EFEKTIVITAS SUPLEMENTASI TEPUNG BATANG PISANG
DAN PROBIOTIK TERHADAP KECERNAAN HASIL
SAMPING KELAPA SAWIT SERTA EMISI METANA
A N T O N I U S
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Efektivitas Suplementasi Tepung Batang Pisang dan Probiotik Terhadap Kecernaan Hasil Samping Kelapa Sawit serta Emisi Metana adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sebagian data penelitian ini telah dipublikasikan di Pakistan Journal of Nutrition Vol 13 Tahun 2014 dengan judul artikel “Digestibility and Methane Emission of Ration Based on Oil Palm By Products Supplemented with Probiotics and Banana Stem: An In vitro Study”. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2014
RINGKASAN
ANTONIUS. Efektivitas Suplementasi Tepung Batang Pisang dan Probiotik Terhadap Kecernaan Hasil Samping Kelapa Sawit serta Emisi Metana. Dibimbing oleh KOMANG G WIRYAWAN, ANURAGA JAYANEGARA dan AMLIUS THALIB.
Salah satu penyebab rendahnya produktivitas ternak ruminansia adalah tidak terpenuhinya asupan nutrien. Penyediaan hijauan berkualitas tinggi seperti rumput unggul dan leguminosa dibatasi oleh kompetisi lahan dengan pertanian dan aktivitas pembangunan. Oleh karena itu, peluang pengembangan usaha peternakan adalah pemanfaatan hasil samping pertanian dan perkebunan sebagai pakan ternak. Hasil samping pertanian dan perkebunan pada umumnya memiliki kualitas rendah yang ditandai dengan serat kasar tinggi dan kadar protein rendah. Karakteristik tersebut ditemukan pada hasil samping kelapa sawit yang tersedia dalam jumlah yang melimpah di Indonesia sebagai negara produsen terbesar di dunia. Hasil samping kelapa sawit diantaranya adalah pelepah, daun, serat perasan buah, tandan kosong, lumpur dan bungkil inti sawit. Pakan berserat tinggi tidak hanya menyebabkan terjadinya penurunan efisiensi penggunaan pakan dan produktivitas ternak, tetapi juga meningkatkan emisi metana sebagai gas rumah kaca. Pembentukan metana di dalam rumen mengakibatkan terjadinya kehilangan energi pakan tercerna.
Tantangan rendahnya produktivitas ternak dan dampak negatif emisi gas metana dari ternak ruminansia perlu dijawab dengan inovasi teknologi yang mampu memperbaiki sistem manajemen pakan. Salah satu pendekatan adalah dengan melakukan optimalisasi dan manipulasi ekosistem mikroba rumen, misalnya dengan menggunakan probiotik. Spesies mikroba yang sering digunakan sebagai probiotik dan telah dilaporkan mampu meningkatkan kecernaan pakan, konversi pakan serta produktivitas ternak adalah Saccharomyces cerevisiae. Selain itu, mikroba acetogen (Acetoanaerobium noterae) juga dilaporkan mampu mengurangi emisi metana enterik. Acetoanaerobium noterae memiliki kemampuan sebagai inhibitor metanogenesis dengan menggunakan hidrogen untuk membentuk asetat di dalam rumen. Pengaruh kombinasi suplementasi Saccharomyces cerevisiae dan Acetoanaerobium noterae terhadap metanogenesis dan fermentasi rumen menjadi menarik untuk diamati. Selanjutnya, suplementasi senyawa fitogenik seperti saponin dan tanin dilaporkan mampu mengurangi emisi metana. Suplementasi mineral Fe, Mn, Zn, Cu, dan Mg terbukti meningkatkan aktivitas mikroba rumen dan keceraan pakan. Spesies tanaman yang mengandung cukup banyak mineral dan senyawa fitogenik adalah tanaman pisang (Musa paradisiaca).
Substrat terdiri dari tepung daun dan pelepah, lumpur, serta bungkil inti sawit dengan perbandingan 60%, 10% dan 30%. Perlakuan yang diberikan adalah suplementasi probiotik (faktor A) dan suplementasi tepung batang pisang (faktor B), yaitu ; substrat, tanpa suplementasi/kontrol (A0B0); substrat + Saccharomyces cerevisiae (A1BO); substrat + Acetoanaerobium noterae (A2B0); substrat + Saccharomyces cerevisiae + Acetoanaerobium noterae (A3B0); substrat + tepung batang pisang (A0B1); substrat + Saccharomyces cerevisiae + tepung batang pisang (A1B1); substrat + Acetoanaerobium noterae + tepung batang pisang (A2B1); substrat + Saccharomyces cerevisiae + Acetoanaerobium noterae + tepung batang pisang (A3B1). Setiap perlakuan diinkubasi in vitro dengan larutan buffer (pH 6.9) dan rumen sebanyak empat ulangan (setiap ulangan diwakili oleh tiga botol inkubasi) selama 48 jam pada suhu 39oC. Produksi gas total dan metana diukur pada interval waktu tertentu. Kecernaan, volatile fatty acids (VFA), ammonia, dan populasi mikroba dihitung setelah inkubasi. Penurunan emisi metana dihitung dengan membandingkan produksi metana dengan produksi gas total. Peningkatan efisiensi energi pakan didekati dengan menghitung produksi gas metana per gram bahan kering tercerna.
Hasil penelitian menunjukkan bahwa secara umum suplementasi tepung batang pisang meningkatkan kecernaan pakan, sedangkan suplementasi probiotik (Saccharomyces cerevisiae atau/dan Acetoanaerobium noterae) efeknya kurang terlihat. Kecernaan bahan kering dan bahan organik substrat terbaik ditunjukkan oleh perlakuan A1B1, yaitu sebesar 20.25% dan 13.66% dibandingkan dengan kontrol. Semua perlakuan secara numerik terlihat memberikan efek terhadap penurunan konsentrasi CH4 pada lama inkubasi 12, 24 dan 48 jam dibandingkan
dengan perlakuan kontrol. Potensi penurunan konsentrasi metana secara umum diperlihatkan oleh perlakuan A2B0 dan A3B0, yaitu sebesar 16.98% (P<0.05) dan 15.58% pada inkubasi 12 jam; 12.51% dan 12.98% pada inkubasi 24 jam; serta 10.55% dan 10.77% pada inkubasi 48 jam. Efisiensi energi pakan terbaik ditunjukkan oleh A0B1, yaitu mengalami peningkatan sebesar 14.16% dibandingkan dengan kontrol. Perlakuan yang lebih toleran untuk tujuan peningkatan kecernaan pakan sekaligus penurunan emisi metana dan peningkatan efisiensi energi pakan adalah A3B1. Perlakuan ini meningkatkan kecernaan bahan kering pakan sebesar 14.58%, menurunkan emisi metana sebesar 7.71% dan meningkatkan efisiensi energi pakan sebesar 11.75% dibandingkan dengan kontrol.
Kesimpulan dari penelitian ini adalah suplementasi tepung batang pisang memiliki interaksi positif dengan Saccharomyces cerevisiae dan Acetoanaerobium noterae dalam meningkatkan kecernaan substrat secara in vitro. Suplementasi Acetoanaerobium noterae dan kombinasinya dengan Saccharomyces cerevisiae berpotensi menurunkan emisi metana, namun peluang kompetisinya perlu ditingkatkan dengan penambahan defaunator. Kombinasi suplementasi terbaik untuk meningkatkan kecernaan dan efisiensi energi pakan serta menurunkan emisi gas metana adalah kombinasi suplementasi tepung batang pisang dengan Saccharomyces cerevisiae dan Acetoanaerobium noterae.
SUMMARY
ANTONIUS. Supplementation Effectivity of Banana Stem Flours and Probiotic on Feed Digestibility and Methane Emissions of ration based on oil palm by-products. Supervised by KOMANG G WIRYAWAN, ANURAGA JAYANEGARA and AMLIUS THALIB.
One of the causes of low ruminant productivity is due to insufficient nutrient intake. Provision of high-quality forages such as grasses and legumes is limited by land competition for various developmental reasons. Therefore, an opportunity to provide forage for livestock development is through utilization of agricultural by-products as animal feeds. Agricultural by-products, however, in general have a low quality which is characterized by high fiber and low protein contents. Such characteristics are found in oil palm by-products which highly available in Indonesia as the biggest producer country worldwide. Typical by-products from oil palm plantation are oil palm midrib, oil palm leaf, oil palm trunk, oil palm frond, empty fruit bunch, oil palm sludge and palm kernel cake. High fiber diet does not only lower feed utilization efficiency and livestock productivity, but also increases the emission of methane as a green-house gases. It has been reported that formation of methane in the rumen causes a loss of digestible energy.
Challenges of low productivity and negative impact of methane emissions of ruminant livestock should be answered with innovation technology that improves feed management system. A promising approach is through optimization and manipulation of rumen microbial ecosystem, for instance, by using probiotics. A species of microbe that has been repeatedly used as probiotics and had been reported to increase feed digestibility, feed conversion and livestock productivity is yeast or Saccharomyces cerevisiae. A part from that, a class of microbes namely acetogen (Acetoanaerobium noterae) has been attempted to reduce ruminal methane emissions. Acetoanaerobium noterae has the ability as methanogenesis inhibitor by using hydrogen to form acetate in the rumen. It would be interesting to observe the effect of a simultaneous addition of Saccharomyces cerevisiae and Acetoanaerobium noterae on methanogenesis and rumen fermentation. Further, supplementation of phytogenic compounds such as saponins and tannins were reported to decrease methane emissions in the rumen. Mineral supplementation such as Fe, Mn, Zn, Cu, and Mg were reported to increase microbes activity and feed digestibility. A plant species that contain a considerable amount of mineral and phytogenic compounds is the banana plant (Musa paradisiaca).
