SENJAYA, KECAMATAN PANGKALAN LADA
KALIMANTAN TENGAH
BANDUNG SAHARI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Struktur Komunitas Parasitoid
Hymenoptera di Perkebunan Kelapa Sawit, Desa Pandu Senjaya, Kecamatan
Pangkalan Lada, Kalimantan Tengah adalah karya saya dengan arahan dari komisi
pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana
pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun
yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan
dalam Daftar Pustaka di bagian akhir disertasi ini.
Palm Plantations, Pandu Senjaya Village, Pangkalan Lada District, Central
Kalimantan.
Under
supervision
of
DAMAYANTI BUCHORI, SJAFRIDA
MANUWOTO, and ALI NURMANSYAH.
The focus of this research was to understanding the community pattern of
lepidopteran pests in oil palm plantation, based on effects of oil palm age gradient
and the present of the flowering plant
Turnera
subulata
J.E.Smith (Parietales:
Turneraceae) on hymenopteran parasitoid community. This research was conducted
in Central Kalimantan between September 2011 and June 2012. In total, as much as
17 species of lepidopteran pests were identified to infest oil palm by surveying 976
trees from three different ages of oil palm plantations. Limacodidae was found to be
the most specious and abundant family inhabiting oil palm plantations in the study
area.
Setora nitens
Walker (Lepidoptera: Limacodidae) was recorded to be the most
abundant species. Abundance structure of lepidopteran pests seems to be different for
different ages of oil palm.
S. nitens
was found to be dominant in less than three years
old oil palms and bagworm was dominant in six years old oil palms. Only five pests
species were parasitized by parasitoids including
Birthamula chara
Swinhoe
(Lepidoptera: Limacodidae), Darna diducta
Snellen (Lepidoptera: Limacodidae),
Darna trima
Moore (Lepidoptera: Limacodidae), Darna bradleyi
Holloway
(Lepidoptera: Limacodidae) and ulat bulu-3 (Lepidoptera: Limantriidae). Parasitizing
parasitoids
were
Braconidae-y
(Hymenoptera: Braconidae),
Euplectrus
sp
(Hymenoptera:
Eulophidae)
and
Tachinidae-1
(Diptera:Tachinidae).
For
hymenopteran parasitoid community, in total, there were 7,675 specimens belonging
to 204 morphospecies.
Brachymeria lasus
Walker (Hymenoptera: Chalcididae) and
Brachymeria
sp (Hymenoptera: Chalcididae) were recorded to be the most dominant
species inhabiting oil palm plantations. Flowers of
T. subulata
only support a certain
number of species of hymenopteran parasitoid. Species richness was not different
between hymenopteran parasitoids visiting flowers of
T. subulata
and inhabiting
inside oil palm plantations, however species composition show different pattern. Oil
palm age has an effect on species richness, abundance, and species composition of
hymenopteran parasitoid in an oil palm plantation. In general, younger oil palm
plantations seem to support higher number of species and individual of hymenopteran
parasitoids than older oil palm plantations.
BANDUNG SAHARI.
Struktur Komunitas Parasitoid Hymenoptera di Perkebunan
Kelapa Sawit, Desa Pandu Senjaya, Kecamatan Pangkalan Lada, Kalimantan Tengah.
Dibimbing oleh DAMAYANTI BUCHORI, SJAFRIDA MANUWOTO, dan ALI
NURMANSYAH.
Tujuan utama penelitian ini adalah (1) mempelajari pola komunitas hama
Lepidoptera yang menyerang tanaman kelapa sawit dan parasitoid yang berasosiasi,
(2) memahami pengaruh umur tanaman kelapa sawit terhadap komunitas parasitoid
Hymenoptera di lapangan, (3) mempelajari respon komunitas parasitoid Hymenoptera
terhadap penanaman tanaman berbunga
Turnera subulata
J.E.Smith (Parietales:
Turneraceae) sebagai sumber pakan bagi parasitoid, dan (4) memahami faktor-faktor
kunci yang mempengaruhi komunitas parasitoid Hymenoptera di perkebunan kelapa
sawit.
Untuk mempelajari struktur komunitas serangga hama Lepidoptera yang
menyerang tanaman kelapa sawit, tingkat parasitisisasi dan keanekaragaman musuh
alaminya, larva Lepidoptera dikoleksi dari setiap tanaman kelapa sawit mengikuti
jalur transek diagonal pada lima blok (dua blok adalah tanaman berumur kurang dari
tiga tahun dan tiga blok adalah tanaman berumur enam tahun) pertanaman kelapa
sawit. Secara keseluruhan, terdapat 17 spesies hama yang ditemukan menyerang
tanaman kelapa sawit. Perbedaan umur tanaman kelapa sawit mempengaruhi struktur
komunitas hama Lepidoptera yang berasosiasi. Ulat api (Limacodidae), terutama
Setora nitens
Walker (Lepidoptera: Limacodidae) merupakan hama yang perlu
diwaspadai untuk tanaman kelapa sawit berumur tiga tahun, sedangkan ulat kantung
(Lepidoptera: Psychidae) dan ulat bulu (Lepidoptera: Limantriidae) untuk tanaman
kelapa sawit yang berumur enam tahun. Hanya lima spesies hama yang ditemukan
terparasit, yaitu
Birthamula chara
Swinhoe (Lepidoptera: Limacodidae), Darna
diducta
Snellen (Lepidoptera: Limacodidae), Darna trima
Moore (Lepidoptera:
Limacodidae), Darna bradleyi
Holloway (Lepidoptera: Limacodidae) dan ulat
bulu-3 (Lepidoptera: Limantriidae). Parasitoid yang ditemukan menyerang kelima spesies
tersebut
adalah Braconidae-y
(Hymenoptera: Braconidae),
Euplectrus
sp
(Hymenoptera: Eulophidae) dan Tachinidae-1 (Diptera: Tachinidae). Kombinasi jenis
parasitoid meningkatkan
parasitisasi secara keseluruhan.
Braconidae-y
dan
Euplectrus
sp. secara bersama memberikan tingkat parasitisasi yang cukup baik pada
D. trima.
dibandingkan dengan jumlah individu pada habitat kebun kelapa sawit. Hal ini tidak
terlepas dari kontribusi
B. lasus
yang jumlahnya sangat melimpah pada bunga
T.
subulata.
Secara umum, penanaman
T. subulata
tidak menyokong seluruh spesies
parasitoid Hymenoptera yang berkunjung, hanya sebagian kecil spesies saja.
Komposisi spesies penyusun komunitas parasitoid terlihat lebih mirip baik menurut
umur tanaman kelapa sawit maupun tipe habitat. Perbedaan komposisi spesies akan
mempengaruhi fungsi-fungsi ekologi dari setiap spesies penyusun komunitas.
Komposisi parasitoid dapat digunakan sebagai indikator yang penting untuk melihat
status fungsi keanekaragaman hayati parasitoid secara keseluruhan.
Untuk
meningkatkan keefektifan pengendalian hayati dengan parasitoid, disarankan untuk
menanam lebih dari satu spesies tanaman berbunga untuk menjaga keanekaragaman
parasitoid.
KALIMANTAN TENGAH
BANDUNG SAHARI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Entomologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dr. Ir. Pudjianto, M.Si.
Nama
: Bandung Sahari
NRP
: A 361070031
Disetujui
Komisi Pembimbing
Dr.Ir. Damayanti Buchori, M.Sc.
Ketua
Prof. Dr. Ir. Sjafrida Manuwoto, M.Sc.
Dr.Ir. Ali Nurmansyah, M.Si.
Anggota
Anggota
Diketahui
Ketua Program Studi
Dekan Sekolah Pascasarjana
Entomologi
Dr. Ir. Pudjianto, M.Si.
Dr. Ir. Dahrul Syah, M.Sc.Agr.
Puji syukur kepada Allah SWT atas segala karunia-Nya, sehingga karya
ilmiah ini dapat diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan
sejak bulan September 2011 hingga Juni 2012 ini adalah pengendalian hayati di
perkebunan kelapa sawit, dengan judul Struktur Komunitas Parasitoid Hymenoptera
di Perkebunan Kelapa Sawit, Desa Pandu Senjaya, Kecamatan Pangkalan Lada,
Kalimantan Tengah. Terimakasih Penulis ucapkan kepada Dr. Damayanti Buchori,
sebagai ketua Komisi Pembimbing, Prof. Dr. Sjafrida Manuwoto dan Dr. Ali
Nurmansyah, sebagai anggota Komisi Pembimbing.
