diketahui dan diteliti, tetapi budidaya ratun kurang diterima di kalangan petani komersial karena hasilnya rendah, kurang tersedianya varietas ratun yang baik, pemasakan yang dipacu oleh penyakit, pemasakan tidak merata, dan kualitas beras yang rendah (Chauhan et al. 1985). Berbagai kelemahan tersebut dapat diperbaiki melalui pemuliaan untuk menciptakan padi ratun tipe baru berdaya hasil tinggi, masak normal dan serentak, serta kualitas beras baik. Potensi untuk perbaikan ratun di lahan pasang surut Kalimantan Tengah dan Kalimantan Selatan cukup besar karena terdapat berbagai kultivar padi yang biasa diratun yang diduga memiliki gen pengendali ratun.
Penelitian ratun padi pasang surut di Indonesia masih sangat terbatas dan belum ditemukan laporan mengenai pemuliaan ratun pada padi pasang surut dan interaksi padi ratun dengan lingkungan. Oleh karena itu, pemuliaan ratun harus dimulai dengan pengenalan karakter yang erat kaitannya dengan ratun sehingga proses pemuliaan lebih terarah. Pengetahuan akan hubungan hasil ratun dengan karakter penting lainnya diperlukan untuk menentukan karakter yang dapat digunakan sebagai kriteria seleksi untuk ratun berdaya hasil tinggi.
berhubungan dengan produktivitas dan diduga terdapat perbedaan kemampuan menghasilkan ratun. Beberapa karakter lain seperti kevigoran, ketebalan batang, kegenjahan, dan sifat daun hijau saat panen (stay green) diduga berhubungan dengan karakter ratun. Karakter-karakter agronomis tersebut dikendalikan secara genetik sehingga dapat dilakukan persilangan untuk menambah keragaman ratun maupun menambahkan karakter ratun kepada padi yang tidak memiliki karakter tersebut. Karakter yang paling dominan mempengaruhi hasil ratun belum diketahui.
Ketebalan batang yang juga merupakan ekspresi dari kevigoran, berkorelasi dengan tingginya kadar karbohidrat dalam jerami. Hal ini dapat menginduksi kemampuan regenerasi ratun yang lebih kuat, menghasilkan ratun yang lebih banyak dan hasil gabah yang lebih tinggi (Palchamy dan Purushothaman 1988). Pertumbuhan ratun padi tergantung pada jumlah total kadar karbohidrat pada pangkal batang, pada awal pertumbuhan. Jumlah karbohidrat yang besar dibutuhkan untuk menghasilkan banyak anakan (Ichii dan Ogaya 1985). Terdapat hubungan antara pertumbuhan batang dengan kemampuan menghasilkan ratun. Pertumbuhan batang yang cepat dan vigor berkaitan dengan jumlah karbohidrat yang banyak pada batang. Tipe pertumbuhan vigor biasanya dimiliki oleh varietas unggul dan varietas hibrida atau kelompok produktivitas tinggi. Hubungan karakter produktivitas tinggi terhadap kemampuannya menghasilkan ratun pada genotipe-genotipe yang vigor masih perlu diteliti untuk mengetahui apakah cadangan karbohidrat di batang tidak habis diserap pada tahap reproduktif untuk menghasilkan biji yang banyak. Menurut Abdullah (2004), tingkat pengisian gabah sangat ditentukan oleh hasil fotosintat (karbohidrat) dalam batang dan daun, dan translokasinya serta akumulasinya dalam gabah.
Potensi ratun pada padi-padi pasang surut perlu dikaji untuk memberi arah pemuliaan padi ratun. Untuk itu diperlukan informasi sumber karakter ratun, karakter yang paling berhubungan dengan ratun yang dapat dijadikan kriteria seleksi, kendali gen, pola pewarisan, maupun stabilitas dari karakter tersebut.
Lahan pasang surut dikelompokkan menjadi empat menurut tipe luapan air, yaitu tipe A, B, C, dan D. Lahan tipe A adalah lahan yang terluapi pada saat pasang besar dan pasang kecil, lahan tipe B hanya terluapi pada saat pasang besar, lahan tipe C tidak terluapi pada saat pasang besar maupun pasang kecil tetapi tinggi permukaan air kurang dari 50 cm, dan lahan tipe D adalah lahan yang tidak terluapi pada saat pasang besar maupun pasang kecil dan tinggi permukaan air lebih dari 50 cm (Widjaja-Adhi et al. 1997). Sehubungan dengan tipe luapan air terdapat 3 hal yang harus diperhatikan dalam budidaya ratun di lahan pasang surut, yaitu: tinggi pemotongan atau jumlah buku yang disisakan pada saat panen, waktu pemupukan, dan waktu panen tanaman utama. Kemampuan tanaman menghasilkan ratun ditentukan waktu panen tanaman utama (Mobasser et al. 2012; Yazdpour et al. 2012) dan pemupukan (Petroudi et al. 2011). Waktu panen dan waktu pemupukan tanaman utama sangat menentukan viabilitas tunas dan penampilan ratun.
