DIVERSITAS LUMUT EPIFIT
DI TIGA PERKEBUNAN TEH JAWA BARAT
HILDA AKMAL
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis yang berjudul Diversitas Lumut Epifit di Tiga Perkebunan Teh Jawa Barat adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain, telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Juni 2012
ABSTRACT
HILDA AKMAL. Diversity of Epiphytic Bryophytes at Three Tea Plantations in West Java. Supervised by SRI SUDARMIYATI TJITROSOEDIRDJO, NUNIK SRI ARIYANTI and SULISTIJORINI.
Study on the epiphytic bryophytes was conducted at three tea plantations located at different elevation in West Java: Gunung Mas (600 m) Bogor, Nirmala (1150 m) Sukabumi, Rancabali (1628 m) Bandung. Three plots were established within each tea plantation, three transects of 30 m were lined in each plot. Samples were collected from the trunk and primary branches of five tea trees in each line transect. The aims of this study were to record the epiphytic bryophytes species of the plantations and to compare diversity of the bryophytes community among the plantations. Ninety species of epiphytic bryophytes were recorded. Checklist of those species were provided in this paper. The bryophytes species richness of the plantations increased with increasing of the elevation. Twenty seven species were found at Gunung Mas, 40 species at Nirmala, and 74 species at Rancabali. The plantations were dominated by different species. The dominant species were Sematophyllum tristiculum (mosses) at Gunung Mas, Lejeunea
anisophylla (liverworts) at Nirmala, and Acanthocoleus javanicus (liverworts) at
Rancabali. The species composition of the bryophytes communities of Gunung Mas was more similar to that of Nirmala compared to that of Rancabali.
RINGKASAN
HILDA AKMAL. Diversitas Lumut Epifit di Tiga Perkebunan Teh Jawa Barat. Dibawah bimbingan SRI SUDARMIYATI TJITROSOEDIRDJO, NUNIK SRI ARIYANTI dan SULISTIJORINI.
Lumut (bryophyte) merupakan kelompok tumbuhan terbesar kedua setelah Angiosperma. Hutan hujan di kawasan tropik merupakan salah satu habitat dengan keanekaragaman lumut yang tinggi. Penelitian tentang flora lumut di pulau Jawa telah berlangsung lama, yaitu sejak hampir dua ratus tahun yang lalu. Sesudah tahun 1960 hanya sedikit studi floristik tentang lumut yang dilakukan di pulau Jawa. Dari beberapa penelitian lumut akhir-akhir ini masih diperoleh catatan baru untuk flora lumut di Jawa. Hal ini menunjukkan bahwa data tentang flora lumut di pulau Jawa sampai saat ini masih belum lengkap. Penelitian dan eksplorasi yang ada umumnya dilakukan pada komunitas lumut di hutan alam, penelitian terhadap lumut di luar kawasan hutan alam masih sangat jarang.
Penelitian ini bertujuan untuk mencatat keanekaragaman jenis lumut epifit di tiga perkebunan teh pada elevasi berbeda di Jawa Barat, dan membandingkan diversitas lumut epifit pada perkebunan teh tersebut. Keanekaragaman jenis lumut disajikan dalam bentuk daftar jenis (checklist) lumut epifit di tiga perkebunan teh. Diversitas komunitas lumut epifit dibandingkan berdasarkan kekayaan, komposisi, dan dominansi jenis, serta kemelimpahan total lumut.
Pengambilan sampel lumut dilakukan di tiga perkebunan teh yaitu di Gunung Mas (600 m dpl), Nirmala (1150 m dpl), dan Rancabali (1628 m dpl). Di setiap perkebunan teh dibuat tiga plot penelitian masing-masing berukuran 30 m x 20 m, jarak antar plot minimal 500 m. Pada setiap plot dibuat tiga transek, jarak antar transek 5 m. Pada setiap transek ditentukan 5 sampel tanaman teh, berjarak 5 m. Persentase penutupan oleh setiap jenis lumut dan penutupan oleh total lumut pada setiap sampel tanaman teh dicatat. Sebagai data pendukung dicatat pula penutupan tanaman teh oleh liken dan tumbuhan paku. Sampel lumut diambil dari batang dan cabang primer pada setiap sampel tanaman teh. Sampel lumut dari lokasi diidentifikasi dengan mengamati ciri-ciri spesifik gametofit dan sporofitnya menggunakan mikroskop, dilanjutkan dengan dokumentasi.