Terimakasih juga disampaikan kepada Ketua Departemen Proteksi Tanaman,
Dr. A. Asih Nawangsih, dan Ketua Program Studi Program Studi Entomologi,
Pascasarjana Institut Pertanian Bogor, Dr. Pudjianto yang telah memberikan
kesempatan kepada saya untuk menyelesaikan studi di IPB. Saya juga berterimakasih
kepada seluruh pengajar IPB yang telah membantu dan memberikan masukan yang
bernilai. Ucapan terimakasih sebesar-besarnya juga disampaikan kepada Bapak
Widya Wiryawan, Bapak Bambang Palgoenadi, Bapak Joko Supriyono, selaku
President Direktur dan Direksi, Bapak Satyoso Harjotedjo, Administratur PT Agro
Menara Rahmat dan seluruh jajaran staf untuk semua dukungan dan fasilitas yang
diberikan selama saya menjalankan penelitian dan menulis disertasi. Ucapan
terimakasih secara khusus disampaikan kepada Prof (Riset) Dr. Rosichon Ubaidillah
untuk dukungan dan masukan yang berarti serta kepada tim R&D PT Astra Agro
Lestari yang telah membantu dalam pelaksanaan penelitian. Secara khusus saya
berterimakasih kepada istri tercinta Lu’lu’ Agustina, anak-anakku tercinta Ghifa and
Hanif, Ibu dan seluruh keluarga atas dukungan yang tidak terhingga. Saya berharap
bahwa hasil penelitian ini bisa bermanfaat untuk perkembangan ilmu pengetahuan
dan dapat diaplikasikan di lapangan.
sebagai anak ke tujuh dari delapan bersaudara, putra dari Bapak Slamet Sutarso
(Alm) dan Ibu Sri Sumijarti. Pendidikan sarjana ditempuh di Program Studi Hama
dan Penyakit Tumbuhan, lulus pada tahun 1999. Pada tahun 2001, melanjutkan
pendidikan Magister Sains, pada program studi Entomologi. Pada tahun 2004, gelar
Magister Sains diperolehnya untuk kemudian tercatat kembali menjadi Mahasiswa
program Doktor mulai tahun 2007.
AB
A
BA
A
A
B
! "#$# % & ' ( ) ) #! ! # * + ,
A
- . '/# (0 -1 # //' ( "2 . *-1 "3 ('/ 0'* (4 !A
!( !/ 0/ 4#. *-1 "% !' 4#.*-1 "+ ( ! ! '& 56 78-trophic Interaction
9: .' * ( ( ( *:/!/ 0 2; !' 4 ## *< .' # ( ('/ / 4# .* -1=B
! >4# !?... ...
22
KOMUNITAS HAMA LEPIDOPTERA DAN
PARASITOIDNYA DI PERKEBUNAN KELAPA SAWIT
Abstrak...
27
Abstract...
27
Pendahuluan...
27
Metode Penelitian...
30
Hasil Penelitian...
32
xiii
PENGARUH UMUR TANAMAN KELAPA SAWIT
TERHADAP KOMUNITAS PARASITOID
HYMENOPTERA
Abstrak...
44
Abstract...
44
Pendahuluan...
45
Metode Penelitian...
47
Hasil Penelitian...
49
Pembahasan...
57
Simpulan...
Daftar Pustaka...
61
61
KOMUNITAS PARASITOID HYMENOPTERA
PENGUNJUNG BUNGA
Turnera subulata
PADA
PERKEBUNAN KELAPA SAWIT, KALIMANTAN
TENGAH
Abstrak...
64
Abstract...
64
Pendahuluan...
65
Metode Penelitian...
67
Hasil Penelitian...
69
Pembahasan...
80
Simpulan...
Daftar Pustaka...
83
83
KOMUNITAS PARASITOID HYMENOPTERA DI
PERKEBUNAN KELAPA SAWIT: GRADIEN UMUR,
TANAMAN BERBUNGA
T. subulata,
METODE
KOLEKSI SERANGGA
Abstrak...
87
Abstract...
87
Pendahuluan...
88
xiv
Simpulan...
Daftar Pustaka...
xv
Tabel 2.1
Distribusi luas kebun sawit menurut pemiliknya
di Indonesia 2004 – 2010...
15
Tabel 2.2
Spesies tumbuhan yang memiliki potensi untuk
menarik parasitoid...
25
Tabel 3.1
Daftar spesies Lepidoptera yang menyerang
tanaman kelapa sawit dan jumlah pokok yang
terserang dari total 976 contoh tanaman kelapa
sawit...
33
Tabel 3.2
Parasitoid yang ditemukan menyerang larva
hama ulat api...
37
Tabel 3.3
Larva Lepidoptera yang dikoleksi, mortalitas,
dan larva terparasit...
38
Tabel 3.4
Waktu pengamatan dan curah hujan...
39
Tabel 4.1
Rata-rata nilai suhu, kelembahan, dan cahaya di
lokasi penelitian...
59
Tabel 5.1
Daftar spesies yang diwakili oleh lebih dari 30
xvi
Halaman
Gambar 1.1
Posisi lokasi kebun penelitian ... ...
9
Gambar 1.2
Peta jalan penelitian... ... ...
11
Gambar 2.1
Proporsi impor 17 minyak nabati dunia pada tahun
2010...
12
Gambar 2.2
Produksi dan impor 17 minyak dan lemak nabati dunia
pada tahun 2010. ... ... ...
13
Gambar 2.3
Negara pengimpor minyak sawit pada tahun 2010...
13
Gambar 2.4
Kontribusi negara produsen dalam minyak sawit
pasokan dunia 2010... ... ...
14
Gambar 2.5
Produksi minyak sawit Indonesia dari tahun 2004
hingga 2010...
14
Gambar 2.6
Luas lahan yang digunakan untuk perkebunan kelapa
sawit dari tahun 2004 hingga 2010...
15
Gambar 3.1
Pola pengambilan contoh serangga hama pada kelapa
sawit...
31
Gambar 3.2
Proporsi kelimpahan famili hama Lepidoptera yang
menyerang tanaman kelapa sawit... ...
34
Gambar 3.3
Distribusi 17 jenis larva Lepidoptera yang menyerang
tanaman kelapa sawit contoh... ...
34
Gambar 3.4
Kelimpahan individu dari 17 spesies Lepidoptera yang
menyerang tanaman kelapa sawit ...
35
Gambar 3.5
Struktur kelimpahan Lepidoptera yang menyerang
kelapa sawit pada umur kurang dari tiga tahun dan
enam tahun...
35
Gambar 3.6
Jumlah larva yang dikoleksi dan larva terparasitid...
37
Gambar 4.1
Kurva akumulasi spesies untuk komunitas parasitoid
yang dikoleksi dari seluruh blok contoh kebun
sawit...
49
Gambar 4.2
Jumlah total individu dan spesies untuk setiap famili
xvii
Gambar 4.4
Rata-rata jumlah spesies parasitoid Hymenoptera yang
dikoleksi dari kelompok umur tanaman kelapa sawit
yang berbeda ...
52
Gambar 4.5
Rata-rata
jumlah
individu
spesies
parasitoid
Hymenoptera yang dikoleksi dari kelompok umur
tanaman kelapa sawit yang berbeda ...
52
Gambar 4.6
Plot skala dua dimensi berdasarkan nilai indeks
Sørensen untuk mengukur kemiripan komposisi
spesies antar plot pengamatan...
53
Gambar 4.7
Fluktuasi
kelimpahan
morfospesies
parasitoid
Hymenoptera yang dikoleksi dari kebun sawit berumur
kurang dari tiga tahun menggunakan nampan kuning
dan jaring serangga. ...
55
Gambar 4.8
Fluktuasi
kelimpahan morfospesies Hymenoptera
parasitoid yang dikoleksi dari kebun sawit berumur
kurang dari enam tahun menggunakan nampan kuning
dan jaring serangga. ...
56
Gambar 4.9
Plot skala dua dimensi berdasarkan nilai indeks
Sørensen untuk mengukur kemiripan komposisi
spesies antar plot pengamatan...
57
Gambar 5.1
Kurva akumulasi spesies untuk komunitas parasitoid
pengunjung bunga
T. subulata...
70
Gambar 5.2
Proporsi kekayaan spesies dari famili Hymenoptera
yang mengunjungi bunga
T. subulata
di perkebunan
sawit... ...
70
Gambar 5.3
Proporsi kelimpahan inividu parasitoid Hymenoptera
dari berbagai famili yang mengunjungi bunga
T.
subulata
di perkebunan kelapa sawit...
71
Gambar 5.4
Peringkat kelimpahan individu dari 16 spesies
parasitoid Hymenoptera yang mengunjungi bunga
T.
subulata... ...
71
Gambar 5.5
Rata-rata jumlah spesies parasitoid Hymenoptera yang
dikoleksi dari tiga kelompok umur kelapa sawit
xviii
dengan masing-masing dua blok contoh...
73
Gambar 5.7
Plot skala dua dimensi berbasis pada nilai indeks
Sørensen untuk melihat kemiripan komposisi spesies
diantara plot-plot pengamatan pada tiga kelompok
umur tanaman yang berbeda...
74
Gambar 5.8
Frekuensi kunjungan parasitoid Hymenoptera pada
bunga
T. subulata
dari kebun kelapa sawit berumur
kurang dari tiga tahun. ... ...
76
Gambar 5.9
Frekuensi kunjungan parasitoid Hymenoptera pada
bunga
T. subulata
dari kebun kelapa sawit berumur
enam tahun. ... ...
77
Gambar 5.10
Frekuensi kunjungan parasitoid Hymenoptera pada
bunga
T. subulata
dari kebun kelapa sawit berumur 18
tahun. ...
78
Gambar 5.11
Rata-rata jumlah spesies yang dikoleksi dengan
metode nampan kuning dan koleksi langsung dengan
jaring serangga...
79
Gambar 5.12
Rata-rata jumlah individu yang dikoleksi dengan
metode nampan kuning dan koleksi langsung dengan
jaring serangga...