The substrate, i.e. oil palm by-products consisted of oil palm midrib and leaf, oil palm kernel cake and oil palm sludge in the ratio of 60%, 30% and 10%, respectively. The treatments given in this study were probiotic supplementation (factor A) and supplementation of banana stem (factor B). The treatments were: substrate, no supplementation/control (A0B0), substrate + Saccharomyces cerevisiae (A1BO), substrate + Acetoanaerobium noterae (A2B0), substrate + Saccharomyces cerevisiae + Acetoanaerobium noterae (A3B0), substrate + banana stem (A0B1), substrate + Saccharomyces cerevisiae + banana stem (A1B1), substrate + Acetoanaerobium noterae + banana stem (A2B1), and substrate + Saccharomyces cerevisiae + Acetoanaerobium noterae + banana stem (A3B1). The treatments were incubated in vitro with buffered-rumen fluid in four replicates (represented by three incubation units per replicate), conducted for 48 h at 39oC. Gas production and methane emission were measured at regular time point intervals. After the incubation, digestibility, volatile fatty acids (VFA), ammonia, and microbial counts were determined. The decrease of methane emissions was calculated by comparing methane production with total gas production. The increase of feed energy efficiency were approximated by calculating the methane production per gram of dry matter digested.
Results showed that in general, supplementation of banana stem improved the digestibility of the diet, but less effect by the addition of probiotics (Saccharomyces cerevisiae and/or Acetoanaerobium) on the digestibility. The highest dry matter and organic matter digestibility were demonstrated by A1B1 treatment, namely; 20.25% and 13.66% compared with control treatment. All other supplemented treatments numerically decreased CH4 emissions at 12, 24 and
48 hours of incubation in comparison to the control treatment. Reduction potential of methane emissions was shown by treatments A2B0 and A3B0, namely; 16.98% (P<0.05) and 15.58% at 12 h of incubations; 12.51% and 12.98% at 24 h of incubations; 10.55% and 10.77% at 48 h of incubations. The best reduction of methane emission was shown by A2B0 at 12 h of incubation. The best feed energy efficiency was shown by A0B1, namely 14.16% compared with control. The best treatment was A3B1, that increased dry matter digestibility by 14.58%, decreased methane emissions by 7.71% and increased feed energy efficiency by 11.75% compared with control.
In conclusion, supplementation of banana stem flour had positive interaction effects with Saccharomyces cerevisiae and Acetoanaerobium noterae to increase feed digestibility. Acetoanaerobium noterae supplementation and its combination with Saccharomyces cerevisiae were potential to decrease ruminal methane emission, but its competition chances should be improved in the rumen, for instance, by using defaunator. The best supplementation that increase feed digestibility and energy efficiency and reduce methane emissions was supplementation combination of banana stem flour with Saccharomyces cerevisiae and Acetoanaerobium noterae.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
EFEKTIVITAS SUPLEMENTASI TEPUNG BATANG PISANG
DAN PROBIOTIK TERHADAP KECERNAAN HASIL
SAMPING KELAPA SAWIT SERTA EMISI METANA
A N T O N I U S
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014 Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Judul Tesis : Efektivitas Suplementasi Tepung Batang Pisang dan Probiotik Terhadap Kecernaan Hasil Samping Kelapa Sawit serta Emisi Metana
Nama : Antonius
NIM : D251120201
Program Study/Mayor : Ilmu Nutrisi dan Pakan
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Komang G Wiryawan Ketua
Diketahui oleh Ketua Program Studi
Ilmu Nutrisi dan Pakan
Dr Ir Dwierra Evvyernie MS MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah MScAgr
Tanggal Ujian: 10 Juli 2014 Tanggal Lulus: Prof Dr Amlius Thalib
Anggota
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahuwata’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian adalah Efektivitas Suplementasi Tepung Batang Pisang dan Probiotik Terhadap Kecernaan Hasil Samping Kelapa Sawit serta Emisi Metana.
Terima kasih penulis ucapkan kepada Bapak Prof Dr Ir Komang G
Wiryawan, Prof Dr Amlius Thalib dan Dr Anuraga Jayanegara SPt MSc selaku pembimbing, serta Dr Ir Yeni Widyastuti MSc dan Dr Ir Elizabet Wina
MSc yang telah banyak memberi saran. Penghargaan penulis sampaikan kepada Bapak Dr Ir Simon Petrus Ginting MSc dari Loka Penelitian Kambing Potong Puslitbang Peternakan yang telah banyak membantu terlaksananya penelitian ini dengan baik serta Ir Nani Irlani dan staf Laboratorium Balai Penelitian Ternak Ciawi yang telah membantu selama pengumpulan data.
Ungkapan terimakasih teristemewa penulis sampaikan kepada ayah, ibu, istri, anak serta seluruh keluarga, atas segala do’a dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2014
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR LAMPIRAN xiii
PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
Hipotesis 2
METODOLOGI PENELITIAN 3
Waktu dan Lokasi Penelitian 3
Peralatan dan Bahan 3
Preparasi Sampel dan Analisis Kimia 3
Inkubasi In Vitro 4
Analisis Pasca Inkubasi 5
Analisis Statistik 6
HASIL DAN PEMBAHASAN 6
Komposisi Nutrien Ransum dan Tepung Batang Pisang 6
Kecernaan Bahan Kering dan Bahan Organik 8
Produksi Gas dan Penurunan Emisi Metana 10
Kondisi Fermentabilitas Rumen dan Populasi Mikroorganisme 15
KESIMPULAN 19
DAFTAR PUSTAKA 20
LAMPIRAN 24
DAFTAR TABEL
1. Kandungan nutrisi substrat ransum perlakuan 6
2. Kandungan mineral dan senyawa fitogenik tepung batang Pisang Ambon 7 3. Kecernaan bahan kering dan bahan organik in vitro dari substrat hasil
samping kelapa sawit yang ditambahkan probiotik dan tepung batang
pisang 8
4. Produksi total gas dan gas metana in vitro dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada
waktu inkubasi 12, 24 dan 48 jam 11
5. Konsentrasi gas metana dalam total gas dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada
waktu inkubasi 12, 24 dan 48 jam 13
6. Produksi total gas dan metana setiap gram bahan kering tercerna dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada waktu inkubasi 12, 24 dan 48 jam 15 7. Produksi Volatile Fatty Acid dari substrat hasil samping kelapa sawit
yang ditambahkan probiotik dan tepung batang pisang 16 8. Konsentrasi pH, N-NH3, populasi bakteri dan protozoa dari substrat hasil
samping kelapa sawit yang ditambahkan probiotik dan tepung batang
DAFTAR LAMPIRAN
1. Analisis ragam kecernaan bahan kering 24
2. Analisis ragam kecernaan bahan organik 24
3. Analisis ragam produksi gas total 24
4. Analisis ragam produksi gas metana 25
5. Analisis ragam konsentrasi gas metana 25
6. Analisis ragam produksi gas total per gram bahan kering tercerna 25 7. Analisis ragam produksi metana per gram bahan kering tercerna 26
8. Analisis ragam populasi bakteri 26
9. Analisis ragam populasi protozoa 26
10. Analisis ragam konsentrasi N-NH3 27
11. Analisis ragam pH cairan rumen 27
12. Analisis ragam VFA total 27
13. Analisis ragam konsentrasi asetat 28
14. Analisis ragam konsentrasi propionat 28
15. Analisis ragam konsentrasi butirat 28
16. Analisis ragam konsentrasi iso butirat 29
17. Analisis ragam konsentrasi valerat 29
1
PENDAHULUAN
Latar Belakang
Kebutuhan pangan hewani yang berasal dari ternak ruminansia di Indonesia belum dapat terpenuhi oleh produksi daging dalam negeri. Salah satu faktor penyebabnya adalah produktivitas ternak yang rendah karena kurangnya asupan nutrisi. Penanaman hijauan pakan ternak unggul memiliki kendala keterbatasan lahan akibat kompetisi dengan tanaman pertanian serta aktivitas pembangunan. Peluang pengembangan usaha peternakan di Indonesia adalah penerapan sistem integrasi, yaitu pemanfaatan hasil samping pertanian/perkebunan sebagai pakan ternak. Luas tanam perkebunan kelapa sawit nasional tahun 2013 adalah sekitar 9 149 919 hektar (Kementerian Pertanian 2013). Setiap hektar lahan perkebunan mampu menampung sekitar 130 pohon kelapa sawit (Puastuti 2008). Setiap pohon menghasilkan hasil samping berupa 22 pelepah per tahun (Diwyanto et al. 2003) dengan rataan bobot pelepah 7 kg/pelepah (Sitompul 2003), sehingga produksi pelepah segar kelapa sawit diperkirakan sekitar 20 ton/ha/tahun. Daun kelapa sawit yang terdapat pada setiap pelepah adalah sekitar 0.5 kg (Mathius et. al. 2004), dengan demikian setiap tahunnya dihasilkan sekitar 1 430 kg daun kelapa sawit per hektar. Industri pengolahan kelapa sawit memproduksi hasil samping yang cukup banyak, diantaranya lumpur sawit sekitar 1 132 kg/ha/t, bungkil inti sawit 514 kg/ha/t, serat perasan 2 681 kg/ha/t dan tandan kosong sebesar 3 386 kg/ha/t (Mathius et al. 2003).
Produksi hasil samping perkebunan dan industri kelapa sawit yang cukup besar ini sangat berpotensi digunakan sebagai bahan pakan ternak ruminansia. Namun, penggunaannya sebagai pakan dibatasi oleh rendahnya tingkat kecernaan dan kualitas nutrisi pakan yang dicirikan dengan tingginya serat kasar dan rendahnya protein. Pakan berserat tinggi menyebabkan terjadinya penurunan kecernaan, efisiensi penggunaan pakan dan produktivitas ternak serta meningkatkan emisi gas metana enterik. Pembentukan metana di dalam rumen membutuhkan sekitar 2-15% energi yang dikonsumsi ternak, tergantung jenis dan tipe ternak, kandungan bahan organik dan komponen serat pakan, tingkat kecernaan serta kondisi lingkungan rumen (Haryanto dan Thalib 2009). Metana memberikan kontribusi emisi gas rumah kaca sekitar 18% dari total gas rumah kaca di atmosfer setelah dieqivalenkan dengan nilai GWP (Global Warming Potency) CO2 per 100 tahun (Kreuzer dan Soliva 2008).