Tujuan Penelitian
padi ratun, dan teknologi budidaya padi tipe baru ratun spesifik lahan pasang surut. Tujuan spesifik dari penelitian ini adalah: 1) Menjelaskan respon genotipe-genotipe padi pasang surut terhadap lingkungan seleksi di lahan pasang surut; 2) Menjelaskan keragaman potensi ratun pada berbagai genotipe padi pasang surut; 3) Memperoleh kriteria seleksi untuk padi ratun dari karakter yang berkorelasi dengan potensi ratun; 4) Menjelaskan dasar fisiologi dan genetik karakter ratun pada empat populasi F2 dan heritabilitas karakter ratun padi pasang surut; 5) Menjelaskan pengaruh genotipe dan jumlah buku yang disisakan pada saat panen tanaman utama terhadap hasil ratun; 6) Menjelaskan respon genotipe terhadap waktu panen tanaman utama dan waktu pemberian pupuk dalam menghasilkan ratun; 7) Menjelaskan daya adaptasi genotipe padi ratun pada berbagai agroekosistem pasang surut; dan 8) Memperoleh genotipe padi pasang surut yang mempunyai kemampuan meratun baik.
Manfaat Penelitian
Penelitian ini menghasilkan genotipe padi tipe baru ratun dan memberikan informasi bagi program pemuliaan untuk menghasilkan padi ratun tipe baru yang spesifik lahan pasang surut melalui informasi keberadaan calon tetua pembawa karakter ratun, pewarisan sifat, kriteria seleksi, maupun teknik budidaya yang dapat mengekspresikan karakter ratun. Selanjutnya hasil penelitian ini diharapkan dapat menjembatani program pemerintah untuk meningkatkan intensitas pertanaman dengan sosial budaya petani dan kondisi alam yang kurang baik setelah panen raya. Dengan demikian petani pasang surut dapat berkontribusi pada peningkatan produksi beras nasional.
Kebaruan Penelitian
Penelitian ini menghasilkan beberapa kebaruan, yaitu: 1) informasi tentang interaksi genotipe-genotipe padi dengan lingkungan seleksi terhadap kemampuan menghasilkan ratun; 2) metode seleksi padi ratun menggunakan karakter sekunder; 3) hubungan kandungan N batang terhadap hasil ratun; 4) informasi aksi gen yang mengendalikan ratun; 5) ideotipe padi ratun tipe baru; 6) teknik budidaya ratun spesifik lahan pasang surut; dan 7) genotipe padi ratun yang stabil.
Ruang Lingkup Penelitian
\
Gambar 1.1. Diagram alir tahapan penelitian
Lingkungan Seleksi dan Keragaman Genetik
dalam Menghasilkan Ratun
Percobaan 1.
Respon Genotipe Padi terhadap Lingkungan Seleksi di Lahan
Pasang Surut
Percobaan 2.
Keragaman Genetik Potensi Ratun Berbagai Genotipe Padi
Komponen Teknologi
Budidaya Ratun
Percobaan 4.
Pengaruh Genotipe dan Jumlah Buku Sisa pada Pemotongan Tanaman Utama terhadap Karakter
Ratun Ratun
Percobaan 5.
Respon Ratun Genotipe Padi terhadap Waktu Panen Tanaman Utama dan Waktu
Pemberian Pupuk
Stabilitas
Produksi Ratun
Uji Multilokasi
Percobaan 6.
Keragaan Ratun Galur-galur Harapan Padi Pasang Surut di Tiga Lingkungan
Tumbuh
Dasar Fisiologi
dan Genetik
Metode Seleksi Genotipe Padi Ratun dan
Teknologi
Budidaya
Percobaan 3.
Dasar Fisiologi dan Genetik Karakter Potensi Meratun Genotipe
2
RESPON GENOTIPE PADI TERHADAP LINGKUNGAN
SELEKSI DI LAHAN PASANG SURUT
ABSTRAK
Penelitian ini bertujuan untuk menjelaskan respon genotipe-genotipe padi terhadap lingkungan pasang surut sebagai lokasi seleksi, yang ditunjukkan oleh jumlah rumpun hidup. Penelitian dilaksanakan di Kabupaten Pulang Pisau (sulfat masam, tipe luapan B), Kabupaten Kapuas (sulfat masam, tipe luapan C), Kabupaten Barito Kuala (bergambut, tipe luapan B) pada bulan Mei-Oktober 2011. Genotipe padi IPB107-F-7-3, IPB107-F-13-1, IPB107-F-40E-1, IPB107-F-18-2, IPB107-F-20-5, IPB107-F-34-1, IPB107-F-77-3, IPB107-F-10-1, dan IPB Dadahup 1R ditanam sesuai rancangan acak kelompok yang diulang tiga kali. Bibit berumur 21 hari sejak semai (hss) ditanam 1 bibit per lubang tanam dengan jarak tanam 20 cm x 20 cm. Tanaman dipupuk dengan Urea 150 kg ha-1, SP36 100 kg ha-1, dan KCl 75 kg ha-1. Panen tanaman utama dilakukan setelah 95% malai menguning. Pemotongan batang saat panen dilakukan pada ketinggian 10 cm dari permukaan tanah. Peubah yang diamati dari tanaman utama adalah jumlah anakan produktif dan hasil biji, sedangkan dari tanaman ratun diamati jumlah rumpun hidup yang dicirikan munculnya tunas ratun pada umur 3-7 hari sejak panen tanaman utama. Hasil penelitian menunjukkan bahwa potensi ratun pada padi dapat diketahui dari kemampuan tunas ratun untuk hidup dan tumbuh beberapa hari setelah panen. Respon genotipe-genotipe padi terhadap ketiga lingkungan seleksi berdasarkan tipologi lahan pasang surut tidak berbeda, yang ditunjukkan oleh jumlah rumpun hidup yang sama antar tipologi lahan. Seleksi untuk kemampuan meratun dapat dilakukan pada tipologi lahan sulfat masam maupun lahan bergambut dengan tipe luapan air B dan C. Genotipe IPB107-F-7-3 mampu menghasilkan rumpun hidup terbanyak sebesar 86.6%.