79
Gambar 5.13
Plot skala dua dimensi proyeksi dari perhitungan
indeks Sørensen untuk kesamaan komposisi spesies
Hymenoptera parasitoid yang tertangkap dengan
nampan kuning... ...
80
Gambar 6.1
Kurva akumulasi komunitas parasitoid Hymenoptera
di kebun sawit... ...
92
Gambar 6.2
Peringkat kelimpahan relatif dari 37 spesies parasitoid
Hymenoptera di kebun kelapa sawit. ... ...
93
Gambar 6.3
Kelimpahan relatif parasitoid Hymenoptera yang
dikoleksi dari dalam kebun (atas) dan dari vegetasi
T.
subulata
(bawah) ...
94
Gambar 6.4
Rata-rata jumlah individu parasitoid Hymenoptera
xix
kelimpahan parasitoid Hymenoptera dari kelompok
umur tanaman yang berbeda...
97
Gambar 6.6
Plot skala dua dimensi berbasis indeks Sørensen untuk
mengukur kemiripan komposisi spesies antar plot-plot
pengamatan dari kelompok umur yang berbeda...
98
Gambar 6.7
Plot skala dua dimensi berbasis pada indeks Sørensen
untuk mengukur kemiripan komposisi spesies antar
xx
Lampiran 1
Pangsa pasar pestisida di Indonesia...
119
Lampiran 2
Gambar beberapa jenis ulat api yang dikoleksi dari
lokasi penelitian... ... ...
120
Lampiran 3
Gambar beberapa jenis ulat api terparasit dan
parasitoidnya...
121
Lampiran 4
Lampiran 5
Lampiran 6
Lampiran 7
Lampiran 8
Lampiran 9
Lampiran 10
Daftar morfospesies/spesies parasitoid Hymenoptera di
kebun sawit. ... ... ...
Hasil analisis ragam jumlah spesies dan jumlah
individu parasitoid yang dikoleksi dari dalam kebun
tanaman kelapa sawit dengan umur yang berbeda...
Hasil analisis ragam jumlah spesies dan jumlah
individu parasitoid Hymenoptera yang dikoleksi
dengan nampan kuning dan jaring serangga untuk
habitat di dalam blok...
Hasil analisis ragam data log jumlah spesies dan log
jumlah
individu
parasitoid
Hymenoptera
yang
mengunjungi bunga
T. subulata
pada umur tanaman
kelapa sawit yang berbeda...
Hasil analisis ragam data log jumlah spesies dan log
jumlah
individu
parasitoid
Hymenoptera
yang
mengunjungi bunga
T. subulata
yang dikoleksi dengan
nampan kuning dan jaring serangga...
Hasil analisis ragam data log jumlah spesies dan log
jumlah
individu
parasitoid
Hymenoptera
yang
dikoleksi dari dua tipe habitat yang berbeda...
Hasil analisis ragam data log jumlah spesies dan log
jumlah
individu
parasitoid
Hymenoptera
yang
dikoleksi baik dari dalam kebun dan
yang
mengunjungi bunga
T. subulata
pada umur tanaman
kelapa sawit yang berbeda...
122
127
128
129
130
131
elakang
Minyak sawit merupakan minyak nabati yang sangat penting di dunia,
terutama jika dilihat dari permintaan pasar terhadap minyak sawit yang jauh lebih
tinggi dibanding minyak nabati lain. Indonesia merupakan produsen utama
dengan memasok sekitar 47% kebutuhan minyak sawit dunia. Pada tahun 2010,
produksi minyak sawit Indonesia mencapai 21,9 juta ton yang dihasilkan dari 8,1
juta hektar luas kebun sawit yang meliputi perkebunan pemerintah, sektor swasta,
dan masyarakat (KMSI 2010). Minyak sawit merupakan produk pertanian terbesar
ke dua di Indonesia, dan sektor pertanian berkontribusi 14,4% dalam produk
domestik bruto (World Growth 2011). Dengan kondisi ini, ekspor minyak sawit
diharapkan dapat memberikan dampak yang positif terhadap pertumbuhan
perekonomian nasional, sehingga kesejahteraan masyarakat baik yang terkait
langsung maupun tidak langsung dengan produksi minyak sawit akan meningkat.
Tanaman kelapa sawit,
fgaei
hg
ijkle
k hi
hJacq. (Arecales: Arecaceae), yang
berasal dari Afrika Barat telah banyak dibudidayakan di berbagai negara tropis di
Asia dan Amerika Selatan (Latiff 2000). Indonesia merupakan lokasi yang sangat
cocok untuk pengembangan perkebunan kelapa sawit. Namun demikian di saat
yang sama, Indonesia juga menjadi surga bagi serangga-serangga herbivora,
termasuk yang menjadi hama kelapa sawit. Tanaman kelapa sawit telah masuk
Asia sejak 100 tahun lalu, dan beberapa spesies herbivora lokal yang berasosiasi
dengan tumbuhan golongan palem telah beradaptasi dengan kelapa sawit, serta
kemudian berkembang menjadi hama penting kelapa sawit (Basri & Kamarudin
2000). Untuk perkebunan kelapa sawit monokultur skala luas, serangan hama bisa
menjadi masalah yang sangat serius, dan dilaporkan mampu mengakibatkan
kehilangan hasil hingga 90% (Sudharto
e
ma
g. 2003). Oleh karena itu, kemudian
serangga hama ini dikenal menjadi faktor pembatas produksi yang sangat
diperhitungkan baik untuk masa lalu, saat ini, dan masa depan.
Masalah hama ini pada umumnya diatasi dengan aplikasi pestisida untuk menjaga
populasi hama tetap rendah. Pada tahun 2010, pangsa pasar pestisida di Indonesia
berdasarkan jenis tanaman, paling besar adalah tanaman padi yang mencapai 42%
diikuti oleh tanaman perkebunan 40%, sedangkan berdasarkan jenisnya,
insektisida adalah yang terbesar, yaitu 41% (Direktorat Pupuk dan Pestisida 2012,
diunduh dari situs Perhimpunan Entomologi Indonesia). Aplikasi bahan kimia
pembunuh hama ini memberikan dampak negatif terhadap musuh alami dan
kesehatan agroekosistem (Garratt
e
na
o. 2011). Serangga penyerbuk kelapa sawit,
po
aed
qrstu va
w xy tzsc
tuFaust. (Coleoptera: Curculionidae), juga dikhawatirkan
dapat menjadi salah satu serangga berguna yang terkena dampaknya sehingga bisa
menurunkan produksi buah segar. Produksi buah sawit ini sangat bergantung pada
keberhasilkan penyerbukan yang dibantu oleh serangga tersebut.
Tantangan lain muncul dari meningkatnya kesadaran global tentang konsep
keberlanjutan (lihat Prinsip dan Kriteria RSPO-
Roundtable for Sustainable Palm
Oil
-; ISPO-
Indonesia Sustainable Palm Oil
-, lihat dokumen
Peraturan
Menteri
Pertanian Nomor 19/Permentan/OT.140/3/2011;
Green Economy Scheme
), yang
secara simultan mendorong perkebunan sawit untuk menerapkan praktik
pengendalian hama yang lebih ramah lingkungan. Disinilah kemudian, prinsip dan
konsep Pengendalian Hama Terpadu (PHT) berperan penting sebagai landasan
pengembangan teknik-teknik pengendalian hama ramah lingkungan. PHT
merupakan sistem penunjang pengambilan keputusan dalam memilih dan
menerapkan taktik pengendalian organisme pengganggu tanaman yang dipadukan
ke dalam strategi pengelolaan usaha tani, dengan berdasarkan pada analisis
biaya/manfaat, dan dengan mempertimbangkan kepentingan serta dampaknya
pada petani, khalayak, dan lingkungan (Kogan 1998). Teknik pengendalian hama
dengan meningkatkan peran musuh alami dipercaya sebagai tulang punggung
PHT. Namun demikian, dalam implementasinya, tidak seluruh teknologi PHT
dapat diterapkan dengan baik, dan hasilnya pun masih jauh dari yang diharapkan.
thuringiensis
menjadi pilihan utama. Namun demikian, teknik ini banyak
mengalami kendala, diantaranya adalah ketidaksesuaian tingkat keasaman
pencernaan serangga target dengan protein kristal dari bakteri sehingga protein
kristal tidak bisa tercerna dan gagal melepaskan racun yang dapat merusak sel-sel
epitel dalam saluran pencernaan. Selain itu masalah ketepatan waktu aplikasi
bakteri juga menentukan keefektifan pengendalian (Basri & Kamarudin 2000).
Untuk skala perkebunan yang luas, optimalisasi peran musuh alami yang
ada di lapangan merupakan strategi yang paling tepat. Selain berfungsi sebagai
pengendalian hama secara alami, pengendalian hayati juga berperan penting untuk
menjembatani terwujudnya pertanian berkelanjutan dan ekosistem yang lebih
sehat. Namun demikian, pengendalian hayati ini tidak bekerja secara optimal
karena peledakan hama masih sering dijumpai, sehingga muncul pertanyaan
mengapa peledakan hama masih saja terjadi?, bagaimana peran musuh alami?,
apakah ada yang salah dari praktik-praktik PHT yang diterapkan?