Tantangan rendahnya produktivitas ternak dan dampak negatif emisi gas metana ini perlu dijawab dengan terobosan-terobosan perbaikan manajemen pakan. Salah satu pendekatan adalah dengan melakukan optimalisasi dan manipulasi kinerja rumen, yaitu menciptakan lingkungan rumen yang ideal untuk proses degradasi dan fermentasi mikrobial serta menghambat terjadinya proses metanogenesis. Berbagai penelitian terkait optimalisasi kinerja rumen dan penurunan emisi metana telah banyak dilakukan, diantaranya adalah suplementasi mineral, teknologi probiotik, defaunasi dan inhibitor metanogenesis.
2
secara tunggal dan kombinasi dapat meningkatkan aktivitas mikroba dan kecernaan pakan (Supriyati et al. 2000; Thalib et al. 2002). Salah satu spesies mikroorganisme yang sering digunakan sebagai probiotik dan dilaporkan dapat meningkatkan kecernaan, konversi pakan serta produktivitas ternak adalah Saccharomyces cerevisiae (Wina 2000; Ando et al. 2004; Guedes et al. 2008).
Intervensi mikroba asetogenik menjadi salah satu pendekatan yang digunakan untuk mengurangi emisi metana karena kemampuannya sebagai inhibitor metanogenesis melalui pemanfaatan H2 dalam pembentukan asetat di
dalam rumen (Fonty et al. 2007; Lopez et al. 1999). Salah satu spesies mikroba asetogenik yang dilaporkan terbukti mampu memperbaiki performa ternak dan menurunkan produksi gas metana enterik adalah Acetoanaerobium noterae (Thalib dan Widiawati 2008). Senyawa fitogenik tanaman dilaporkan mampu menurunkan emisi metana enterik, yaitu senyawa tanin dan saponin. Tanin menurunkan emisi metana dengan cara mengurangi populasi mikroba metanogen (Jayanegara et al. 2011), sedangkan saponin memiliki kemampuan untuk mendefaunasi protozoa yang menjadi media tumbuh mikroba metanogen (Makkar et al. 2007). Defaunasi protozoa memberikan efek positif terhadap penurunan konsentrasi H2 yang menjadi senyawa pembentuk metana di dalam rumen (Thalib
2004).
Salah satu tanaman yang yang mengandung banyak mineral serta senyawa fitogenik adalah tanaman pisang. Batang pohon pisang memiliki kandungan tanin, mineral K, Mg, Fe, Ca, Mn, Zn, Cu dan mineral lain (Wina 2001). Produksi buah pisang nasional pada tahun 2012 adalah sekitar 6 189 043 ton dengan sentra produksi di Jawa Timur, Jawa Barat, Lampung, Jawa Tengah, Sumatera Utara, Banten, Bali dan beberapa daerah lainnya (Kementrian Pertanian 2013). Setiap panen, budidaya pohon pisang menghasilkan hasil samping berupa batang, daun dan tandan pisang. Hasil samping ini sangat potensial untuk dijadikan feed supplement yang mengandung mineral dan senyawa fitogenik.
Teknologi suplementasi mineral, probiotik, defaunasi dan inhibitor metanogenesis sudah dilakukan secara terpisah, namun interaksi penggunaan komponen teknologi tersebut secara bersamaan belum banyak dilakukan. Oleh sebab itu, penelitian ini bertujuan untuk menginvestigasi efek kombinasi komponen-komponen tersebut terhadap kecernaan pakan dan emisi metana secara in vitro.
Tujuan
Tujuan penelitian ini adalah menginvestigasi efek kombinasi suplementasi probiotik Acetoanaerobium noterae, Saccharomyces cerevisiae dan tepung batang pisang terhadap kecernaan substrat berbasis hasil samping kelapa sawit dan emisi gas metana enterik secara in vitro.
Hipotesis
3
METODOLOGI PENELITIAN
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan pada bulan September sampai dengan Desember 2013. Perbanyakan Saccharomyses cerevisiae dilakukan di Laboratorium Mikrobiologi Fakultas Peternakan IPB. Perbanyakan Acetoanaerobium noterae, uji invitro dan analisis peubah dilakukan di Laboratorium Balai Penelitian Ternak, Ciawi Bogor.
Peralatan dan Bahan
Peralatan yang digunakan pada penelitian ini diantaranya adalah botol inkubasi, inkubator, hot plate, laminar flow, tube roller, mikroskop, centrifuge, spektrofotometer UV-vis, vortex, tabung reaksi, labu reaksi, erlenmeyer, timbangan digital, pipet, gelas ukur, freezer, sonicator (ultrasonic water bath), oven, cawan Conway dan peralatan pendukung lainnya. Bahan yang digunakan diantaranya biakan Saccharomyses cerevisiae dan Acetoanaerobium noterae, basal medium, vitamin solution, trace element sol, basal solution, reducing agen, media PDA dan PDB, tepung batang pisang, pelarut aseton dan methanol, cairan rumen kambing, serta substrat (pakan) yang terdiri dari bungkil inti sawit, lumpur sawit, serta tepung pelepah dan daun sawit.
Preparasi Sampel dan Analisis Kimia
Substrat in vitro terdiri dari tepung pelepah dan daun sawit, bungkil inti sawit serta lumpur sawit. Pelepah dan daun diambil dari beberapa pohon, dicacah, dioven 60o C selama 3 hari hingga kering dan digiling halus (1 mm). Tepung pelepah dan daun diaduk secara homogen dengan solid dan bungkil inti sawit dengan formulasi 60: 10: 30. Kandungan nutrien sampel pakan dianalisa dengan menggunakan metode AOAC (1997) dan Van Soest et al. (1991).
4
ditumbuhkan pada media cair yang terdiri dari basal medium, vitamin solution dan trace element solution sesuai prosedur Ogimoto dan Imai (1981).
Inkubasi In Vitro
Inkubasi in vitro dilakukan berdasarkan metode Theodorou dan Brooks (1990). Substrat dimasukkan kedalam setiap botol inkubasi sebanyak 1 gram dan tepung batang pisang sebanyak 100 mg sesuai perlakuan. Media inkubasi untuk setiap botol adalah 90 ml larutan buffer (pH 7.9) yang terdiri dari 86 ml basal solution dan 4 ml reducing agent, serta 10 ml cairan rumen kambing. Cairan rumen diambil pada pagi hari sebelum pemberian pakan dari 3 ekor kambing jantan PE dengan metode stomach tube. Kambing yang digunakan mengkonsumsi rumput gajah, silase daun tebu dan konsentrat komersial SP-25. Koleksi cairan rumen dibawa ke laboratorium, disaring dengan saringan nilon
berukuran 100 μm dan ditambahkan ke dalam larutan buffer (100 ml/900 ml larutan buffer). Media inkubasi dijenuhkan dengan gas CO2 hingga kondisi media
menjadi anaerob, kemudian dimasukkan ke dalam setiap botol inkubasi sebanyak 100 ml. Sediaan cair Acetoanaerobium noterae dan Saccharomyces cerevisiae ditambahkan ke dalam botol sebanyak 1 ml (mengandung 109 dan 106 cfu/ml) sesuai perlakuan. Botol inkubasi segera ditutup dan dimasukkan ke dalam water bath incubator (39oC) dan diinkubasi selama 48 jam.
Perlakuan yang diberikan adalah suplementasi probiotik (faktor A) dan suplementasi tepung batang pisang (faktor B), terdiri dari : substrat, tanpa suplementasi (A0B0); substrat + Saccharomyces cerevisiae (A1BO); substrat + Acetoanaerobium noterae (A2B0); substrat + Saccharomyces cerevisiae + Acetoanaerobium noterae (A3B0); substrat + tepung batang pisang (A0B1); substrat + Saccharomyces cerevisiae + tepung batang pisang (A1B1); substrat + Acetoanaerobium noterae + tepung batang pisang (A2B1); substrat + Saccharomyces cerevisiae + Acetoanaerobium noterae + tepung batang pisang (A3B1).
Pengambilan total gas dan gas metana dilakukan setiap 3 jam sekali hingga 12 jam pertama dan selanjutnya setiap 6 jam hingga 48 jam inkubasi dengan menggunakan metode Fievez et al. (2005). Gas diambil dengan menggunakan syringe (Sigma-Aldrich Z314382-1EA, Poulten & Graf GmbH Wertheim, Germany) dan dialirkan ke dalam erlenmeyer (kapasitas 1 liter) yang berisi larutan NaOH 5 N, terhubung dengan syringe pengukur volume gas metana. Gas CO2 terperangkap dalam larutan NaOH 5 N, sedangkan gas metana masuk ke
5
Analisis Pasca Inkubasi
Kecernaan bahan kering dan bahan organik diukur setelah 48 jam inkubasi. Sinterglass dikeringkan terlebih dahulu dengan oven 105o C selama 24 jam dan ditimbang untuk mengetahui berat masing-masingnya. Residu inkubasi dikeringkan dengan oven 105o C selama 24 jam untuk penghitungan kecernaan bahan kering dan ditanur 550o C selama 6 jam untuk penghitungan kecernaan bahan organik. Sampel pada setiap botol in vitro disaring menggunakan sinterglass dengan bantuan vacuum pump. Supernatan hasil saringan diambil sebagai sampel analisa volatile fatty acids (VFA), ammonia (NH3), populasi
bakteri dan protozoa.
