RESPON MORFOLOGIS DAN FISIOLOGIS
BIBIT BERBAGAI GENOTIPE JARAK PAGAR (JATROPHA
CURCAS L.) TERHADAP CEKAMAN KEKERINGAN
SARTIKA SYAFI
PROGRAM STUDI AGRONOMI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan sebenar-benarnya, bahwa segala pernyataan dalam tesis yang berjudul: Respon Morfologis dan Fisiologis Bibit Berbagai Genotipe Jarak Pagar (Jatropha curcas L.) terhadap cekaman kekeringan adalah karya saya sendiri dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2008
ABSTRACT
SARTIKA SYAFI. Morphological and Physiological Response of Six Genotypes of Jatropha curcas L. seedling to water stress. Under direction of ENDANG MURNIATI and ENDAH RETNO PALUPI.
Development of Jatropha in Indonesia is focused on marginal lands to avoid competition with food crops. Therefore, drought tolerant varieties/clones with high productivity are needed in order to meet the target of 1.5 ha jatropha plantation in 2010. The research was aimed at: 1) studying morphological and physiological response of six genotypes of Jatropha curcas L. seedling to drought stress, 2) determining drought tolerance of the six genotypes. The research was conducted in Seed Science and Technology Laboratory at Leuwikopo. Morphological and physiological response was analyzed at Ecophysiology Laboratory and RGCI Laboratory from July to Oktober 2007.
Split plot with three replication was used as experimental designed, with soil moisture content, ie. K1=40%, K2=36%, K3=32% as the main plot and genotype as sub-plot, ie. A1=Karanganyar, A2=Sukabumi, A3=NTB (Dompu), A4=IP-1P, A5=IP-1M dan A6= IP-1A. The data was analysed using SAS and DMRT at α=0.05. The results show that soil moisture content significantly affected leaf area, number of opened stomata, shoot dry weight, root dry weight, shoot/root ratio, root length, leaf water content, net assimilation rate, chlorophyll content and carbon content. Leaf area, shoot dry weight, root length and net assimilation rate were also affected by genotype. The interaction of soil moisture content and genotype only affected plant height, leaf thickness and root volume. The increment of plant height averaged of 14.07, 11.02 and 9.73 cm was obtained 8 week after the seedlings were grown on 40, 36 and 32% soil moisture content respectively, with genotype from Sukabumi had the highest increment. Although soil moisture affected the number of opened stomata in which the lower soil moisture resulted in lower number, however, it was not vary among genotypes. Likewise was the shoot dry weight.
The low soil moisture (32%) reduced root length. The lower the soil moisture the lower the root length, so as root volume. The reduction of root volume was lowest in IP-1M. Leaf water content was reduced only on those planted in 32% soil moisture. Soil moisture of 32% was considered as drought stress condition therefore was used to evaluate drought tolerance of the six genotypes. The sensitivity index show that NTB was the most tolerant genotypes, followed by IP-1M, Karanganyar, IP-1P, Sukabumi and IP-1A respectively.
Ringkasan
SARTIKA SYAFI. Respon Morfologis dan Fisiologis Bibit Berbagai Genotipe Jarak Pagar (Jatropha curcas L.) Terhadap Cekaman Kekeringan. Dibimbing oleh ENDANG MURNIATI dan ENDAH RETNO PALUPI.
Tanaman jarak pagar merupakan tanaman tahunan yang tahan kekeringan. Jarak pagar dapat beradaptasi pada lahan ataupun agroklimat di Indonesia bahkan tanaman ini dapat tumbuh baik pada lahan dengan tingkat kesuburan rendah (lahan kritis). Budidaya tanaman jarak pagar sebaiknya lebih difokuskan ke lahan kering agar tidak berkompetisi dengan budidaya tanaman pangan, sehingga pengembangan tanaman jarak yang paling sesuai untuk dilakukan yaitu di lahan marginal seperti di wilayah Indonesia Timur. Penelitian ini bertujuan untuk: (1) mempelajari respon morfologis dan fisiologis bibit berbagai genotipe tanaman jarak pagar terhadap cekaman kekeringan dan (2) mendapatkan informasi toleransi terhadap kekeringan masing-masing genotipe yang digunakan. Penelitian ini dilaksanakan di rumah kaca Laboratorium Ilmu dan Teknologi Benih Leuwikopo. Analisis fisiologis dan morfologis di laboratorium Ekofisiologi dan RGCI, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor, pada bulan Juli sampai dengan November 2007.
Rancangan yang digunakan adalah rancangan petak terbagi (split plot) dengan tiga ulangan. Faktor pertama, sebagai petak utama adalah kadar air media terdiri atas K1 : kadar air media 40%, K2 : kadar air media
36%, K3 : kadar air media 32%. Faktor kedua, sebagai anak petak adalah
genotipe terdiri atas A1 : Karanganyar, A2 : Sukabumi, A3 : NTB, A4 : IP-1P,
A5 : IP-1M dan A6 : IP - 1A. Peubah morfologis yang diamati adalah tinggi
tanaman, luas daun, ketebalan daun, jumlah stomata terbuka, defisit air, bobot kering pucuk, bobot kering akar, ratio pucuk akar, panjang akar dan volume akar, sedangkan peubah fisiologi yang diamati adalah kadar air daun, laju asimilasi bersih, kandungan klorofil, kandungan karbon dan kandungan prolin. Data dianalisis dengan menggunakan SAS, bila berpengaruh nyata dilanjutkan dengan uji Duncan pada taraf 5%.
Kadar air media yang rendah (32%) menurunkan panjang akar. Semakin rendah kadar air media maka panjang akar juga makin rendah, demikian juga volume akar. Penurunan volume akar paling rendah terjadi pada IP-1M. Kadar air daun tanaman yang ditanam pada kadar air media 32% lebih rendah dibandingkan dengan tanaman pada kadar air media 40 dan 36%. Kadar air media 32% dikategorikan sebagai kondisi cekaman kekeringan, sehingga digunakan dalam evaluasi toleransi kekeringan pada keenam genotipe yang diamati. Indeks sensitivitas keenam genotipe menunjukkan bahwa genotipe NTB yang paling toleran terhadap kekeringan diikuti berturut-turut oleh IP-1M, Karanganyar, IP-1P, Sukabumi dan IP-1A.
©Hak cipta milik IPB, tahun 2008 Hak cipta dilindungi
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan ini hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan keritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugiikan kepentingan yang wajar IPB.
RESPON MORFOLOGIS DAN FISIOLOGIS BIBIT BERBAGAI GENOTIPE JARAK PAGAR (Jatropha curcas L.) TERHADAP
CEKAMAN KEKERINGAN
SARTIKA SYAFI
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Departemen Agronomi dan Hortikultura
PROGRAM STUDI AGRONOMI SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : RESPON MORFOLOGIS DAN FISIOLOGIS BIBIT BERBAGAI GENOTIPE JARAK PAGAR (Jatropha curcas L.) TERHADAP CEKAMAN KEKERINGAN Nama Mahasiswa : Sartika Syafi
Nomor Pokok : A 351050031 Program Studi : Agronomi
Disetujui
Komisi Pembimbing
Dr.Ir.Endang Murniati, MS. Dr.Ir.Endah Retno Palupi, M.Sc. Ketua Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana IPB
Dr. Ir. Munif Ghulamahdi, MS. Prof. Dr. Ir. Khairil A. Notodiputro, M.S.
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala karunia dan hidayah-Nya sehingga penulis dapat menyelesaikan penulisan tesis ini. Penelitian yang dilaksanakan dari bulan Juli sampai dengan November 2007 ini berjudul: Respon morfologis dan fisiologis bibit berbagai genotipe jarak pagar (Jatropha curcas L.) terhadap cekaman kekeringan.
Pada kesempatan ini penulis menyampaikan terimakasih yang setulus-tulusnya dan penghargaan yang tinggi kepada:
1. Dr.Ir. Endang Murniati, MS sebagai ketua komisi pembimbing dan Dr.Ir. Endah Retno Palupi, MSc selaku anggota komisi pembimbing, atas segala bantuan, arahan dan bimbingan yang dengan sabar mereka berikan kepada penulis sejak awal penelitian sampai penulisan tesis ini.
2. Dr.Ir.Suwarto, MS selaku dosen penguji yang telah memberi saran dan masukan dalam penyempurnaan penulisan tesis ini.
3. Pimpinan dan Staf di lingkup Sekolah Pascasarjana Institut Pertanian Bogor, atas segala pendidikan, layanan administrasi dan bantuan yang telah diberikan.
4. Pimpinan, staf dan pegawai Laboratorium Ekofisiologi Fakultas Pertanian IPB, atas segala bantuan yang telah diberikan selama aktivitas di laboratorium.
5. Pimpinan, staf dan pegawai Laboratorium Research Group on Crop Improvement (RGCI) Fakultas Pertanian IPB, atas segala bantuan yang telah diberikan selama aktivitas di laboratorium.
6. Pimpinan, staf dan pegawai Laboratorium Ekofisiologi Tanaman, BALITRO. Cimanggu, atas segala bantuan yang telah diberikan selama aktivitas di laboratorium.
7. Rektor Universitas Muhammadyah yang telah memberikan izin untuk melanjutkan study.
8. Ayahanda dan Ibunda tercinta atas doa dan restunya yang selalu menyertai penulis dalam keadaan suka maupun duka yang semuanya tidak akan terbalaskan, hanya Yang Maha Kuasa penulis serahkan segalanya.
9. Kakak-kakak dan Adikku tersayang atas dukungannya baik moril, spirituil dan materil, semoga semuanya mendapat tempat di sisi Yang Maha Kuasa.
10. Teman-teman Pascasarjana Fakultas Pertanian Ir.Ince MS, Ir Iskandar MS, Ir Bambang MS, Safrizal SP.Msi. yang telah memberikan dukungannya baik dalam keadaan suka maupun duka.
Tesis ini merupakan upaya maksimal yang dapat penulis sajikan. Namun demikian sebagai manusia biasa yang tidak luput dari Khilaf, salah dan keliru dengan senang hati menerima kritik dan saran yang bermanfaat dari semua pihak demi penyempurnaannya. Semoga segala bantuan yang diberikan mendapat balasan yang setimpal dari Allah SWT. Amin.
