DAEMONOROPS FISSA
COMPLEX IN WEST MALESIA
DWI PUTRI RAMADHANI
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
STATEMENT OF RESEARCH ORIGINALITY AND
INFORMATION SOURCE
This is to verify that my thesis entitled: Daemonorops fissa complex in West Malesia is my own work and never been submitted to any institution before. All the incorporated data and information are valid and stated clearly in the text, and listed in the references.
Bogor, May 2015
Dwi Putri Ramadhani
SUMMARY
DWI PUTRI RAMADHANI. Daemonorops fissa complex in West Malesia. Supervised by TATIK CHIKMAWATI and HIMMAH RUSTIAMI.
Daemonorops fissa complex is highly varied in West Malesia, but information about this complex is limited. The objective of the research was to reveal species diversity of Daemonorops fissa complex in West Malesia based on morphological and anatomical characters.
Evaluation of morphological characters was conducted in Herbarium Bogoriense (BO) Cibinong Science Centre LIPI, Bogor (West Java). A total of 355 herbarium specimens were observed in this study representing all species of
Daemonorops fissa complex. This study explored Daemonorops fissa complex, including the following: 1) species diversity, (2) distribution, (3) morphological variation, (4) anatomical variation and (5) phenetic relationships. Data obtained then was analized descriptively and phenetically.
Species of Daemonorops fissa complex in West Malesia share some unique characters such as erect peduncle (to ± 2 cm), alternate leaves, length of involucrum 1 cm, globose fruits, and ruminate endosperm. This complex species consists of eight species namely, D. angustifolia, D. calicarpa, D. fissa, D. grandis, D. lewisiana, D. melanochaetes, D. monticola, D. stenophylla. Daemonorops binnendijkii, D.palembanica and D. trichroa proposed as new synonymies. Daemonorops binnendijkii and D. trichroa are new synonymsto D. angustifolia, and D. palembanica is a new synonym to D. melanochaetes. All species found in lowland rain forest, especially along stream and other wet places or more open areas, swampy area, ridgetop hill Dipterocarp forest, bamboo forest, and river side on altitude from 250 m – 1900 m asl.
Morphological variation of Daemonorops fissa complex in West Malesia could be differentiated into 20 morphological characters. Cluster analysis separated all species into two main groups with coefficient similarity of 36%. Group I consisted of D. angustifolia, D. fissa, D. grandis, D. melanochaetes, D. monticola and D. stenophylla, while group II consisted of D. calicarpa and D. lewisiana. They were separated from five characters, total height, number of leaflets, width of leaflets, type of inflorescence and type of spine in leafsheath.
Leaf anatomical characters of paradermal section could not be used to separate among species within D. fissa complex, but leaf anatomical characters of transverse section could be used to identify among species. All examined species of D. fissa complex have three similar characters, tetracytyc stomata, cryptopor type with sunken position and cell wall of sinous, but they have different pattern of bulliform cell and palisade tissues. Both characters are considered as two valuable new characters for supporting morphological data for species delimitation.
RINGKASAN
DWI PUTRI RAMADHANI. Daemonorops fissa kompleks di Kawasan Malesia Barat. Dibimbing oleh TATIK CHIKMAWATI dan HIMMAH RUSTIAMI.
Daemonorops fissa kompleks memiliki keanekaragaman yang tinggi di Malesia Barat, tetapi informasi tentang kompleks ini sangat terbatas. Penelitian ini mempelajari keanekaragaman jenis Daemonorops fissa kompleks di kawasan Malesia Barat berdasarkan karakter morfologi dan anatomi.
Evaluasi karakter morfologi dilakukan di Herbarium Bogoriense, Pusat Penelitian Biologi, LIPI dengan mengamati 355 lembar spesimen herbarium yang mewakili semua jenis Daemonorops fissa kompleks. Kajian yang dilakukan meliputi 1) keanekaragaman jenis, (2) persebaran, (3) variasi morfologi, (4) variasi anatomi dan (5) kekerabatan fenetik. Analisis data meliputi analisis deskriptif dan analisis fenetik.
Jenis-jenis dari Daemonorops fissa kompleks di kawasan Malesia Barat memiliki karakter bersama yaitu tanpa tangkai bunga atau memiliki tangkai bunga sampai ± 2 cm, susunan anak daun berseling, panjang daun pembalut 1 cm, bentuk buah bulat dan bentuk endosperma termamah. Daemonorops fissa
kompleks di kawasan Malesia Barat terdiri dari delapan jenis yaitu D. angustifolia, D. calicarpa, D. fissa, D. grandis, D. lewisiana, D. melanochaetes, D. monticola, D. stenophylla. Daemonorops binnendijkii, D.palembanica dan D. trichroa diusulkan sebagai sinonim. Daemonorops binnendijkii dan D. trichroa
adalah sinonim dari D. angustifolia, sedangkan D. palembanica adalah sinonim dari D. melanochaetes. Semua jenis ditemukan di berbagai habitat seperti hutan hujan tropis, tempat terbuka, hutan Dipterocarpaceae, hutan primer, hutan bambu dan sepanjang aliran sungai dengan ketinggian sekitar 250 m sampai 1900 mdpl.
Variasi morfologi Daemonorops fissa kompleks dapat dibedakan ke dalam 20 karakter yang digunakan untuk menyusun deskripsi dan kunci identifikasi. Analisis gugus memisahkan semua jenis ke dalam dua kelompok utama pada koefisien keserupaan 36 %. Kelompok I terdiri dari D. angustifolia, D. fissa, D. grandis, D. melanochaetes, D. monticola dan D. stenophylla. Kelompok II terdiri dari D. calicarpa dan D. lewisiana.Kedua kelompok ini memisahkan berdasarkan karakter tinggi total, jumlah anak daun, lebar anak daun, tipe perbungaan dan bentuk duri pada batang.
Karakter anatomi daun D. fissa kompleks dari irisan paradermal tidak dapat digunakan sebagai pembeda jenis dalam D. fissa kompleks, tetapi irisan melintang dapat digunakan sebagai pembeda jenis. Semua jenis D. fissa kompleks yang diamati memiliki tiga karakter yang sama, bentuk stomata tetrasitik, stomata tenggelam dan tepi sel epidermis bergelombang, tetapi memiliki bentuk sel kipas dan jaringan palisade yang berbeda. Kedua karakter tersebut dijadikan sebagai karakter baru untuk menguatkan karakter morfologi sebagai batasan jenis D. fissa
kompleks.
Copyright@ 2015, Bogor Agricultural University
All Right Reserved
It is prohibited to cite all or part of this thesis without referring to and mentioning the source. Citation only permitted for the sake of education, research, scientific writing, report writing, critical writing or reviewing scientific problems. Citation does not inflict the name and honour of Bogor Agricultural University.
A thesis submitted to fulfill one of the requirements for the Master Degree in Plant Biology Graduate Program
DAEMONOROPS FISSA
COMPLEX IN WEST MALESIA
THE GRADUATE SCHOOL
BOGOR AGRICULTURAL UNIVERSITY BOGOR
2015
Examiner of the examination: Dr. Rugayah, M.Sc.
Herbarium Bogoriense, Botany Division,
Thesis Title : Daemonorops fissa Complex in West Malesia Name : Dwi Putri Ramadhani
NIM : G353120231
Certified by Supervisor Commitee
Dr Ir Tatik Chikmawati, MSi Chairman
Dr Himmah Rustiami, SP, MSc Member
Approved by
The Head of Plant Biology Graduate Program
Dr Ir Miftahudin, MSi
The Dean of Postgraduate School
Dr Ir Dahrul Syah, MScAgr
PREFACE
I am grateful to my supervisors, Dr. Tatik Chikmawati M.Si., and Dr. Himmah Rustiami, SP, M.Sc from Herbarium Bogoriense (BO) LIPI for their advice, kindness, patience and valuable discussion throughout this study.
Many people have helped me through good advice, suggestions and supports. In particular, I would like to thank Mahya, Gilang, Zulfan and Aini Qomariah for their help during field, Dian and Hariri for their help in GIS Mapping for this study. I would like to thank Dr. Sunaryo, Drs. Erlin Rachman, Eka Fatmawati T, S.Si., Ujang Hapid, Widoyanti and Darius for their help and technical supports during my anatomical study.