Kandungan VFA dianalisis dengan menggunakan Gas Liquid Chromatography (GLC Bruker Scion 436-GC, Bruker Daltonik GmbH, Bremen, Jerman) menggunakan kolom (BR-Wax fame, mmlD 0.32, 0.25 lm df) dan detector FID. Sediaan supernatan yang diperoleh dari inkubasi in vitro ditambahkan dengan 3 mg sufosalicylic acid dihydrate, disentrifugasi selama 10 menit dengan kecepatan 12 000 rpm pada suhu 7o C dan kemudian diinjeksikan ke dalam kolom GLC. Kuantifikasi individu VFA dilakukan dengan membandingkannya dengan standar eksternal. Satuan VFA individu yang
diperoleh adalah dalam μmol/ml atau mM. Kandungan total VFA didapatkan melalui penjumlahan masing-masing VFA individu penyusunnya.
Analisa kandungan ammonia menggunakan metode Conway (1950). Asam borat 3% diinjeksikan ke bagian tengah cawan Conway sebanyak 3 ml dan kemudian ditambahkan 1 tetes larutan indikator BCG (brom cresol green) dan red methyl indicator. Supernatan dan larutan NaOH diinjeksikan sebanyak 1 ml pada bagian yang berbeda (kiri atau kanan) cawan Conway. Cawan ditutup dan kemudian digoyangkan perlahan untuk mencampurkan NaOH 20% dan supernatan. Sampel dibiarkan selama 24 jam dan dititrasi dengan HCl hingga asam borat berubah warna dari biru menjadi merah muda. Konsentrasi NH3
dihitung dengan mengalikan volume HCl yang diperlukan untuk titrasi dengan konsentrasi HCl dan faktor pengenceran.
6
Analisis Statistik
Percobaan dilakukan dengan rancangan acak kelompok pola faktorial (4 x 2) dengan 4 kelompok sebagai ulangan dan setiap kelompok diwakili oleh 2 botol inkubasi. Data dianalisis menggunakan software statistik SPSS versi 16.0. dengan prosedur general linear model (GLM). Nilai rata-rata antara perlakuan diuji lanjut dengan Duncan’s Multiple Range Test.
HASIL DAN PEMBAHASAN
Komposisi Nutrien Substrat dan Tepung Batang Pisang
Komposisi nutrien substrat berdasarkan analisis proksimat dan analisis komposisi serat diperlihatkan pada Tabel 1. Berdasarkan nilai nutrisinya, ransum ini hanya cocok untuk ternak perbesaran, penggunaan untuk ternak potong dan perah perlu perbaikan komposisi dan tambahan bahan pakan lain. Substrat yang digunakan pada penelitian ini merupakan hasil samping perkebunan dan industri kelapa sawit tanpa pengolahan dan peningkatan kualitas nutrisi. Hal ini dilakukan untuk mengetahui kemampuan optimum dari Saccharomyces cerevisiae, Asetoanaerobium noterae dan tepung batang pisang dalam mempengaruhi kecernaan pakan dan emisi metana enterik pada substrat yang memiliki kandungan serat yang tinggi.
Tabel 1. Kandungan nutrisi substrat ransum perlakuan
Nutrien Kandungan selulosa dan lignin), TDN (total digestible nutrient : total nutrisi tercerna).
7
pada penelitian ini dinilai cukup tinggi, yaitu sebesar 81.28%. Sebagian besar (67.58%) dari NDF ini merupakan fraksi selulosa dan lignin yang biasanya membentuk ikatan lignoselulosa dan sulit dicerna oleh mikroorganisme rumen. Selain mempengaruhi kecernaan, kandungan NDF juga berpengaruh terhadap emisi metana. Hasil akhir fermentasi selulosa dan hemiselulosa di dalam rumen adalah berupa asam asetat. Pembentukan asam asetat menghasilkan gas hidrogen (H2) yang digunakan sebagai substrat pada reaksi metanaogenesis. Semakin
banyak asam asetat yang terbentuk, maka H2 yang dihasilkan menjadi meningkat
dan produksi metana semakin tinggi. Produksi metana meningkat seiring dengan meningkatnya kandungan NDF (Jayanegara et al. 2009).
Selain nutrisi substrat, kandungan mineral dan senyawa fitogenik yang terkandung dalam tepung batang pisang juga menentukan kecernaan dan emisi metana enterik. Suplementasi tepung batang pisang diharapkan mampu memberikan kontribusi terhadap optimalisasi fermentasi rumen melalui dua mekanisme, yaitu 1) memberikan sumbangan mineral yang dibutuhkan oleh mikroba yang menguntungkan di dalam rumen, dan 2) mendefaunasi protozoa serta mengurangi populasi mikroba metanogen melalui intervensi senyawa fitogenik yang dimilikinya. Kandungan mineral dan senyawa fitogenik tepung batang pisang disajikan pada Tabel 2.
Kandungan tanin tepung batang pisang pada penelitian ini relatif lebih kecil dari pada tanin fraksi cair batang pisang yang dilaporkan Wina (2001), yaitu 0.09% vs 0.46%. Perbedaan kandungan tanin ini diduga karena berbedanya daerah pengambilan batang pisang, yaitu dari Propinsi Bali, sementara pada penelitian ini sampel diambil di daerah Ciawi Bogor Jawa Barat. Selain mengandung tanin, tepung batang pisang juga mengandung saponin sebesar 0.54%. Dua senyawa fitogenik yang terkandung dalam tepung batang pisang ini diduga memberikan pengaruh terhadap kecernaan substrat, emisi metana, populasi mikroba dan kondisi fermentabilitas rumen.
Tabel 2. Kandungan mineral dan senyawa fitogenik tepung batang Pisang Ambon
8
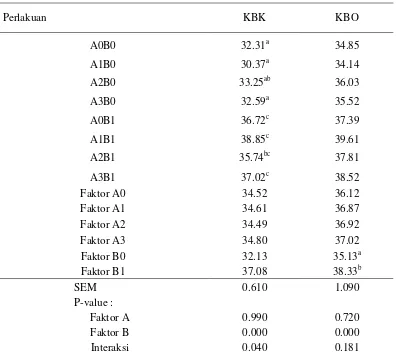
Kecernaan Bahan Kering dan Bahan Organik
Kecernaaan bahan kering dan bahan organik substrat sebagaimana tersaji pada Tabel 3 dinilai cukup rendah, yaitu di bawah 40%. Rendahnya tingkat kecernaan diduga terjadi karena seluruh komponen bahan pakan yang digunakan berasal dari hasil samping kelapa sawit yang memiliki kualitas nutrisi yang rendah (Tabel 1). Selain itu, nilai kecernaan dalam penelitian ini hanya mewakili proses pencernaan dalam rumen saja tanpa mempertimbangkan pencernaan pasca-rumen. Secara umum, suplementasi tepung batang pisang (faktor B) meningkatkan kecernaan substrat, sedangkan pemberian probiotik (faktor A) tidak memberikan pengaruh yang nyata.
Tabel 3. Kecernaan bahan kering dan bahan organik in vitro dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang
Perlakuan KBK KBO
A0B0 32.31a 34.85
A1B0 30.37a 34.14
A2B0 33.25ab 36.03
A3B0 32.59a 35.52
A0B1 36.72c 37.39
A1B1 38.85c 39.61
A2B1 35.74bc 37.81
A3B1 37.02c 38.52
Faktor A0 34.52 36.12
Faktor A1 34.61 36.87
Faktor A2 34.49 36.92
Faktor A3 34.80 37.02
Faktor B0 32.13 35.13a
Faktor B1 37.08 38.33b
SEM 0.610 1.090
P-value :
Faktor A 0.990 0.720
Faktor B 0.000 0.000
Interaksi 0.040 0.181
Superskrip yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P<0,05); P-value (tingkat signifikansi); SEM (Standard Error Mean); A0 (tanpa probiotik); A1 (S. cerevisiae); A2 (A. noterae); A3 (S. cerevisiae dan A. noterae ); B0 (tanpa tepung batang pisang); B1 (tepung batang pisang); KBK (Kecernaan Bahan Kering); KBO (Kecernaan Bahan Organik).
9
yaitu 38.85% dan 39.61% atau mengalami peningkatan kecernaan sebesar 20.25% dan 13.65% dibandingkan dengan kontrol. Peningkatan kecernaan bahan kering dan bahan organik pakan diduga terjadi karena sumbangan nutrien yang dibutuhkan untuk pertumbuhan dan aktivitas mikroba rumen oleh tepung batang pisang dan Saccharomyces cerevisiae. Batang pisang memiliki beberapa kandungan mineral yang dibutuhkan oleh mikroba rumen, diantaranya adalah mineral K, Fe, Zn dan Ca (Wina 2001). Kandungan mineral tepung batang pisang yang digunakan dalam penelitian ini sebagaimana yang tersaji pada Table 2 diantaranya adalah kalium (1.62% bahan kering), magnesium (0.13% bahan kering), besi (150 ppm), mangan (99.53 ppm), zink (20 ppm), tembaga (1.98 ppm) dan diperkirakan masih banyak mineral lain yang dikandungnya. Thalib et al. (2002) melaporkan bahwa suplementasi Faktor Pertumbuhan Mikroba yang salah satu komposisinya adalah Zn dan Cu terbukti meningkatkan kecernaan pakan yang terdiri dari rumput gajah dan konsentrat hingga 49% dibandingkan dengan pakan tanpa suplementasi.