kemelimpahan totalnya sangat berbeda dari jenis-jenis lainnya. Komunitas lumut epifit di Rancabali dan Gunung Mas memiliki kesamaan komposisi yang paling kecil (38%) dibandingkan antara Rancabali dan Nirmala (51%), maupun antara Nirmala dan Gunung Mas (54%). Hal ini menunjukkan bahwa semakin besar perbedaan elevasi antar lokasi semakin kecil kesamaan komunitas lumut antar keduanya. Di perkebunan teh Rancabali dijumpai 74 jenis lumut epifit, 41 jenis di antaranya tidak ditemukan di lokasi perkebunan lainnya. Acanthocoleus javanicus
merupakan jenis lumut hati yang dominan di Rancabali, Lejeunea anisophylla di Nirmala, dan Sematophyllum tristiculum dominan di Gunung Mas. Rata-rata
penutupan tanaman teh oleh lumut paling rendah di Nirmala. Hal ini diduga berhubungan dengan ukuran tajuk tanaman teh
© Hak Cipta milik IPB, Tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
DIVERSITAS LUMUT EPIFIT
DI TIGA PERKEBUNAN TEH JAWA BARAT
HILDA AKMAL
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Biologi Tumbuhan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Diversitas Lumut Epifit di Tiga Perkebunan Teh Jawa Barat Nama : Hilda Akmal
NIM : G353090301
Disetujui Komisi Pembimbing
Dr. Sri Sudarmiyati Tjitrosoedirdjo, M.Sc Ketua
Dr. Nunik Sri Ariyanti, M.Si Dr.Ir.Sulistijorini, M.Si Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana Biologi Tumbuhan
Dr.Ir. Miftahudin, M.Si Dr.Ir.Dahrul Syah, M.Sc.Agr
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat ALLAH SWT atas segala kasih sayang NYA sehingga karya ilmiah ini dapat diselesaikan.
Penelitian berjudul “Diversitas Lumut Epifit di Tiga Perkebunan Teh Jawa Barat” ini merupakan salah satu syarat untuk memperoleh gelar Magister Sains pada Program Studi Biologi Tumbuhan, Sekolah Pascasarjana IPB. Penelitian ini dibiayai oleh proyek Fundamental DIKTI tahun 2010 atas nama Dr. Nunik Sri Ariyanti, MSi.
Pada kesempatan ini penulis ingin menyampaikan terima kasih yang sebesar-besarnya kepada:
1. Dr. Sri Sudarmiyati Tjitrosoedirdjo, MSc; Dr. Nunik Sri Ariyanti, MSi; dan Dr. Ir. Sulistijorini, MSi; sebagai komisi pembimbing yang telah memberikan bimbingan, saran dan masukan selama penelitian dan penulisan laporan ini. 2. Dr. Ir. Aris Tjahyoleksono, DEA, sebagai wakil Program Studi Biologi
Tumbuhan, yang telah memberikan saran dan masukan.
3. Dr. Harry Wiriadinata selaku penguji, yang telah banyak memberikan saran dan masukan.
4. Departemen Biologi FMIPA-IPB, yang telah memberikan kesempatan dan
penugasan, sekaligus menjadi penyandang dana bagi penulis untuk melanjutkan studi di Sekolah Pascasarjana IPB.
5. Dekan FMIPA-IPB, Rektor IPB, yang telah memberikan kesempatan, izin dan
penugasan bagi penulis untuk melanjutkan studi di Sekolah Pascasarjana IPB. 6. Ketua Program Studi Biologi Tumbuhan Sekolah Pascasarjana dan Dekan
Sekolah Pascasarjana IPB, yang telah memberikan kesempatan kepada penulis untuk menyelesaikan studi.
7. Seluruh Staf Pengajar dan Staf Penunjang di Departemen Biologi, yang telah memberikan bantuan dan dukungannya.
8. Pimpinan serta staf Perkebunan Teh Gunung Mas, Nirmala, dan Rancabali, atas izin dan bantuan yang diberikan kepada penulis untuk pengambilan sampel lumut di lokasi perkebunan.