Dalam kaitan dengan strategi untuk mengatasi masalah hama di lapangan,
kita harus memahami apa yang disebut konsep interaksi tiga tingkat trofik (
tri-trophic interaction
) antara tanaman inang, hama dan musuh alaminya.
Pemahaman terhadap konsep ini merupakan dasar dari pengembangan strategi
pengendalian hama yang efektif. Dalam konsep interaksi tiga tingkat trofik,
tanaman sangat mempengaruhi interaksi antara hama dan musuh alaminya.
Fenologi tanaman, karakteristik, dan senyawa volatil memiliki peran penting
dalam
evolusi
dan
ekologi
perilaku
pada
interaksi
inang/mangsa-parasitoid/predator. Tanaman dapat mempengaruhi parasitisasi, baik melalui
senyawa volatil yang dihasilkannya, maupun dengan menyediakan pakan alami
seperti embun madu atau nektar untuk parasitoid (Godfray 1994). Perkembangan
terakhir dari hipotesis interaksi tiga tingkat trofik dibahas lebih lanjut oleh
Mooney
et al
. (2012).
konsekuensi terhadap proses-proses ekologi dan interaksi antar tingkat trofik yang
terlibat seperti parasitoid/predator, serangga herbivora, dan tanaman inang. Di
perkebunan kelapa sawit, peledakan hama diduga merupakan akibat dari
menurunnya keanekaragaman musuh alami karena miskinnya kualitas habitat di
agroekosistem (Kruess &Tscharntke 1994; With
et al
. 2002). Penurunan
keanekaragaman hayati musuh alami menyebabkan banyak serangga herbivora
terhindar dari parasitisasi (Kruess & Tscharntke 1994) dan predasi (With
et al
.
2002).
Dalam konteks pengendalian hayati di perkebunan kelapa sawit, penelitian
yang ada lebih banyak terkonsentrasi pada pendataan musuh alami (Sankaran &
Syed 1972; Mexzon & Chincilla 1991), status serangan hama (Mexzon &
Chincilla 1991; Fee & Cheong 1991), pemantauan hama, dan alternatif
pengendaliannya (Sudharto
et al
. 2003). Informasi yang terkait dengan pola
interaksi tiga tingkat trofik dan implikasinya terhadap serangan dan pengendalian
hama belum banyak dilaporkan. Jadi ide dasar dari penelitian ini adalah
memahami faktor-faktor yang berpengaruh terhadap pola interaksi tiga tingkat
trofik antara musuh alami, serangga hama, dan tanaman inang, serta
keterkaitannya dengan pengendalian hayati di lapangan.
Dalam penelitian ini, kegiatan difokuskan pada kelompok parasitoid
Hymenoptera di perkebunan kelapa sawit. Diantara berbagai kelompok musuh
alami, parasitoid merupakan agens pengendalian hayati paling efektif dalam
menekan populasi hama di lapangan (Hawkins
et al.
1997). Kelompok
Hymenoptera parasitika merupakan kelompok serangga penting
dalam
parasitoid Hymenoptera ditemukan menyerang hama ulat kantung (Psychidae)
(Syankaran & Syed 1972; Basri
et al
. 1995; Cheong
et al
. 2010) dan ulat api
(Limacodidae) (Desmier de Chenon
et al
. 1989). Walaupun demikian, bukan
berarti keberadaan parasitoid secara otomatis akan menurunkan populasi hama
seperti yang diharapkan. Hasil penelitian sebelumnya menunjukkan bahwa
meskipun keberadaan parasitoid dapat mempengaruhi populasi ulat kantung di
lapangan, tetapi hasilnya masih belum seperti yang diharapkan (Cheong
et al
.
2010).
Efektifitas pengendalian hayati secara alami di lapangan sangat dipengaruhi
oleh kompleksitas komunitas parasitoid yang ada. Kompleksitas ini menjadi kunci
yang menentukan tingkat parasitisasi di lapangan (Kruess & Tschartke 1994;
Menalled
et al
. 1999). Di perkebunan kelapa sawit, kompleksitas habitat sangat
dipengaruhi oleh praktik agronomi yang diterapkan. Pada umumnya, tanaman
tidak ditanam secara serempak, tetapi secara bertahap mengikuti proses
land
clearing
, sehingga menciptakan keanekaragaman umur (gradien) tanaman kelapa
sawit di lapangan. Gradien umur tanaman pada perkebunan kelapa sawit ini akan
membawa konsekuensi terhadap tingkat kompleksitas vegetasi yang hidup di
dalamnya. Artinya bahwa peningkatan umur kelapa sawit akan mengubah pola
vegetasi lain karena semakin tua umur kelapa sawit maka kanopi terbentuk,
naungan meningkat dan intensitas cahaya yang masuk semakin berkurang,
sehingga iklim mikro, jenis vegetasi, dan ketersediaan pakan untuk parasitoid juga
akan berubah (Perovic
et al.
2010). Hal ini juga didukung oleh hasil penelitian
lain yang melaporkan bahwa keberadaan tanaman herba berbunga di suatu habitat
dapat menyediakan pakan yang berlimpah untuk parasitoid (Lee & Heimpel 2005;
Rohrig
et al
. 2008; Perovic
et al
. 2010).
agroekosistem diharapkan dapat meningkatkan peluang parasitoid untuk bertahan
dan berkembang (Gillespie
et al
2011). Penelitian sebelumnya melaporkan bahwa
keberadaan tanaman berbunga memainkan peran penting dalam mempertahankan
komunitas parasitoid di lapangan (Basri
et al
. 1999, 2001; Basri & Kamarudin
2002; Desmier de Chenon
et al
. 2002) dengan menyediakan pakan berupa nektar
yang sangat dibutuhkan parasitoid sebagai sumber energi (Hogg
et al
. 2011),
diharapkan dapat menjaga populasi parasitoid untuk bertahan di lapangan.
Saat ini, penanaman tanaman berbunga untuk konservasi musuh alami dan
optimalisasi pengendalian hayati banyak mendapat perhatian dari berbagai
peneliti di dunia (Wratten
et al
. 2003; Lee & Heimpel 2005; Fiedler & Landis
2007). Untuk tanaman kelapa sawit, eksplorasi berbagai spesies herba yang dapat
menyediakan pakan untuk parasitoid telah banyak dilakukan, dan disebutkan
bahwa
Casia cobanensis
Linnaeus (Fabales: Leguminosae) dan
Euphorbia
heterophilla
Linnaeus (Malpighiales: Euphorbiaceae) merupakan tanaman terbaik
(Desmier de Chenon
et al
. 2002; Tuck
et al.
2003; Kamarudin & Basri 2010 ).
Namun demikian, di lapangan termasuk di Sumatera, Kalimantan, dan Sulawesi,
Turnera subulata
J.E.Smith. (Parietales: Turneraceae) merupakan jenis tanaman
berbunga yang banyak ditanam. Padahal dari berbagai jenis yang dieksplorasi,
T.
subulata
adalah jenis yang belum banyak diteliti terutama dari potensinya sebagai
tanaman yang dapat menarik parasitoid. Pertanyaannya kemudian adalah, apakah
T. subulata
dapat menarik semua jenis parasitoid, atau hanya jenis tertentu saja
yang akan diuntungkan?
Jika dilihat dari sisi lain, parasitoid Hymenoptera sangat selektif dalam
gradien umur kelapa sawit juga menjadi salah satu faktor yang berpengaruh?
Data yang menjelaskan tentang aspek-aspek di atas sangat penting untuk
membangun suatu pendekatan pengelolaan kebun sawit ramah lingkungan,
terutama dalam kaitan dengan strategi pengendalian hama yang efektif dan
mampu menjembatani terwujudnya pertanian berkelanjutan.
Penelitian ini dimaksudkan untuk menjawab berberapa pertanyaan seperti
diuraikan dibawah ini:
(1) Apakah kompleksitas parasitoid Hymenoptera memiliki dampak yang
signifikan terhadap komunitas hama Lepidoptera yang berasosiasi dengan
kelapa sawit?
(2) Apakah gradien umur kelapa sawit mempengaruhi komposisi spesies,
keanekaragaman, dan kelimpahan parasitoid Hymenoptera?
(3) Apakah
T. subulata
yang ditanam di kebun sawit hanya dapat menyokong
takson tertentu dari parasitoid Hymenoptera?
(4) Apakah faktor-faktor kunci yang mempengaruhi dinamika pola komunitas
parasitoid Hymenoptera?
{ |
juan Penelitian
Tujuan akhir penelitian ini adalah untuk memberikan informasi mendasar
dan baru mengenai pola interaksi tanaman, serangga hama dan komunitas
parasitoid di perkebunan kelapa sawit, sehingga dapat dibangun suatu strategi
pengendalian hayati hama yang lebih baik berdasarkan informasi yang dihasilkan.