Kata Kunci: ratun, lingkungan, genotipe padi, pasang surut, interaksi
RESPONSE OF RICE GENOTYPES TO SELECTION ENVIRONMENT IN TIDAL LAND
ABSTRACT
days after sowing (das), one seedling per planting hole with a spacing of 20 cm x 20 cm. The plants were fertilized with Urea 150 kg ha-1, SP-36 100 kg ha-1, and KCl 75 kg ha-1. The main crops were harvested after 95% of panicles ripening. Cutting height at harvest was 10 cm from the soil surface. Variables measured from the main crop were the number of productive tillers and grain yield while the characters observed from the ratoon crops was number of living clumps that characterized by the emergence of shoots of ratoon at 3-7 days after the main crop harvested. The results showed that the rice ratooning ability can be seen from the ability of bud in node to grow a few days after main crop harvested. Response of genotypes shown by the number of ratoon living clumps did not differ between the typology of acid sulfate soil or peat land with the overflow type B or C. The different of soil environment does not cause different in ratooning ability of rice. Selection can be done on acid sulfate soil and peatlands with overflow type B and C. Genotype IPB107-F-7-3 was able to produce the higest living clumps of 86.6%.
Keywords: ratoon, environment, rice genotype, tidal land, interaction
PENDAHULUAN
Lingkungan pasang surut beragam berdasarkan tipe luapan air, tipologi lahan, maupun jenis dan kadar unsur beracun. Berdasarkan tipe luapan air dikenal empat tipe lahan pasang surut yaitu tipe A, B, C, dan D. Lahan tipe A adalah lahan yang terluapi pada saat pasang besar dan pasang kecil, lahan tipe B hanya terluapi pada saat pasang besar, dan lahan tipe C tidak terluapi pada saat pasang besar maupun pasang kecil tetapi tinggi permukaan air 50 cm di bawah permukaan tanah, dan lahan tipe D memiliki kedalaman air lebih dari 50 cm di bawah permukaan tanah. Berdasarkan tipologi lahan terdapat empat kategori lahan pasang surut, yaitu: 1) lahan potensial yaitu lahan yang kedalaman piritnya >50 cm di bawah permukaan tanah, 2) lahan sulfat masam, yaitu lahan yang lapisan piritnya 0-50 cm di bawah permukaan tanah, 3) lahan gambut, yaitu lahan yang mengandung lapisan gambut dengan kedalaman beragam minimal 50 cm, 4) lahan salin, yaitu lahan yang mendapat intrusi air laut sehingga mengandung garam dengan konsentrasi tinggi terutama pada musim kemarau (Widjaja-Adhi et al. 1997). Jenis dan kadar unsur beracun seperti Al dan Fe yang beragam sesuai dengan tingkat oksidasi-reduksi menyebabkan lingkungan semakin beragam. Keragaman lingkungan akan semakin tinggi jika di antara komponen-komponen penyebab keragaman tersebut terjadi kombinasi dan interaksi.
Cekaman lingkungan biasanya mempengaruhi pertumbuhan dan hasil tanaman utama padi. Cekaman lingkungan yang berat dapat menyebabkan tanaman utama segera memasuki masa menua lalu mati dan tidak menghasilkan ratun. Lingkungan seperti ini tidak menguntungkan sebagai lingkungan seleksi untuk padi ratun. Tipe lingkungan yang dapat mempertahankan kesegaran batang tanaman utama dan viabilitas tunas ratun masih perlu dikaji.
Keracunan besi dan kekeringan akan mempercepat penuaan tanaman. Lingkungan pasang surut berhubungan dengan dua hal tersebut. Lahan yang mengandung kadar besi tinggi dapat menyebabkan akar diselimuti oleh plak (Wang dan Peverly 1999, Liang et al. 2006), sehingga pada saat panen dimana biasanya terjadi pengeringan lahan, plak yang menutupi akar akan menyebabkan absorpsi air terhambat. Hal ini diduga dapat mengganggu pertumbuhan tunas ratun. Selanjutnya di lahan tipe C periode kering dapat terjadi pada fase penting pertumbuhan tanaman dan berlangsung lebih lama. Kondisi lahan kering dan basah di lahan pasang surut masing-masing memiliki kelebihan dan kekurangan terhadap budidaya padi ratun. Pada lahan tipe luapan C yang kering pada saat panen menyebabkan tajuk tanaman segera mengering, sedangkan pada lahan tipe luapan A dan B tajuk tanaman lebih lambat mengering, tetapi pada lahan yang selalu tergenang, cekaman besi menjadi ancaman karena pada tingkat yang cukup parah tanaman dapat mengalami gejala layu seperti kekurangan air.