9. Rekan-rekan di Laboratorium Anatomi Tumbuhan dan Laboratorium
Taksonomi Tumbuhan Departemen Biologi: bapak Sunaryo, bapak Suparman, Saiful Bachri, SSi, Marinda Sari Sofiyana, SSi, Fibo dll, atas bantuan, kerjasama, dan semangat yang telah diberikan.
10. Sdri. Indah Wahyuni, SSi dan bapak Setiabudi S.Hut di Herbarium BIOTROP
(BIOT), atas bantuan dan kerjasamanya.
11. Hadisunarso, Okkie Senna Maladi, mama Damuniar, ibu Umi Kalsum, ibu Sumarsinah, kakak-kakak dan adik-adik atas dorongan semangat, pengorbanan, doa dan kasih sayangnya selama penulis menempuh studi.
12.Semua pihak yang terlibat dalam proses pendidikan, yang tidak dapat penulis sebutkan satu-persatu.
Harapan penulis, semoga penelitian ini bermanfaat bagi para pembaca yang memerlukan.
Bogor, Juni 2012
RIWAYAT HIDUP
Penulis dilahirkan di Sawahlunto pada tanggal 1 September 1954 dari ayah Drs. Akmal Yunus dan ibu Damuniar Kamal. Penulis adalah anak keempat dari tujuh bersaudara.
Pada tahun 1974 penulis lulus dari SMA Gabungan Jayapura, dan pada tahun yang sama diterima di Fakultas Biologi Universitas Gadjah Mada Jogjakarta. Selama kuliah penulis menjadi asisten praktikum mata kuliah Fisiologi Hewan dan Histologi, selain itu penulis juga aktif sebagai anggota Himpunan Mahasiswa Islam (HMI). Pada tahun 1980 penulis lulus sebagai sarjana dari Fakultas Biologi Universitas Gadjah Mada.
Pada tahun 1981 sampai dengan 1982 penulis menjadi staf pengajar tidak tetap untuk mata kuliah Biologi dan Genetika di FKIP Universitas Cendrawasih Jayapura. Selanjutnya pada tahun 1982 penulis diterima menjadi staf pengajar di
DAFTAR ISI
Halaman
DAFTAR ISI ... xi
DAFTAR TABEL ... xii
DAFTAR GAMBAR... xii
PENDAHULUAN ... 1
TINJAUAN PUSTAKA ... 3
Peranan Ekologi dan Potensi Lumut... 4
Keanekaragaman Jenis Lumut di Berbagai Habitat... 4
Keanekaragaman Jenis Lumut pada Elevasi yang Berbeda... 6
Penelitian Lumut di Perkebunan Teh ... 7
METODE PENELITIAN ... 9
Lokasi Penelitian ... 9
Pengambilan Sampel Lumut ... 10
Identifikasi Sampel Lumut ... 12
Analisis Data ... 12
HASIL ... 15
Checklist Lumut Epifit di Perkebunan Teh Jawa Barat: Gunung Mas, Nirmala dan Rancabali... 15 Diversitas Lumut Epifit di Tiga Perkebunan Teh pada Elevasi Berbeda ... 18
PEMBAHASAN ... 25
Checklist Lumut Epifit di Perkebunan Teh Jawa Barat: Gunung Mas, Nirmala, dan Rancabali ... 25
Diversitas Lumut Epifit di Tiga Perkebunan Teh pada Elevasi yang Berbeda... 26
SIMPULAN ... 33
DAFTAR TABEL
Halaman
1. Elevasi, kisaran kelembapan, kisaran suhu, dan rata-rata curah
hujan di perkebunan teh Gunung Mas, Nirmala, dan Rancabali... 9
2. Checklist lumut epifit di tiga perkebunan teh Gunung Mas (GM),
Nirmala (NR), dan Rancabali (RB), Jawa Barat ... 15
3. Daftar jenis lumut sejati dan lumut hati dengan indeks nilai penting (INP) lebih dari 10% di perkebunan teh Gunung Mas,
Nirmala, dan Rancabali... 21
DAFTAR GAMBAR
Halaman
1. Lokasi pengambilan sampel lumut di tiga perkebunan teh
Jawa Barat: Gunung Mas (A), Nirmala (B), dan Rancabali (C)... 9