Bogor, Agustus 2008
RIWAYAT HIDUP
Penulis dilahirkan di Ternate pada tanggal 18 Maret 1978 dari Ayah Hi. Syafi Do. Hi. Thalib dan Ibu Robo Adam. Penulis merupakan putri ke enam dari tujuh bersaudara.
Penulis menyelesaikan pendidikan sekolah dasar di Sekolah Dasar Negeri Dufa-Dufa 2 Ternate (1992), sekolah menengah pertama di Sekolah Menengah Pertama Negeri 2 Ternate (1995) dan sekolah menengah atas di Sekolah Menengah Atas Negeri 5 Ternate (1998).
Pendidikan sarjana ditempuh di Jurusan Budidaya Pertanian, Universitas Khairun Ternate, lulus pada tahun 2002. Selanjutnya pada tahun 2005 penulis mendapatkan kesempatan untuk melanjutkan pendidikan ke Program Magister pada Program Studi Agronomi di sekolah Pascasarjana IPB. Beasiswa pendidikan pascasarjana diperoleh dari BPPS.
DAFTAR ISI Respon Morfologis Tanaman terhadap Cekaman Kekeringan... Respon Fisiologis Tanaman terhadap Cekaman Kekeringan...
BAHAN DAN METODE Pengaruh Perlakuan terhadap Peubah yang diamati...
Respon Morfologis Enam Bibit Genotipe Jarak Pagar terhadap Cekaman Kekeringan... Respon Fisiologis Enam Bibit Genotipe Jarak Pagar terhadap Cekaman Kekeringan... Toleransi terhadap Cekaman Kekeringan...
KESIMPULAN DAN SARAN
Kesimpulan... Saran...
DAFTAR PUSTAKA...
LAMPIRAN... 49 49
50
DAFTAR TABEL
Kadar air benih, daya berkecambah dan tinggi bibit genotipe jarak pagar yanng digunakan dalam penelitian ..
Rekapitulasi sidik ragam pengaruh kadar air media dan genotipe terhadap karakter morfologis dan fisiologis tanaman jarak pagar (Jatropha curcas L.) pada akhir pengamatan...
Pengaruh interaksi antara kadar air media dan genotipe terhadap tinggi tanaman (cm) jarak pagar pada akhir pengamatan...
Pertambahan tinggi tanaman selama 8 minggu setelah perlakuan pada masing-masing genotype dan kadar air media………...
Pengaruh kadar air media terhadap luas daun (cm2) tanaman jarak pagar pada akhir pengamatan...
Pengaruh genotipe terhadap luas daun (cm2) tanaman jarak pagar pada akhir pengamatan...
Pengaruh interaksi antara kadar air media dan genotipe terhadap ketebalan daun (mm) jarak pagar pada akhir pengamatan………
Pengaruh kadar air media terhadap jumlah stomata terbuka, stomata tertutup dan total stomata tanaman jarak pagar pada akhir pengamatan...
Pengaruh genotipe terhadap jumlah stomata terbuka, tertutup dan total stomata tanaman jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap defisit air (%) jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap bobot kering pucuk (g) jarak pagar pada akhir pengamatan...
Pengaruh genotipe terhadap bobot kering pucuk (g) tanaman jarak pagar pada akhir pengamatan...
14.
Pengaruh kadar air media terhadap rasio pucuk akar jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap panjang akar (cm) jarak pagar pada akhir pengamatan...
Pengaruh genotipe terhadap panjang akar (cm) jarak pagar pada akhir pengamatan...
Pengaruh interaksi antara kadar air media dan genotipe terhadap volume akar (cm3) jarak pagar pada akhir pengamatan...
Perbedaan volume akar (cm3) pada masing-masing genotype dan kadar air media (40% - 36% dan 36% - 32%).………
Pengaruh kadar air media terhadap kadar air daun (%) jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap laju asimilasi bersih (g cm2hari-1) jarak pagar pada akhir pengamatan...
Pengaruh genotipe terhadap Laju asimilasi bersih (g m2hari-1) jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap kandungan klorofil (mg/g) jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap kandungan karbon (%) jarak pagar pada akhir pengamatan...
Pengaruh kadar air media terhadap kandungan prolin (μg prolin/g bobot segar) tanaman jarak pagar pada akhir pengamatan...
Rekapitulasi hasil pengamatan rata-rata pengaruh kadar air media terhadap berbagai peubah yang diamati...
Matriks tingkat toleransi pada 15 peubah berdasarkan nilai indeks sensitivitas yang diamati pada masing-masing genotipe...
DAFTAR GAMBAR
No
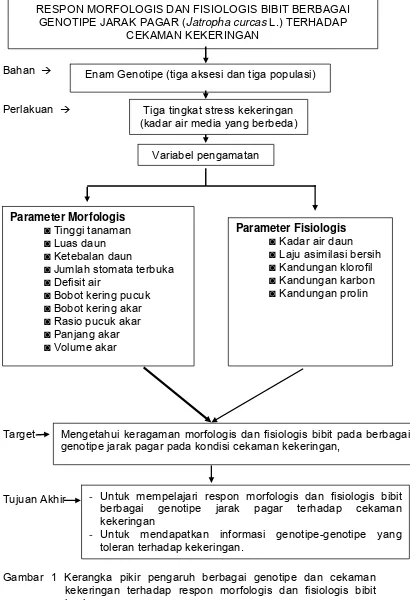
1. Kerangka pikir pengaruh berbagai genotipe dan cekaman kekeringan terhadap respon morfologis dan fisiologis bibit jarak pagar...
Halaman
DAFTAR LAMPIRAN
Sidik ragam pengaruh kadar air media dan genotipe terhadap tinggi tanaman pada akhir pengamatan…………
Sidik ragam pengaruh kadar air media dan genotipe terhadap luas daun pada akhir pengamatan……….
Sidik ragam pengaruh kadar air media dan genotipe terhadap ketebalan daun pada akhir pengamatan………..
Sidik ragam pengaruh kadar air media dan genotipe terhadap jumlah stomata pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap defisit air pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap bobot kering pucuk jarak pagar pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap bobot kering akar jarak pagar pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap rasio pucuk akar jarak pagar pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap panjang akar pada akhir pengamatan…………...
Sidik ragam pengaruh kadar air media dan genotipe terhadap volume akar pada akhir pengamatan………
Sidik ragam pengaruh kadar air media dan genotipe terhadap kadar air daun pada akhir pengamatan…………
Sidik ragam pengaruh kadar air media dan genotipe terhadap laju asimilasi bersih (NAR) pada akhir pengamatan...
Sidik ragam pengaruh kadar air media dan genotipe terhadap klorofil daun pada akhir pengamatan...
14.
Sidik ragam pengaruh kadar air media dan genotipe terhadap kandungan karbon pada akhir pengamatan…….
Sidik ragam pengaruh kadar air media dan genotipe terhadap kandungan prolin pada akhir pengamatan...
Nilai IS berdasarkan peubah tinggi tanaman...
Nilai IS berdasarkan peubah luas daun...
Nilai IS berdasarkan peubah ketebalan daun...
Nilai IS berdasarkan peubah jumlah stomata...
Nilai IS berdasarkan peubah defisit air...
Nilai IS berdasarkan peubah bobot kering pucuk...
Nilai IS berdasarkan peubah bobot kering akar...
Nilai IS berdasarkan peubah ratio pucuk akar...
Nilai IS berdasarkan peubah panjang akar...
Nilai IS berdasarkan peubah volume akar...
Nilai IS berdasarkan peubah kadar air daun...
Nilai IS berdasarkan peubah laju asimilasi bersih (NAR)...
Nilai IS berdasarkan peubah kandungan klorofil...
Nilai IS berdasarkan peubah kandungan karbon...
Nilai IS berdasarkan peubah kandungan prolin...
Absorbansi Larutan Standar – Prolin...
PENDAHULUAN
Latar Belakang
Cadangan minyak mentah dunia semakin hari semakin berkurang sedangkan kebutuhan semakin meningkat seiring dengan pertambahan penduduk dunia. Meningkatnya konsumsi minyak dunia telah berakibat pada meningkatnya harga minyak bumi hingga mencapai di atas $ 130 per barel. Harga ini diperkirakan akan terus meningkat seiring terjadinya kelangkaan sumber energi tersebut. Menghadapi kenaikan harga minyak pada tahun 2005, pemerintah Indonesia melakukan kebijakan pengembangan bahan bakar nabati (BBN) yang merupakan sumber energi yang dapat diperbaharui dan ramah lingkungan.
Saat ini berbagai cara digunakan untuk mengantisipasi hal tersebut, maka sumber energi alternatif seperti bahan bakar nabati perlu dicari bahkan terus diperbaharui setiap waktu sebagai persediaan sepanjang waktu. Bahan bakar nabati dapat digunakan baik secara murni maupun dicampur dengan petrodiesel tanpa terjadi perubahan pada mesin diesel kendaraan atau mesin lain yang menggunakannya (Mahmud 2006). Penggunaan bahan bakar nabati sebagai sumber energi semakin mendesak untuk direalisasikan. Bahan bakar nabati yang merupakan salah satu solusi menghadapi kelangkaan bahan bakar minyak pada masa mendatang yang bersifat ramah lingkungan, dapat diperbaharui serta mampu menurunkan gas buang dan efek gas rumah kaca. Beberapa tanaman yang mempunyai potensi sebagai bahan bakar nabati (BBN) antara lain kelapa sawit, kelapa, ubi kayu, ubi jalar, tebu, kedelai, jagung dan jarak pagar (Prawitasari et al. 2006).
2
mengendalikan keong emas dan sebagai obat, minyak jarak dapat digunakan untuk meredakan rasa sakit karena rematik. Pada industri tekstil tanaman jarak digunakan sebagai bahan pewarna (Heyne 1987).