Perlakuan budidaya seperti pengeringan lahan menjelang panen diduga berpengaruh terhadap viabilitas tunas ratun. Ada lahan yang mudah dikeringkan menjelang panen seperti lahan tipe luapan C, tetapi ada juga yang tidak dapat dikeringkan seperti lahan tipe luapan A. Bagaimana respon mata tunas genotipe padi terhadap kondisi tersebut masih perlu dipelajari.
Kondisi lingkungan seperti kesuburan tanah (Dev et al. 2013), pemupukan N (Petroudi et al. 2011), padat tanam (Luo et al. 2007; Huang 2011), kondisi iklim (Yang et al. 2011), persiapan lahan, tata air, dan pengendalian hama, penyakit, dan gulma (Santos et al. 2003), mempengaruhi viabilitas tunas dan pertumbuhan ratun. Lingkungan yang sesuai akan menyebabkan batang padi tetap segar pada saat panen dan mata tunas pada buku tetap viable, sehingga tidak lama setelah panen, tunas sudah mulai tumbuh. Menurut Susilawati et al (2010) tunas ratun sudah tumbuh pada hari kedua hingga hari ketujuh sejak panen tanaman utama.
Penentuan lingkungan seleksi yang tepat penting bagi program pemuliaan. Seleksi pada lingkungan optimal akan menyebabkan ekspresi genotipe optimal dan menghasilkan genotipe berdaya hasil tinggi. Seleksi pada lingkungan suboptimal akan menghasilkan genotipe yang daya hasilnya tidak tinggi, tetapi memiliki ketahanan terhadap cekaman lingkungan target.
tanah dan air adalah faktor penting dalam menentukan cepat atau lambat tanaman utama memasuki masa menua dan kuncup ratun menjadi tidak viabel. Penelitian untuk mengetahui lingkungan seleksi dan lingkungan target yang tepat masih perlu dilaksanakan. Penelitian ini bertujuan untuk menjelaskan respon genotipe-genotipe padi terhadap lingkungan pasang surut sebagai lokasi seleksi, yang ditunjukkan oleh jumlah rumpun hidup.
BAHAN DAN METODE
Penelitian dilaksanakan pada lahan petani (on farm trial) di Desa Mentaren 2, Kecamatan Kahayan Hilir, Kabupaten Pulang Pisau, Provinsi Kalimantan Tengah; Desa Petak Batuah, Kecamatan Kapuas Murung, Kabupaten Kapuas, Provinsi Kalimantan Tengah; dan Desa Danda Jaya, Kecamatan Rantau Badauh, Kabupaten Barito Kuala, Provinsi Kalimantan Selatan. Penelitian dilaksanakan pada bulan Mei-Oktober 2011.
Lahan di Kapuas merupakan tanah sulfat masam, pH 3.93 (sangat masam), kadar besi 148 mg kg-1 (sangat tinggi), tipe luapan air C, ditanami dua kali setahun, sudah dibatasi oleh pematang, dan dikelilingi oleh parit yang menjadi sumber air pada saat musim kemarau dengan sistem pompa. Petani melakukan pengolahan tanah sempurna satu kali bajak, satu kali garu, dan satu kali meratakan tanah. Pengendalian hama tikus dengan menggunakan beberapa cara yaitu: pemasangan pagar plastik dan parit air (trap barrier system), umpan beracun, pengemposan, dan pemasangan kawat beraliran listrik di sawah.
Lahan di Barito Kuala adalah lahan pasang surut bergambut, pH 4.23 (sangat masam), kadar besi 255.64 mg kg-1 (sangat tinggi), tipe luapan B, sawah sistem surjan yaitu setiap petak sawah dibatasi oleh pematang yang tinggi dan lebar yang ditanami dengan tanaman tahunan atau sayur-sayuran, sawah ditanami dua kali setahun tetapi petani disekitarnya masih dominan menanam padi satu kali setahun, pengolahan tanah satu kali bajak dan satu kali garu. Sistem pengairan sudah cukup baik dan ketinggian air mudah dikendalikan yaitu dengan membuka dan menutup pintu air. Pengendalian hama tikus menggunakan pagar plastik dan umpan beracun.
Lahan di Pulang Pisau merupakan tanah sulfat masam, tipe luapan C, pH 4.08 (sangat masam), kadar besi 313 mg kg-1 (sangat tinggi), hamparan tanpa pematang sehingga air pasang bebas keluar masuk, ditanami satu kali setahun, pengolahan tanah minimum, dan pengairan mengandalkan luapan air pada saat pasang besar serta air hujan. Pengendalian hama tikus dengan memasang umpan beracun. Di lahan tersebut lazim ditemukan bronzing yaitu daun padi mengalami nekrosis akibat keracunan Fe.
tanaman utama dilakukan setelah 95% malai menguning. Pemotongan batang saat panen dilakukan pada ketinggian 10 cm dari permukaan tanah.
Peubah yang diamati dari tanaman utama adalah jumlah anakan produktif dan hasil dan dari tanaman ratun diamati jumlah rumpun hidup yang dicirikan munculnya tunas ratun pada umur 3 – 7 hari sejak panen tanaman utama. Data dianalisis ragam dan diuji lanjut menurut Tukey.