Tujuan utama penelitian ini adalah:
(1). Mempelajari pola komunitas hama Lepidoptera yang menyerang tanaman
kelapa sawit dan parasitoid yang berasosiasi
(2). Memahami pengaruh umur tanaman kelapa sawit terhadap komunitas
parasitoid Hymenoptera di lapangan
(3). Mempelajari respon komunitas
parasitoid
Hymenoptera terhadap
penanaman tanaman berbunga
T. subulata
sebagai sumber pakan bagi
parasitoid
}~
faat Penelitian
Penelitian ini diharapkan dapat memberikan informasi baru mengenai
aspek-aspek yang harus diperhatikan dalam meningkatkan peran pengendalian
hayati menggunakan parasitoid pada perkebunan kelapa sawit skala besar. Salah
satu pertanyaan utama yang ingin dijawab dalam penelitian ini adalah apakah
semua parasitoid dapat disokong oleh penanaman tanaman
T. subulata
yang
sekarang ini banyak ditanam? Melalui penelitian diharapkan dapat dihasilkan
informasi tentang jenis-jenis parasitoid Hymenoptera yang dapat disokong oleh
tanaman berbunga
T. subulata.
Selain itu, faktor-faktor yang mempengaruhi pola
dinamika komunitas parasitoid Hymenoptera yang hidup di perkebunan kelapa
sawit juga dapat diidentifikasi sehingga dapat dikembangkan strategi yang lebih
tepat dalam meningkatkan peran parasitoid di lapangan.
Lokasi Penelitian
Gambar 1.1 Posisi lokasi kebun penelitian, Desa Pandu Senjaya, Kecamatan
Pangkalan Lada, Kalimantan Tengah
enelitian
dilakukan serangkaian penelitian, pertama adalah struktur komunitas hama
Lepidoptera yang menyerang perkebunan kelapa sawit dan parasitoidnya.
P
P
R
O
D
U
K
T
E
K
N
O
L
O
G
I
P
E
N
E
L
IT
IA
N
D
A
N
P
E
N
G
E
M
B
A
N
G
A
N
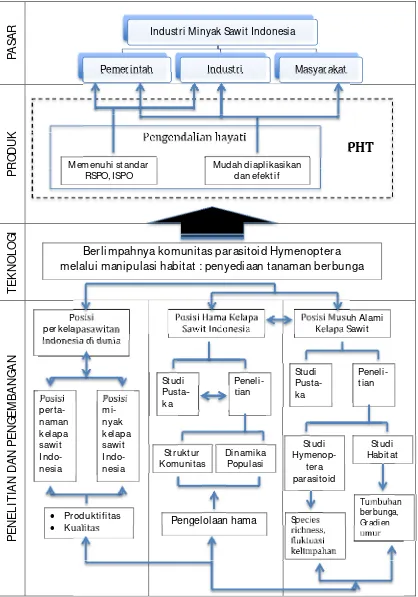
Gambar 1.2 Peta jalan penelitian
Pemer intah
Industr i
Masyar akat
Ber limpahnya komunitas par asitoid Hymenopter a
melalui manipulasi habitat : penyediaan tanaman ber bunga
Posisi
per kelapasawitan
Indonesia di dunia
Posisi
per
ta-naman
kelapa
sawit
Indo-nesia
Posisi
mi-nyak
kelapa
sawit
Indo-nesia
Pengendalian hayati
Mudah diaplikasikan
dan efektif
Memenuhi standar
RSPO, ISPO
PHT
Pr oduktifitas
Kualitas
Posisi Hama Kelapa
Sawit Indonesia
Studi
Pusta-ka
Peneli-tian
Str uktur
Komunitas
Dinamika
Populasi
Pengelolaan hama
Posisi Musuh Alami
Kelapa Saw it
[image:33.561.55.473.100.697.2]
r
u
k
s
n
w
t Du
n
n
In
o
n
s
inyak sawit merupakan minyak nabati terpenting di dunia
antara 1
¡minyak nabati yang dikonsumsi oleh masyarakat dunia
¢pangsa pasar minyak
sawit merupakan yang terbesar
¢yaitu 5
£¤¢diikuti oleh minyak kedelai
¢dan
minyak bunga matahari (
¥¦mbar 2
§¨¦n
¥¦mbar 2
©)
ªina
¢« ¬ ®opa
¢dan
¯ndia
merupakan tiga negara pengimpor terbesar dengan menyerap 52% produksi
minyak sawit dunia (
¥ambar 2
°)
±¦rena tanaman kelapa sawit hanya cocok
tumbuh di wilayah tropis
¢maka negara produsen terbesar minyak sawit dunia
pada umumnya berasal dari wilayah ini
¯ndonesia merupakan negara produsen
terbesar dengan memasok sekitar 4
¡¤kebutuhan minyak sawit dunia di tahun
2010
¢yang diikuti oleh
alaysia dengan 3
¡% (
¥ambar 2
²)
¶ ·
mbar 2
¸¹ ºroduksi dan impor 1
»minyak dan lemak nabati dunia pada tahun
2010 (
¼umber
½¾omisi
¿inyak
¼awit
Àndonesia 2010)
ÄÅ
mbar 2
ÆÇ È ÉÊËribusi negara produsen dalam minyak sawit pasokan dunia 2010
(
Ìumber
ÍÈomisi
Îinyak
Ìawit
Ïndonesia 2010)
Ð
roduksi minyak sawit di
Ïndonesia meningkat dari tahun ke tahun
Æ Ðada
tahun 2004
Ñproduksi minyak sawit hanya sekitar 12
ÆÒÓÔÆÇÕÖton
Ñdan mencapai
angka fantastik sekitar 21
ÆÖ×ØÆÕÓÙton pada tahun 2010 (
ÄÅmbar 2
Æ ×)
Æ ÚÅl ini
sejalan dengan meningkatnya luas perkebunan kelapa sawit dari 5
ÆÛÕÛÆ02
Ô ÜÅÝÅÞÅËÅÜß Ê ÓÙÙÇ à á ÊâÅÞ ã ØÆÕÕÙÆÇÇÛÜÅ ÝÅÞÅËÅÜßÊÓÙÕÙ
(
Ä ÅàäÅåÓÆÔ)
Æ æÅ äáç ÓÆÕ àá Êß ÊâßèèÅ ÊäÅÜéÅ çáäãÜÞÅ åã ×ÙêçÅÜÅ Ê èá çÅ ÝÅëÅéã Ë Þãèá çÉçÅÉçáÜ Ýá åßëÅÜÅÅ ÊÑ èá àßÞã ÅÊ Ýáåèáäß ÊÅÊ èá çÅ ÝÅ ëÅéãË åÅèìÅ Ë Þ ÅÊ Ëá åÅèÜã å ìÅ Êí Þãèáç É çÅ ÉçáÜ Ýá åßëÅÜÅ ÅÊàã çãèÊáíÅ åÅÆî ï
mbar 2
ðñ òuas lahan yang digunakan untuk perkebunan kelapa sawit dari
tahun 2004 hingga
2010 (
óumber
ô õomisi
öinyak
óawit
÷ndonesia 2010)
ø ù
eh karena itu
úperusahaan
ûperusahaan perkebunan kelapa sawit
memegang peran penting bukan hanya dalam konteks produksi nasional
útetapi
juga sebagai teladan dalam pencapaian produksi minyak sawit berkelanjutan
ðüý
dak mengherankan jika kemudian
úskema
ûskema pertanian berkelanjutan seperti
þóÿ ø ïn
÷óÿ øùebih ditekankan kepada perusahaan
ðü ï
bel 2
ð1
ýstribusi luas kebun sawit menurut pemiliknya di
÷ndonesia pada
tahun 2004
2010
ü ï
hun
ò
uasan area (ha)
ö
asyarakat
ÿemerintah
ÿerusahaan
2004
2
ð220
ð33
(3
ú%)
ñ ð(
ú)
ð ð(
úñ)
ðñð
(
ú)
ñð(
ú)
ððñ(
ú)
ñ ðñð
(
úñ)
ñ ð(
ú)
ðñð(
úñ)
ð ð
(
ú)
ñð(
ú)
ðñðññ(
ú)
ðð
(
ú)
ñ ñðñññ(
ú)
ð ð(
ú)
ðñ
(
ú)
ñð ñ(
ú)
ð ñðñ(
ú)
ððñ
(
ú)
ñð(
ú)
ð ð(
ú)
l
t
Kelapa sawit bukanlah tanaman asli Indonesia, oleh karena itu hama yang
berasosiasi merupakan serangga-serangga lokal yang telah beradaptasi sejak
introduksinya (Basri & Kamarudin 2000). Serangga-serangga yang saat ini
dipandang sebagai hama pada tanaman kelapa sawit diantaranya adalah ulat
kantung (Lapidoptera: Psychidae) (Sankaran & Syed 1972; Basri & Kamarudin
2000; Susanto
t
. 2010), ulat api (Lepidoptera: Limacodidae), ulat buah kelapa
sawit
r
!"r
#$v
%Walker (Lepidoptera:Pyralidae),
kumbang badak
&'
y
(t
s r
!%)(!r
)*Linnaeus (Coleoptera: Scarabaeidae) (Basri & Kamarudin
2000; Susanto
t
. 2010),
+,)-)%x
,.t
)%s
Ritsema (Coleoptera:
Scarabaeidae),
/!0%(!),)'u
s
sp.
(Coleoptera: Curculionidae),
+.)'tu
s
()1,'
ssu
s
Weber (Coleoptera: Scarabaeidae), belalang kembara
2)(u
st
1 -')'Linnaeus (Orthoptera: Acrididae), dan
3 %-%-'()'%s
Burmeister
(Orthoptera: Acrididae) (Susanto
t
. 2010).