I would also like to thank all people in Herbarium Bogoriense for the information and facilities when I carried out specimen examinations. I sincerely thank all my friends of Plant Biology Graduate Program for their friendship.
I express deepest appreciation to Indonesian Government through DIKTI for providing financial support for my research. Finally, my deepest appreciation is also to my family, Erlis Suriani (mother) and Djulias Muhar (father), Sitha Muriani (my sister) and Doni Tri Hariansyah (my brother) who has given me love and moral supports. Undoubtedly, without them it is going to be impossible for me to finally finish this study.
Bogor, May 2015
CONTENTS
LIST OF TABLES vi
LIST OF FIGURES vi
LIST OF APPENDIXES vii
1 INTRODUCTION 1
2 MATERIALS ANDMETHODS 3
Time and Place 3
Plant Materials 3
Research Methods 3
Data Analysis 6
3 RESULT AND DISCUSSION 6
Species Diversity 6
Distribution 7
Morphological Variations 8
Anatomical Variations Anatomical Key to Daemonorops fissa complex in West Malesia 14
16
Phenetic Analysis 17
Taxonomic Treatment 19
Morphological Key to Daemonorops fissa complex in West Malesia 19
Species Description of Daemonorops fissa complex in West Malesia 20
4 CONCLUSION 35
REFERENCES 35
LIST OF TABEL
1 Part of rattan. [A] Vegetative parts; [B] Generative parts. 1. cirrus, 2. flagellum, 3. stem bearing a crownshaft, 4. stem bearing open sheaths, 5. knee, 6. ocrea, 7. rachilla, 8. fruit
4
2 Distribution of Daemonorops fissa complex in West Malesia
D.angustifolia ( ), D.grandis ( ), D.calicarpa ( ),D.fissa (
D. lewisiana ( ), D. melanochaetes ( ),D. monticola ( ),
D. stenophylla ( )
7
3 Spine types on the leaf sheath of D. fissa complex [A] oblique circle; [B] scattered
8 4 Type of knee Daemonorops fissa complex [A] bulgy; [B]
elongate-folded; [C] elongate-flat; [D] elongate inflated.
9 5 Density of spine in prophyll of Daemonorops fissa complex
[A] velutinose with triangular spine; [B] velutinose with net-like spine; [C] setose; [D] net-like spine; [E] triangular spine.
gynoecium; [9] corolla; [10] calyx; [11] empty anther.
11
7 Fruit scale surface of Daemonorops fissa complex [A] concave; [B] flat two vascular bundle; [B] near midvein; [C] midvein.
15 13 Type of bulliform cell Daemonorops fissa complex [A] letter V; [B]
curved upward; [C] parallel; [D] curved downward.
14 Palisade tissue Daemonorops fissa complex [A] with one layer of palisade tissue (dorsiventral); [B] with two layer of palisade tissue (isobilateral). [1] upper palisade; [2] under palisade.
16
15 Dendogram of Daemonorops fissa complex in West Malesia based on morphological characters
17 16 Daemonorops angustifolia (Griff.) Mart. [A] herbarium type
collection SING; [B] female and sterile male flower; [C] fruit; [D] endosperm; [E] knee; [F] latex; [G] cane.
22
17 Daemonorops calicarpa (Griff.) Mart. [A] herbarium type collection K; [B] spine oblique circle in leaf sheaths; [C] flower; [D] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
23
18 Daemonorops fissa Blume [A] herbarium collection BO; [B] knee; [C] cirrus; [D] fruit; [E] flower; [F] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
25
19 Daemonorops grandis (Griff.) Mart.[A] herbarium type collection SING; [B] knee; [C] male flower; [D] endosperm. [1] corolla; [2] stamen; [3] calyx.
27
20 Daemonorops lewisiana (Griff.) Mart. [A] herbarium type collection K; [B] spine in leaf sheaths; [C] female and male flower; [D] seed; [E] endosperm; [F] transversal veinlets.
28
21 Daemonorops melanochaetes Blume. [A] herbarium type collection K; [B] knee; [C] prophyll; [D] flower; [E] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
31
22 Daemonorops monticola (Grift) Mart., [A] herbarium type collection K; [B] prophyll; [C] spine in leaf sheath; [D] male flower; [E] flower; [F] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
33
23 Daemonorops stenophylla Becc. [A] herbarium type collection K; [B] knee; [C] prophyll; [D] fruit.
1 INTRODUCTION
Daemonorops is a rattan genus included in the subtribe Calamoideae. It is the second largest rattan genus after Calamus, which was consisted of about 120 species (Rustiami 2011). The genus Daemonorops was first described by Blume (1849), based on the type specimen Daemonorops melanochaetes. Daemonorops
was distributed from India and South China through the Malay Archipelago to New Guinea where was represented by one species. Twenty-two species occur in Southern Asia (Beccari 1911; Dransfield 1979, 2001; Evan et al. 2001; Hodel 1998). Daemonorops species are highly variable, but they are homoplasious in many morphological characters (Umapathy et al. 2014).
The greatest morphological variation of Daemonorops was found in Malay Peninsula, Sumatra and Borneo (Dransfield et al. 2008). West Malesia consisted of Borneo, Sumatra, Malay Peninsula and Java (Raes & van Welzen 2009). High diversity in West Malesia is supported by geological conditions and climate history of the complex as well as the fact that this region is an archipelago with a variety of obstacles. This condition opens up opportunities for the development of its type in the isolated region. Rising sea levels during the Pleistocene as well as the isolated islands causes the distribution patterns of plants including rattan is unique in the West Malesia (Rustiami 2009).
The species of Daemonorops are mostly confined to primary tropical rain forest on a great various soils, but some species had narrow ecological requirements. A few species are a rather more weedy nature, found abundant in forest habitats with high-light intensities such as riverbanks. One species in Borneo, Daemonorops longispatha, grows on the landward margin of mangrove forest. Some species are strictly montane, occurring at altitude up to ca. 2500 m above sea level (m asl). Several species in Borneo are confined to heath forest, or to limestone or serpentine rock (Dransfield et al. 2008).
Daemonorops spesies have been recognized having many uses. structure, section Daemonorops and Piptospatha (Beccari 1911; Rustiami et al.
2014). Species within section Daemonorops have concave boat-shaped bracts, which at anthesis are completely enclosed by the prophyll and split longitudinally to expose their flowers. All bracts are usually persistent. In contrast to section
2
Daemonorops fissa complex was classified into the section Daemonorops
with the sheath upright and directly attaching to the stem by peduncle length of ± 2 cm (pers. com 2013). Based on observed specimen in herbarium and compared to the previous studies, species concept of Daemonorops fissa complex remains unclear. Daemonorops fissa complex in West Malesia consists of 9 species namely D. angustifolia, D. binnendijkii, D. fissa, D.grandis, D. melanochaetes, D. palembanica, D. sepal, D. stenophylla, and D. trichroa (Beccari 1911). The later study mentioned only 3 species in Malaya, namely D. angustifolia, D. grandis and
D. melanochaetes (Furtado 1953). However, the last study reported that 5 species were recognized, D. angustifolia, D. fissa, D. grandis, D. melanochaetes and D. sepal in Malay Peninsula, Sabah and Sarawak (Dransfield 1979; 1984, 1992).
Rattan classification was mainly based on discontinued of morphological characters, the basic data for separating among species (Rustiami 2009). However, morphological characters sometimes have disadvantage where placement of a taxon in the classification is doubtful (Tellu 2005), therefore another approach is needed, such as anatomy. Because of uncertain in distinguishing each species within D. fissa complex, this species complex in this region needs to be re-examined to obtain clear species delimitation, and to describe the relationships among species within this complex using phenetic approach, based on morphological and anatomical characters.
Little work has been done on the leaf anatomy of Daemonorops. Anatomy of Daemonorops can be distinguished from Calamus based on three characters: guard cells, the presence of longer sklereid cells and commissure, and large cells contained of tannins (Tomlinson 1961). Another study found that Daemonorops
spp. from Malaya can be distinguished from their sclereid characters (Tomlinson 2006).
3
2 MATERIALS AND METHODS
Time and Place
This study was conducted in August 2013 until October 2014 starting with field exploration in three locations in North Sumatra; Sibolangit, Mount Sinabung and Deleng Lancuk. Observation of herbarium specimens was done in the Herbarium Bogoriense (BO) Cibinong Science Centre LIPI, Bogor (West Java).