HASIL
Selama musim tanam tidak terjadi kekeringan di Kapuas dan Barito Kuala tetapi lokasi penelitian di Pulang Pisau mengalami kemarau pada fase vegetatif sehingga air harus dipompa dari saluran sekunder. Tanaman memperoleh air yang cukup hingga musim panen. Kondisi lingkungan secara umum kondusif sehingga tanaman utama tumbuh baik.
Hasil analisis ragam gabungan menunjukkan bahwa interaksi lingkungan dengan genotipe berpengaruh tidak nyata terhadap jumlah anakan produktif dan hasil tanaman utama serta jumlah rumpun hidup ratun. Faktor genotipe berpengaruh sangat nyata terhadap ketiga peubah tersebut. Faktor lingkungan hanya berpengaruh terhadap hasil panen tanaman utama (p<0.01).
Interaksi yang tidak nyata antara genotipe dengan lingkungan berarti tidak terjadi perubahan peringkat suatu genotipe antar lingkungan, baik berdasarkan karakter jumlah anakan produktif dan hasil tanaman utama maupun jumlah rumpun hidup ratun. Genotipe dapat mempertahankan peringkatnya pada lingkungan yang berbeda. Hal ini menunjukkan semua genotipe yang diuji sudah tergolong stabil. Genotipe-genotipe juga sudah menunjukkan adaptasi yang baik di lingkungan pasang surut berdasarkan hasil tanaman utama dan kemampuannya menghasilkan ratun. Semua genotipe dapat ditanam di lahan dengan tipe luapan B dan C, baik lahan bergambut maupun sulfat masam untuk mengetahui kemampuan meratun.
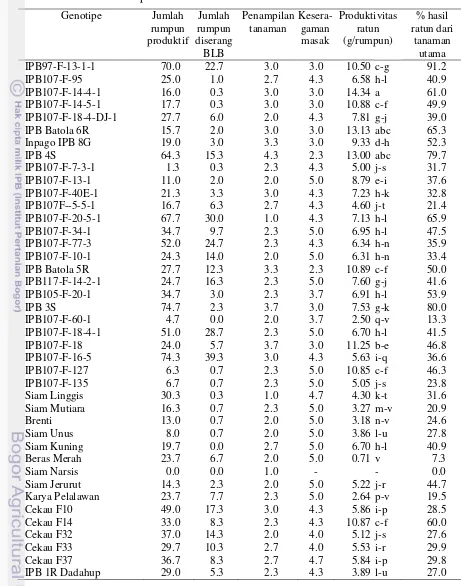
Tabel 2.1 menunjukkan bahwa karakter jumlah anakan produktif tidak berbeda nyata antar lokasi. Secara umum semua genotipe menghasilkan jumlah anakan produktif yang rendah. Jumlah anakan produktif yang tertinggi dihasilkan oleh genotipe IPB107-F-34-1 sebanyak 11.1 anakan. Jumlah anakan produktif penting untuk dua hal, yaitu: menentukan hasil panen tanaman utama dan menentukan jumlah tunas ratun yang akan tumbuh setelah panen tanaman utama. Semakin banyak jumlah anakan produktif diharapkan semakin banyak pula tunas ratun yang dihasilkan. Menurut Hairmansis et al. (2010), jumlah anakan produktif memberikan pengaruh langsung yang positif terhadap hasil gabah
tertinggi 3.4 t ha-1. Di lingkungan Pulang Pisau, adaptasi genotipe tersebut lebih baik dibandingkan genotipe-genotipe lainnya. Lingkungan seleksi mampu memperlihatkan perbedaan karakter hasil panen tanaman utama tetapi tidak mampu memperlihatkan perbedaan jumlah rumpun hidup, yang berarti bahwa ketiga tipologi lingkungan tersebut dapat digunakan sebagai lokasi untuk menyeleksi kemampuan meratun padi.
Tabel 2.1. Pengaruh genotipe padi terhadap jumlah anakan produktif tanaman utama
Genotipe Jumlah anakan produktif Rata-rata
genotipe Kapuas Barito Kuala Pulang Pisau
1. IPB107-F-7-3 7.0 6.8 6.3 6.7 c
2. IPB107-F-13-1 7.6 7.0 7.1 7.2 bc
3. IPB107-F-40E-1 7.4 8.3 7.6 7.8 bc
4. IPB107-F-18-2 7.6 7.6 6.3 7.1 bc
5. IPB107-F-20-5 7.9 7.1 5.7 6.9 bc
6. IPB107-F-34-1 11.9 12.2 9.2 11.1 a
7. IPB107-F-77-3 10.0 8.7 7.8 8.8 abc
8. IPB107-F-10-1 8.4 7.9 10.9 9.1 ab
9. IPB Dadahup 1R 8.3 9.2 7.7 8.4 bc
Rata-rata lingkungan 8.5 8.3 7.6
KK = 19.09%
Keterangan: Angka yang diikuti oleh huruf kecil yang sama pada kolom yang sama berarti tidak berbeda nyata menurut uji Tukey 0.05
Tabel 2.2. Pengaruh genotipe padi dan lingkungan terhadap hasil panen tanaman utama
Genotipe Hasil (t ha-1) Rata-rata
genotipe
Kapuas Barito Kuala Pulang Pisau
1. IPB107-F-7-3 3.4 3.2 1.5 2.7 bc
2. IPB107-F-13-1 3.3 3.0 2.2 2.8 abc
3. IPB107-F-40E-1 3.1 2.7 1.6 2.5 c
4. IPB107-F-18-2 3.4 3.3 2.2 3.0 abc
5. IPB107-F-20-5 3.5 3.1 2.0 2.9 abc
6. IPB107-F-34-1 4.0 3.6 2.3 3.3 ab
7. IPB107-F-77-3 3.5 3.9 2.8 3.4 a
8. IPB107-F-10-1 3.3 3.2 1.2 2.6 c
9. IPB Dadahup 1R 3.0 3.3 2.3 2.9 abc
Rata-rata lingkungan 3.4 a 3.2 a 2.0 b
KK = 15.13%
Cekaman lingkungan yang lebih berat di Pulang Pisau ternyata tidak berpengaruh secara nyata terhadap jumlah rumpun hidup ratun (Tabel 2.3). Tingkat kemasaman tanah, kadar besi, dan tipe luapan air yang berbeda tidak menyebabkan rumpun hidup menjadi lebih sedikit di lokasi yang bercekaman lebih berat. Rata-rata jumlah rumpun hidup di semua lokasi sudah cukup tinggi.