Komposisi kimia sel Saccharomyces cerevisiae terdiri atas protein kasar 50-52%, karbohidrat 30-37%, lemak 4-5% dan mineral 7-8% (Reed dan Nagodawithana 1991). Saccharomyces cerevisiae merupakan sumber vitamin, enzim, nutrien dan kofaktor lain bagi mikroba dan pencernaan di dalam rumen (Dawson 1990). Chevaux dan Fabre (2007) menjelaskan bahwa selain menyediakan nutrisi dan kofaktor penting yang merangsang aktivitas mikroba rumen, Saccharomyces cerevisiae juga berperan dalam mengontrol lingkungan rumen agar lebih anaerob. Mekanisme kerja Saccharomyces cerevisiae di dalam rumen secara singkat dijelaskan oleh Yoon dan Stern (1996) bahwa aktivitas respiratory Saccharomyces cerevisiae mengkonsumsi oksigen di dalam rumen, sehingga kondisi anaerob dapat dipertahankan. Kondisi ini menstimulasi pertumbuhan populasi mikroba rumen yang diikuti dengan meningkatnya pemanfaatan amonia dan asam laktat sehingga pH rumen menjadi stabil. Kondisi anaerob dan pH rumen yang stabil meningkatkan populasi dan aktivitas mikroba, sehingga proses fermentasi menjadi optimal dan kecernaan pakan menjadi meningkat.
Suplementasi Saccharomyces cerevisiae secara tunggal tidak memberikan pengaruh terhadap peningkatan kecernaan bahan kering dan bahan organik pakan. Hal ini diduga terjadi karena kurangnya dosis pemberian pada kondisi pakan rendah nutrisi yang terdiri dari hasil samping kelapa sawit. Protozoa memiliki sifat fagositosit terhadap mikroba rumen seperti bakteri dan fungal zoospores Theodorou dan France (2005). Aktivitas fagositosit protozoa akan semakin tinggi pada kondisi pakan yang mengandung sedikit karbohidrat mudah dicerna (Erwanto 1995). Rendahnya kualitas nutrisi substrat memicu protozoa untuk mengkonsumsi Saccharomyces cerevisiae, sehingga aktivitas respiratory dan sumbangan nutrien yang diharapkan dari Saccharomyces cerevisiae menjadi tidak bisa dimanfaatkan oleh bakteri dan mikroba rumen lain pencerna substrat. Suplementasi Saccharomyces cerevisiae secara tunggal diduga meningkatkan aktivitas dan pertumbuhan protozoa, sehingga aktifitas fagositosit menjadi semakin tinggi juga dan berefek negatif terhadap kecernaan substrat.
10
fagositosit dan pertumbuhan protozoa dihambat oleh senyawa fitogenik saponin yang terkandung dalam tepung batang pisang. Sehingga peran Saccharomyces cerevisiae menjadi lebih optimal untuk meningkatkan kecernaan.
Suplementasi tepung batang meningkatkan kecernaan bahan organik pakan secara numerik sebesar 7.29 %. Namun, peningkatan yang terjadi belum signifikan. Hal ini terjadi karena adanya interaksi tanin dengan bakteri yang mengakibatkan pemanfaatan mineral menjadi kurang optimal. Tanin dapat berikatan dengan membran sel bakteri rumen serta menghambat pertumbuhan dan aktivitas enzim yang dihasilkannya (Smith et al. 2005). Tanin menurunkan beberapa aktivitas enzim rumen seperti carboxymethyl cellulase, urease, protease, glutamine dehydrogenase dan alanine transferase (Makkar et al. 1998). Selain itu, tanin juga memiliki kemampuan untuk membentuk senyawa komplek dengan protein, selulosa, hemiselulosa, pektin dan mineral sehingga menyebabkan kecernaan zat makanan menjadi turun (Wiryawan dan Marliana 2008).
Kombinasi pemberian tepung batang pisang dengan Saccharomyces cerevisiae memiliki dampak yang positif terhadap peningkatan kecernaan bahan organik pakan, yaitu dari 7.29% menjadi 13.65% dibandingkan dengan perlakuan kontrol. Peningkatan kecernaan bahan organik ini diduga terjadi melaui beberapa mekanisme, yaitu ; 1) aktivitas respiratory Saccharomyces cerevisiae mengkondisikan rumen menjadi lebih anaerob, sehingga aktivitas mikroba untuk memanfaatkan mineral tepung batang pisang dan mencerna pakan menjadi lebih optimal, 2) Saccharomyces cerevisiae menjadi sumber nutrien yang dibutuhkan oleh mikroba rumen, sehingga kemampuan mikroba dalam mencerna pakan menjadi lebih baik, 3) enzim dan kofaktor lain yang dihasilkan oleh Saccharomyces cerevisiae berkompetisi dengan tanin tepung batang pisang dalam mempengaruhi kinerja mikroba rumen dan enzim yang dihasilkannya, sehingga efek negatif tanin menjadi berkurang. Saccharomyces cerevisiae meningkatkan populasi dan pertumbuhan mikroba selulolitik (Miller dan Wolin 2002), mikroba pengguna asam laktat (Jouany 2001) dan bakteri proteolitik (Yoon dan Stern 1996). Suplementasi Saccharomyces cerevisiae merangsang metabolisme mikroba rumen (Oeztuerk et al. 2005), meningkatkan kestabilan pH rumen (Thrune et al. 2009), menurunkan konsentrasi asam laktat rumen (Guedes et al. 2008) dan
meningkatkan konsentrasi asam asetat dan propionat di dalam rumen (O’connor et al. 2002).
Beberapa penjelasan diatas memberikan gambaran bahwa sebagaimana halnya Saccharomyces cerevisiae, suplementasi tepung batang pisang pada pakan yang memiliki kualitas nutrisi rendah sebaiknya tidak diberikan secara tunggal. Pengkombinasian suplementasi memberikan efek yang saling menguntungkan. Saponin tepung batang pisang melindungi Saccharomyces cerevisiae dari aktivitas fagositosit protozoa dan sebaliknya efek negatif tanin yang menghambat pemanfaatan mineral tepung batang pisang menjadi berkurang karena adanya aktivitas enzimatis Saccharomyces cerevisiae.
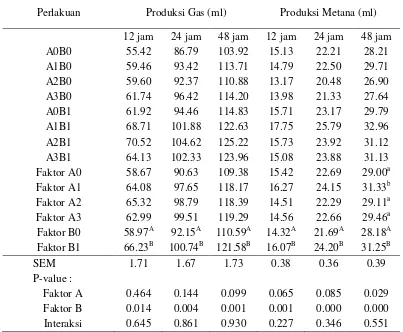
Produksi Gas dan Penurunan Emisi Metana
11
(suplementasi tepung batang pisang) memberikan pengaruh secara nyata (P<0.05) terhadap peningkatan produksi gas, sedangkan faktor A (probiotik) dan interaksi antar faktor memberikan pengaruh yang tidak signifikan. Hal ini bisa dipahami karena produksi gas merupakan gambaran dari tingkat kecernaan pakan. Produksi gas adalah hasil fermentasi substrat secara langsung berupa CO2 dan CH4 serta
secara tidak langsung melalui mekanisme buffering yakni gas CO2 yang
dilepaskan dari buffer bikarbonat yang diproduksi selama proses fermentasi (Getachew et al. 1998). Secara numerik, setiap perlakuan suplementasi probiotik terlihat memberikan efek terhadap peningkatan produksi gas, baik pada waktu inkubasi 12, 24 maupun 48 jam. Tidak signifikannya peningkatan produksi gas diduga karena jumlah pemberian probiotik sebanyak 1 ml/100 ml larutan inkubasi in vitro masih belum cukup untuk substrat yang terdiri dari hasil samping kelapa sawit. Secara umum seperti terlihat pada Tabel 4, produksi gas merupakan gambaran tingkat kecernaan pakan, semakin lama proses inkubasi, maka semakin banyak total gas yang dihasilkan. Laju produksi gas akan semakin berkurang seiring dengan lamanya inkubasi dan berkurangnya substrat yang bisa dicerna. Tabel 4. Produksi total gas dan gas metana in vitro dari substrat hasil samping
kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada waktu inkubasi 12, 24 dan 48 jam
Perlakuan Produksi Gas (ml) Produksi Metana (ml)
12 jam 24 jam 48 jam 12 jam 24 jam 48 jam
A0B0 55.42 86.79 103.92 15.13 22.21 28.21
A1B0 59.46 93.42 113.71 14.79 22.50 29.71
A2B0 59.60 92.37 110.88 13.17 20.48 26.90
A3B0 61.74 96.42 114.20 13.98 21.33 27.64
A0B1 61.92 94.46 114.83 15.71 23.17 29.79
A1B1 68.71 101.88 122.63 17.75 25.79 32.96
A2B1 70.52 104.62 125.22 15.73 23.92 31.12
A3B1 64.13 102.33 123.96 15.08 23.88 31.13
Faktor A0 58.67 90.63 109.38 15.42 22.69 29.00a
Faktor A1 64.08 97.65 118.17 16.27 24.15 31.33b
Faktor A2 65.32 98.79 118.39 14.51 22.29 29.11a
Faktor A3 62.99 99.51 119.29 14.56 22.66 29.46a
Faktor B0 58.97A 92.15A 110.59A 14.32A 21.69A 28.18A
Interaksi 0.645 0.861 0.930 0.227 0.346 0.551
12
Peningkatan produksi gas biasanya diikuti oleh peningkatan produksi CH4.
Hal ini terjadi karena CH4 merupakan komponen gas hasil fermentasi substrat
pakan di dalam rumen. Semakin tingggi produksi gas, maka produksi CH4 juga
akan semakin tinggi. Oleh sebab itu, parameter yang lebih relevan untuk menilai tingkat penurunan emisi CH4 asal ternak ruminansia adalah perbandingan
produksi gas CH4 dengan total produksi gas atau konsentrasi CH4 dalam total gas
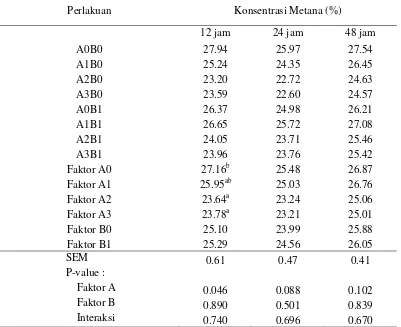
yang diproduksi. Semakin rendah nilai perbandingan menunjukkan semakin rendah konsentrasi CH4 dan semakin menurun tingkat emisi metana.