Tanaman jarak pagar (Jatropha curcas L.) merupakan salah satu tanaman primadona sebagai sumber energi alternatif yang sangat prospektif dan dapat dimanfaatkan sebagai bahan bakar pengganti solar. Minyak jarak pagar tidak termasuk dalam kategori minyak makan (edible oil), sehingga pemanfaatannya sebagai biodiesel tidak akan mengganggu penyediaan kebutuhan minyak makan nasional, kebutuhan industri dan ekspor CPO (Prawitasari et al. 2006).
Tanaman jarak pagar merupakan tanaman tahunan yang tahan kekeringan. Jarak pagar dapat beradaptasi pada lahan ataupun agroklimat di Indonesia bahkan tanaman ini dapat tumbuh baik pada lahan dengan tingkat kesuburan rendah (lahan kritis). Budidaya tanaman jarak pagar sebaiknya lebih difokuskan ke lahan kering agar tidak berkompetisi dengan budidaya tanaman pangan, sehingga pengembangan tanaman jarak yang paling sesuai untuk dilakukan yaitu di lahan marginal seperti di wilayah Indonesia Timur (Mulyani et al. 2006).
Walaupun tergolong tanaman yang tahan kekeringan, jarak juga dapat tumbuh dimana saja dengan cepat dan kuat, baik itu didataran rendah yang beriklim basah maupun sedang dan juga pada lahan yang tandus dan berbatu. Kelengasan tanah bukan menjadi faktor pembatas (misalnya irigasi atau curah hujan cukup merata), tetapi jarak yang hidup di daerah basah produksinya rendah karena bunganya mudah gugur. Jarak pagar dapat berproduksi sepanjang tahun, tetapi tidak dapat bertahan dalam kondisi tanah jenuh air. Iklim yang kering akan meningkatkan kadar minyak biji, akan tetapi kekeringan yang berkepanjangan menyebabkan gugur daun sehingga pertumbuhan tanaman menjadi terhambat (Mulyani et al. 2006).
3
sederhana yang kurang memadai tanpa memperhatikan mutu bahan tanaman. Tanaman jarak merupakan tanaman tahunan yang dapat berproduksi selama beberapa tahun, seyogyanya bahan tanaman yang digunakan adalah yang jelas asal-usulnya dan bermutu tinggi.
Pusat Penelitian dan Pengembangan Perkebunan telah mengembangkan tanaman jarak pagar yaitu IP-1A, IP-1M dan IP-1P. Ketiga IP tersebut masing-masing direkomendasikan sebagai sumber benih untuk daerah beriklim kering, sedang dan basah. Karena pengembangan jarak pagar diarahkan ke daerah marginal yang umumnya kering, maka diperlukan bahan tanam yang toleran kekeringan. Melalui penelitian ini diharapkan akan diperoleh informasi toleransi bibit beberapa genotipe jarak pagar yang toleran terhadap kekeringan berdasarkan respons morfologis dan ekofisiologisnya. Pemilihan bahan tanam untuk pengembangan tanaman jarak pagar di daerah tertentu dapat disesuaikan dengan keadaan iklimnya.
Tujuan Penelitian
Tujuan dari penelitian ini yaitu:
1. Mempelajari respon morfologis dan fisiologis bibit berbagai genotipe jarak pagar terhadap cekaman kekeringan
2. Mendpatkan informasi genotipe-genotipe yang toleran terhadap kekeringan.
Hipotesis
Adapun hipotesis yang diajukan dalam penelitian ini yaitu:
1. Kadar air media berpengaruh terhadap respon morfologis dan fisiologis bibit jarak pagar
2. Terdapat keragaman respon morfologis dan fisiologis bibit pada beberapa genotipe jarak pagar yang diamati
4
Bahan Æ
Perlakuan Æ
Target
Tujuan Akhir
Gambar 1 Kerangka pikir pengaruh berbagai genotipe dan cekaman kekeringan terhadap respon morfologis dan fisiologis bibit jarak pagar.
Enam Genotipe (tiga aksesi dan tiga populasi)
Tiga tingkat stress kekeringan (kadar air media yang berbeda)
RESPON MORFOLOGIS DAN FISIOLOGIS BIBIT BERBAGAI GENOTIPE JARAK PAGAR (Jatropha curcas L.) TERHADAP
CEKAMAN KEKERINGAN
Mengetahui keragaman morfologis dan fisiologis bibit pada berbagai genotipe jarak pagar pada kondisi cekaman kekeringan,
Variabel pengamatan
Parameter Morfologis ◙ Tinggi tanaman ◙ Luas daun ◙ Ketebalan daun
◙ Jumlah stomata terbuka ◙ Defisit air ◙ Laju asimilasi bersih ◙ Kandungan klorofil ◙ Kandungan karbon ◙ Kandungan prolin
- Untuk mempelajari respon morfologis dan fisiologis bibit berbagai genotipe jarak pagar terhadap cekaman kekeringan
TINJAUAN PUSTAKA
Tanaman Jarak Pagar
Jarak pagar diperkirakan berasal dari Amerika Tengah, khususnya
Meksiko. Di daerah tersebut, tanaman tumbuh secara alami di kawasan
hutan pinggiran pantai. Di Afrika dan Asia, jarak pagar hanya ditemukan
sebagai tanaman pagar atau pembatas lahan pertanian. Jarak pagar
menyebar di Malaka setelah tahun 1700 dan di Filipina sebelum tahun
1750. Di Malaka, jarak pagar disebut sebagai Dutch castor oil dan di Jawa
sebagai Chinese castor oil. Di Afrika dan Asia, jarak pagar disebut sebagai
castor oil plant yang menunjukkan bahwa tanaman ini ditanam untuk
diambil minyaknya. Sebutan jarak sebagai hedge castor oil plant
menunjukkan bahwa tanaman ini biasanya ditanam sebagai pagar.
Penyebaran jarak pagar di Thailand terjadi lebih dari dua abad yang lalu
oleh saudagar-saudagar Portugis yang menggunakan biji jarak untuk
membuat sabun cuci dan lainnya. Di Thailand terdapat lima spesies jarak
pagar yaitu J. curcas, J. gossypifolia, J. multifida, J. integrrima, dan J.
podagrica (Mulyani et al. 2006).
Tidak ada catatan yang pasti kapan jarak pagar masuk ke wilayah
Indonesia, tetapi diperkirakan bersamaan dengan di Malaysia. Jarak pagar
dapat ditemukan di berbagai tempat, namun umumnya tumbuh di pagar
atau tepi jalan di pedesaan. Jarak pagar dikenal dengan berbagai nama
daerah, antara lain nawaih nawas di Aceh, jarak wolanda di Manado, jirak
di Minangkabau, jarak kosta di Jawa Barat, jarak budeg, jarak gundul,
jarak iri, jarak pager, jarak cina, kaleke di Madura, jarak pageh di Bali,
tangang-tangang kali kanjoh di Makassar, malate (hoti) di Seram Timur,
dan balacai di Maluku Utara (Mulyani et al. 2006).
Tanaman jarak pagar dapat menyebar luas dan tumbuh subur di
daerah yang cocok sebagai tempat tumbuhnya yaitu di dataran rendah
hingga ketinggian 300 m dpl. Namun, sebaran tumbuh dapat mencapai
6
lahan kritis di Indonesia. Wilayah yang sesuai untuk tempat
pengembangan jarak pagar adalah di wilayah Indonesia Timur, dengan
curah hujan 200-1500 mm per tahun (Prawitasari et al. 2006).
Tanaman jarak pagar termasuk famili Euphorbiaceae, satu famili
dengan karet dan ubi kayu. Tanaman jarak berbentuk perdu dengan tinggi
1-7 m, bercabang tidak teratur. Batangnya berkayu, silindris dan bila
terluka mengeluarkan getah. Tanaman jarak pagar berakar tunggal yang
mampu menahan air dan tanah sehingga tahan terhadap kekeringan,
selain itu juga berfungsi sebagai tanaman penahan erosi. Daun tersebar di
sepanjang batang, permukaan atas dan bawah daun berwarna hijau
dengan bagian bawah lebih pucat dibandingkan permukaan atas.
Daunnya lebar dan berbentuk jantung atau bulat telur melebar dengan
panjang 5-15 cm. Tulang daun menjari dengan jumlah 5-7 tulang daun
utama. Daunnya dihubungkan dengan tangkai daun. Panjang tangkai
daun antara 4-15 cm (Prawitasari et al. 2006).
Tanaman jarak mempunyai bunga majemuk berbentuk malai,
berwarna kuning kehijauan, berkelamin tunggal dan berumah satu
(monoecious), yaitu bunga jantan dan bunga betina terdapat pada satu
tanaman dan tersusun dalam rangkaian berbentuk cawan yang tumbuh
diujung batang atau ketiak daun. Bunga betina lebih besar dari bunga
jantan terdiri atas ovari (bakal buah) yang beruang lima. Tangkai putik
lepas atau melekat pada pangkal, kepala putik terpecah tiga, berwarna
coklat. Buahnya berbentuk elips dengan panjang satu cm, memiliki dua
hingga tiga biji dengan kadar minyak dalam inti biji 54,2% atau sekitar
31,5% dari berat total biji. Pada umur lima bulan sudah mulai berbuah dan
produktivitas tertinggi dicapai ketika tanaman berumur lima tahun. Umur
produktif jarak pagar mencapai 50 tahun (Syah 2005).
Respon Morfologis Tanaman terhadap Cekaman Kekeringan Pada tanaman, cekaman kekeringan merupakan istilah untuk
menyatakan bahwa tanaman mengalami kekurangan suplai air akibat
keterbatasan air dari lingkungannya yaitu media tanam. Menurut Morgan
7
fluktuasi kelembaban udara, radiasi matahari yang diterima tanaman
cukup tinggi sampai pada lahan bermasalah yang mengalami defisit air,
dan kelembaban udara sangat rendah di lingkungan yang kering.
Kekurangan air secara internal pada tanaman berakibat langsung pada
penurunan pembelahan dan pembesaran sel. Pada tahap pertumbuhan
vegetatif, air digunakan oleh tanaman untuk pembelahan dan pembesaran
sel yang terwujud dalam pertambahan tinggi tanaman, perbanyakan daun
dan pertumbuhan akar (Kramer 1969).