2 Skema plot dan penentuan sampel tanaman teh dalam setiap plot
di perkebunan teh... 10
3 Sampel tanaman teh dengan jenis-jenis lumut epifit (a) dan
sampel lumut yang dikoleksi sebagai spesimen herbarium (b)... 11
4 Alur identifikasi spesimen lumut di laboratorium ...11
5 Total jenis lumut di perkebunan teh Gunung Mas, Nirmala, dan Rancabali (a), rata-rata jumlah jenis lumut per tanaman teh dan rata-rata jumlah jenis lumut per plot (b) di masing-masing perke-
bunan teh... 19
6. Total jumlah jenis lumut epifit dan indeks similaritas Sorensen (ISS) pada dua perkebunan teh yang dibandingkan: Gunung Mas (GM) vs. Nirmala (NR), Gunung Mas vs. Rancabali (RB), dan
Nirmala vs. Rancabali... 20
7. Rata-rata persentase penutupan oleh total lumut, liken, dan tumbuhan paku, terhadap batang dan cabang primer tanaman teh di perkebunan teh Gunung Mas, Nirmala, dan Rancabali ... 23
8. Rata-rata diameter tajuk sampel tanaman teh di perkebunan teh
Gunung Mas, Nirmala, dan Rancabali ... 23
Lumut (bryophyte) merupakan kelompok tumbuhan terbesar kedua setelah
Angiosperma. Saat ini diperkirakan ada sekitar 15000 jenis lumut meliputi lumut hati (liverworts), lumut sejati (mosses), dan lumut tanduk (hornworts) (Gradstein
et al. 2001). Diperkirakan sebanyak 1196 jenis lumut ada di pulau Jawa, meliputi
554 jenis lumut hati dan 14 jenis lumut tanduk (SÖderstrÖm et al. 2010), serta 628
jenis lumut sejati (Tan & Iwatsuki 1999).
Penelitian tentang flora lumut di pulau Jawa telah berlangsung lama, dilakukan sejak hampir dua ratus tahun yang lalu, terutama dilakukan pada masa penjajahan Belanda (Schiffner 1900; Verdoorn 1930), namun tidak berlanjut dalam waktu yang cukup lama. Sesudah tahun 1960 hanya sedikit studi floristik tentang lumut yang dilakukan di pulau Jawa, oleh karena itu pengetahuan tentang flora lumut di pulau Jawa sampai saat ini belum lengkap. Beberapa penelitian lumut yang dilakukan akhir-akhir ini melaporkan masih ada catatan baru untuk flora lumut di Jawa (Tan etal. 2006; Haerida et al. 2010; Gradstein et al. 2010).
Lumut dijumpai di semua tipe habitat kecuali di laut (Tan & PÖcs 2002),
tumbuh di berbagai substrat baik di tanah, di bebatuan, pada kayu yang melapuk, tumbuh sebagai epifit di batang pohon, dan substrat lainnya (Gradstein et al.
2001). Hutan hujan di kawasan tropik merupakan salah satu habitat dengan keanekaragaman lumut yang tinggi. Dua pertiga dari total keanekaragaman lumut dunia dijumpai di hutan hujan tropik (Gradstein & PÖcs 1989). Selain dipengaruhi
iklim persebaran lumut juga ditentukan oleh iklim mikro (suhu, kelembapan
udara, intensitas cahaya, dan ketersediaan substrat). Oleh karena itu dijumpai diversitas lumut yang bervariasi di berbagai tipe habitat pada iklim yang sama, seperti di kawasan Asia tropik pada habitat hutan alam (Gradstein & Culmsee 2010), pada hutan perkebunan (Sporn et al. 2007), pada perkebunan monokultur (Ariyanti et al. 2008; Suleiman et al. 2009), dan di hutan kota (kebun raya)
(Apriana 2010; Junita 2010; Wahyuni 2010).