Nilai perbandingan produksi CH4 terhadap total produksi gas tersaji pada
Tabel 5. Pemberian tepung batang pisang tidak memberikan efek yang positif terhadap penurunan konsentrasi metana. Suplementasi A. noterae dan kombinasinya dengan S. cerevisiae pada 12 jam inkubasi menurunkan konsentrasi CH4 secara signifikan (P<0,05). Suplementasi Asetoanaerobium noterae selama
12 jam inkubasi menurunkan konsentrasi CH4 (P<0.05) sebesar 16.98%, yaitu dari
27.97% (kontrol) menjadi 23.20%. Penurunan konsentrasi CH4 diduga terjadi
karena aktivitas inhibisi pembetukan CH4 oleh Asetoanaerobium noterae melalui
mekanisme pemanfaatan hidrogen bebas di dalam rumen untuk membentuk asetat. Mikroba asetogenik (termasuk Asetoanarobium noterae) memanfaatkan CO2 dan
H2 untuk pertumbuhannya dan menghasilkan asam asetat sebagai produk
metabolit (Lopez et al. 1999). Mikroba asetogenik termasuk golongan mikroba hidrogenotropik yang mampu mereduksi karbondioksida membentuk asetat, mereduksi sulfat membentuk hydrogen sulfida, dan mereduksi fumarat membentuk suksinat (Morvan et al. 1996).
Suplementasi Asetoanaerobium noterae memberikan pengaruh yang nyata terhadap penurunan emisi metana pada 12 jam inkubasi, namun pengaruh suplementasi semakin berkurang seiring dengan lama ikubasi. Hal ini diduga berkaitan dengan daya kompetisi Asetoanaerobium noterae terhadap mikroba metanogenik. Oleh sebab itu, penggunaan Asetoanaerobium noterae sebagai pakan tambahan perlu mempertimbangkan kombinasi teknologi yang mampu meningkatkan daya kompetisi Asetoanaerobium noterae di dalam rumen. Salah satu pendekatan adalah penggunaan saponin sebagai agen defaunator protozoa. Penurunan populasi protozoa berimplikasi terhadap menurunnya populasi mikroba metanogenik. Kandungan saponin di dalam tepung batang pisang diduga belum memadai untuk mendefaunasi protozoa. Penambahan senyawa saponin pada perlakuan suplementasi menjadi topik yang menarik untuk diteliti lebih lanjut.
Secara numerik, semua perlakuan terlihat memberikan efek terhadap penurunan konsentrasi CH4 pada lama inkubasi 12, 24 dan 48 jam jika
13
Tabel 5. Konsentrasi gas metana dalam total gas dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada waktu inkubasi 12, 24 dan 48 jam
Perlakuan Konsentrasi Metana (%)
Superskrip yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P<0,05); P-value (tingkat signifikansi); SEM (Standard Error Mean); A0 (tanpa probiotik); A1 (S. cerevisiae); A2
alternatif elektron akseptor yang memiliki kemampuan yang lebih baik dari CO2
dalam mengikat H2. Senyawa fitogenik yang terdapat dalam tepung batang pisang
menghambat pertumbuhan dan populasi mikroba metanogen secara langsung serta mendefaunasi protozoa yang menjadi media tumbuh mikroba metanogen secara tidak langsung. Selain mengandung senyawa fitogenik, tepung batang pisang juga mengandung banyak mineral yang diantaranya berpotensi untuk mengikat H2
seperti Fe. Ion Fe (Fe3+) memiliki afinitas lebih tinggi terhadap hidrogen dibandingkan CO2 (Thalib, 2004).
Suplementasi Asetoanaerobium noterae berperan sebagai agen inhibitor metanogenesis dengan memanfaatkan hidrogen untuk pembentukan asetat. Saccharomyces cerevisiae dapat merangsang pertumbuhan dan aktivitas mikroba asetogenik dalam memanfaatkan H2 di dalam rumen untuk membentuk asam
14
mikroba metanogenik dalam memanfaatkan hidrogen di dalam rumen diduga meningkat karena sumbangan vitamin, enzim dan nutrisi oleh Saccharomyces cerevisiae.
Peran Asetoanaerobium noterae, Saccharomyces cervisiae, senyawa fitogenik dan mineral tepung batang pisang secara umum belum mampu memberikan pengaruh yang signifikan terhadap penurunan konsentrasi metana. Hal ini diduga terjadi karena rendahnya jumlah pemberian kedua jenis probiotik dan tepung batang pisang. Satu ml probiotik per 100 ml larutan inkubasi in vitro dan 1 mg tepung batang pisang per 1 ml larutan inkubasi in vitro pada substrat pakan bernutrisi rendah seperti hasil samping kelapa sawit belum mampu menurunkan konsentrasi metana secara signifikan. Peningkatan jumlah pemberian dan imbangan ke tiga komponen merupakan topik yang menarik untuk diteliti lebih lanjut, baik pada substrat yang sama maupun pada substrat yang memiliki nutrisi yang lebih baik.
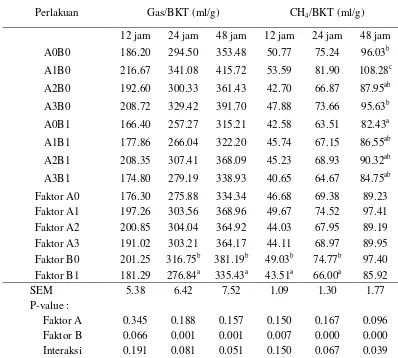
Hubungan peningkatan efisiensi energi pakan dengan produksi gas total dan metana dapat dinilai dengan membandingkan produksi gas total dan metana dengan setiap gram bahan kering tercerna. Semakin rendah nilai produksi (ml/g) menunjukkan semakin tinggi tingkat efisiensi energi pakan. Produksi gas total secara umum memiliki tren yang sama dengan produksi gas metana pada setiap perlakuan, sebagaimana yang tersaji pada Tabel 6.
Fokus kajian pada penelitian ini adalah upaya mengurangi kehilangan energi akibat pembetukan metana di dalam rumen. Oleh sebab itu, pembahasan lebih dititik beratkan pada efisiensi energi akibat pembentukan metana. Nilai produksi metana terendah (P<0.05) ditunjukkan oleh suplementasi tepung batang pisang, yaitu 82.43 ml/gram bahan kering tercerna. Tingkat produksi metana pada perlakuan ini menggambarkan terjadinya peningkatan efisiensi energi pakan sebesar 14.16% dibandingkan dengan kontrol. Potensi penurunan produksi metana secara numerik juga ditunjukkan oleh perlakuan suplementasi kombinasi tepung batang pisang, Asetoanaerobium noterae dan Saccharomyces cerevisiae, yaitu sebesar 40.65 ml/gram pada inkubasi 12 jam, 64.67 ml/gram pada inkubasi 24 jam dan 84.75 ml/gram pada inkubasi 48 jam. Perlakuan ini menyebabkan terjadinya efisiensi energi pakan sebesar 19.93% pada inkubasi 12 jam, 14.05% pada inkubasi 24 jam dan 11.75% pada inkubasi 48 jam.
15
cerevisiae. Perlakuan ini meningkatkan kecernaan bahan kering pakan sebesar 14.58%, menurunkan emisi metana sebesar 7.71% dan meningkatkan efisiensi energi pakan sebesar 11.75% dibandingkan dengan kontrol.
Tabel 6. Produksi total gas dan metana setiap gram bahan kering tercerna dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang pada waktu inkubasi 12, 24 dan 48 jam
Perlakuan Gas/BKT (ml/g) CH4/BKT (ml/g)
12 jam 24 jam 48 jam 12 jam 24 jam 48 jam
A0B0 186.20 294.50 353.48 50.77 75.24 96.03b
A1B0 216.67 341.08 415.72 53.59 81.90 108.28c
A2B0 192.60 300.33 361.43 42.70 66.87 87.95ab
A3B0 208.72 329.42 391.70 47.88 73.66 95.63b
A0B1 166.40 257.27 315.21 42.58 63.51 82.43a
A1B1 177.86 266.04 322.20 45.74 67.15 86.55ab
A2B1 208.35 307.41 368.09 45.23 68.93 90.32ab
A3B1 174.80 279.19 338.93 40.65 64.67 84.75ab
Faktor A0 176.30 275.88 334.34 46.68 69.38 89.23
Faktor A1 197.26 303.56 368.96 49.67 74.52 97.41
Faktor A2 200.85 304.04 364.92 44.03 67.95 89.19
Faktor A3 191.02 303.21 364.17 44.11 68.97 89.95
Faktor B0 201.25 316.75b 381.19b 49.03b 74.77b 97.40
Faktor B1 181.29 276.84a 335.43a 43.51a 66.00a 85.92
SEM 5.38 6.42 7.52 1.09 1.30 1.77
P-value :
Faktor A 0.345 0.188 0.157 0.150 0.167 0.096
Faktor B 0.066 0.001 0.001 0.007 0.000 0.000
Interaksi 0.191 0.081 0.051 0.150 0.067 0.039
Superskrip yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P<0,05); P-value (tingkat signifikansi); SEM (Standard Error Mean); A0 (tanpa probiotik); A1 (S. cerevisiae); A2 (A. noterae); A3 (S. cerevisiae dan A. noterae ); B0 (tanpa tepung batang pisang); B1 (tepung batang pisang); BKT (Bahan Kering Tercerna).