Plant Materials
Plant materials used to provide morphological data for the present study were herbarium specimens including preserved material deposited at Herbarium Bogoriense. All plant materials were represented the species of Daemonorops fissa complex such as D. angustifolia, D. binnendijkii, D. callicarpa, D. fissa, D. grandis, D. melanochaetes, D. monticola. D. palembanica, D. trichroa, and D. stenophylla. A total of 355 herbarium specimens were observed for morphological examination in this study.
Research Methods
Procedure for observation in this study followed the method described by Rifai (2012), and de Vogel (1987). The procedure included 14 steps:(1) Selection of taxon and determination of scope in the study and work to carried out; (2) Collecting the material as much as possible (3) Reviewing literatures and records of all the scientific names and data that included in the important taxa; (4) Selecting the specimens that have been collected in taxonomic units based on observable characteters; (5) Scrutinizing the available specimens based on morphological approach; (6) Testing the characters already used by previous researches, and performing cross check for justifying whether the existing correlation between various characters could be confirmed or not; (7) Delimiting the taxa in question according to the study results and resting the plausibility of newly discovered characters; (8) Searching and determining the relationships among taxa; (9) Solving the problem concerned to nomenclature; (10) Constructing a key for identification of acceptable taxa; (11) Labeling on each specimen examined according to the name of the concluded study; (12) Making description of each taxa and note for individual specimens analyzed such as ecology data and distribution; (13) Making draws of organ parts for publication as needed; (14) Compiling a scientific report to be published.
Field Sampling
4
Specimen collection followed standard procedures developed by Dransfield (1986). Data and information recorded from the field included: general data (location, habitat, height, use of species, and date of collection); habit (cluster/ solitary), stem (total height, with or without leaf sheath diameter, length and internodes, colour), leaves (number of leaflets, leaflets arrangement, the length and width of leaves), additional organ (cirrus); inflorescences (length, number, colour of rachilla), flower (colour, length and widths) ; fruit and seeds. The plants were then documented, collected and processed for herbarium specimens.
Preparation of herbarium specimens followed standard procedure by Dransfield (1986). The collection of complete rattan specimens has many representative characters including:
1- Vegetative parts: Leaf sheath has a knee, ocrea, spine and sometimes a flagellum (Figure 1A). Leaves of many species are long and large, collected only a portion of the leaf base, middle section and apex. If the leaf ends with cirrus, collected the leaf apex with cirrus. If the leaflet is large which does not fit to the folder, cutting the leaflets off one side (leaving the leaflet bases in place) and the other side is folded to fit the folder.
Figure 1 Part of rattan. [A] Vegetative parts. [B] Generative parts. 1. cirrus, 2. flagellum, 3. stem bearing a crownshaft, 4. stem bearing open sheaths, 5. knee, 6. ocrea, 7. rachilla, 8. fruit
2. Generative parts: Many rattan species have large and long inflorescence which could not be collected as a whole specimen (Figure 1B). To collect such specimens, a part of the inflorescence base was taken, including primary axis, bract, and the whole partial inflorescence were collected, and then collected the last partial inflorescence to see the range sizes.
Anatomical Observation
The plant materials used in the present study were mostly dried and pressed specimens. Samples were taken from the middle leaves located in the middle and cut along 10cm2. Each species represents of one to two collection numbers.
A B
1
2
3 4
8
7
5 Anatomical observations were prepared from paradermal and transverse leaf sections. Leaf blade was boiled in HNO3 to manufacture paradermal section then
stained with 1% safranin (Sass 1951). Observed characters included shape and size epidermal cells and stomata. For preparing leaf transverse section, leaf were cut using freezing microtome Yamato RV-240 as thin as 20-25 µm. Leaf slices then were dripped with sodium hypochlorite to remove chlorophyll, then stained with safranin and then added few drops of glycerin on slide. Observed character included the number of palisade tissue, palisade thickness, the presence of tannin cells and the type, number, location and length of bulliform cells. After obtaining permanent slides, the slide were observed using Nikon Eclipse E100 and documented using OptiLab Viewer 2.2.
Morphological Observation
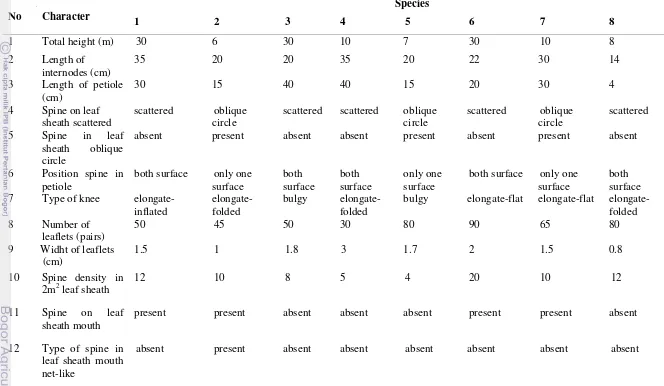
Twenty observed morphological characters were described in Table 1. Morphological character includes several characters from internodes, petiole, spine, knee, leaflets, inflorescence, fruit and endosperm. (Beccari 1911, Furtado 1953, Dransfield 1979; 1984, 1992, Rustiami 2011).
Table 1 Morphological characters of Daemonorops fissa complex in West Malesia No Characters State of character (score)
1 Total height (m) ≤ 5 tall (0), 6 –10 tall (1), ≥10 tall (2) 2 Length of internodes (cm) ≤20 (0), 21-30 (1), >30 (2)
3 Length of petiole (cm) 4 (0), ≤30 (1), > 30 (2) 4 Spine in leaf sheath scattered absent (0), present (1) 5 Spine in leaf sheath oblique
circles
absent (0), present (1)
6 Position spine in petiole only one surface (0), on both surface (1) 7 Type of knee elongate-folded (0), elongate-flat (1),
elongate inflated (2). bulgy (3)
14 Type of inflorescence pleonanthic (0), hapaxanthic (1) 15 Length of prophyll (cm) ≤ 30 (0), > 30 (1)
16 Surface of prophyll setose (0), velutinose (1) 17 Length of peduncle (cm) 1 cm (0), > 1 cm (1) 18 Spine of peduncle absent (0), present (1) 19 Surface of fruit flat (0), concave (1)
6
Data Analysis
A species cluster was constructed based on morphological characters observed for each species. Coefficient of similarity was analyzed using Simple Matching (SM). Clustering analysis was analyzed using unweighted pair-group method with arithmetic average (UPGMA) method (Rohlf 2002). Analysis was grandis, D. lewisiana, D. melanochaetes, D. monticola and D. stenophylla. Three species are proposed as new synonym, D. binnendijkii and D. trichroa are synonym to D. angustifolia, whereas D. palembanica as a synonym to D. melanochaetes.
Daemonorops binnendijkii was named based on two types, cultivated plant at Buitenzorg and fruits from the Utrecht Herbarium, but the collection number of the specimen types were unknown (Beccari 1911). On September, 25 in 1914, Grashoff collected a specimen named Daemonorops cf. binnendijkii in Blinjoe, Bangka Island. This spesimen deposits in Herbarium Bogoriense as a specimen type. Until now, D. binnendijkii was never collected by researches. Because of lesser data could be collected, D. binnendijkii is rated as Data Deficient (Bachman 2013). Based on morphological observation of specimen herbarium, D. binnendijkii is similar to D. angustifolia, and later study based on anatomical characters showed that anatomical characters of D. binnendijkii is same to D. angustifolia.
Fruit collection by Teysmann no. 3582 collected in Muara Dua, Palembang is the type of Daemonorops trichroa, and it deposits in Herbarium Bogoriense. The fruit is extremely similar to fruit of D. angustifolia from Malayan Peninsula (Beccari 1911). Based on characters armature of the petiole, leaf sheath, rachis and spathae, Furtado reduce D. trichroa Miq. as a synonym of
Daemonorops angustifolia (Furtado 1937).