2
suhu rata-rata lebih dari 15OC (Ohsawa 1982). Penelitian lumut di perkebunan teh belum banyak dilakukan. Informasi lumut di perkebunan teh yang ada berasal dari eksplorasi singkat yang dilakukan Pancho (1979), Tan et al. (2006), dan
Gradstein et al. (2010). Penelitian tentang kemelimpahan dan jenis-jenis lumut
umum dan dominan di perkebunan teh belum dilakukan.
Perkebunan teh di Indonesia umumnya berada pada ketinggian 600 m dpl sampai sekitar 2000 m dpl. Beberapa penelitian tentang keanekaragaman lumut pada elevasi berbeda telah dilaporkan namun semuanya dilakukan pada tipe habitat hutan alam (Enroth 1990; Grau et al. 2007; Gardstein & Culmsee 2010;
Chantanaorrapint 2010; Ariyanti & Sulistijorini 2011). Penelitian tentang keanekaragaman lumut pada elevasi berbeda di habitat perkebunan belum dilakukan. Penelitian ini bertujuan untuk mencatat keanekaragaman jenis lumut epifit di tiga perkebunan teh di Jawa Barat, dan membandingkan diversitas lumut
umumnya berukuran kecil, memiliki profil yang umumnya rendah dengan tinggi sekitar 1-2 cm. Lumut pada umumnya tidak mempunyai jaringan pembuluh dan tanpa kutikula atau dengan kutikula yang sangat tipis, dinding sel tanpa diperkuat oleh lignin. Lumut adalah tumbuhan poikilohidrik, bergantung pada kelembapan yang ada di lingkungannya. Pada periode kering lumut bersifat dorman, pada saat kondisi lingkungan menjadi basah lumut segera menyerap air untuk aktivitas fotosintesis. Lumut berkembang biak dengan spora, pergiliran generasi didominasi oleh gametofit (Gradstein et al. 2001).
Dewasa ini diperkirakan terdapat lebih dari 15000 jenis lumut yang termasuk ke dalam 1200 marga dengan persebaran yang luas. Tiga divisi lumut meliputi: Marchantiophyta (lumut hati/liverworts) terdiri atas 6000-8000 jenis
(Crandall-Stotler & Stotler 2002; Gradstein et al. 2001), Bryophyta (lumut sejati/mosses) lebih dari 10000 jenis (Buck & Goffinet 2002), dan Anthophyta (lumut tanduk/hornworts) sekitar 100 jenis (Renzaglia & Vaughn 2002). Kekayaan jenis lumut di kawasan Indonesia telah dilaporkan oleh beberapa peneliti. Tan dan Iwatsuki (1999) melaporkan ada 426 jenis lumut di Irian Jaya dan 650 jenis di Borneo. Dari checklist yang disusun oleh Gradstein et al. (2005), ada 476 jenis lumut di Sulawesi. Ho et al. (2006) melaporkan 490 jenis lumut sejati di Sumatera. Di pulau Jawa, ada 568 jenis lumut hati dan lumut tanduk yang dilaporkan oleh SÖderstrÖm et al. (2010), dan 628 jenis lumut sejati (Tan & Iwatsuki 1999), sebagian besar ditemukan di Jawa Barat.
4
Peranan Ekologi dan Potensi Lumut
Lumut di kawasan hutan hujan pegunungan terdapat melimpah dengan keanekaragaman yang tinggi, berperan dalam konservasi tanah. Lapisan lumut yang tebal di permukaan lahan yang terbuka ataupun di lantai hutan, dapat memperlambat aliran air sehingga mencegah erosi. Lumut epifit yang tumbuh di batang pohon dapat memperlambat aliran air di permukaan batang (stem flow).
Hal ini dimungkinkan karena kemampuan lumut yang tinggi dalam menyerap sekaligus menahan air hujan, mencapai 5-25 kali bobot keringnya (Gradstein et al.
2001).
Lumut dapat digunakan sebagai bioindikator polutan. Kondisi ini dimungkinkan karena dengan tidak terdapatnya kutikula pada lumut memudahkan larutan dan gas, termasuk gas dan larutan pencemar, mencapai sel-sel sekaligus terakumulasi dalam jumlah besar (Glime 2007; Dey & De 2012).
Sebagian besar polutan tersebut diakumulasi dalam sel-sel daun (Gradstein et al.