Menurut Levitt (1980) dan Bray (1997), cekaman kekeringan yang
biasa disebut drought stress pada tanaman dapat disebabkan dua hal
yaitu (1) kekurangan suplai air di daerah perakaran dan (2) permintaan air
yang berlebihan oleh daun akibat laju evapotranspirasi melebihi laju
absorpsi air walaupun keadaan air tanah cukup tersedia. Menurut Fitter
dan Hay (1991), keadaan cekaman air menyebabkan penurunan turgor
pada sel tanaman dan berakibat pada menurunnya proses fisiologi.
Potensial turgor akan menurun hingga dapat mencapai nol dan
mengakibatkan kelayuan jika kehilangan air dari tanaman ini berlangsung
terus-menerus di luar batas kendalinya (Naiola 1996).
Hasil penelitian Pangaribuan (2001) pada tanaman kelapa sawit
menunjukkan bahwa kadar air media mengakibatkan perbedaan yang
nyata terhadap tinggi tanaman. Bibit kelapa sawit pada umur 20 minggu
setelah ditanam pada kondisi media 75% KL sebesar 48.80 cm lebih
tinggi dibandingkan bibit yang ditanam pada media 25% KL yaitu 34.36
cm. Demikian juga luas daun pada kondisi media yang rendah 25% KL
menurun sebesar 55.5% cm2.
Wijana (2001) melakukan penelitian yang sama pada tanaman
kelapa sawit menyatakan bahwa, pada kondisi kadar air tanah sekitar
10,6% dapat menyebabkan terjadinya cekamam kekeringan. Pengaruh
cekaman kekeringan lebih lanjut akan menyebabkan pengurangan luas
daun sekitar 0,022% yang erat kaitannya dengan penurunan potensial air
daun. Penurunan tersebut menyebabkan luas permukaan fotosintesis
8
Keadaan yang sangat kering pada tanaman akan dapat
mempengaruhi fase pertumbuhan dan produksi tanaman. Bila keadaan
kering terjadi selama fase vegetatif maka akan berpengaruh terhadap luas
daun dan panjang batang sehingga dapat menurunkan laju fotosintesis.
Boyer (1970) menyatakan bahwa menurunnya laju fotosintesis pada
tanaman kedelai yang mengalami kekeringan terutama disebabkan oleh
meningkatnya resistensi stomata terhadap CO2, sedangkan menurunnya
fotosintesis secara langsung pada tanaman yang mengalami kekeringan
juga akibat protoplasma dan kloroplas mengalami dehidrasi sehingga
mempunyai kemampuan yang rendah untuk proses fotosintesis.
Pada kondisi kekeringan, stomata daun menutup atau menutup
sebagian dan mengurangi aktivitasnya, sehingga menghambat masuknya
CO2 didalam ruang interseluler daun yang secara langsung mengurangi
aktivitas fotosintesis. Kekurangan air pada tanaman yang menghambat
terjadinya proses fotosintesis juga diteliti oleh Gerik et al. (1996) yang
telah membuktikan bahwa kekurangan air pada tanaman kapas sangat
berpengaruh terhadap kapasitas fotosintesis. Terjadi penurunan kapasitas
fotosintesis dan peningkatan penuaan daun yang berpengaruh buruk
terhadap produksi kapas. Pengaruh negatif lainnya akibat kekurangan air
adalah terjadinya penurunan pertumbuhan dan pembesaran sel,
perluasan daun, translokasi, dan transpirasi tanaman. Perluasan daun
pada 5 hari cekaman memiliki luas daun sekitar 20.4 cm2, setelah mengalami cekaman yang lebih lanjut sekitar 9 hari memiliki luas daun
yang lebih kecil yaitu 16.5 cm2.
Tardieu dalam Sopandie (2006) mengatakan ada dua pendekatan
utama yang sering digunakan untuk melihat kemampuan tanaman dalam
menghadapi cekaman kekeringan. Pendekatan pertama adalah dengan
melihat kemampuan pengambilan air secara maksimal dengan perluasan
dan kedalaman sistem perakaran. Pendekatan kedua dengan melihat
kemampuan tanaman mempertahankan turgor melalui penurunan
potensial osmotik, mengingat tekanan turgor mutlak diperlukan bagi
9
Dedywiryanto (2006) yang melakukan penelitian pada kelapa sawit,
menunjukkan bahwa, ketersediaan air tanah yang semakin rendah akan
mengakibatkan ketersediaan air bagi tanaman semakin berkurang,
sementara proses metabolisme dan transpirasi masih terus berlangsung,
sehingga mengurangi jumlah air yang tersimpan pada tajuk tanaman yang
dapat menyebabkan terjadinya penurunan kadar air daun pada tanaman
yang ditanam didalam media 25% KL. Hal yang sama juga diteliti oleh
Pangaribuan (2001) menunjukkan bahwa, pada kondisi kekurangan air,
bobot kering akar tanaman kelapa sawit menurun setelah mengalami
perlakuan cekaman pada 25% KL sekitar 4.57 g dari 9.22 g (100% KL).
Respon Fisiologis Tanaman terhadap Cekaman Kekeringan
Prolin merupakan salah satu metabolit yang terbentuk dan
terakumulasi pada daun dalam jumlah yang lebih banyak apabila tanaman
mengalami cekaman kekeringan. Peningkatan kadar prolin pada awal
terjadinya kekeringan relatif lambat dan meningkat dengan cepat setelah
tanaman mengalami kekeringan lebih lanjut. Prolin yang terakumulasi
didalam sitoplasma berperan memelihara keseimbangan air antara
vakuola, sitoplasma dan dengan lingkungan (Yang dan Kao 1999).
Prolin mempunyai salah satu fungsi yaitu untuk melindungi dari
denaturasi protein, sebagai sumber energi dari kelompok amino dan
merupakan protektan bagi enzim akibat pengaruh toksik biologi seperti
urea. Oleh karena itu prolin dikenal sebagai salah satu osmoprotektan
(Notle et al. 1997). Sebagai osmoprotektan, prolin diduga sangat terlibat
dalam osmoregulasi, menjaga kelarutan protein, kestabilan membran
fosfolipid dan juga sebagai sumber cadangan karbon, nitrogen dan energi
(Walton et al. 1998).
Menurut Levitt (1980) peranan prolin tidak hanya terbatas pada
penyesuaian osmotik yang dikaitkan dengan status air, tetapi juga
menjaga peranan lain seperti menetralisir pengaruh toksik NH3 hasil
hidrolisis protein, sebagai sumber energi dan sumber N bagi pemulihan
10
Menurut Yang dan Kao (1999) prolin merupakan salah satu asam amino
yang di sintesis dan diakumulasi pada berbagai jaringan tanaman yang
dicekam kekeringan, terutama pada bagian daun. Menurut Yoshida et al.
(1997) prolin dijumpai terakumulasi lebih banyak pada tanaman yang lebih
toleran terhadap cekaman kekeringan dibandingkan dengan tanaman
yang peka.
Bates et al. (1973) mengatakan bahwa kandungan prolin pada
tanaman meningkat secara proporsional lebih cepat dibandingkan dengan
asam amino lain pada kondisi cekaman kekeringan. Indikasi ini dapat
dimanfaatkan sebagai suatu tolok ukur untuk mengevaluasi
varietas-varietas yang tahan terhadap kondisi kekeringan. Prolin disintesis dan
diakumulasi dari asam glutamat serta diduga selama cekaman kekeringan
prolin berfungsi sebagai cadangan makanan. Wijana (2001) yang
melakukan penelitian pada kelapa sawit mengatakan bahwa prolin
mengalami peningkatan setelah tanaman dicekam kekeringan sejalan
dengan penurunan kandungan air daun.
Informasi mengenai karakter toleransi tanaman terhadap
cekaman kekeringan berdasarkan karakteristik morfofisiologi sudah cukup
banyak dilaporkan pada tanaman pangan seperti jagung (Frederique et al.
1998). Pada genotipe kopi arabika (Coffea arabica) dan kopi robusta
(Coffea canophora) yang toleran, perlakuan cekaman kekeringan
menyebabkan terjadinya peningkatan akumulsi prolin dua kali lebih
banyak dibandingkan dengan tanpa perlakuan kekeringan (Maestri et al.
1995). Hal yang sama juga dinyatakan oleh Hanson et al.(1986) bahwa
secara keseluruhan tanaman dapat menunjukkan tingkat toleransi
terhadap cekaman kekeringan melalui penekanan proses fisiologis antara
lain pengurangan dehidrasi sel, misalnya rendahnya konduktan stomata
terhadap penguapan air, tekanan osmotik atau osmoregulasi dan laju
fotosintesis bersih.
Penelitian Chirtine et al. (1996) membuktikan bahwa konsentrasi
asam amino pada tanaman alfalfa meningkat (1.8 kali) ketika potensial air
11
tanaman untuk menghasilkan asam amino bervariasi untuk setiap
potensial air yang berbeda. Konsentrasi asam amino pada tanaman alfalfa
ini meningkat sejalan dengan menurunnya potensial air.
Konsentrasi prolin yang dihasilkan mencapai 150 mM pada
tanaman alfalfa untuk merespon penurunan potensial air tanah sebesar
-1.0 sampai -2.0 MPa. Diketahui juga bahwa dari beberapa asam amino
yang terdapat pada tanaman alfalfa tersebut, prolin merupakan asam
amino yang paling berfluktuasi sehubungan dengan adanya perbedaan
potensial air, sedangkan asparagin yang merupakan asam amio utama
dengan kandungan 75% pada tanaman alfalfa, tidak menunjukkan respon
terhadap perubahan potensial air, sehingga kadarnya tetap tinggi baik
dalam keadaan tanpa cekaman kekeringan maupun dalam keadaan
tercekam (Chirtine 1996).
Cekaman kekeringan mempengaruhi berbagai aspek pertumbuhan
tanaman. Pada tanaman yang layu akibat kekeringan, terjadi peningkatan
hidrolisi protein yang dibarengi dengan peningkatan asam amino (Barnett
dan Naylor 1966). Kekurangan air akan menurunkan kandungan protein
larut dan kandungan asam amino arginin, sebaliknya kandungan asam
amnino prolin meningkat dalam jumlah yang banyak (Kramer 1969).