Dari berbagai jenis hama tersebut, ulat api dan ulat kantung merupakan
hama terpenting kelapa sawit yang dapat menyebabkan kehilangan hasil yang
signifikan (Kalshoven 1981; Sankaran & Syed 1972; Wood 2002, Sudharto
t
.
2003; Kamarudin & Basri 2010; Cheong
t
. 2010). Dari tujuh spesies ulat
kantung yang berasosiasi dengan kelapa sawit (Basri & Kamarudin 2000), tiga
spesies diantaranya menjadi hama utama kelapa sawit termasuk
4t
s
p
%Walker
5 4!*% ( )'"tt
Tams
5
dan
6r
)1 ,% .#Joanis (Lepidoptera:
Psychidae) (Sankaran & Syed 1972; Wood 2002; Cheong
t
. 2010).
Hama-hama ini dilaporkan cukup penting di wilayah Sabah (Sankaran & Syed 1972) dan
mampu menyebabkan kehilangan hasil mencapai 43% setelah serangan yang
cukup serius (Kamarudin & Basri 2010). Jenis ulat kantung yang merugikan
secara ekonomi akan berbeda untuk lokasi yang berbeda (Basri & Kamarudin
2000). Ulat kantung dan dua spesies ulat api,
7t
)' %t
%*Walker dan
8 '%tr
1
Moore (Lepidoptera: Limacodidae) dapat meledak populasinya jika berada
dalam kondisi yang kondusif (Wood 2002). Di Indonesia, ledakan hama ulat api
dapat menyebabkan kehilangan hasil mencapai 70% pada tahun pertama serangan,
(Sudharto
9t
:;. 2003). Oleh karena itu penanganan terhadap hama-hama tersebut
secara tepat menjadi penting. Struktur komunitas hama dan kaitannya dengan
status musuh alaminya di lapangan merupakan informasi yang diperlukan untuk
mengembangkan strategi pengendaliannya.
<=>=? @AB@CD
ym
En
o
pt
Er
= EF=Gs
=@HG En
s
<En
G En
C=I@=JD=K=A@ C @<Erk
EFu
n
=JK
El
=L =M =N @t
Hymenoptera parasitika merupakan kelompok takson yang memiliki
kekayaan spesies cukup berlimpah (LaSalle 1993; Mason & Huber 1993; Quicke
1997; Whitfield 1998). Mayoritas anggota dari Hymenoptera parasitika berperan
sebagai parasitoid dan berfungsi dalam mengatur populasi serangga herbivora
(Quick 1997; Godfray 1994). Banyak spesies dari parasitoid Hymenoptera telah
digunakan untuk pengendalian hayati di lansekap pertanian (Noyes & Hayat 1984;
Quicke
1997).
Diantara
berbagai
kelompok
parasitoid
Hymenoptera,
Ichneumonidae dan Braconidae adalah famili terpenting yang ditemukan
menyerang berbagai jenis serangga dari ordo Lepidoptera dan Coleoptera
(Clausen 1940).
Peranan parasitoid Hymenoptera dalam pengendalian hayati di perkebunan
kelapa sawit telah banyak diteliti, diantaranya adalah mengenai potensinya dalam
pengendalian ulat api dan ulat kantung. (Sankaran & Syed 1972; Basri
9t
:;.
1995; Idris 2001; Desmier de Chenon
9t
:;. 1989). Desmier de Chenon
9t
:;O
(2002) melaporkan bahwa 36 jenis parasitoid dari famili Trichogrammatidae,
Eulophidae,
Encytidae,
Chalcididae,
Braconidae,
Ceraphronidae
dan
Ichneumonidae ditemukan berasosiasi dengan hama Lepidoptera yang menyerang
tanaman kelapa sawit termasuk
Limacodidae, Psychidae, Lymantriidae,
Noctuidae, Hesperidae, dan Amathusiidae. Beberapa parasitoid penting yang
diidentifikasi dari penelitian ini adalah
Pr
QRSTUV:WW:X TQY9:t
STZ9:9Nagaraja
(Hymenoptera: Trichogrammatidae) yang menyerang telur Limacodidae,
[ \Q]:VQ:sp. (Hymenoptera: Braconidae) yang secara spesifik menyerang inang
[O ]Qt
9]Z,
^ TV ]QRQ:
sp. (Hymenoptera: Braconidae) yang menyerang ulat api, dan
_\:]X9;9s
sp. (Hymenoptera: Braconidae) yang menyerang ulat kantung.
parasitoid yaitu
b cdefgeu
s
eh gi jcs
Walker
(Hymenoptera:
Eulophidae)
(parasitisasi 67,4%),
kflemnfdcoedcphc
t
pq pc
Nixon (Hymenoptera: Braconidae)
(parasitisasi 17,8%),
br clpqhe(Hymenoptera: Eulophidae) (parasitisasi 10,2%),
stnp ofuhjq
t
nvl pw(Hymenoptera: Ceraphronidae)
(parasitisasi 7,4%),
sjlfqp tncs p
sy
mnedev
fru
s
(Hymenoptera: Braconidae)(parasitisasi 9,6%),
xjtclh jq mpyfwp oy npcFerriere (Hymenoptera: Eupelmidae)(parasitisasi 3,2%),
xjiyt
fhpsp. (Hymenoptera:Eurytomidae) (parasitisasi 2,1%),
zchcljmnpsp.
(Hymenoptera: Ichneumonidae) (parasitisasi 1,1%),
{fiy
tnjqsp. (Hymenoptera:
Ichneumonidae) (parasitisasi 1,1%) (Hymenoptera: Ichneumonidae).
|r tl popdiparasit oleh
br e hgiu
cs
dan
{fiy
tnjqsp. (25%), sedangkan
|r mfi gctt
ehanya
diserang oleh
sr bqy
mnedev
fiu
s
. Selain dikenal sebagai parasitoid,
br p ofhplu
s
Gahan dan
br e hgi cu
s
juga diidentifikasi sebagai hiperparasitoid dari
kr hct
pqpc(Basri
ct
plr1995; Cheong
ct
pl. 2010).
Pada penelitian-penelitian sebelumnya, kegiatan lebih banyak dipusatkan
pada inventarisasi parasitoid (Sankaran & Syed 1972), potensinya di lapangan
(Cheong
ct
pl. 2010), dan peran tanaman berbunga yang sengaja ditanam agar
dapat meningkatkan populasi parasitoid di lapangan (Sankaran & Syed 1972;
Basri
ct
pl. 1995; 2002; Desmier de Chenon
ct
pl. 2002; Tuck
ct
pl. 2003;
Kamarudin & Basri 2010). Namun demikian, penelitian untuk melihat struktur
komunitas parasitoid, kondisi lansekap, dan implikasinya terhadap pengendalian
hayati masih sangat terbatas. Penelitian yang dilakukan oleh Idris (2001),
menunjukkan bahwa parasitoid dari Braconidae terlihat lebih melimpah dan
beraneka ragam dibandingkan Ichneumonidae di perkebunan kelapa sawit, hal ini
mungkin terkait dengan preferensi habitat yang berbeda dari dua kelompok
parasitoid ini. Di perkebunan kelapa sawit, belum ditemukan publikasi ilmiah
yang melaporkan pelepasan parasitoid untuk pengendalian hayati di lapangan.
Pengelolaan habitat melalui penanaman tanaman berbunga menjadi pilihan utama
}~~
n
lo
l
}~rk
~u
n
n
K
~l
t
Kelapa sawit merupakan salah satu tanaman pertanian penting di Indonesia
yang telah ditanam secara luas sekitar 8,1 juta ha hingga tahun tahun 2010 (KMSI
2010). Untuk perkebunan kelapa sawit skala besar. Praktik pengelolaan secara
intensif dengan sistem yang tepat selalu dikembangkan untuk menjamin
produktivitas yang semakin meningkat. Tanaman kelapa sawit pada umumnya
ditanam dalam sistem blok seluas 25 - 40 hektar untuk memudahkan pengelolaan,
baik dari sisi agronomi maupun pengaturan sumberdaya manusia (Dolmat MT
2000). Siklus hidup tanaman kelapa sawit berkisar antara 25 dan 35 tahun.
(peremajaan) dilakukan tergantung pada fase produktif tanaman
kelapa sawit. Penanaman untuk kebun yang luas tidak dilakukan secara serempak
tetapi secara bertahap sehingga menciptakan variasi umur kelapa sawit dari yang
muda hingga tua (gradien). Kompleksitas habitat dalam kebun diduga dipengaruhi
oleh proses ini.
Tanaman legum yang berfungsi sebagai tanaman penutup tanah pada
umumnya ditanam segera setelah
dilakukan. Tanaman legum ini
diharapkan dapat berfungsi untuk menjaga kelembahan tanah, melindungi tanah
dari curahan air hujan secara langsung, membantu mempertahankan bahan
organik
t
s
, proses ekologi dalam tanah, dan infiltrasi air (Chew
t
. 2009).
Pada tanaman muda, tanaman penutup tanah,
u
u
t
DC.ex Kurz
(Fabales: Fabaceae) mendominasi vegetasi habitat lahan kebun sawit. Komposisi
dan kelimpahan vegetasi ini akan berubah mengikuti pertumbuhan kelapa sawit.