Tabel 2.3. Pengaruh genotipe padi terhadap persentase rumpun hidup ratun pada umur 3-7 hari setelah tanaman utama dipanen
Genotipe
Persentase rumpun hidup Rata-rata
genotipe
Kapuas Barito Kuala Pulang Pisau
1. IPB107-F-7-3 86.3 88.7 84.9 86.6 a
2. IPB107-F-13-1 87.3 79.5 80.4 82.4 a
3. IPB107-F-40E-1 85.9 69.1 74.6 76.6 ab
4. IPB107-F-18-2 85.7 83.6 76.1 81.8 a
5. IPB107-F-20-5 73.5 79.8 74.4 75.9 ab
6. IPB107-F-34-1 80.5 87.2 77.4 81.7 a
7. IPB107-F-77-3 75.3 89.8 88.2 84.4 a
8. IPB107-F-10-1 68.3 74.8 64.8 69.3 b
9. IPB Dadahup 1R 82.9 83.7 72.3 79.6 ab
Rata-rata lingkungan 80.6 81.8 77.0
KK = 9.12 %
Keterangan: Angka yang diikuti oleh huruf kecil yang sama pada kolom yang sama berarti berbeda tidak nyata menurut uji Tukey 0.05
Pengaruh genotipe yang sangat nyata terhadap persentase rumpun hidup menunjukkan adanya perbedaan kemampuan meratun antar genotipe (Tabel 2.3) sebagai akibat adanya keragaman kemampuan mempertahankan viabilitas mata tunas antar genotipe. Hal tersebut menunjukkan kemungkinan memperoleh genotipe ratun yang lebih baik melalui seleksi. Di antara sembilan genotipe yang diuji, hanya satu genotipe yang memiliki persentase rumpun hidup rendah, yaitu IPB107-F-10-1.
Faktor tanah yang berbeda tidak menyebabkan potensi meratun genotipe berbeda jika air cukup tersedia. Lahan tidak harus tergenang untuk menghasilkan tunas ratun. Kadar air tanah pada kapasitas lapang seperti di Pulang Pisau (Gambar 2.1 C) masih mampu menghasilkan ratun yang baik.
Gambar 2.1 Penampilan rumpun hidup genotipe padi pada 3-7 hari setelah panen tanaman utama di Kapuas (A), Barito Kuala (B), dan Pulang Pisau (C).
PEMBAHASAN
Tinggi rendahnya hasil tanaman utama tidak menyebabkan jumlah rumpun hidup ratun antar lokasi berbeda. Hasil tanaman utama lebih rendah di Pulang Pisau tetapi jumlah rumpun yang bertahan hidup hingga musim panen masih tinggi, sama dengan lingkungan lainnya. Efek cekaman lingkungan diduga akan nyata seiring dengan pertambahan umur tanaman. Durasi pertumbuhan dapat menunjukkan tingkat keparahan cekaman atau penurunan hasil karena efek cekaman bersifat kumulatif.
Rendahnya hasil panen tanaman utama di Pulang Pisau berhubungan dengan tingkat cekaman lingkungan yang lebih berat, yaitu kadar besi yang lebih tinggi dibandingkan lokasi lainnya dan tanah lebih masam. Kadar besi yang tinggi tersebut disebabkan oleh oksidasi pirit yang sudah terjadi sebelumnya. Lahan pasang surut tipe luapan C rentan terhadap oksidasi pirit karena sering mengalami kekeringan.