Produksi total fotosintat dtentukan oleh interaksi antara laju
fotosintesis dan luas daun atau permukaan fotosintetik (Sinha ey al. 1982).
Kekeringan akan mengakibatkan penurunan produksi fotosintat bersih,
oleh karena reduksi luas daun dan peningkatan konduktansi stomata
(Kramer 1969). Penurunan produksi bahan kering atau produksi fotosintat
tersbut disebabkan rendahnya laju fotosintesis, sebagaimana dijelaskan
oleh Mayoral et al. (1981) bahwa rendahnya laju fotosintesis akibat
kekeringan, disebabkan berkurangnya kandungan klorofil daun dan
terhambatnya aktivitas enzim-enzim yang terlibat dalam fotosintesis, yaitu
enzim RuBP karboksilase dan PEP karboksilase.
Cekaman air dapat mempengaruhi perangkat fotosintesis yaitu
menurunkan kandungan klorofil dalam kloroplas, mesofil pada sel yang
12
kandungan klorofil juga yang diteliti oleh Yusnaeni (2002) pada tanaman
Hoya (Asclepiadaceae) yang mengatakan bahwa, kandungn klorofil
menurun sekitar 0.46 mg/g daun segar (penyiraman setiap minggu) jika
dibandingkan dengan penyiraman setiap hari yang memiliki klorofil sekitar
0.54 mg/g daun segar.
Fotosintesis pada hakekatnya merupakan satu-satunya mekanisme
masuknya energi ke dalam dunia kehidupan. Sebagaimana reaksi
oksidasi penghasil energi, yaitu tempat bergantungnya semua kehidupan,
maka fotosintesis meliputi reaksi oksidasi dan reduksi. Proses
keseluruhan adalah pemindahan elektron (oksidasi) disertai dengan
pelepasan O2 sebagai hasil samping dan reduksi CO2 untuk membentuk
senyawa organik, misalnya karbohidrat (Salisbury dan Ross 1992).
BAHAN DAN METODE
Tempat dan Waktu Peneltian
Penelitian ini dilaksanakan di rumah kaca Laboratorium Ilmu dan Teknologi Benih Leuwikopo, analisis morfologid dan ekofisiologis dilakukan di Laboratorium Ekofisiologi dan RGCI. Departemen Agronomi dan Hortikultura Fakultas Pertanian, Institut Pertanian Bogor. Penentuan luas daun dengan leaf area meter di Laboratorium Ekofisiologi Tanaman, BALITRO. Cimanggu. Penelitian berlangsung pada bulan Juli sampai dengan November 2007.
Bahan dan Alat
Bahan tanaman yang digunakan pada penelitian ini adalah benih jarak pagar yang terdiri atas tiga populasi yaitu Pakuwon (IP-1P), Muktiharjo (IP-1M) dan Asembagus (IP-1A). Ketiganya masing-masing direkomendasikan sebagai sumber benih untuk daerah basah, sedang dan kering yang merupakan hasil seleksi dari Puslitbang Perkebunan. Selain itu juga benih jarak pagar yang belum terseleksi terdiri atas tiga aksesi yaitu Sukabumi, Karanganyar dan NTB yang masing-masing mewakili daerah basah, sedang dan kering. Media tumbuh yang digunakan adalah tanah lapisan atas dan pasir dengan perbandingan 3:1 berdasarkan volume. Tanaman dipupuk dengan menggunakan pupuk Urea dengan dosis 1.3 g/4 kg media. Bahan kimia untuk analisis prolin dan kandungan klorofil adalah asam asetat glasial, asam fosfat, asam sulfosalisilat, ninhidrin, toluen dan aseton.
14
Metode Penelitian Rancangan Percobaan
Penelitian ini disusun secara petak terbagi (split plot) menggunakan rancangan acak kelompok (RAK) dengan dua faktor, yaitu:
1. Faktor pertama, kadar air media (KAM) sebagai petak utama, yang terdiri atas tiga taraf:
K1 : Kadar air media 40 %
K2 : Kadar air media 36 %
K3 : Kadar air media 32 %
2. Faktor kedua, genotipe yang terdiri dari aksesi dan populasi sebagai anak petak, sebagai berikut
A1 : Karanganyar A4 : IP-1P
A2 : Sukabumi A5 : IP-1M
A3 : NTB A6 : IP-1A
Dengan demikian, secara keseluruhan diperoleh 18 kombinasi perlakuan. Tiap perlakuan diulang tiga kali, sehingga terdapat 54 satuan percobaan. Tiap percobaan terdiri dari 8 bibit, maka jumlah bibit yang digunakan sebanyak 432 bibit jarak pagar. Data yang diperoleh dianalisis dengan uji F menggunakan program SAS (Statistical Analysis System) dan jika berpengaruh nyata secara statistik (pada α = 5%) maka dilakukan uji lanjut menggunakan Duncan Multiple Range Test (DMRT)
Model rancangan percobaan yang digunakan adalah sebagai berikut: Yijk = µ + Rk+ Pi + δik + Kj+ (PK)ij + ijk
dimana :
Yijk = nilai pengamatan pada perlakuan cekaman kekeringan ke-i, dan
perlakuan genotipe ke-j dan ulangan ke-k. µ = nilai rataan umum
Rk = pengaruh ulangan ke-k
Pi = pengaruh perlakuan cekaman kekeringan pada taraf ke-i δik = pengaruh galat pada perlakuan cekaman kekeringan ke-i dan ulangan ke-k
Kj = pengaruh perlakuan genotipe pada taraf ke-j
PKij = pengaruh interaksi antara cekaman kekeringan pada taraf ke-i dan
genotipe ke-j.
ijk = pengaruh galat pada perlakuan cekaman kekeringan pada taraf
15
Pelaksanaan Penelitian
Persemaian Benih
Benih jarak pagar dikecambahkan terlebih dahulu pada bedengan persemaian dengan media semai terdiri dari tanah lapisan atas dan pasir yang terletak di dalam rumah kaca. Pemeliharaan dilakukan dengan melakukan penyiraman setiap hari pada pagi dan sore hari. Kecambah dipelihara dalam persemaian selama 21 hari, sebelum dipindahkan ke pembibita yang berukuran 20 cm x 20 cm dan dipelihara pada kondisi yang optimum selama 2 bulan. Selanjutnya bibit yang dipindahkan ke polybag yang berukuran 35 cm x 35 cmyang berisi 4 kg tanah. Sebelum dipindahkan bibit dikondisikan pada kondisi yang sub optimum (selama 3 hari tidak di lakukan penyiraman).
Penyiapan Media Tanaman
Mula-mula campuran tanah dikeringanginkan dan diayak dengan menggunakan ayakan berdiameter lubang ± 5 mm untuk membuang sisa-sisa kayu, akar dan batu. Selanjutnya tanah dicampur pasir dengan perbandingan 3:1 berdasarkan volume, kemudian dicampur pupuk urea sebanyak 1.3 g/4 kg tanah. Selanjutnya media tanam dimasukkan kedalam polibag dengan ukuran 35 x 35 cm, masing-masing diisi 4 kg tanah kering angin.
Penetapan Kapasitas Lapang dan Titik Layu Permanen pada Media Tanam
Penetapan kapasitas lapang dan titik layu permanen pada media tanam dilakukan dengan menggunakan alat ”Pressure Plate Apparatus” dan ”Pressure Membran Apparatus” masing-masing pada pF 2.54 dan pF 4.20. Untuk penetapan kapasitas lapang dan titik layu permanen digunakan tanah kering udara yang digunakan untuk media tanam.
16
contoh tanah ini disiram air sampai berlebihan dan dibiarkan selama 48 jam. Alat ditutup rapat-rapat, kemudian diberi tekanan sesuai dengan pF yang dikehendaki (untuk pF 2.54 dengan tekanan 1/3 bar dan 1.5 bar untuk pF 4.20). Keseimbangan tercapai kira-kira 48 jam setelah diberi tekanan. Kemudian contoh tanah dikeluarkan dan ditetapkan kadar airnya dengan metode gravimetri, dengan menggunakan rumus:
KA = X100%. Dari hasil perhitungan didapatkan kadar air pada
kondisi tanah kapasitas lapang yaitu 40.35 % dan titik layu permanen adalah 25.56 % dengan demikian selang kadar air tersedia adalah selisih antara kapasitas lapang dan titik layu permanen yaitu 14.79 %. Kadar air tersedia ini digunakan untuk menentukan level cekaman kekeringan dalam penelitian. Hasil penentuan level cekaman diperoleh K1 (KL) = kadar air media 40% , K2 = kadar air media 36% dan K3 = kadar air media 32%. Kadar air media tanaman pada level cekaman tertinggi yaitu 25.56 + (0.5 x 14.79 ) = maka diperoleh 32% kadar tersedia.
Banyaknya air yang harus ditambahkan pada awal percobaan dihitung sebagai berikut :
a. Untuk menghitung bobot tanah kering angin (X) berdasarkan bobot tanah kering mutlak digunakan rumus sebagai berikut :
Bobot tanah kering mutlak (BKM) =
X = BKM x (1 + K.A tanah kering angin) X = 5 x (1+37,37) = 191. 85
b. Untuk menghitung bobot tanah kapasitas lapang (Y) berdasarkan bobot tanah kering mutlak digunakan rumus sebagai berikut :
Bobot tanah kering mutlak (BKM) =
Y = BKM x (1 + K.A tanah kapasitas lapang) Y = 5 x (1+40) = 205
c. Banyaknya air yang harus ditambahkan untuk contoh tanah 5000 g adalah Y – X = 205 – 191.85 = 13.15.