Kondisi Fermentabilitas Rumen dan Populasi Mikroorganisme
Produk akhir dari pencernaan fermentatif di dalam rumen menghasilkan asam lemak terbang (VFA), NH3 dan gas yang terdiri dari CH4, CO2, serta gas
16
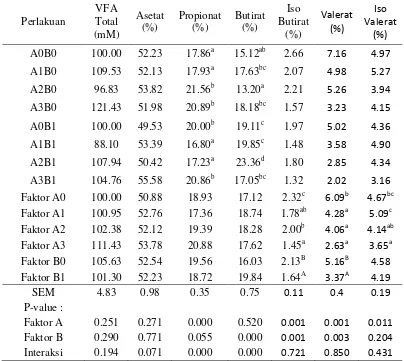
berupa iso butirat dan iso valerat. Kandungan Volatile Fatty Acid (VFA) dan komposisi penyusunnya tersaji pada Tabel 7.
Tabel 7. Produksi Volatile Fatty Acid dari substrat hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang
Perlakuan
A0B0 100.00 52.23 17.86a 15.12ab 2.66 7.16 4.97
A1B0 109.53 52.13 17.93a 17.63bc 2.07 4.98 5.27
A2B0 96.83 53.82 21.56b 13.20a 2.21 5.26 3.94
A3B0 121.43 51.98 20.89b 18.18bc 1.57 3.23 4.15
A0B1 100.00 49.53 20.00b 19.11c 1.97 5.02 4.36
A1B1 88.10 53.39 16.80a 19.85c 1.48 3.58 4.90
A2B1 107.94 50.42 17.23a 23.36d 1.80 2.85 4.34
A3B1 104.76 55.58 20.86b 17.05bc 1.32 2.02 3.16
Faktor A0 100.00 50.88 18.93 17.12 2.32c 6.09b 4.67bc
Faktor A1 100.95 52.76 17.36 18.74 1.78ab 4.28a 5.09c
Faktor A2 102.38 52.12 19.39 18.28 2.00b 4.06a 4.14ab
Faktor A3 111.43 53.78 20.88 17.62 1.45a 2.63a 3.65a
Faktor B0 105.63 52.54 19.56 16.03 2.13B 5.16B 4.58
Faktor B1 101.30 52.23 18.72 19.84 1.64A 3.37A 4.19
SEM 4.83 0.98 0.35 0.75 0.11 0.4 0.19
P-value :
Faktor A 0.251 0.271 0.000 0.520 0.001 0.001 0.011
Faktor B 0.290 0.771 0.055 0.000 0.001 0.003 0.204
Interaksi 0.194 0.071 0.000 0.000 0.721 0.850 0.431
Superskrip huruf kecil atau kapital yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P<0,05); P-value (tingkat signifikansi); SEM (Standard Error Mean); A0 (tanpa probiotik); A1 (S. cerevisiae); A2 (A. noterae); A3 (S. cerevisiae dan A. noterae ); B0 (tanpa tepung batang pisang); B1 (tepung batang pisang).
Komposisi asetat, butirat dan propionat berinteraksi satu sama lain dan menghasilkan perbedaan VFA total yang tidak signifikan. Kandungan VFA total dari setiap perlakuan berkisar antara 88.10 s/d 121.43 mM. Interaksi antar komponen ini juga memberikan efek terhadap tingkat emisi metana dan efisiensi energi pakan. Pembentukan propionat di dalam rumen membutuhkan hydrogen (H2), sedangkan pembentukan asetat dan butirat menghasilkan H2 (Martin et al.
2008). Hidrogen yang diproduksi di dalam rumen dimanfaatkan oleh mikroba methanogen dalam mereduksi CO2 untuk membentuk CH4. Pembentukan asetat
dan butirat meningkatkan akumulasi H2 di dalam rumen serta meningkatkan emisi
17
Perlakuan yang diberikan tidak memberikan pengaruh yang nyata terhadap komposisi asetat, namun berpengaruh nyata (P<0.05) terhadap komposisi propionat, butirat, iso butirat, valerat dan iso valerat. Secara umum, faktor A meningkatkan komposisi propionat dan faktor B meningkatkan komposisi butirat. Kedua faktor saling berinteraksi untuk meningkatkan propionat dan butirat. Peningkatan komposisi propionat terbaik ditunjukkan oleh suplementasi Asetoanaerobium noterae. Data ini memberikan gambaran bahwa selain memanfaatkan H2, Asetoanaerobium noterae juga berkontribusi dalam
mengarahkan jalur fermentasi ke arah pembentukan propionat. Asam piruvat yang dihasilkan dari fermentasi karbohidrat substrat diubah menjadi laktat untuk menghasilkan propionat.
Peningkatan komposisi propionat dan butirat diduga terjadi karena berkembangnya bakteri Prevotella ruminicola, Treponema bryantii dan Butyrivibrio fibrisolvens. akibat berkurangnya populasi protozoa (Suharti 2010). Suplementasi Asetoanaerobium noterae memberikan pengaruh yang nyata terhadap penurunan populasi protozoa (Tabel 8). Bakteri Butyrivibrio fibrisolvens adalah bakteri penghasil butirat, bakteri Prevotella ruminicola merupakan bakteri penghasil propionat melalui jalur akrilat, sedangkan Treponema bryantii termasuk bakteri yang aktif mendegradasi turunan xylan dan pektin menjadi suksinat yang merupakan prekursor propionat (Hobson dan Stewart 1997).
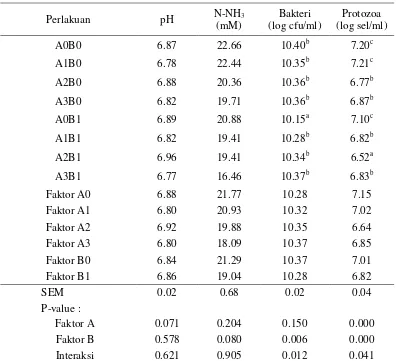
VFA (Volatile Fatty Acid) berfungsi sebagai sumber energi bagi ternak ruminansia dan merupakan sumber kerangka karbon bagi pembentukan protein mikroba (Damron 2006). Amonia, asam amino bebas, dan oligopeptida digunakan oleh bakteri untuk membentuk protein tubuhnya (McDonald et. al. 2002). Konsentrasi optimum amonia di dalam rumen berkisar antara 6-21 mM (McDonald et al. 2002). Konsentrasi VFA dan amonia harus dalam keadaan cukup untuk menunjang sintesis protein mikroba. Amonia yang diperoleh pada penelitian ini relatif sama, yaitu berkisar antara 16.46 s/d 22.66 mM. Kondisi pH rumen juga relatif sama dan berada dalam kondisi nyaman bagi mikroorganisme rumen, yaitu 6.77 s/d 6.96. Oleh sebab itu, perbedaan populasi mikroba rumen diduga terjadi karena mekanisme lain yang tidak berkaitan dengan konsentrasi VFA, amonia dan pH rumen.
Rataan populasi bakteri dan protozoa disajikan pada Tabel 8. Suplementasi probiotik (faktor A) tidak memberikan pengaruh yang nyata (P>0.05) terhadap populasi bakteri, sedangkan suplementasi tepung batang pisang (faktor B) terlihat memberikan pengaruh yang signifikan (P<0.05). Suplementasi tepung batang pisang secara tunggal menurunkan populasi bakteri dari 10.40 log cfu/ml (kontrol) menjadi 10.15 log cfu/ml. Penurunan populasi bakteri diduga terjadi karena aktivitas senyawa fitogenik yang terkandung dalam batang pisang, terutama tanin. Tanin berikatan dengan membran sel bakteri rumen, menghambat pertumbuhan dan aktivitas enzimatisnya (Smith et al. 2005).
18
menurunkan populasi protozoa (P<0.05) dari 7.20 log sel/ml menjadi 6.77 log sel/ml. Tingkat penurunan populasi protozoa menjadi lebih tinggi jika suplementasi Asetoanaerobium noterae dikombinasikan dengan tepung batang pisang, yaitu menjadi 6.52 log sel/ml. Hal ini menggambarkan bahwa Saccharomyces cerevisiae dan Asetoanaerobium noterae memiliki interaksi yang positif dengan tepung batang pisang dalam menurunkan populasi protozoa. Tabel 8. Konsentrasi pH, N-NH3, populasi bakteri dan protozoa dari substrat
hasil samping kelapa sawit yang ditambahkan probiotik dan tepung batang pisang
Superskrip yang berbeda pada kolom yang sama menunjukkan perbedaan nyata (P<0,05); P-value (tingkat signifikansi); SEM (Standard Error Mean); A0 (tanpa probiotik); A1 (S. cerevisiae); A2 (A. noterae); A3 (S. cerevisiae dan A. noterae ); B0 (tanpa tepung batang pisang); B1 (tepung batang pisang).
19
pakan hasil samping kelapa sawit belum mampu mengurangi populasi protozoa secara signifikan. Tidak berbedanya populasi protozoa pada perlakuan suplementasi tepung batang pisang diduga karena jumlah pemberiannya yang masih kurang untuk pakan yang terdiri dari hasil samping kelapa sawit. Kemampuannya baru terlihat dengan mengkombinasikannya dengan Saccharomyces cerevisiae. Enzim yang dihasilkan oleh Saccharomyces cerevisiae diduga mempercepat dan menjadi katalisator reaksi saponin tepung batang pisang dengan sterol protozoa, sehingga kemampuan saponin tepung batang pisang mendefaunasi protozoa menjadi meningkat.