2001).
Lumut sudah dimanfaatkan dalam berbagai bidang di banyak negara. Di bidang kesehatan, dari berbagai penelitian dilaporkan adanya sejumlah kandungan bahan aktif pada lumut, beberapa di antaranya bersifat sebagai antijamur dan antibakteri (Bodade et al. 2008; Asakawa 2008; Beike 2010; Dey & De 2012). Pada lumut juga ditemukan berbagai senyawa kimia sebagai metabolit sekunder, di antaranya terpenoid dan komponen aromatik, yang dimanfaatkan di bidang industri (Mues 2002; Dey & De 2012). Taman lumut dengan koleksi berbagai jenis lumut digunakan untuk edukasi di samping dari segi estetika bermanfaat untuk menambah keindahan dan kesejukan. Selain itu lumut juga banyak digunakan dalam bidang kesenian (Glime 2007).
Keanekaragaman Jenis Lumut di Berbagai Habitat
& Sulistijorini 2011). Penggundulan hutan khususnya di daerah tropik menyebabkan hilangnya jenis lumut endemik. Tingkat kehilangan jenis lumut bervariasi bergantung kepada skala perubahan habitat yang terjadi. Tumbuhan epifit yang ternaung (shade epiphyte), termasuk lumut epifit, paling terpengaruh
oleh gangguan hutan tersebut (Gradstein 1992).
Alih fungsi hutan primer menjadi hutan sekunder atau perkebunan akan menurunkan keanekaragaman lumut epifit. Gradstein (1992) menyatakan bahwa sekitar 20% marga lumut hati shade epiphyte di hutan primer tidak dijumpai di
hutan sekunder ataupun di perkebunan, akibat terbukanya kanopi hutan primer. Adanya pembukaan area hutan yang digantikan oleh perkebunan dengan tanaman eksotik, menyisakan sekitar 10% jenis lumut, sedangkan pada hutan sekunder tersisa sekitar 50-70% jenis lumut dibandingkan hutan primer (Gradstein 1992).
Informasi dan data tentang kekayaan lumut epifit di luar hutan primer, termasuk kekayaan lumut yang ada di perkebunan, masih sangat jarang (Gradstein 1992). Kekayaan jenis lumut yang terdapat di daerah monokultur seperti perkebunan, tidak setinggi yang terdapat di hutan primer sebagai habitat alaminya. Sungguhpun demikian jenis lumut epifit yang tumbuh di perkebunan, dapat beradaptasi terhadap habitat mikro yang relatif kering dan tetap mempunyai peluang hidup yang baik (Gradstein et al. 2001).
Penelitian yang dilakukan Ariyanti et al. (2008) di Sulawesi Tengah pada tiga habitat yang berbeda yaitu di hutan primer, hutan sekunder, dan perkebunan kakao menunjukkan kekayaan jenis lumut sejati terendah ditemukan di perkebunan kakao, diikuti hutan sekunder. Kekayaan jenis lumut sejati tertinggi ditemukan di hutan primer. Lumut yang dominan di perkebunan kakao adalah
Caudalejeunea revurvistipula, Lopholejeunea subfusca, Mastigolejeunea
auriculata, Floribundaria floribunda, dan Chaetomitrium lanceolatum.
Suleiman et al. (2009) melaporkan, di perkebunan kelapa sawit Sabah
ditemukan lumut sejati sebanyak 56 jenis dalam 31 marga dan 14 suku. Jumlah
tersebut sekitar 7,7% dari total lumut sejati yang ada di Borneo dan 9,3% dari lumut sejati yang ada di Sabah. Dalam penelitian tersebut ditemukan Acroporium
convolutum sebagai catatan baru untuk Borneo dan Ectropotheciella
6
kelapa sawit adalah Syrrhopodon ciliatus dan Atrhocormus schimperi, dan pada permukaan tanah di area perkebunan kelapa sawit umumnya adalah lumut
Vesicularia dubyana.