1 + kadar air tanah kering angin Bobot tanah kering angin (X)
1 + kadar air tanah kapasitas lapang Bobot tanah kapasitas lapang (Y)
17
d. Berdasarkan perhitungan pada butir a, b dan c, maka banyaknya air yang harus ditambahkan sesuai perlakuan cekaman adalah:
Kadar air media pada kondisi KL K1, 40 % yaitu 13.15 x 40 = 526.0 ml
Kadar air media pada level cekaman K2, 36 % yaitu 13.15 x 36 = 473.4 ml
Kadar air media pada level cekaman K3, 32 % yaitu 13.15 x 32 = 420.8 ml
Masing-masing media K1, K2 dan K3 ditimbang untuk menentukan bobot awal masing-masing media dan untuk mempertahankan kondisi masing-masing kadar air media selama penelitian, penambahan air setiap hari dikembalikan ke bobot awal media tanam diatas timbangan.
Koreksi terhadap pertambahan bobot tanaman, dilakukan dengan menimbang bobot tanaman sesuai kombinasi perlakuan setiap 2 minggu sekali dengan menggunakan sampel tidak tetap yang disediakan khusus untuk koreksi bobot basah tanaman.
Indeks sensitivitas kekeringan
Indeks sensitivitas kekeringan (IS) bertujuan untuk menentukan genotipe kedalam tingkat toleransi cekaman.
Indeks sensitivitas kekeringan (IS), dihitung menggunakan Rumus yang dikemukakan oleh Fischer dan Maure dalam Diah (2005), yaitu
IS = Keterangan :
IS = Indeks sensitivitas kekeringan
Y = Nilai respon genotipe kondisi stres kekeringan (KAM 32 %) Yp = Nilai respon genotipe kondisi non stres kekeringan (KAM 40%) X = Nilai respon rata-rata enam genotipe pada kondisi stres kekeringan
(KAM 32 %)
XP = Nilai respon rata-rata enam genotipe pada kondisi non stres kekeringan (KAM 40%)
(1-Y/Yp)
18
Kriteria untuk menentukan tingkat sensitivitas suatu genotipe terhadap stres kekeringan adalah jika :
IS ≤ 0.5 = toleran 0.5 < IS ≤1.00 = agak toleran IS > 1.00 = peka (sensitif)
Penanaman Bibit
Bibit yang digunakan adalah bibit hasil perkecambahan dari masing-masing genotipe pada polibag 1 kg yang berukuran 2 cm x 2 cm yang telah berumur 2 bulan. Bibit dipindahkan pada media tanam yang telah diisi tanah 4 kg pada polybag berukuran 5 kg. Pemindahan dengan cara bibit dibuat stress selama 3 hari, agar bibit dapat beradaptasi dengan perlakuan yang diberikan (cekaman kekeringan) yaitu dengan cara menyobek polybag 1 kg, kemudian media beserta tanaman pada 1 kg dipindahkan pada polybag yang telah terisi tanah 4 kg agar tidak merusak media tanam berupa akar tanaman agar tidak terganggu atau rusak. Media tanam disusun dirumah kaca sesuai dengan perlakuan yang dilakukan sebelumnya.
Pengamatan
19
Respon morfologis yang diamati adalah sebagai berikut:
1. Tinggi tanaman. Diukur dari pangkal batang (batas leher akar) sampai titik tumbuh. Pengukuran dilakukan dengan menggunakan standar mistar besi dan dilakukan pada umur 0, 2, 4, 6 dan 8 MSP.
2. Luas daun. Pengamatan luas daun dilakukan pada daun yang terletak pada buku ke-3. Pengamatan dilakukan pada pagi hari dan berumur 0, 2, 4, 6 dan 8 MSP. Luas daun ditentukan dengan menggunakan leaf area meter di Laboratorium Ekofisiologi Tanaman, BALITRO. Cimanggu.
3. Ketebalan daun. Diukur menggunakan preparat segar. Pengamatan dan pengukuran daun yang telah diiris tipis dilakukan dengan menggunakan mikroskop pada skala mikrometer pada daun paling muda yang telah membuka sempurna.
4. Jumlah stomata terbuka. Pengamatan jumlah stomata dilakukan pada umur 1, 4 dan 7 minggu setelah perlakuan. Dihitung dari preparat yang telah dibuat dari permukaan epidermis bawah. Preparat dibuat dengan cara melapisi permukaan bawah daun jarak dengan menggunakan larutan cutex pucat dan menempelkan lapisan cutex pucat tersebut pada gelas objek menggunakan selatip. Jumlah stomata yang diperiksa dengan menggunakan mikroskop perbesaran 10 kali dengan luas jaringan yang diamati berdiameter 2 mm.
20
sehingga didapatkan bobot kering (bk). Defisit air dihitung dengan rumus sebagai berikut:
6. Bobot kering pucuk. Tanaman dibersihkan dan dipotong hingga batas leher akar, kemudian bobot kering tajuk ditimbang setelah dikeringkan dalam oven selama 72 jam dengan suhu 80oC diukur pada akhir pengamatan.
7. Bobot kering akar. Bobot kering akar diukur pada akhir pengamatan ditimbang setelah dikeringkan dalam oven selama 72 jam dengan suhu 80oC diukur pada akhir pengamatan.
8. Rasio pucuk akar. Rasio pucuk akar ditentukan dengan membandingkan berat kering pucuk dengan berat kering akar tanaman yang dilakukan pada akhir pengamatan.
9. Panjang akar. Panjang akar yang diukur dari pangkal batang sampai ujung akar terpanjang dilakukan pada akhir pengamatan. 10. Volume Akar. Volume akar diukur pada akhir pengamatan
dengan menggunakan gelas ukur 1000 ml yang telah diisi air hingga volume tertentu, kemudian dimasukkan akar yang telah dipotong ke dalam gelas ukur. Pertambahan volume yang ditunjukkan skala pembacaan pada gelas ukur saat seluruh akar terendam dalam air adalah volume akar yang terukur.
Respon fisiologis tanaman, terdiri atas:
1. Kadar air daun. Pengamatan Kadar air daun dilakukan pada umur 1, 4 dan 7 minggu setelah perlakuan. Diukur dengan cara menimbang daun segar (bobot segar = bs), selanjutnya daun dikeringkan dengan oven (pada 80oC) selama 48 jam (bobot kering = bk). Kadar air daun dihitung dengan rumus: Kadar air daun
21
2. Laju asimilasi bersih. Laju asimilasi bersih dilakukan pada umur 1, 4 dan 7 minggu setelah perlakuan yang dihitung dengan menggunakan rumus yang dikembangkan oleh Sitompul dan Guritno (1995):
3. Kandungan klorofil. Analisis kandungan klorofil daun dilakukan dengan cara: klorofil diekstrak menggunakan metode baku yang dikembangkan oleh Arnon dalam Ardie (2006) yaitu 1 g jaringan daun segar ditimbang dan digerus hingga halus dalam mortar, Kemudian ditambahkan aseton 10 ml sehingga jaringan menjadi homogen. Jaringan tersebut diaduk hingga menghasilkan supernatan lalu ditambahkan aseton 80%. Suspensi dituangkan kedalam tabung sentrifius dan diputar selama 10 menit dengan kecepatan 5000 rpm. Secara perlahan supernatan dituangkan kedalam tabung sentrifius yang lain dan volume dibuat 3 ml. kemudian dilakukan pengukuran absorban dengan menggunakan spectrofotometer Spectonic 21 DV Bausch 7 Lomb pada panjang gelombang 663 nm dan 645 nm atau 652 nm. Penetapan konsentrasi klorofil (mg/g) didasarkan pada metode McKinney and Arnon dalam Ardie sebagai berikut:
Konsentrasi klorofil a = 12. 7 A663 – 2.69 A645
c = volume ekstrak yang diambil dari ekstrak awal d = konversi dari liter ke ml
22
3. Kandungan karbon. Analisis kandungan karbon daun dilakukan dengan cara: daun dikeringkan selama 2 x 24 jam pada suhu 50oC kemudian digerus hingga halus. Ditimbang 1 g contoh dalam cawan porselin, lalu diabukan pada suhu 600oC sekitar 5 jam dengan menggunakan tanur, setelah itu diangkat dan didinginkan, selanjutnya ditimbang lagi dalam keadaan kering, dan dicari kadar abunya. Dengan menggunakan rumus sebagai berikut:
% karbon
4. Kandungan prolin. Analisis kandungan prolin daun dilakukan berdasarkan metode Bates et al. (1973). Mula-mula potongan daun ditimbang sebanyak 0.5g, kemudian dihaluskan ditambah 5 ml asam sulfosalisilat 3%, diaduk dan disentrifuge (600 rpm selama 3 menit) kemudian diambil supernatannnya, residunya ditambah lagi dengan 4 ml asam sulfosalisilat 3%, diaduk dan disentrifuge kembali, kemudian supernatan yang diperoleh dicampur dengan supernatan sebelumnya. Supernatan yang telah diperoleh lalu ditepatkan/ditera hingga 10 ml. Sebanyak 2 ml supernatan contoh ditambah 2 ml asam ninhidrin dan 2 ml asam asetat glasial, dipanaskan pada penangas air suhu 100oC. Setelah 60 menit reaksi segera dihentikan dengan merendamnya di dalam es. Setelah dingin ditambah 4 ml toluen, diaduk dengan vortex selama 15 detik, dan didiamkan pada suhu kamar hingga fase toluen terpisah dari fase airnya. Absorban fase toluen dibaca dengan spektrofotometer pada panjang gelombang 520 nm dengan toluen sebagai blanko. Jumlah prolin dapat dihitung berdasarkan kurva standar (kurva standar cukup dibuat dengan lima titik, masing-masing 0, 200, 400, 600 dan 800 μg/ml). Kandungan prolin dapat dihitung berdasarkan persaman:
HASIL DAN PEMBAHASAN
Keadaan Umum Penelitian
Keadaan pertanaman pada umur 0 – 2 bulan menunjukkan pertumbuhan yang sehat, tingkat serangan hama dan penyakit relatif rendah. Namun setelah tanaman berumur lebih dari 2 bulan, terdapat berbagai serangan hama yang menyerang tanaman jarak pagar, yaitu belalang (Valanga spp), ulat daun, ulat bulu dan rayap. Akibat yang ditimbulkan oleh serangan belalang dan ulat daun tidak terlalu mengganggu pertumbuhan dan perkembangan tanaman. Belalang menimbulkan kerusakan dengan meningkatkan bekas gigitan yang berupa lubang-lubang pada daun. Serangan ulat dapat ditekan dengan pengendalian secara mekanis.