Mekanisme defaunasi protozoa oleh Asetoanaerobium noterae belum banyak dipahami. Salah satu pendekatan yang bisa digunakan adalah karena media tumbuh Asetoanaerobium noterae. Salah satu media tumbuh pembuatan probiotik Asetoanaerobium noterae adalah CuSO4. Jouany (1991) menyatakan
bahwa CuSO4 memiliki kemampuan sebagai defaunator protozoa. Pendekatan lain
adalah karena aktivitas fisik Asetoanaerobium noterae terhadap protozoa. Protozoa menjadi media tumbuh mikroba metanogen di dalam rumen. Hal ini diduga karena permukaan tubuh protozoa memberikan kondisi yang nyaman bagi mikroba metanogen baik lingkungan maupun penyediaan makanan. Mikroba asetogenik relatif memiliki kebutuhan yang sama dengan mikroba metanogenik, sehingga sebagian Asetoanaerobium noterae juga menjadikan protozoa sebagai media tumbuh di dalam rumen. Asetoanaerobium noterae diduga memiliki sifat yang berbeda dengan mikroba metanogen yang bersimbiosis mutualisme dengan protozoa. Aktifitas Asetoanaerobium noterae secara fisik diduga memberikan efek yang negatif terhadap protozoa. Mekanisme defaunasi Asetoanaerobium noterae terhadap protozoa merupakan topik yang menarik untuk diteliti lebih lanjut.
KESIMPULAN
20
DAFTAR PUSTAKA
Ando S, Khan RI, Takahasi J, Gamo Y, Morikawa R, Nishiguchi Y, Hayasaka K. 2004. Manipulation of rumen fermentation by yeast: The effect of dried beer yeast on the in vitro degradability of forages and methane production. Asian-Aust J Anim Sci. 17:68-72.
[AOAC] Association of Official Analytical Chemists. 1997. Official Methods of Analysis. Virginia (US): Arlington.
Campbell JR, Douglas M, Kenealy. 2003. Animal Sciences 4th Edition. New York (US): McGraw-Hill.
Chaucheryras F, Fonty G, Bertin G, Gouet P. 1995. In vitro utilization by a ruminal acetogenic bacterium cultivated alone or in association with an Archea metanogen is stimulated by a probiotic strain of Saccharomyces cerevisiae. Appl Environ Microbiol. 61:3466-3467.
Chevaux E, Fabre MM. 2007. Probiotic yeast in small ruminants. Feed Mix. 15(1):28029.
Conway EJ. 1950. Microdiffusion Analysis and Volumetric Error. London (GB): Crosby Lokswood and Sons Ltd.
Damron WS. 2006. Introduction to Animal Science. Ohio (JP): Prentice Hall. Dawson KA, Newman KE, Boling JA. 1990. Effect of microbial supplements
containing yeast and Lactobacili on roughage-fed ruminal microbial activities. J Anim Sci. 68:3392-3398.
De Boever JL, Aerts JM, Vanacker JM, De-Brabander DL. 2005. Evaluation of the nutritive value of maize silages using a gas production technique. Anim Feed Sci Technol. 123:255-265.
Diwyanto K, Sitompul DM, Manti I, Mathius IW, Soentoro. 2003. Pengkajian pengembangan usaha sistem integrasi kelapa sawit-sapi. Prosiding Lokakarya Nasional Sistem Integrasi Kelapa Sawit-Sapi; Bengkulu, Indonesia. Bogor (ID): Puslitbang Peternakan. hlm11-22.
Erwanto, 1995. Optimalisasi system fermentasi rumen melalui suplementasi sulfur, defaunasi, reduksi emisi metan dan stimulasi pertumuhan mikroba pada ternak ruminansia [Disertasi]. Bogor (ID): Institut Pertanian Bogor. Fievez VA, Babayemi OJ, Demeyer D. 2005. Estimation of direct and indirect gas
production in syringes: A tool to estimate short chain fatty acid production that requires minimal laboratory facilities. Anim Feed Sci Technol. 123-124:197-210.
Fonty G, Joblin K, Chavarot M, Roux R, Naylor G, Michallon F. 2007. Establishment and development of ruminal hydrogenotrophs in metanogen-free lambs. Appl Environ Microbiol. 73:6391-6403.
Getachew G, Blummel M, Makkar HPS, Becker K. 1998. In vitro gas measuring techniques for assessment of nutritional quality of feeds: a review. Anim Feed Sci Technol. 72:261-281.
21
Hart KJ, Yanez-Ruiz DR, Duval SM, McEwan NR, Newbold CJ. 2008. Plant extracts to manipulate rumen fermentation. Anim Feed Sci Technol. 147:8-35.
Haryanto B, Thalib A. 2009. Emisi metana dari fermentasi enterik: kontribusinya secara nasional dan faktor-faktor yang mempengaruhinya pada ternak. Wartazoa. 19(4):157-165.
Hobson PN, Stewart CS. 1997. The Rumen Microbial Ecosystem. Second Ed. Blackie Academic & Professional, London.
Hiai S, Oura H, Nakajima T. 1976. Color Reaction of somesapogenins and saponins with vanillin and sulfuric acid. Planta Med. 29:116-122.
Jayanegara A, Sofyan A, Makkar HPS, Becker K. 2009. Kinetika produksi gas, kecernaan bahan organik dan produksi gas metana in vitro pada hay dan jerami yang disuplementasi hijauan mengandung tanin. Med Pet. 32:120-129.
Jayanegara A, Wina E, Soliva CR, Marquardt S, Kreuzer M, Leiber F. 2011. Dependence of forage quality and metanogenic potential of tropical plants on their phenolic fractions as determined by principal component analysis. Anim Feed Sci Technol. 163: 231-243.
Jouany JP. 1991. Defaunation of the rumen. Di dalam: Jouany JP, editor. Rumen Microbial Metabolism and Ruminant Digestion. Paris (FR): INRA. hlm 239-261.
Jouany JP. 2001. 20 years of research and now more relevant than ever- the coming of age of yeast cultures in ruminant diets. Di dalam: Jouany JP, editor. Responding to a Changing Agricultural Landscape. Paris (FR):
Alltech’s European. hlm 44-69.
Kementerian Pertanian. 2013. Statistik Pertanian 2013. Sutiyorini S, Waryanto B, editor. Jakarta (ID): Pusat Data dan Sistem Informasi Pertanian Kementerian Pertanian.
Kreuzer M, Soliva CR. 2008. Nutrition: key to methane mitigation in ruminants. Proc Soc Nutr Physiol. 17: 168-171.
Lopez S, Mcintosh FM, Wallace RJ, Newbold CJ. 1999. Effect of adding acetogenic bacteria on methane production by mixed rumen microorganisms. Anim Feed Sci Technol. 78:1-9.
Makkar HPS, Becker K. 1998. Do tannins in leaves of trees and shrubs from Africa and Himalayan regions differ in level and activity? Agroforestry Syst. 40: 59-68.
Makkar HPS. 2003. Quantification of Tannins in Tree and Shrub Foliage a Laboratory Manual. Dordrecht (NL): Kluwer Academic Publishers. Makkar HPS, Francis G, Becker K. 2007. Bioactivity of phytochemicals in some
lesser-known plants and their effects and potential applications in livestock and aquaculture production systems. Animal. 1:1371-1391.
Martin C, Doreau M, Morgavi DP. 2008. Methane mitigation in ruminants: from rumen microbes to the animal. Livestock and Global climate Change Conference; Hammamet. Hammamet (TN).
22
Mathius IW, Azmi, Manurung BP, Sitompul DM, Pryatomo E. 2004. Integrasi sawit-sapi : Imbangan pemanfaatan produk samping sebagai bahan dasar pakan. Prosiding Sistem Integrasi Tanaman-Ternak; 2004 Juli; Denpasar, Indonesia. Bogor (ID): Puslitbang Peternakan. hlm: 439-446.
McAllister TA, Newbold CJ. 2008. Redirecting rumen fermentation to reduce methanogenesis. Aust J Exp Agric. 48:7-13.
McDonald P, Edward RA, Greenhalgh JFD, Morgan CA. 2002. Animal Nutrition. New York (US): Scientific and Tech John Willey & Sons. Inc.
Miller TL, Wolin MJ. 2001. Inhibition of growth of methane-producing bacteria of the ruminant forestomach by hydroxymethylglutaryl-SCoA reductase inhibitors. J Dairy Sci. 84:1445-1448.
Morvan B, Bonnemoy F, Fonty G, Gouet P. 1996. Quantitative determination of H2-utilizing acetogenic and sulfate-reducing bacteria and metanogenic archaea from digestive tract of different mammals. Curr Microbiol. 32:129-133.
O’connor MH, Martin SA, Hill GM. 2002. Effects of Saccharomyces cerevisiae on in vitro mixed ruminal microorganism fermentation. Prof Anim Sci. 18:358-362.
Oeztuerk H, Schroeder B, Beyerbach M, Breves G. 2005. Influence of living and autoclaved yeasts of Saccharomyces boulardii on in vitro ruminal microbial metabolism. J Dairy Sci. 88:2594-2600.
Ogimoto K, Imai S. 1980. Atlas of Rumen Microbiology. Tokyo (JP): Jap. Sci. Soc. Press.
Puastuti W. 2008. Optimasi penggunaan produk samping kelapa sawit sebagai pakan ruminansia. Prosiding Lokakarya Nasional Optimalisasi Hasil Samping Perkebunan Kelapa Sawit dan Industri Olahannya sebagai Pakan Ternak; Bogor, Indonesia. Bogor (ID) Puslitbang Peternakan. hlm 143-151.
Reed G, Nagodawithana TW. 1991. Yeast Technology. Newyork (US): Rein Hold. Sitompul D. 2003. Desain pengembangan kebun dengan sistem usaha terpadu ternak sapi Bali. Prosiding Sistem Integrasi Tanaman-Ternak; Denpasar, Indonesia. Bogor (ID) Puslitbang Peternakan. hlm. 11-22.
Smith AH, Zoetendal E, Mackie RI. 2005. Bacterial mechanisms to overcome inhibitory effects of dietary tannins. Microb Ecol. 50:197-205.
Suharti S. 2010. Modifikasi keragaman mikroba dan fermentasi rumen sapi dengan pemberian saponin lerak (Sapindus rarak) [Disertasi]. Bogor (ID): Institut Pertanian Bogor.