Pirit (FeS2) pada tanah sulfat masam akan stabil dan tidak berbahaya pada kondisi tergenang (anaerob). Pirit menjadi berbahaya jika permukaan air tanah turun akibat pasang surutnya air laut, musim kemarau atau pembuatan saluran drainase yang menyebabkan oksigen masuk ke pori tanah dan mengoksidasi pirit (van Breemen 1976). Pada saat pasang naik atau saat musim hujan yang menyebabkan sawah tergenang, tanah berada dalam kondisi reduksi sedangkan pada musim kemarau tanah berada dalam kondisi oksidasi. Menurut Fairhurst et al. (2007), oksidasi Fe2+ menjadi Fe3+ akibat pelepasan oksigen oleh akar menyebabkan pemasaman dalam rizosfer padi dan membentuk lapisan kecoklatan pada perakaran. Konsentrasi tinggi Fe2+ dalam tanah dapat mengganggu penyerapan unsur P dan K. Di bawah kondisi reduksi yang kuat, produksi H2S dan FeS dapat berperan dalam keracunan Fe dengan melemahkan daya oksidasi akar. Faktor penyebab keracunan besi adalah: 1) tingginya konsentrasi Fe2+ dalam larutan tanah karena kondisi reduksi dan/atau pH rendah, 2) status hara tanaman rendah, 3) daya pengeluaran Fe2+ yang buruk karena kahat P, Ca, Mg, atau K, dan adanya akumulasi asam organik, H2S, dan FeS di rizosfer yang bersifat menghambat respirasi.
Peningkatan konsentrasi ion H+ dalam larutan tanah menyebabkan laju absorpsi kation turun akibat dari kompetisi antar ion yang bermuatan sama, sehingga mempengaruhi keseimbangan ionik. Hal tersebut merupakan pengaruh langsung pH terhadap pertumbuhan tanaman. Kondisi tanah yang sangat masam akan menginduksi toksisitas Al, Fe dan Mn, serta ketidakcukupan Ca, Mg, P dan Mo (Alam et al. 1999). Unsur makro seperti N, P, K, Ca dan Mg menjadi tidak tersedia bagi tanaman pada pH < 4. Di sisi lain kadar Al3+ yang bersifat toksik meningkat sehingga menjadi faktor pembatas (Kochian 1995 dan Matsumoto 2000). Ion logam merupakan faktor stress yang dapat mengubah reaksi fisiologis (strain), sehingga vigor tanaman menurun atau dapat secara total menghambat pertumbuhan jika konsentrasinya sangat tinggi (Levit 1980).
Wheeler 1993). Bronzing disebabkan oleh aktivitas polifenol oksidase yang meningkat akibat keracunan Fe (Peng dan Yamauchi 1993).
Tanaman mempunyai batas toleransi terhadap besi. Sebagai gambaran, varietas Margasari yang menurut Suprihatno et al. (2009) toleran terhadap keracunan besi, ternyata menurut Koesrini dan William dalam Amnal (2009), varietas tersebut sudah menunjukkan gejala keracunan besi pada konsentrasi Fe dalam larutan tanah 400 ppm, dan tanaman mati pada konsentrasi Fe sekitar 600 ppm. Keracunan besi ini terlihat dari gejala bronzing pada daun, tanaman menjadi kerdil, pertumbuhan akar terhambat, biomasa tanaman rendah, umur panen terlambat dan hasil menurun.
Panen tanaman utama ketika tanaman belum kering atau pada saat tanaman masih aktif melaksanakan metabolisme akan menyebabkan kuncup tunas segera tumbuh menjadi ratun. Ratun akan tumbuh selama tidak ada cekaman yang menyebabkan tanaman utama mati. Pengaruh lingkungan maupun interaksi genotipe x lingkungan yang tidak nyata terhadap kemampuan meratun menunjukkan bahwa kemampuan meratun dikendalikan faktor genetik. Pada musim kemarau pun ratun masih dapat tumbuh jika tanaman utama diairi.
Potensi ratun padi dapat diketahui dari kemampuan tunas ratun untuk hidup dan tumbuh beberapa hari setelah panen. Seleksi dapat dilakukan di tiga tipologi lahan karena lingkungan tanah tidak berpengaruh nyata terhadap jumlah rumpun hidup.
SIMPULAN
3
KERAGAMAN GENETIK POTENSI RATUN PADA 68
GENOTIPE PADI
ABSTRAK
Penelitian bertujuan untuk: a) menjelaskan keragaman genetik potensi ratun pada 68 genotip padi, b) memperoleh kriteria seleksi untuk padi ratun dari karakter yang berkorelasi dengan potensi ratun, dan c) memperoleh genotipe yang mempunyai kemampuan meratun baik. Penyaringan genotipe-genotipe ratun dilaksanakan di Bogor pada bulan Oktober 2011 hingga Mei 2012. Sebanyak 68 genotipe ditanam menggunakan rancangan acak kelompok lengkap yang diulang tiga kali. Setiap genotipe ditanam sebanyak empat baris sepanjang 5 m dengan jarak tanam 20 cm x 20 cm saat bibit berumur 20 hari. Setiap lubang ditanam 1 bibit. Pupuk dasar Urea 100 kg ha-1, SP36 150 kg ha-1, KCl 50 kg ha-1 diberikan saat tanam. Pupuk susulan Urea 50 kg ha-1 dan KCl 50 kg ha-1 diberikan pada umur 35 hst. Panen dilakukan setelah 95% malai menguning dengan memotong batang padi pada ketinggian 10 cm. Satu hari setelah panen, lahan diairi dan ditaburkan pupuk Urea 50 kg ha-1, SP36 30 kg ha-1, dan KCl 25 kg ha-1. Hasil penelitian menunjukkan bahwa keragaman genetik dari kemampuan meratun genotipe padi luas. Seleksi untuk kemampuan meratun dapat dilakukan melalui karakter tanaman utama sebagai karakter sekunder, yaitu: diameter batang, tumbuh cepat dan vigor, daun hijau saat panen (stay green), anakan produktif banyak, dan produktivitas tanaman utama tinggi. Seleksi dengan menggunakan indeks seleksi menghasilkan sembilan genotipe dengan kemampuan meratun baik, yaitu: IPB97-F-13-1-1, IPB107-F-14-4-1, IPB107-F-14-5-1, IPB Batola 6R, Inpago IPB 8G, IPB 4S, IPB Batola 5R, IPB 3S, dan IPB107-F-18.