Serangan rayap cukup mengganggu pertumbuhan dan perkembangan tanaman bahkan ada tanaman yang mati akibat serangan rayap. Rayap menyerang akar tanaman selanjutnya batang tanaman, sehingga batang menjadi keropos dan akhirnya tanaman rebah dan mati. Gejala serangan lain adalah daun menguning dan ranting mengering.
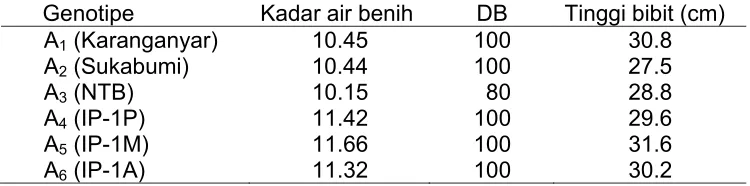
Tabel 1 menunjukkan bahwa kadar air benih masing - masing genotipe dengan genotipe A5 (IP-1M) paling tinggi yaitu 11.66% diikuti
oleh genotipe A4 (IP-1P) 11.42%, A6 (IP-1A), A1 (Karanganyar), A2
(Sukabumi) dan genotipe A3 (NTB) yang memiliki kadar air benih yang
lebih rendah yaitu 10.15%. Daya berkecambah benih yang digunakan dalam penelitian mencapai 100% untuk genotipe A1, A2, A4, A5 dan A6,
sedangkan genotipe A3 memiliki daya berkecambah hanya 80%.
Tabel 1 Kadar air benih, daya berkecambah dan tinggi bibit genotipe jarak pagar yang digunakan dalam penelitian
Genotipe Kadar air benih DB Tinggi bibit (cm) A1 (Karanganyar) 10.45 100 30.8
A2 (Sukabumi) 10.44 100 27.5
A3 (NTB) 10.15 80 28.8
A4 (IP-1P) 11.42 100 29.6
A5 (IP-1M) 11.66 100 31.6
24
Tinggi bibit pada saat perlakuan dimulai cukup bervariasi antar genotipe. Genotipe A5 (IP-1M) mempunyai tinggi bibit rata-rata sekitar
31.6 cm yang tertinggi diantara semua genotipe, sedangkan genotipe A2
(Sukabumi) yang terendah sekitar 27.5 cm.
Pengaruh Perlakuan terhadap Peubah yang diamati
Hasil pengamatan menunjukkan bahwa kadar air media berpengaruh sangat nyata terhadap tinggi tanaman, luas daun, ketebalan daun, jumlah stomata terbuka, bobot kering pucuk, bobot kering akar, panjang akar, volume akar, kadar air daun, laju asimilasi bersih dan kandungan karbon, berpengaruh nyata pada rasio pucuk akar dan kandungan klorofil dan tidak nyata terhadap defisit air dan kandungan prolin (Tabel 2).
Tabel 2 Rekapitulasi sidik ragam pengaruh kadar air media dan genotipe
terhadap karakter morfologis dan fisiologis tanaman jarak pagar (Jatropha curcas L.) pada akhir pengamatan
No Peubah KAM Genotipe KAMxGenotipe KK
A Karakter Morfologis:
Tinggi tanaman ** ** * 2.35
Luas daun ** ** tn 14.02
Ketebalan daun ** tn ** 4.73
Jumlah stomata terbuka ** tn tn 4.54
Defisit air tn tn tn 4.75
Bobot kering pucuk ** * tn 21.16
Bobot kering akar ** tn tn 21.06
Ratio pucuk akar * tn tn 31.89
Panjang akar ** ** tn 8.26
Volume akar ** ** ** 8.76
B Karakter Fisiologis:
Kadar air daun ** tn tn 5.30
Laju asimilasi bersih ** * tn 19.61
Kandungan klorofil * tn tn 11.46
Kandungan karbon ** tn tn 3.01
Kandungan prolin tn tn tn 4.87
Keterangan : * = nyata, ** = sangat nyata, tn = tidak nyata
25
jumlah stomata terbuka, defisit air, bobot kering akar, ratio pucuk akar, kadar air daun, kandungan klorofil, kandungan karbon dan kandungan prolin. Interaksi antara kadar air media dan genotipe hampir tidak berpengaruh pada semua karakter yang diamati kecuali terhadap tinggi tanaman, ketebalan daun dan volume akar.
Respon Morfologis Enam Bibit Genotipe Jarak Pagar terhadap Cekaman Kekeringan
Respon morfologis tanaman yang diamati adalah tinggi tanaman, luas daun, ketebalan daun, jumlah stomata terbuka, defisit air, bobot kering pucuk, bobot kering akar, rasio pucuk akar, panjang akar dan volume akar.
Tinggi Tanaman
Respon tinggi tanaman dipengaruhi secara nyata oleh interaksi kadar air media dan genotipe, tinggi tanaman yang dicapai semua genotipe pada kadar air media 36% lebih rendah daripada kadar air media 40%, kecuali genotipe A3 (NTB) (Tabel 3).
Tabel 3 Pengaruh interaksi antara kadar air media dan genotipe terhadap tinggi tanaman (cm) jarak pagar pada akhir pengamatan
Genotipe Kadar air media
40% 36% 32% A1 (Karanganyar) 44.5 ab 41.0 ef 41.1 ef
A2 (Sukabumi) 43.5 bcd 39.7 fg 37.8 h
A3 (NTB) 41 ef 39.8 fg 37.8 h
A4 (IP-1P) 43.8 abc 40.5 ef 39.8 fg
A5 (IP-1M) 45.5 a 42.1 cde 41.9 de
A6 (IP-1A) 45.1 ab 41.5 ef 38.5 gh
Ket: Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf DMRT 5%.
Walaupun genotipe A3 (NTB) memiliki tinggi tanaman yang paling
26
yang ditunjukkan oleh tinggi tanaman juga dilaporkan oleh Kartika (2006) pada tanaman kelapa sawit. Tinggi tanaman kelapa sawit menurun sebesar 35.5% bila diberi cekaman kekeringan (25% air tersedia).
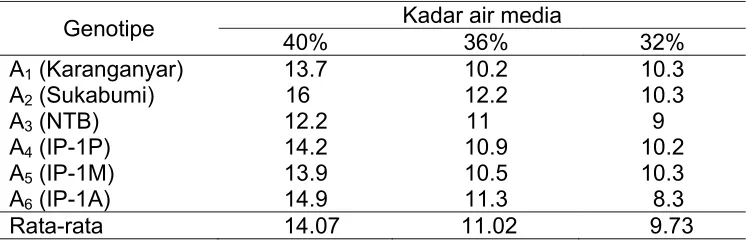
Tabel 4 menunjukkan rata-rata pertambahan tinggi tanaman pada kadar air media 36% dan 32% masing-masing berkisar antara 11.02 cm dan 9.73 cm, sedangkan rata-rata pertambahan tinggi tanaman pada kadar air media 40% sekitar 14.07 cm. Tinggi tanaman sangat bervariasi antar genotipe pada kadar air media 40%, tetapi pada kadar air media 36% pertambahan tinggi semua genotipe tidak terlalu bervariasi. Genotipe Sukabumi tumbuh paling cepat pada kondisi optimum (40%) dan kurang optimum (36%) sebagaimana ditunjukkan oleh pertambahan tinggi masing-masing sebesar 16 dan 12.2 cm selama 8 minggu pengamatan. Sebaliknya genotipe NTB paling lambat pertumbuhannya bahkan pada semua kadar air media. Pada kadar air media 40% pertambahan tinggi tanaman paling besar, diduga karena pembelahan, perbesaran dan pemanjangan sel berjalan dengan baik. Pada kadar air media 36% dan 32% diduga pembesaran dan pemanjangan sel terhambat karena terbatasnya air tersedia sehingga pertambahan tinggi tanaman lebih rendah daripada pada kadar air media 40%.
Tabel 4 Pertambahan tinggi tanaman selama 8 minggu setelah perlakuan pada masing-masing genotipe dan kadar air media
Genotipe Kadar air media
40% 36% 32%
A1 (Karanganyar) 13.7 10.2 10.3
A2 (Sukabumi) 16 12.2 10.3
A3 (NTB) 12.2 11 9
A4 (IP-1P) 14.2 10.9 10.2
A5 (IP-1M) 13.9 10.5 10.3
A6 (IP-1A) 14.9 11.3 8.3
Rata-rata 14.07 11.02 9.73
Luas daun
27
daun, namun penurunan kadar air media selanjutnya menjadi 32% menurunkan luas daun sekitar 12.62 cm2. Hal ini sesuai dengan penelitian yang dilakukan oleh Pangaribuan (2001) pada kelapa sawit, bahwa kelapa sawit yang diberi cekaman menurunkan luas daun 430.60 cm2 pada (25% KL) dari 922.59 cm2 (100% KL). Penelitian ini juga sesuai dengan yang dilakukan oleh Sasli (1999) pada tanaman kakao, bahwa kakao yang diberi cekaman kekeringan menurunkan luas daun sekitar 1642.50 cm2 (55% air tersedia) dari 2642.90 cm2 (100% air tersedia).
Tabel 5 Pengaruh kadar air media terhadap luas daun (cm2) tanaman jarak pagar pada akhir pengamatan
Perlakuan Nilai tengah
K1 (KAM 40 %) 36.34 a
K2 (KAM 36 %) 35.39 a
K3 (KAM 32 %) 23.72 b
Ket : Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf DMRT 5 %.
Luas daun tanaman menurun seiring dengan terjadinya peningkatan cekaman kekeringan. Pengurangan luas daun merupakan salah satu reaksi tanaman yang dapat dideteksi dari kekurangan kelembaban pada awal perkembangan vegetatif. Ukuran luas daun yang mengecil merupakan salah satu respon tanaman terhadap kekurangan air, hal ini merupakan mekanisme penghindaran tanaman untuk menekan kehilangan air, yaitu dengan mengurangi terjadinya transpirasi pada tanaman. Semakin mengecil luas daun, berarti menurunkan intersepsi cahaya matahari, yang menyebabkan luas bidang permukaan yang mengalami proses fotosintesis menyempit. Hal ini diduga menyebabkan terhambatnya pertumbuhan tanaman pada penelitian ini. Menurut Lakitan (1995), jika kadar air diturunkan 90%, maka pembesaran sel daun menjadi terhambat dan pembesaran sel daun akan terhenti sama sekali jika kadar air turun 70 -75 %.