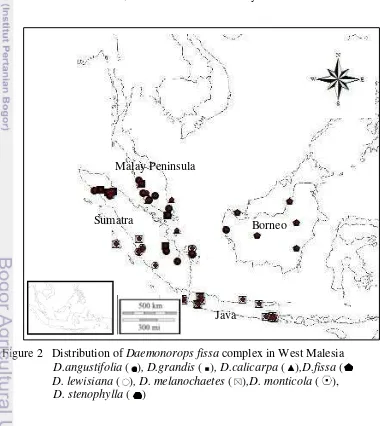
7 Distribution
The distribution of the Daemonorops fissa complex is given in Figure 2.
D. angustifolia, D. callicarpa, D. fissa, D. grandis, D. lewisiana, D. monticola
and D. melanochaetes are the most wide spread species, extending throughout in Malay Peninsula, Sumatra, Java and Borneo. D. stenophylla is known only from the forest of North Sumatra and west Sumatra. D. callicarpa in Sumatra found in lowland Dipterocarp forest and growed with Calamus castaneus. D. fissa is a new record from Sumatra. Dransfield (1979; 1984, 1992), has investigated this species widespread throughout in Borneo and endemic. D. melanochaetes is widespread in Java. Generally, the forest in Malay Peninsula and Sumatra are comparatively rich in Daemonorops fissa complex population, each of them having 6 species.
Daemonorops fissa complex grew in lowland rain forest especially along stream and other wet places or more open areas, swampy area, ridgetop hill Dipterocarp forest, bamboo forest and river side on altitude from 70 m – 1900 m sl. All species of Daemonorops fissa complex was found in lowland forest on altitude 250 – 1900 m asl, but D. monticola was only found above 1000 m asl.
Figure 2 Distribution of Daemonorops fissa complex in West Malesia
D.angustifolia ( ), D.grandis ( ), D.calicarpa ( ),D.fissa ( D. lewisiana ( ), D. melanochaetes ( ),D. monticola ( ),
D. stenophylla ( )
Malay Peninsula
Sumatra Borneo
8
Morphological Variation
Habit
All species of Daeomonorops fissa complex from West Malesia are climb clustering rattan. Three species, D. calicarpa, D. lewisiana and D. monticola are found climbing up to 5 m high or more. Two other species, D. grandis and D. stenophylla are found climbing to 10 m. Daemonorops angustifolia, D. binnendijkii, D. fissa and D. melanochaetes are the highest climbers, up to 20 m or higher.
Leaf sheath
Leaf sheath of Daemonorops fissa complex are varied in colour, spine position and type. The colour of leaf sheath are dull greenish brown, pale green to pale brownish, and brownish green. Spine position in leaf sheath are scattered or oblique circle (Figure 3). Species with spine arranged in oblique circle are D. calicarpa, D. lewisiana and D. monticola, whereas species with scattered spines include D. angustifolia, D.fissa, D. grandis, D. melanochaetes and D. stenophylla. Spine type are net-like spine to triangular spines. All species of D. fissa complex have triangular spines, but only D. calicarpa has a net-like spine.
Figure 3 Spine types on the leaf sheath of D. fissa complex. [A] Oblique circle; [B] scattered
Leaves
The colour of D. fissa complex leaves are dark green. Both upper and under leaf surfaces are glabrous. The leaflets arrangement is pinnate with narrow and elliptic shape, but leaflets of D. grandis is broad with lanceolate shape. The number of leaflets on D. fissa complex is generally up to 50 – 90 pairs on each side, but D. grandis has up to 30 pairs.
Cirrus
Cirrus is extending from leaf apex. It is varied in length. All species D. fissa complex have cirrus length up to 100 cm – 125 cm, but D. calicarpa have short cirrus, 30 cm.
Petiole
Cross section of D. fissa complex petiole are channeled, the petiole adaxial surface is concave and ridged while the petiole abaxial surface convex.
Daemonorops stenophylla has shorter petiole (4 cm) than that of other species B
9 (10-30 cm). Daemonorops angustifolia has the longest petiole length up to 35 cm.
Knee
Type of knee is an important character for identification rattan species. Baja-lapis (2010) described 5 knee types, including bulgy, elongate-folded, elongate-flat, elongate inflated and cylindrical with ant holes. Daemonorops calicarpa, D. grandis and D. stenophylla have elongate-folded knees. D.
melanochaetes and D. monticola has elongate-flat knee. D. angustifolia has elongate-inflated knee; whereas D. fissa and D. lewisiana have bulgy knee (Figure 4). Knees are covered with spine like the leaf sheath.
Figure 4 Type of knee Daemonorops fissa complex. [A] bulgy; [B] elongate- folded; [C] elongate-flat; [D] elongate inflated.
Prophyll
Daemonorops fissa complex has persistent prophyll. The prophylls are covered many spines, but the density spine is varied. The spine density on prophyll are rarely to densely spines. Species with rarely spines (setose species) can be found on D. angustifolia, D. fissa, D. grandis, D. lewisiana, D. monticola,
and D. stenophylla (Figure 5). Two species, D. calicarpa and D. melanochaetes,
have densely spines (velutinose). There are two spine types among Daemonorops, net-like spine and triangular spine. However, D. calicarpa and D. melanochaetes
have different types of spines, D. calicarpa with net-like spine and D. melanochaetes with triangular spine. The prophyll of D. fissa complex are tubular and up to 45 cm long, but D. calicarpa is scarcely longer than 25 cm.
Figure 5 Density of spine in prophyll of Daemonorops fissa complex. [A] velutinose with triangular spine; [B] velutinose with like spine; [C] setose; [D] net-like spine; [E] triangular spine.
A B C E
C D
D
10
Inflorescence
Daemonorops is dioecious plants. There are two inflorescence types, pleonanthic and hapaxanthic. Stems will die after flowering in hapaxhantic type. Their inflorescence comprise of numerous lateral inflorescence arising from the axils of, often markedly, reduced leaves on the upper part of stem. While in pleonanthic type stem continues to grow even after flowering. Their inflorescence grow from the lower part of stem which then is continue to grow vegetatively, and reproduce over a relatively long period throughout its adult life (Baker et. al.
1999).All species of D. fissa complex have pleonanthic inflorescences, but D. calicarpa and D. lewisiana have hapaxhantic inflorescence.
Flower
Male flowers are up to 6 mm long with corolla splitted to the base into 3 petals. The petals have lanceolate shape and rounded at the base with 4 mm long. Corolla is twice longer than calyx. Calyx is 2 mm long. Each flower has 6 stamen, and with anther 2 mm long, female rachilla bears many flowers in dyads consisted of one female flower and one sterile male flower with empty anthers. Female flowers are 4 mm long with ovoid, corolla consisted of 3 petals, corolla as long as calyx. Calyx is truncate or shallowly 3 lobed. Sterile flower (4 mm long) is always shorter than male flower with empty anthers (Figure 6).
Figure 6 Flower of Daemonorops fissa complex. [A] flower of D. melanochaetes; [B] Male flower of D. angustifolia; [C] Female flower of D. melanochaetes; [D] Sterile male flower of D. melanochaetes. [1] sterile male flower; [2] female flower; [3] male flower; [4] calyx; [5] corolla; [6] androecium; [7] corolla; [8]
gynoecium; [9] corolla; [10] calyx; [11] empty anther. Fruit
All species of Daemonorops fissa complex have globose fruit, but their fruits are differed in fruit scale surface. Only Daemonorops monticola has
A B
5 6
4
7 8
1 2 3
C D
11 9
11 concave scale surface, the other species have flat scale surface (Figure 7). The scale of D. fissa complex are arranged in 15 – 18 vertical rows, but the scale of D. grandis from Aceh is arranged in 21 vertical rows.
Figure 7 Fruit scale surface of Daemonorops fissa complex. [A] concave; [B] flat Seed
The seed of nearly all rattan has an outer fleshy layer. This fleshy layer is the outer seed-coat and hence is properly called sacrotesta. sarcotesta varies greatly in thickness from species to species (Dransfield 1979). All species of
Daemonorops fissa complex have smooth seed surface, but there are two collections, D. grandis from Mount Kemiri, Aceh and D. angustifolia from Deleng Lancuk, Sumatra, having reticulate seed surfaces (Figure 8).