Kata Kunci: keragaman, padi pasang surut, kriteria seleksi, fenotipe, batang
GENETIC VARIABILITY OF RATOONING ABILITY OF 68 RICE GENOTYPES
ABSTRACT
harvested after 95% of panicles ripening with cutting height 10 cm. One day after harvest, the land was irrigated and sown with Urea 50 kg ha-1, TSP 30 kg ha-1, and KCl 25 kg ha-1. The results showed that the variability of ratooning ability of rice genotypes was high. Selection for ratooning ability can be done by using a secondary character, namely: culm diameter, growing fast and vigor, green leaves at harvest (stay green), many productive tillers, and high productivity of major crops. By using some of the characters as the selection criteria, nine potential genotypes for ratoon were selected, namely: IPB97-F-13-1-1, IPB107-F-14-4-1, IPB107-F-14-5-1, IPB Batola 6R, Inpago IPB 8G, IPB 4S, IPB 5R Batola, IPB 3S and IPB107-F-18.
Keywords: diversity, tidal rice, selection criteria, phenotype, rice culm
PENDAHULUAN
Persilangan untuk memperbaiki karakter padi lokal pasang surut dan menambahkan karakter ratun telah dilaksanakan dan menghasilkan galur-galur yang kemampuannya menghasilkan ratun masih perlu diuji. Susilawati et al (2010) menemukan sembilan genotipe memiliki kemampuan menghasilkan ratun tinggi dari 18 genotipe yang diuji dan pada pengujian lima genotipe di lahan pasang surut, Susilawati et al. (2012) memperoleh hasil ratun berkisar 38.1-56.6% dari hasil tanaman utama. Genotipe yang diuji sedikit sehingga informasi keragaman ratun padi pasang surut belum diketahui.
Beberapa karakter seperti tumbuh cepat dan vigor, batang tebal, genjah, dan sifat daun masih berwarna hijau pada saat panen “stay green” diduga berhubungan dengan karakter ratun dan dikendalikan secara genetik, tetapi belum diketahui karakter yang paling dominan mempengaruhi hasil ratun. Karakter ketebalan batang berkorelasi dengan tingginya kadar karbohidrat dalam jerami. Karbohidrat dapat menginduksi kemampuan regenerasi ratun yang lebih kuat dan hasil ratun yang lebih banyak (Palchamy dan Purushothaman, 1988). Pertumbuhan ratun padi tergantung dari jumlah total kadar karbohidrat pada pangkal batang, pada awal pertumbuhan. Jumlah karbohidrat yang besar dibutuhkan untuk menghasilkan banyak anakan (Ichii dan Ogaya 1985). Terdapat keragaman kemampuan meratun antar kultivar akibat kendali genetik (Bahar dan de Datta 1977) dan pengaruh lingkungan.
Padi tipe baru memiliki pertumbuhan vegetatif yang lebih baik dan memberikan hasil ratun yang lebih tinggi dibandingkan varietas lain, tetapi varietas hibrida lebih vigor dibandingkan inbrida (Susilawati et al. 2010). Di antara 30 varietas/galur yang diuji untuk mengetahui kemampuan meratun, hanya 3 galur yang berpotensi ratun dan vigor (Oad dan Cruz 2002). Durasi pertumbuhan adalah salah satu karakter genetik yang berpengaruh nyata terhadap kemampuan meratun dan hasil padi (Vergara et al. 1988).
jika batang tanaman utama dipotong dengan meninggalkan 2-3 buku. Keragaman tanah, air, cahaya, dan suhu sangat berpengaruh terhadap kemampuan meratun (Krishnamurthy 1998).
Aspek fisiologis seperti laju fotosintesis netto pada fase pengisian biji yang berbeda antar kultivar, menyebabkan hasil ratun berbeda (Zhang et al. 2011). Oleh karena itu seleksi genotipe-genotipe ratun akan dapat menghasilkan ideotipe ratun yang baik. Di sisi lain, kemampuan meratun varietas sangat dipengaruhi oleh teknik budidaya. Menurut Liu et al. (2012), sistem tanam pindah menghasilkan ratun yang lebih baik dibandingkan sistem tebar benih atau tanam benih langsung.
Informasi penyaringan genotipe ratun khususnya di lahan pasang surut masih sangat terbatas. Tujuan penelitian ini adalah untuk mengetahui keragaman potensi meratun pada berbagai genotipe padi, memperoleh kriteria seleksi untuk padi ratun dari karakter yang berkorelasi dengan potensi ratun, dan memperoleh genotipe yang