Tabel 6 menunjukkan bahwa diakhir pengamatan genotipe A5
(IP-1M) memiliki luas daun yang paling besar sekitar 36.1 cm2, tidak berbeda dengan genotipe A1 (Karanganyar) dan A6 (IP-1A), tetapi berbeda dengan
28
paling rendah sekitar 28.9 cm2 sama dengan genotipe A2 (Sukabumi), A4
(IP-1P) dan A6 (IP-1A).
Tabel 6 Pengaruh genotipe terhadap luas daun (cm2) tanaman jarak pagar pada akhir pengamatan.
Genotipe Nilai tengah
A1 (Karanganyar) 34.5 ab
A2 (Sukabumi) 28.9 c
A3 (NTB) 28.9 c
A4 (IP-1P) 30.3 bc
A5 (IP-1M) 36.1 a
A6 (IP-1A) 32.2 abc
Ket : Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf DMRT 5 %.
Ketebalan daun
Peningkatan ketebalan daun merupakan salah satu bentuk pertumbuhan tanaman yang merupakan hasil dari aktivitas pembelahan dan pemanjangan sel (Salisbury dan Ross 1995). Pembelahan dan pemanjangann sel tanaman yang dipengaruhi oleh ketersediaan unsur hara. Respon ketebalan daun sangat dipengaruhi oleh interaksi kadar air media dan genotipe. Genotipe A3 (NTB) memiliki daun yang paling tebal
sebesar 2.25 mm pada perlakuan kadar air media 40% dan tidak berbeda dengan genotipe A2 (Sukabumi) dan genotipe A4 (IP-1P), sedangkan
genotipe A5 (IP-1M) memiliki ketebalan daun yang paling tipis yaitu sekitar
1.85 mm dan tidak berbeda dengan genotipe A1 (Karanganyar), A2
(Sukabumi) dan A6 (IP-1A) (Tabel 7).
Tabel 7 Pengaruh interaksi antara kadar air media dan genotipe terhadap ketebalan daun (mm) jarak pagar pada akhir pengamatan
Genotipe Kadar air media
40% 36% 32%
A1 (Karanganyar) 1.94 ef 2.06 cbde 2.17 abcd
A2 (Sukabumi) 2.04 bcdef 2.10 abcde 2.14 abcde
A3 (NTB) 2.25 ab 2.00 cdef 2.31 a
A4 (IP-1P) 2.13 abcde 2.13 abcde 2.28 a
A5 (IP-1M) 1.85 f 2.29 a 2.17 abc
A6 (IP-1A) 1.96 def 2.19 abc 2.30 a
29
Penurunan kadar air media dari 40% menjadi 36% meningkatkan ketebalan daun hampir pada semua genotipe kecuali genotipe A3 (NTB)
dan A4 (IP-1P). Genotipe A5 (IP-1M) mengalami peningkatan tebal yang
paling tinggi menjadi 2.29 dari 1.85 mm . Respon yang nyata terhadap peningkatan ketebalan daun akibat penurunan kadar air media dari 40% menjadi 32% terlihat pada genotipe A1 (Karanganyar), A5 (IP-1M) dan A6
(IP-1A) yang memiliki peningkatan ketebalan daun masing-masing sebesar 0.23 mm, 0.32 mm dan 0.34 mm. Data ini menunjukkan bahwa perlakuan kadar air media 32% menghasilkan daun yang lebih tebal dibandingkan dengan perlakuan kadar air media 40%.
Menurut Gardner et al. (1991) terjadinya respon penebalan daun pada suatu tanaman yang tercekam kekeringan yaitu untuk mengurangi terjadinya kehilangan air pada tanaman. Respon tersebut juga telah diteliti oleh Yusnaeni (2002) pada tanaman Hoya. Ketebalan daun tanaman hoya meningkat 1.60 mm pada tanaman yang diberi cekaman (penyiraman tiap minggu) dibandingkan dengan yang tidak diberi cekaman (penyiraman tiap hari) sekitar 1.58 mm. Hasil penelitian ini juga menunjukkan respon peningkatan ketebalan daun pada berbagai genotipe yang tercekam kekeringan.
Jumlah stomata terbuka
Tabel 8 menunjukkan bahwa respon jumlah stomata terbuka dan tertutup dipengaruhi oleh cekaman kekeringan dan berbeda nyata pada setiap kadar air media. Jumlah stomata tertutup lebih banyak dari pada jumlah stomata terbuka. Jumlah stomata terbuka pada kadar air media 32% lebih rendah sekitar 7.4%, jauh lebih rendah dibandingkan dengan kondisi 40% dan 36% sebesar 17.3 dan 14.4%.
Tabel 8 Pengaruh kadar air media terhadap jumlah stomata terbuka, stomata tertutup dan total stomata tanaman jarak pagar pada akhir pengamatan
Perlakuan Stomata terbuka Stomata tertutup Total stomata K1 (KAM 40 %) 42.5 a (17.3%) 202.5 c (82.7%) 245.00
K2 (KAM 36 %) 35.4 b (14.4%) 210.26 b (85.6%) 245.66
K3 (KAM 32 %) 18.1 c ( 7.4%) 227.87 a (92.6%) 245.97 Ket : Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf
30
Semakin rendah kadar air media dapat menyebabkan stomata semakin banyak yang menutup sehingga mengakibatkan berkurangnya asimilasi CO2. Hasil penelitian ini menunjukkan terjadinya peningkatan
jumlah stomata yang tertutup dengan meningkatnya cekaman kekeringan. Pada perlakuan kadar air media 40%, jumlah stomata tertutup sekitar 82.7%. Penurunan kadar air media dari 40% menjadi 36% meningkatkan jumlah stomata tertutup sekitar 85.6%. Jika kadar air media diturunkan lebih lanjut menjadi 32% terjadi peningkatan jumlah stomata yang tertutup lebih banyak yaitu 92.6%. Hal ini sesuai dengan penelitian yang dilakukan oleh Pangaribuan (2001) tentang kelapa sawit, bahwa kelapa sawit yang diberi cekaman menurunkan jumlah stomata terbuka menjadi 8.70 pada 25% KL dari 9.00 pada 100% KL.
Tabel 9 menunjukkan bahwa pada semua genotipe memiliki total stomata yang sama, Genotipe A3 (NTB) yang memiliki jumlah stomata
terbuka yang paling rendah sekitar 12.2% dan tidak berbeda dengan genotipe A4 (IP-1P). Genotipe A6 (IP-1A) mempunyai jumlah stomata
terbuka paling tinggi sekitar 13.5%, kemudian diikuti oleh genotipe A1
(Karanganyar), A2 (Sukabumi), A5 (IP-1M), dan A4 (IP-1P). Genotipe A3
(NTB) memiliki jumlah stomata terbuka lebih sedikit tetapi tidak berbeda dengan A4 (IP-1P), namun berbeda dengan genotipe A1 (Karanganyar), A2
(Sukabumi), A5 (IP-1P) dan A6 (IP-1A). Respon jumlah stomata yang
tertutup menunjukkan bahwa genotipe A3 (NTB) lebih banyak
dibandingkan dengan genotipe yang lainnya.
Tabel 9 Pengaruh genotipe terhadap jumlah stomata terbuka, tertutup dan total stomata tanaman jarak pagar pada akhir pengamatan. Genotipe Stomata terbuka Stomata tertutup Total stomata A1 (Karanganyar) 32.4 a (13.2) 213.4 ab (86.8) 245.8
A2 (Sukabumi) 32.3 a (13.3) 210.4 b (86.7) 242.7
A3 (NTB) 30.1 b (12.2) 216.6 a (87.8) 246.7
A4 (IP-1P) 31.9 ab (12.9) 215.5 a (87.1) 247.4
A5 (IP-1M) 32.3 a (13.2) 213.4 ab (86.9) 245.7
A6 (IP-1A) 33.0 a (13.5) 211.9 b (86.5) 244.9 Ket : Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf
31
Genotipe tidak berpengaruh nyata terhadap jumlah stomata terbuka (Tabel 2) akan tetapi Tabel 9 menunjukkan adanya kecenderungan bahwa genotipe NTB mempunyai jumlah stomata terbuka yan paling rendah dibandingkan genotipe lainnya. Ada kemungkinan hal ini karena genotipe NTB termasuk daerah kering, sehingga genotipe tersebut sudah teradaptasi untuk membuka sebagian kecil stomatanya pada kondisi yang kurang menguntungkan.
Defisit air
Defisit air menunjukkan besarnya kehilangan air dalam jaringan dibandingkan dengan kandungan air pada kondisi turgor penuh. Semakin besar defisit air maka semakin rendah air yang tersedia untuk proses metabolisme tanaman. Respon defisit air tidak dipengaruhi oleh kadar air media (Tabel 2), akan tetapi data pada Tabel 10 menunjukkan bahwa tanaman yang ditanam pada kadar air media 32% mengalami defisit air lebih besar (74.8%) dari pada 40 dan 36%, masing-masing sebesar 72.9 dan 74.0%. Perbedaan defisit air tanaman yang ditanam di media berkadar air 40% dengan 36% setelah delapan minggu perlakuan hanya sebesar 1.1%, sedangkan perbedaan defisit air dari media berkadar air 40% dengan 32% sebesar 1.9%.
Tabel 10 Pengaruh kadar air media terhadap defisit air (%) jarak pagar pada akhir pengamatan
Perlakuan Nilai tengah
K1 (KAM 40 %) 72.9 b
K2 (KAM 36 %) 74.0 ab
K3 (KAM 32 %) 74.8 a
Ket : Angka yang diikuti oleh huruf yang sama tidak berbeda nyata pada taraf DMRT 5 %.