Figure 8 Seed surface of Daemonorops fissa complex. [A] smooth; [B] reticulate
Endosperm
Sarcotesta is easily separable from the rest of the seed. Endosperm varies from homogeneous in most genera to shallowly or deeply ruminate in
Daemonorops. A previous study showed that all Daemonorops species have ruminate endosperm (Dransfield 2008), but endosperms of Daemonorops fissa
complex from West Malesia are varied from slightly ruminate (D. calicarpa, D. fissa, D. lewisiana and D. monticola) to deeply ruminate endosperm (D. angustifolia, D. grandis, D. melanochaetes and D. stenophylla) (Figure 9).
Figure 9 Endosperm of Daemonorops fissa complex. [A] slightly ruminate; [B] deeply ruminate
A A
B
B
A B
12
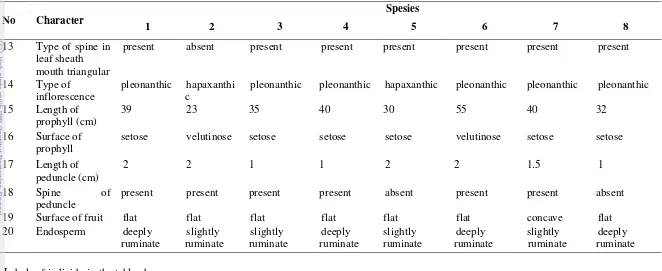
Table 2. Morphological variation of Daemonorops fissa complex in West Malesia N
scattered scattered oblique circle
absent present absent absent present absent present absent
6 Position spine in
present present absent absent absent present present absent
12 Type of spine in leaf sheath mouth net-like
13
present absent present present present present present present
14 Type of inflorescence
pleonanthic hapaxanthi c
pleonanthic pleonanthic hapaxanthic pleonanthic pleonanthic pleonanthic
15 Length of prophyll (cm)
39 23 35 40 30 55 40 32
16 Surface of prophyll
setose velutinose setose setose setose velutinose setose setose
17 Length of peduncle (cm)
2 2 1 1 2 2 1.5 1
18 Spine of
peduncle
present present present present absent present present absent
19 Surface of fruit flat flat flat flat flat flat concave flat
Labels of individu in the table above are:
1. D. angustifolia 5. D. lewisiana
2. D. callicarpa 6. D. melanochaetes
3. D. fissa 7. D. monticola
4. D. grandis 8. D. stenophylla
14
Anatomical Variation
Stomata type of Daemonorops fissa complex on paradermal section have been studied. The stomata types of all species are tetracytic (Figure 10). The type has two guard cells surrounded by four subsidiary cells (or cells neighbouring of the guard cells), two lateral and two polar ones (Metcalfe 1961, Dickison 1999). Stomata are infrequent in adaxial surface, and are restricted to intercostals region in abaxia surface. Terminal subsidiary cells are not well differentiated, sometimes having short size. Almost all stomata are present on abaxial leaf surface, but they are only fewer in adaxial surface.
1
2
Figure 10 Epidermal cell of Daemonorops. [A] abaxial surface; [B] adaxial surface, 1 stomata, 2 epidermis
Adaxial epidermis is always uniform, having cell rectangular with cell walls markedly sinuous but they do not have thickening anticlinal walls. Tellu (2006) showed that the stomata of Daemonorops species have kriptopor type with sunken position in the epidermis or sub-epidermis. In contrast to stomatal type, stomatal size shows variation within Daemonorops fissa complex. Daemonorops stenophylla has the smallest stomata size, 135.25-193.22 µm2, whereas D. monticola has the largest stomata size, 270.51-318.81 µm2.
Figure 11 Transverse section of leaf Daemonorops. [1] epidermis; [2] sclerenchym; [3] bulliform cell; [4] palisade; [5] sponge; [6] tannin; [7] phloem strands; [7] xylem.
100 µm
B A
100µm
1
4
8 2
3
5
7
15 Bulliform cells are commonly found in Monocotyledon, which have function for storing water. Bulliform, so called motor cells, are lose turgory under water deficit conditions, and thus constrict in upon themselves, causing lamina to fold or roll inward edge to edge (Dickison 1999).
The bulliform cells of Daemonorops fissa complex have thin walls, and their size are larger than that of the adjacent epidermal cells. There are usually 6-17 bulliform cells on the adaxial leaf surfaces. From the middle to the margin leaf, the bulliform cells on the abaxial epidermis are gradually getting smaller, and becoming the same size to the abaxial epidermal cells. The position of bulliform cell is varied, in adaxial and abaxial epidermis, between two vascular bundles, near midvein and mid-vein (Figure 12).
Figure 12 Position of bulliform cell in Daemonorops fissa complex. [A] between two vascular bundle; [B] near midvein; [C] midvein. Bulliform cell pointed by arrow.
All eight species of Daemonorops fissa complex have many bulliform cells on the upper and lower epidermis. The shape of bulliform cells on the upper epidermis are varied. Bulliform cells were curved downward found in D. angustifolia, D. binnendijkii, and D. calicarpa. Daemonorops fissa, D. lewisiana, D. monticola and D. stenophylla have bulliform cells parallel to epidermis, D. grandis has V shape, whereas the bulliform cells of D. melanochaetes are curved upward (Figure 13).
The mesophylls of Daemonorops fissa complex are differentiated into palisade and sponge tissues. Mesophylls usually consist of one layer of palisade tissue and three to five layers of sponge tissue. Daemonorops melanochaetes and
D. fissa only have one palisade tissue in upper side (dorsiventral), but D. calicarpa, D. lewisiana, D. grandis, D. melanochaetes and D. stenophylla have two palisade tissues (isobilateral) (Figure 14).
C
16
Figure 13 Type of bulliform cell in Daemonorops fissa complex. [A] letter V; [B] curved upward; [C] parallel to epidermis; [D] curved downward
Figure 14 Palisade tissue in Daemonorops fissa complex. [A] with one layer of
palisade tissue (dorsiventral); [B] with two layer of palisade tissue (isobilateral); 1. upper palisade, 2. under palisade.
Anatomical key to Daemonorops fissa complex in West Malesia
1. a. Stomata size ≤ 193.22 µm2 …………...………D. stenophylla
b. Stomata size > 193.22 µm2………....2 2. a. Number of tannin sac 3 –5………...…….3 b. Number of tannin sac 6 –8………..D. monticola
3. a. Have one type of bulliform cell...4 b. Have two type of bulliform cell ...………...D. grandis
4. a. Number of bulliform cell in upper epidermis ≤ 10………5 b. Number of bulliform cell in upper epidermis 12 –17…...D. lewisiana
5. a. Leaves isobilateral type………...D. fissa
b. Leaves dorsiventral type………...6 6. a. Length of upper palisade 20.33 µm2 ...D. calicarpa
b. Length of upper palisade > 23.56 µm2...7 7. a. Type of bulliform cell curved upward...D. melanochaetes
b. Type of bulliform cell curved downward...D. angustifolia D
A B
C
A B
1 1
17
In this study, 20 characters, mostly qualitative characters were selected for phenetic analysis (Table 1). A data matrix of characters was scored according to the species descriptions.
Cluster analysis separated all samples into two major groups with similarity coefficient of 0.36 (Figure 15). Group I are composed of 6 species which are D. angustifolia, D. fissa, D. grandis, D. melanochaetes, D. monticola
and D. stenophylla. Grup II contained, D. lewisiana and D. callicarpa. They were separated from six characters, total height, number of leaflets, width of leaflets, type of spine in leafsheath, position spine in petiole and type of inflorescence.
Based on the similarity coefficient, Daemonorops angustifolia, D. binnendijkii and D. trichroa are identic with similarity coefficient of 100%. Therefore, it is concluded that D. binnendijkii and D. trichroa are synonym of D. angustifolia. D. palembanica is a synonym of D. melanochaetes based on three characters: density spine in leaf sheath mouth, spine in prophyll velutinose and deeply endosperm. Grup I grouped by spine in leafsheath scattered, position spine in petiole only one surface and type of inflorescence is pleonanthic. D. lewisiana
and D. callicarpa grouped by total height ≤ 5 m tall, spine in leafsheath oblique circles and type of inflorescence hapaxanthic.
Strong character in taxonomic research is a character that only belongs to one group of taxa that can be used to differentiate it to other taxa (Wiley 1981). In this research, the type of knees, spines and width of leaflets are considered as strong characters. Type of knee is an important character for identification of rattan (Baja-lapis 2010). Spine and width of leaflets can be used as a diagnostic character to species. Based on dendogram, width of leaflets is a diagnostic characters to separated D. grandis from others. Previous study also showed that leaflet size is a diagnostic character (Dransfield 1976).