Dengan meningkatnya umur kelapa sawit, penutupan kanopi akan
meningkat diikuti dengan menurunnya intensitas cahaya, meningkatnya
kelembahan, dan perubahan arsitektur vegetasi. Penyiangan gulma dilakukan
secara terjadwal, sehingga arsitektur vegetasi akan selalu dinamis. Basset (1991)
menyatakan bahwa perbedaan struktur vegetasi akan mempengaruhi iklim mikro
yang pada akhirnya berdampak terhadap kelimpahan dan keanekaragaman
artropoda. Vegetasi yang lebih kaya akan menyediakan sumberdaya yang lebih
baik untuk menyokong keanekaragaman serangga yang lebih tinggi (Hunter
2002), serta dapat memberikan dampak signifikan terhadap pola komposisi
Di lokasi penelitian, pengendalian hama didekati dengan konsep PHT yang
berbasis pada informasi hasil pemantauan populasi hama di lapangan. Pemantauan
hama dilakukan secara rutin sebulan sekali untuk menentukan tingkat serangan.
Jika populasi meningkat di atas ambang pada blok-blok tertentu, maka interval
pemantauan populasi hama pada blok-blok tersebut akan ditingkatkan dan
dilakukan pengendalian. Untuk hama Lepidoptera, pengendalian dilakukan
menggunakan beberapa cara, yaitu (1) pengasapan dengan bioinsektisida berbahan
aktif
t
s
, (2) kutip kepompong dan kutip ulat secara intensif, (3)
perangkap lampu, (4) aplikasi insektisida kimia sintetik dilakukan jika memang
sangat diperlukan (PT Astra Agro Lestari Tbk 2011)
¡ ¢ £¤
r
¥¦s
§¨ §©¥¨ §n
©k
¥£¨r
ª« §k
(
Tri-trophic Interaction
), Ko
¤mpl
k
s
§¥¬t
¥¢s
¤k
¥®,
¯ ¥n
°¤n
©¤n
¯ ¥± § ¥¢² ¥³ ¥£§.
Dalam menangani persoalan hama di lapangan, pola interaksi yang terjadi
antara tanaman sebagai inang serangga hama dan serangga hama sebagai inang
parasitoid perlu dipahami, serta perlu diketahui pengaruh tanaman inang terhadap
parasitoid. Hubungan antar tingkat trofik inilah yang menjadi dasar
pengembangan strategi pengendalian hama di lapangan. Tidak diragukan bahwa
tanaman memiliki pengaruh yang sangat kuat terhadap evolusi dan ekologi
perilaku interaksi inang-parasitoid (Price
t
´µ. 1980; Godfray 1994). Peranan
parasitoid dan predator dalam interaksi trofik antara tanaman inang dan serangga
herbivora telah mendapat perhatian serius dari para ahli setelah tahun 1980-an
(Money
t
´µ. 2012). Pengaruh tanaman terhadap interaksi antara parasitoid dan
inangnya telah banyak dibahas oleh Price
t
´µ. (1980). Dalam kenyataanya,
semua komunitas yang hidup di wilayah daratan paling tidak tersusun oleh
tingkatan trofik yang berbeda: tanaman, herbivora dan musuh alami dari
herbivora. Price
t
´µ. (1980) berargumentasi bahwa teori interaksi
serangga-tanaman tidak dapat berlangsung tanpa mempertimbangkan keterlibatan tingkat
trofik ke-tiga. Tingkat trofik ke tiga, harus dipertimbangkan sebagai bagian dari
pemicu pertahanan tanaman melawan serangga herbivora.
Parasitoid menggunakan stimulus yang dikeluarkan oleh tanaman untuk
mempengaruhi terjadinya parasitisme. Fase pertumbuhan dan sistem pertahanan
tanaman sangat berpengaruh terhadap serangga-serangga yang memakannya, yang
pada akhirnya berdampak terhadap terjadinya parasitisasi oleh parasitoid terhadap
serangga herbivora. Variasi karakteristik tanaman sangat mempengaruhi dinamika
interaksi parasitoid dan inangnya (van Emden 1966; Lawton & McNeill 1979).
Tanaman juga bisa sangat berpengaruh pada terjadinya parasitisasi, baik melalui
senyawa volatil tanaman atau dengan menyediakan pakan untuk parasitoid
melalui embun madu atau nektar (Price
¶t
·¸¹1980).
Perkembangan terakhir tentang konsep interaksi tiga tingkat trofik dibahas
oleh Mooney
¶t
·¸. (2012). Para peneliti ini kemudian mengajukan hipotesis
ºr
» ¼tr
½¾¿ »À »Á¶
r
·Àt
»½Á. Hipotesis ini merupakan pengembangan dari pendekatan
multitrofik yang melihat berbagai aspek yang mempengaruhi pola hubungan
tanaman, serangga herbivora dan parasitoid. Hipotesis ini dibangun dengan
mengintegrasikan dan membandingkan hipotesis-hipotesis yang sangat dikenal
sebelumnya:
t
¿¶ ¾¿ys
»½¸½ à »À·¸¶Ä Ä»À» ¶ÁÀy
(herbivora spesialis lebih baik dibanding
generalis dalam menggunanan tanaman yang sama sebagai pakan, dan kualitas
tanaman inang berpengaruh besar pada generalis dibanding spesialis)
Ås
¸½ Æ ¼Ãǽ Æ
t
¿È¿»Ã¿É½Çt
·¸ »ty
(kualitas inang yang rendah meningkatkan pengaruh musuh
alami)
Ådan
¶Á¶Éy
¼Är
¶¶s
¾·À¶¿Ê¾½Â ¿¶s
¶s
(herbivora spesialis lebih baik dibanding
generalis dalam menggunakan inang yang sama untuk menghindari predator).
Hipotesis-hipotesis tersebut menekankan tentang interaksi trofik dari kombinasi
yang spesifik antara serangga herbivora-kualitas tanaman, musuh alami, dan
variasi pakan herbivora mempengaruhi kebugaran (
¾¶r
Ä½Ç É ·ÁÀ¶) serangga
herbivora (Mooney
¶t
·¸. 2012). Teori tentang interaksi tiga tingkat trofik yang
telah teruji menunjukkan bahwa kualitas tanaman inang, musuh alami, dan variasi
pakan akan sangat mempengaruhi kebugaran serangga herbivora. Secara spesifik,
hipotesis ini menduga bahwa herbivora spesialis (jika dibandingkan dengan
generalis) memiliki peluang lebih tinggi untuk terhindar dari musuh alami dan
mampu berkompetisi dengan baik karena laju pertumbuhannya lebih cepat.
Kualitas tanaman inang dan musuh alami sangat mempengaruhi pola makan
menjelaskan apa yang akan terjadi jika kedua kelompok herbivora ini berbagi
jenis pakan yang sama.
Transformasi ekosistem alami menjadi agroekosistem membawa banyak
konsekuensi terhadap proses-proses ekologi yang ada, khususnya interaksi inang
–parasitoid dan
dampaknya terhadap pengendalian hayati. Menurunnya
keanekaragaman parasitoid karena rendahnya kompleksitas habitat dalam
agroekosistem diduga menjadi kunci gagalnya pengendalian hayati (Kruess &
Tscharntke 1994). Hubungan antara kompleksitas lansekap dan komunitas
parasitoid telah didokumentasi oleh banyak laporan penelitian (Idris 2001; Marino
Ë
t
ÌÍ. 2006; Buchori
Ët
ÌÍ. 2008). Hal tersebut dijelaskan oleh hipotesis yang
menyatakan bahwa kompleksitas lansekap meningkatkan keanekaragaman
parasitoid dan laju parasitisme (lihat Menalled
Ët
ÌÍ. 1999), yang saat ini menjadi
sebuah fenomena yang sangat umum. Beberapa studi juga mendukung hipotesis
bahwa kompleksitas habitat memiliki dampak yang signifikan terhadap kekayaan,
keanekaragaman dan kelimpahan spesies (Idris 2001; Marino
Ët
ÌÍ. 2006; Buchori
Ë
t
ÌÍ. 2008).
Pola komunitas parasitoid dalam suatu lansekap dapat dipengaruhi oleh
salah satu dari empat faktor sebagai berikut: (1) karakteristik inang (Hawkins
1994); (2) interaksi ekologi; (3) karakteristik habitat seperti habitat alami versus
agroekosistem, dan implikasinya pada stabilitas habitat dan keanekaragaman
(Hawkins 1994); dan (4) pola evolusi, termasuk didalamnya filogeni inang,
tingkat suksesi dari habitat inang, dan strategi pertahanan anti herbivora (Godfray
1994).
ÎÏ
n
ÐÏlo
l
ÑÑÒ Ó ÑÔ Õt
ÑÖu
n
tu
k
×ÏÏmp
rt
ÑØÑÒk
ÑÒKo
mu
Õt
ÑÙn
ÎÑr
ÑÙ Õt
ÚÕ ÛÛÕ ÎÏrk
ÏÔu
n
ÑÒK
Ïl
ÑÜ ÑÝ ÑÞÕt:
ÎÏr
ÑÒß ÑÒ Ñm
ÑÒB
ÏÔr
u
n
ÐÑPerkebunan kelapa sawit dalam skala besar sangat rentan terhadap
peledakan hama karena dukungan musuh alami yang terbatas, terutama parasitoid.