Figure 15 Dendogram of Daemonorops fissa complex in West Malesia based on morphological characters
II I
18
Table 3. Anatomical characters of Daemonorops fissa complex in West Malesia
Character
Spesies
D.angustifolia D.calicarpa D.fissa D.grandis D.lewisiana D.melanochaetes D.monticola D.stenophylla
Shape of epidermis Rectangular rectangular rectangular rectangular rectangular rectangular rectangular rectangular
Epidermis size (µm2) 20.00-36.75 23.00-39.68 30.00-74.40 24.58-50.00 23.00-39.68 20.00-36.75 24.58-50.00 30.00-74.40
Type of stomata Tetracytic tetracytic tetracytic tetracytic tetracytic tetracytic tetracytic tetracytic
Stomata size (µm2) 241.53-265.68
169.07-Position sclerenchym Palisade palisade palisade palisade palisade palisade palisade palisade
Palisade in lower epidermis
Present present absent present present absent absent absent
Number of tannin
parallel letter v parallel curved upward parallel parallel
Number of bulliform
19
Taxonomic Treatment
Daemonorops
Daemonorops Blume in J.A. & J.H. Schultes, Syst. Veg. 7(2):1333 (1830). – Type: Daemonorops melanochaetes Blume. West Sumatra, Padang near Ayer Manchor, August 1878. Beccari 831 (isotype K)
Solitary or clustering rattans, acaulescent to high climbing hapaxanthic (then always very short stemmed) or pleonanthic, dioecious. Leaf sheaths heavily armed with spines, spines frequently highly organized. Flagellum absent. Knee frequently present. Leaves ecirrate in acaulescent species or long cirrate. Leaflets variously arranged. Inflorescence male and female superficially similar, but within the genus of two basic types: one with all bracts enclosed within the outermost bract or prophyll, splitting along their length to expose the flowers (section
Cymbospatha) or the other with bracts splitting along their entire length to leave no tubular portion and frequently falling (section Piptospatha). Bracts variously armed. In the section Piptospatha partial inflorescences longer than the subtending bract; bracteoles and involucres inconspicuous. Male rachilla bearing male solitary flowers; male flowers with small cup shaped, calyx with three small lobes; corolla split to the base into 3 petals; stamen 6, slightly epipetalous; pistillode minute. Sterile male flower found with each female flower, as the fertile male, but stamens with empty anthers. Female rachilla bearing many flowers in dyads consists of one female flower and one sterile male flower. Female flower calyx truncate or shallowly 3 lobed; corolla with 3 petals; gynoecium with 3 stigmas and with 3 loculs. Sterile flower smaller or at least more slender than the female, with well formed calyx and corolla, 6 sterile stamens, an abortive ovary. Fruit variously shaped, tipped with stigmatic remains and covered with reflexed scales. Seed only one, covered by thin to thick sweet or sour sarcotesta. Endosperm deeply ruminate. Embryo basal.
Distribution. Geographical distribution of Daemonorops is more restricted than the genus Calamus. The center of their distribution are similar, from China, India to New Guinea, specifically Sumatra, Java, Borneo and Malay Peninsula. Daemonorops does not occur in Africa, Himalaya, Peninsular India, Sri Lanka and Australia (Dransfield
et al., 2008).
Habitat. Daemonorops grows in rather disturbed primary forest, on alluvial soil near rivers, terrain flat to gently sloping, ridge top, lowland forest, and steep hill slope primary forest on volcanic soils.
Uses. One species is recorded to have sweet, edible young shoot.
Morphological key to Daemonorops fissa complex in West Malesia
1. a. Stem pleonanthic climbing up to 30 m……….2 b. Stem hapaxanthic erect to 6 m………...7 2. a. Petiole absent or up to 4 cm long………D. stenophylla
b. Petiole present 20-30 cm long………...3 3. a. Leaflets broad ca, with the number of leaflets 30 on each side of the rachis,
20
b. Leaflets narrow, with the number of leaflets >50 on each side of rachis, peduncle >1 cm and armed……….………..4 4. a. Length of internodes ca <30 cm, surface of prophyll with velutinose spine
………..……….D. melanochaetes
b. Length of internodes ca >30 cm, surface of prophyll with setose spine…...5 5. a. Spine in leafsheath scattered and scale surface of fruit flat……...6 b. Spine in leafsheath oblique circle and scale surface of fruit concave………...D. monticola
6. a. Endosperm of seed slightly ruminate……….……...D. fissa
b. Endosperm of seed deeply ruminate………...…D. angustifolia
7. a. Knee bulgy, spine in leaf sheath mouth absent……...D. lewisiana
b. Knee elongate folded, spine in leaf sheath mouth with net-like shape……….…………..D. calicarpa
Species Description of Daemonorops fissa complex in West Malesia
Daemonorops angustifolia(Griff.) Mart.
Daemonorops angustifolia (Griff.) Mart. Hist, Nat. Palm. 3: 327. 1853. Daemonorops hygrophila (Griff.) Mart., Hist. nat. Palm. 3: 328. 1853.
Daemonorops carcharodon Ridl., Mat. Fl. Malay. Penins. 2: 178. 1907.
Daemonorops angustispatha Furtado, Gard. Bull. Straits. Settlem. 9: 161. 1937. – Type: Malay Peninsula, Province Wellesley, Bukit Juru, Desember 1845, H.N Ridley 7002 (holotype SING)
Daemonorops binnendijkii Becc. Ann. Roy. Bot. Gard. Calcutta 12 (1):67. 1911. Type: Bangka, Blinjoe island. 15 September 1914, Grashoof 7 (holotype BO!).
synon. nov.
Daemonorops trichroa Miq., Fl. Ned. Ind., Eerste Bijv.: 952.1861. Daemonorops trichroa Miq. Ann. Roy. Bot. Gard. Calcutta 12 (1):71. 1911.
Type: Palembang, Muara Dua. Teysmann 3582 (holotype BO!) synon. nov
21 longer than calyx, calyx 2 mm long, stamen 6. Female rachilla bearing many flowers in dyads consisted of one female flower and one sterile male flower with empty anthers. Pistillate inflorescences pendulous, to 36 cm long; peduncle 2 cm long, armed distally with solitary spines. Prophyll papery, erect, 36 cm long, 5 cm wide, ellipsoid oblong, armed with scattered spines, some spines in groups of 3; each inflorescence bearing up to 9-11 partial inflorescence. Female flowers 4 mm long, ovoid, calyx truncate or shallowly 3 lobed, corolla as long as calyx, corolla with 3 petals. Mature fruits globose, 2 x 1.4 cm2, covered by 15 – 19 (-18) vertical rows of scales. Seed smooth and reticulate, ovoid, 10 x 9 mm2. Endosperm deeply ruminate (Figure 16).
Leaves dorsiventral. Epidermal cell size 20 - 36.75 µm2, rectangular, cell walls sinous, no thickening anticlinal walls. Stomata on abaxial leaf surface, fewer in adaxial surface, tetracytyc, epidermal depressions or crypts; 241.53 - 265.68 µm2, terminal subsidiary cells not well differentiated, sometimes having short size. Bulliform cell in upper and lower epidermis, with curve downward type, 37.2 - 44.64 µm long, with 8 - 9 cell in upper, 8 - 9 cell in lower. Mesophyll consisted of one palisade layer and four sponge tissues layers. Sclerenchym supporting tissue well distributed in palisade. Tannin sac distributed in sponge with 3
– 5 cell. Vascular bundle sheath without bundle sheath extension.
Distribution. Malay Peninsula (Johor, Malaka, Negeri Sembilan, Perak, Selangor, Pahang), Singapore, Sumatra (widely distributed), Borneo (Pontianak)
Habitat and Ecology. This species found in lowland rain forest especially along stream and other wet places or more open areas to 1000 m elevation. In Malaya
Daemonorops angustifolia is widespread throughout the lowland, but there was no recorded in Perlis and Penang.
Vernacular name. Rotan getah
Uses. Umbut uses it as medicine, while Suku Anak Dalam uses it for appetite enhancer magh (Jumiati et al. 2012).
Notes. D. angustifolia is similar to D. binenndijkii, but it has different length of internodes. There are only two collections of D. binnendijkii from two collectors in Herbarium Bogoriense but type specimen is not found. This species has only been collected once in Indonesia (Grashoff 7, 1914) in Blinjoe, Bangka island. Therefore, D. binnendijkii is rated as Data Deficient (Bachman 2013). But, from this study, we found that transverse leaf section of D. binnendijkii is very similar to that of D. angustifolia.
Spesimen examined: Malay Peninsula: Seremban, Kuala Pilah Road, 18 Jul 1937, Furtado 33134; Ulu Kujang, 13 Nov 1935, EJH Corner 30447; Sungai kayu, 26 Oct 1936, Kiah 32173; Pahang, Sungai Bera in Tasek Bera, 17 Oct 1930, MR Henderson 24620; Lawn W, Singapore Botanic Garden, 13 Feb 1936,
22
Figure 16 Daemonorops angustifolia (Griff.) Mart. [A] herbarium type collection SING; [B] female and sterile male flower; [C] fruit; [D] endosperm; [E] knee; [F] latex; [G] cane
Daemonorops calicarpa (Griff.) Mart.
Daemonorops calicarpa (Griff.) Mart. Hist, Nat. Palm 3:326.1853. Daemonorops petiolaris (Griff.) Mart. Hist. Nat. Palm 3:326.1850. Daemonorops microthamnus
Becc. Rec. Bot. Surv. Ind.2:221. 1902. Daemonorops calicarpa (Griff.) Mart. Ann. Roy. Bot. Gard. Calcutta 12 (1):94. 1911.
Type: Malay Peninsula, Malacca, Griffith 6380 (isotype K)
Clustering rattan. Sheathed stem up to 1.5 cm in diam., stem without sheath up to 0.9 cm in diam. Leaf sheaths dull with oblique circle spines, triangular black spines up to 3 cm long, weak attach, with brown scaly indumentum, leaf sheath mouth armed with spine up to 3 cm. Knee elongate-folded, very conspicuous, armed as the rest of sheath. Ocrea poorly develop. Petiole 15 cm long, armed adaxially with short, erect, solitary, scattered spines to 1 cm long, abaxially armed with erect, very rare solitary spines, up to 1 mm long; rachis armed. Leaf blade to 2 m long, leaflets mostly arranged regularly, 45 on each rachis side, stiff, horizontal; lanceolate, papery, apex acuminate with an extension hair, up to 35 cm long, 1 cm wide, short bristles along the main nerve on lower surface, transverse veinlets conspicuous, short cirrus, armed with regularly arranged groups of grapnel-like spines. Inflorescence hapaxanthic. Staminate inflorescence pendulous 18 cm long. Prophyll erect 18 cm long, 4 cm width, armed with scattered, velutinose, net-like spine. Peduncle 1 cm long with net-like spine. Male flowers 6 mm long, corolla split to the base into 3 petals, lanceolate, 4 mm long, twice longer than calyx, calyx 2 mm long, stamen 6. Female rachilla bearing many flowers in dyads consisted of one female and one sterile male flower with empty anthers. Pistillate inflorescences pendulous to 23 cm long; peduncle 2 cm long, armed distally with solitary spine. Prophyl papery, erect, 24 cm long, 4 cm width, ellipsoid oblong, armed with net-like spines, each
B C D
E F
A
G
23 inflorescence bearing up to 11 partial inflorescence. Female flowers 4 mm long, ovoid, calyx truncate or shallowly 3 lobed, corolla as long as calyx, 3 petals. Mature fruits globose, 1.9 x 1.5 cm2, covered by 18 vertical rows of scales. Seed smooth, ovoid. Endosperm slightly ruminate (Figure 17).
Leave dorsiventral. Epidermal cell size 23 - 39.68 µm2 long, rectangular, cell walls sinous, no thickening anticlinal walls. Stomata on abaxial leaf surface, fewer in adaxial surface, tetracytyc, epidermal depressions or crypts; 169.07-217.37 µm2 long, terminal subsidiary cells not well differentiated, sometimes having short size. Bulliform cell in upper and lower epidermis, curve downward type, 24.8 µm, 7 cell in upper, 9 cell in lower. Mesophyll consisted of one palisade layer and four sponge tissues layers. Sclerenchym supporting tissue well distributed in palisade. Tannin sac distributed in sponge with 3 – 6 cells. Vascular bundle sheath without bundle sheath extension.
Distribution. Malay Peninsula (Perak, Pahang, Selangor, Negri Sembilan, Malacca, Johore, Kemaman), North Sumatra.
Habitat and Ecology. This species found in lowland rain forest at 50 m to 1900 m elevation
Vernacular name. unknown
Uses. The leaves are used for thatch, and the leaf rachis cleaned of thorn is for some forms of basketry (Dransfield 1979)
Notes. D. calicarpa is similar to D. lewisiana, but it can be distinguished by three characters i.e type of knee is elongate-folded, present spine in leafsheath mouth and prophyllsurface is velutinose.
Spesimen examined: Malay Peninsula: Pahang, Bentong, Sungai Kesar, 14 Jun 1937. CX Furtado 33118. Ulu, Bendong, Kajang, Kemaman, 30 Oct 1935. EJH Corner 30077. Johore, Gunung kajang, Pulau Tioman, 17 May 1927. Md Nur 18938. Malaya, 13 Feb 1936, CX Furtado 30661. Negeri Sembilan, Beremban, 18 Jun 1937, CX Furtado 33131. Sumatra: Lingga Archipelago, 11 Aug 1919 HAB Bunnenmeyer 7451. North Sumatra: Ulu Besitang, Tanjung Pura, 14 Aug 1971 J. Dransfield 1831.
Figure 17 Daemonorops calicarpa (Griff.) Mart. [A] herbarium type collection K; [B] spine oblique circle in leaf sheaths; [C] flower; [D] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
A B C
D
2 3
24
Daemonorops fissa Blume.
Daemonorops fissa Blume, Rumphia. 3: 17. 1847. Daemonorops cinnamomea
Schaedtler, Hamburger Garten-Blumenzeitung 31: 161. 1875. Daemonorops fissa
var. cinnamomea Becc., Rec. Bot. Surv. India 2: 219. 1902. nom. nud. Daemonorops hallieriana Becc., Ann. Roy. Bot. Gard. (Calcutta) 12(1): 218 1911.Daemonorops fissa Blume. Ann. Roy. Bot. Gard. (Calcutta) 12(1): 65. 1911.
Type: River Dusson in South Borneo, Miquel sn (holotype L)
Clustering rattan. Sheathed stem up to 4 cm in diam., stem without sheath up to 3 cm in diam. Leaf sheaths greenish brown with scattered and grouped spines, triangular black spines up to 2 cm long, strong attach, with brown indumentum, leaf sheath mouth smooth. Knee bulgy, very conspicuous, smooth to armed as the rest of sheath. Ocrea poorly develop. Petiole 40 cm long, armed adaxially with short, erect, solitary, scattered spines to 1 cm long, abaxially armed with erect, very rare solitary spines, up to 1 mm long; rachis armed. Leaf blade to 3 m long, leaflets mostly arranged regularly, 100 on each rachis side, stiff, horizontal, lanceolate, papery, apex acuminate with an extension hair, up to 45 cm long, 2.2 cm width, short bristles along the main nerve on lower surface, transverse veinlets conspicuous, cirrus up to 112 cm long, armed with regularly arranged groups of grapnel-like spines. Inflorescence pleonanthic. Staminate inflorescence pendulous, 28 cm long. Prophyll erect, 28 cm long, 6 cm width, armed with scattered, setose, triangular and solitary spine; peduncle 1 cm, armed to smooth. Male flowers 6 mm long, calyx 2 mm long, corolla split to the base into 3 petals, twice longer than calyx, lanceolate, 4 mm long, stamen 6. Female rachilla bearing many flowers in dyads consisted of one female flower and one sterile male flower with empty anthers. Pistillate inflorescences pendulous, to 32 cm long; peduncle 1 cm long, armed distally with solitary spines; prophyl papery, erect, 32 cm long, 5 cm width, ellipsoid oblong, armed with scattered spines,
Leaves isobilateral. Epidermal cell size 30-74.4 µm2 long, rectangular, cell walls sinous, no thickening anticlinal walls. Stomata on abaxial leaf surface, fewer in adaxial surface, tetracytyc, epidermal depressions or crypts; 217.37-265.68 µm2, terminal subsidiary cells are not well differentiated, sometimes having short size. Bulliform cell in upper and lower epidermis, with parallel type, 32.24-49.6 µm, 8-10 cell in upper, 8 cell in lower. Mesophyll consisted of one palisade layer and four sponge tissues layers. Sclerenchym supporting tissue well distributed in palisade. Tannin sac distributed in sponge with 5 – 7 cells. Vascular bundle sheath without bundle sheath extension.