Habitat yang kompleks dapat menyediakan iklim mikro yang cocok serta sumber
pakan untuk parasitoid, sehingga dapat mempertahankan keanekaragaman
parasitoid Hymenoptera (Fiedler & Landis 2007; Silvinski
Ët
ÌÍ. 2006; Marino
Ët
memberikan sumber pakan, merupakan kunci untuk meningkatkan keefektifan
pengendalian hayati. Hubungan antara komunitas parasitoid dan tanaman
berbunga telah banyak dilaporkan (lihat Baggen & Gurr 1998; Wratten
àt
áâã2003; Lee & Heimpel 2005). Keanekaragaman parasitoid lebih tinggi pada habitat
yang memiliki banyak tumbuhan berbunga dibandingkan yang kurang memiliki
tumbuhan berbunga (Jervis
àt
áâ. 1993).
Dalam kaitannya dengan perkebunan kelapa sawit, beberapa peneliti telah
melaporkan beberapa spesies tumbuhan berbunga yang berpotensi digunakan
untuk menarik parasitoid diperkebunan sehingga pengendalian hayati menjadi
lebih efektif (Basri
àt
áâ. 1999; Desmier de Chenon
àt
áâ. 2002; Tuck
àt
áâ. 2003;
Kamarudin & Basri 2010). Teh (1996), Basri
àt
áâã(1999) dan Ho & Teh (1999)
telah menguji beberapa jenis tanaman yang memiliki potensi untuk menarik
parasitoid. Tabel 2.2 menunjukkan terdapat 10 famili dan 16 spesies tanaman
yang dapat menarik parasitoid pada perkebunan kelapa sawit. Basri
àt
áâ. (1999)
melaporkan bahwa
ä áås
æá çèéáêàêå æs
(Fabales: Leguminosae) dapat menarik
parasitoid
ëèìy
íîïåéïêèîGauld (Hymenoptera: Ichneumonidae) dan
ðïñòáêæás
èó
y
òèìáTosquinet (Hymenoptera: Ichneumonidae). Hasil ini didukung oleh
penelitian lain yang melaporkan bahwa
ä ã ç èéáêàês
æs
dapat menarik banyak
parasitoid hama ulat kantung,
ôã íâ áêá(Basri & Kamaruddin 2002). Namun
demikian Ho & Teh (1999) justru melaporkan bahwa
õïíîèìéæ á îàt
àr
èíîñâ âáLinnaeus (Malpighiales: Euphorbiaceae) adalah tumbuhan yang paling baik dalam
menarik parasitoid. Tuck
àt
áâã(2003) mengkonfirmasi bahwa baik
õãîà
t
àr
èíîñââádan
ä ãçèéáêàêå æs
keduanya dapat meningkatkan aktivitas parasitoid
di lapangan.
Penelitian-penelitian
tersebut
terkonsentrasi
untuk
meningkatkan
pengendalian hama ulat kantung di Malaysia, sebaliknya di Indonesia, dimana ulat
api yang menjadi hama utama, penelitian yang mengarah pada eksplorasi tanaman
berbunga untuk menarik parasitoid belum banyak dilakukan (Desmier de Chenon
et
áâ. 2002; Sudharto
àt
áâ. 2003). Salah satu studi untuk melihat peranan berbagai
tanaman berbunga dalam menyokong parasitoid dari ulat api dilakukan oleh
Desmier de Chenon
àt
áâ. pada tahun 2002. Dalam penelitian tersebut,
öãs
demikian, fakta di lapangan menunjukkan bahwa
ø ùs
úû úü ýþ ýadalah tanaman
berbunga yang paling banyak ditanam, walaupun penelitian untuk melihat
potensinya masih sangat terbatas. Parasitoid Hymenoptera sangat selektif dalam
memilih tanaman yang akan dikunjungi, bahkan cenderung memiliki preferensi
tertentu terhadap jenis tanaman berbunga (Coley & Luna 2000). Hal ini juga
didukung oleh Desmier de Chenon
ÿt
ýü. (2002) yang melaporkan bahwa terdapat
hubungan yang spesifik antara spesies tanaman berbunga dan parasitoid
pengunjungnya. Suatu spesies tumbuhan berbunga mungkin hanya akan
dikunjungi oleh sekelompok parasitoid tertentu. Hal ini berarti, pola komunitas
parasitoid yang hidup dalam suatu agroekosistem mungkin akan sangat
dipengaruhi oleh keanekaragaman jenis tanaman berbunga yang ada di dalamnya.
Dengan demikian, pemilihan tanaman berbunga secara tepat sangat penting untuk
berperannya musuh alami secara efektif dalam menekan populasi hama tertentu di
( 3 2 $ '. /* #%4 # ,% #$ 2
P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . # 6" %#( ) '" * +$ ," -$ # .$%$ / # $ 0 1 2B
#* &.$ " # * # " , #% #3 2 $ '. /* #%4 # ,% #$ 2
P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
6+$ ,"* ' #%#< * % . #+$ ,"* '# % #< : ; ; = 6 ," > '" #3 ? * % . #+$ ,"* '# % #< : ; ; = (
C
# //" #* ! # ,$, /" /9 ) '" * +$ ," -$ # .$%$ / # $ 0 1 2B
#* &.$ " # * # " , #% #3 2 $ '. /* #%4 # ,% #$ 2
P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
C
%# '# " #@# ,@" ! # " *#7 ) '" * +$ ," -$ # .$%$ / # $ 0 1 2B
#* &.$ " # * # " , #% # 3 2 $ '. /* #%4 # ,% #$ 2P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
C
6/ # #. $ , /" /9 ) '" * +$ ," -$ # .$%$ / # $ 0 1 2B
#* &.$ " # * # " , #% # 3 2 $ '. /* #%4# ,% #$ 2P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
A B/& /% # /" #" ,% / #9 ) '" * +$ ," -$ # .$%$ / # $ 0 1 2
B
#* &.$ " # * # " , #% # 3 2 $ '. /* #%4 # ,% #$ 2P
$ -" ! " / ". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
A B+$ #% .* ,&@" -$ / (
) '" * +$ ," -$ # .$%$ / # $ 0 1 2
B
#* &.$ " # * # " , #% # 3 2 $ '. /* #%4 # ,% #$ 2P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
C
C
'$ .$ %" - / $ .#DC ) '" * +$ ," -$ # .$%$ / # $ 0 1 2B
#* &.$ " # * # " , #% # 3 2 $ '. /* #%4 # ,% #$ 2P
$ -" ! " /". ! $ / 5 2P
6# , . # ' /7 28 $% # /%" * / 2 &% . #9 $%# ' : ; ; ;
GHIJ KJ L M NOJN O PHQHOJ R SJ TUHV WT JHI HRJ PN VNKJRJ X Y Z J H[NHN \]^ _` ab cd `e_f g ` ch
iJV V HNY O
j `kb dl ` m] nb^ ]h f] a] ch ] oJ pS Vq
B
ehd l_f] ebh dh eb nh ah rS ONM s qtu v ]kfu cdh a `whn alh ] GN QQJ N QNqP
]^ b ` gb u cbfg eu] c x HKyN QqP
zh n `fh k u c{ HsHVq| ]a eh c ab d lu cOM qtu e_ a`fhOM} HOQJ]ah k ~
\ z]e ab dbkk h ah iHIHQy j `kb dl ` m] nb^ ]h f] a] ch ] oJ pS Vq
B
ehd l_f] ebh dh eb nh ah rS ONM s qtu v ]kfu cdh a `whn alh ] GN QQJ N QNqP
]^ b ` gb u cbfg eu] c x HKyN QqP
zh n `fh k u c{ HsHVq| ]a eh c ab d lu cOM qtu e_ a`fhOM} HOQJ]ah k ~
B
` e e] eb hkh ab `kb hY Z K }QH[ S VJT HNq s HK [JT J T HN SN s PS KUWS V H[N HN n ab m ` n ` nk]v a`v u c S S yQV
` e_v lu c gu n ` l {HY KT q
B
u c_fh nbh `w_f` eh S OY JVNR q j `kb dl ` m] nb^ ]h f] a] ch ] oJ pS VqB
ehd l_f] ebh dh eb nh ah rS ONM s qtu v ]kfu cdh a `whn alh ] GN QQJ N QNqP
]^ b ` gb u cbfg eu] c x HKyN QqP
zh n`f h ku c {Hs HVq |] aeh cabd lu c OM q tue_ a`f h OM q u k` ch v l] cv c_d lb^ b ` euc Y NONZ N[yY [y]ah k ~
iHIJ H[NHN Y QVN QH[NHN
\_v ab cg e]b v ]cP SJ R | u en] ehcu gu k h ah
}QH[ S VJT HNq s HK [JT J T HN
` e_v lu c gu n ` l {HY KT q
B
u c_fh nbh `w_f` eh S OY JVNR q j `kb dl ` m] nb^ ]h f] a] ch ] oJ pS VqB
ehd l_f] ebh dh eb nh ah rS ONM s qtu v ]kfu cdh a `whn alh ] GN QQJ N QNq <