Distribution. Culta, Sumatra (widely distributed in Jambi, Bengkulu, Bangka), Singapore, Borneo (Sabah, Sarawak, Pontianak, Samarinda, Banjarmasin)
25 Vernacular name. Rotan Kotok
Uses. Unknown
Notes. Daemonorops fissa from Borneo and Sumatra have a little differences. Spine on leafsheath from Borneo is stronger than that from Sumatra. Peduncle from Sumatra individual is unarmed, while peduncle from Borneo is armed. Spesimen examined: Sumatra: Bengkulu, Desa Air Putih, Lebong Utara, Rejang Lebong, Taman Nasional Kerinci Seblat, 23 Agu 2000, JW 180. Bangka, Desa Menjelang, Lundang, Kitok Mangkura, Sertong, 8 Mei 2011, V3 002,
Bangka, Desa Tanjung Gunung, Kabupaten Tengah, 3 April 2011, ZA 22, Bangka, Bukit Permis Kecamatan Simpang Rimba, 23 Maret 2011, SHER 006. Borneo: Kalimantan Barat, Kabupaten pasir, Kecamata Batu Kajang, 1 Des 1979, JPM 1884. Kalimantan Selatan, District upper tabalong area, Muara Pasuang Ayu, 3 Agu 2000, JPM 7390. Kalimantan Selatan, District upper tabalong area, Muara Pasuang Ayu, 3 Agu 2000, JPM 7393. Kalimantan Selatan, Barabai, Datar alai, 26 Jul 1976, JPM 702. Kalimantan Timur, Tabang, Gunong Mendam, 15 Jan 1979, JPM 1617. Sarawak, Padawan, hills near Belimbang, 12 Apr 1981, JD 5901. Kalimantan Barat, Sintang, Bukit Baka National Park, 0037’S 112015E alt 330m, 17 Feb 1994, JPM 441.
Figure 18 Daemonorops fissa Blume [A] herbarium collection BO; [B] knee; [C] cirrus; [D] fruit; [E] flower; [F] endosperm. [1] sterile male flower; [2] female flower; [3] male flower.
Daemonorops grandis (Griff.) Mart.
Daemonorops grandis (Griff.) Mart., Hist. Nat. Palm. 3: 327. 1853.
Daemonorops intermedia (Griff.) Mart., Hist. Nat. Palm. 3: 327. 1853. Daemonorops kirtong Griff., Palms Brit. E. Ind.: 102. 1850. Daemonorops malaccensis Mart., Hist. Nat. Palm. 3: 327. 1853.Daemonorops grandis var.
megacarpus Furtado, Gard. Bull. Singapore 14: 67. 1953. Daemonorops laciniatus Furtado, Gard. Bull. Singapore 14: 75. 1953.
Type: Malay Peninsula, Perak, Bukit Chom, Kroh, 24 May 1934, CX Furtado 33016 (holotype SING)
1
F E
D C
B A
2
26
Clustering rattan. Sheathed stem up to 3 cm in diam., stem without sheath up to 2 cm in diam. Leaf sheaths pale green to pale brownish with scattered and grouped spines, triangular black spines up to 5 cm long, strong attach, with brown indumentum, leaf sheath mouth smooth. Knee elongate-folded, very conspicuous, smooth. Ocrea poorly develop. Petiole 40 cm long, armed adaxially with short, erect, solitary, scattered spines to 1 cm long, abaxially armed with erect, very rare solitary spines, up to 1 mm long; rachis armed. Leaf blade to 3.5 m, leaflets mostly arranged regularly, 60 on each rachis side, stiff, horizontal; leaflets lanceolate, papery, apex acuminate with an extension hair, up to 42 cm long, 3.2 cm width, short bristles along the main nerve on lower surface, transverse veinlets conspicuous, cirrus up to 125 cm long, armed with regularly arranged groups of grapnel-like spines. Inflorescence pleonanthic. Staminate inflorescence pendulous 36 cm long. Prophyll erect 36 cm long, 6 cm width, armed with scattered, setose, triangular and solitary spine, peduncle 1 cm, smooth. Male flowers 6 mm long, calyx 2 mm long, corolla split to the base into 3 petals, lanceolate, 4 mm long, corolla twice longer than calyx, stamen 6. Female rachilla bearing many flowers in dyads consisted of one female flower and one sterile male flower with empty anthers. Pistillate inflorescences pendulous to 27 cm long, peduncle 1 cm long, smooth, prophyl papery, erect, 27 cm long, 5 cm width, ellipsoid oblong, armed with scattered spines, some spines in groups of 3, each inflorescence bearing up to 11 partial inflorescence. Female flowers 4 mm long, ovoid, calyx truncate or shallowly 3 lobed,corolla as long as calyx, 3 petals. Mature fruits globose, 1.9 x 2.1 cm2, covered by 15 –17 vertical rows of scales. Seed smooth, ovoid, 14 x 15 mm2. Endosperm deeply ruminate (Figure 19).
Leaves dorsiventral. Epidermal cell size 24.58-50 µm2 long, rectangular, cell walls sinous, no thickening anticlinal walls. Stomata on abaxial surface, fewer in adaxial surface, tetracytyc, epidermal depressions or crypts, 217.37-318.81µm2, terminal subsidiary cells well differentiated, sometimes having short size. Bulliform cell in upper and lower epidermis, with letter V type, 24.8-49.6 µm, 5-8 cell in upper and 14 cell in lower. Mesophyll consisted of one palisade layer and four sponge tissues layers. Sclerenchym supporting tissue well distributed only in palisade. Tannin sac distributed in sponge with 5 – 8 cell. Vascular bundle sheath without bundle sheath extension.
Distribution. Malay Peninsula (Perak, Pahang, Selangor, Negri Sembilan, Malacca). Singapore, Sumatra (Aceh, North Sumatra)
Habitat and Ecology. Daemonorops grandis is common in lowland rain forest to 800 m elevation.
Vernacular name. Rotan relang (sakai),
Uses. Provides a poor-quality cane, which is seldom used; the palm hearts are used for medicinal (Henderson 2009).
Notes. Daemonorops grandis is easily distinguished by its broad leaflets, the number of leaflets 30 on each sides, rachis and petiole tinged crimson when young.
![Figure 1 Part of rattan. [A] Vegetative parts. [B] Generative parts. 1. cirrus, 2.](https://thumb-ap.123doks.com/thumbv2/123dok/593652.438311/18.595.86.481.326.531/figure-rattan-vegetative-parts-b-generative-parts-cirrus.webp)

![Figure 5 Density of spine in prophyll of Daemonorops fissa [A] velutinose complex](https://thumb-ap.123doks.com/thumbv2/123dok/593652.438311/23.595.105.509.87.813/figure-density-spine-prophyll-daemonorops-fissa-velutinose-complex.webp)

![Figure 9 Endosperm of Daemonorops fissa complex. [A] slightly ruminate; [B]](https://thumb-ap.123doks.com/thumbv2/123dok/593652.438311/25.595.117.460.156.234/figure-endosperm-daemonorops-fissa-complex-slightly-ruminate-b.webp)
![Figure 11 Transverse section of leaf Daemonorops. [1] epidermis; [2]](https://thumb-ap.123doks.com/thumbv2/123dok/593652.438311/28.595.88.455.516.696/figure-transverse-section-leaf-daemonorops-epidermis.webp)
![Figure 12 Position of bulliform cell in Daemonorops fissa complex. [A] between](https://thumb-ap.123doks.com/thumbv2/123dok/593652.438311/29.595.114.494.236.479/figure-position-bulliform-cell-daemonorops-fissa-complex.webp)