Eksplorasi dan checklist lumut di Kebun Raya Bogor masih terus
dilakukan. Junita (2010) mengidentifikasi sebanyak 42 jenis lumut sejati epifit
yang termasuk dalam 21 marga dan 11 suku. Apriana (2010) melaporkan sejauh ini ditemukan sebanyak 92 jenis lumut hati epifit di Kebun Raya Bogor, dari hasil eksplorasi dan dari koleksi spesimen yang ada di Herbarium Bogoriense. Wahyuni (2010) melaporkan sebanyak 43 jenis lumut terestrial meliputi 33 jenis lumut sejati dan 10 jenis lumut hati pada lokasi yang sama.
Keanekaragaman Jenis Lumut pada Elevasi yang Berbeda
Secara umum di kawasan hutan hujan tropik kekayaan jenis lumut hati meningkat pada elevasi yang lebih tinggi sedangkan kekayaan jenis lumut sejati justru semakin menurun. Gradstein dan Culmsee (2010) dalam penelitian di Sulawesi Tengah, menemukan sekitar 55% lumut sejati dan 45% lumut hati di hutan dataran rendah. Pada hutan pegunungan bawah lumut hati memiliki kekayaan jenis yang lebih tinggi (60%) dibandingkan lumut sejati (40%). Grau et
al. (2007) dari penelitian di Nepal, melaporkan kekayaan jenis lumut meningkat dengan bertambahnya elevasi. Pada penelitian tersebut, kekayaan lumut hati tertinggi dijumpai pada elevasi 2800 m dpl dan lumut sejati pada ketinggian 2500 m dpl. Enroth (1990) meneliti lumut di Huon Peninsula Papua New Guinea, dan melaporkan adanya kecenderungan kekayaan jenis lumut yang meningkat sampai dengan elevasi 2300 m dpl, pada elevasi yang lebih tinggi kekayaan jenis lumut mulai menurun. Menurut Richards (1984) setiap kenaikan elevasi sekitar 100 m di daerah tropik akan menyebabkan penurunan suhu sekitar 0.4-0.7OC dan kenaikan
curah hujan tahunan. Umumnya lumut tumbuh optimal pada kisaran suhu 15-25O
C dengan intensitas cahaya yang rendah, dan pada lingkungan yang lembap dan
Penelitian Lumut di Perkebunan Teh
Teh (Camellia sinensis) diperkirakan berasal dari China yang telah
memanfaatkannya sejak 2000 tahun yang lalu. Pada masa itu teh masih ditanam secara tradisional. Budidaya teh di perkebunan baru dilakukan mulai tahun 1820 di India. Sejak tahun 1870 teh mulai menyebar ke beberapa negara tropik, termasuk Indonesia. Dewasa ini areal perkebunan teh sudah menyebar dari daerah beriklim sedang sampai daerah tropik (Ohsawa 1982). Perkebunan teh di
Indonesia umumnya dijumpai di dataran tinggi, mulai ketinggian 600 m dpl sampai sekitar 2000 m dpl.
Tanaman teh termasuk jenis pohon tetapi umumnya dipelihara sebagai tanaman perdu dengan tinggi kurang dari 150 cm. Lingkungan perkebunan yang
hangat dan lembap dengan suhu rata-rata lebih dari 15OC, dan curah hujan minimal 50 mm/bulan, diperlukan untuk pertumbuhan tanaman teh. Di perkebunan teh juga ditanam berbagai jenis tumbuhan yang berfungsi sebagai
wind breaker yaitu pemecah dan penahan angin untuk mempertahankan iklim
mikro. Pemeliharaan tanaman teh (mencakup pemupukan, pemberantasan hama dan penyakit, pengendalian gulma, dan pemangkasan), dapat menentukan kualitas dan kuantitas teh. Pengendalian hama dan penyakit dilakukan dengan penyemprotan, pengendalian gulma dengan cara tradisional ataupun menggunakan herbisida. Pemangkasan umumnya dilakukan 3-4 tahun sekali untuk merangsang pertumbuhan tunas muda, mempertahankan bidang pemetikan agar tetap rendah, dan meningkatkan produktivitas (Ohsawa 1982).
Penelitian tentang lumut di perkebunan teh belum banyak dilakukan. Informasi yang ada berasal dari penelitian singkat yang dilakukan Pancho (1979) di perkebunan teh Pagilaran. Dalam penelitian tersebut ditemukan 15 jenis lumut dari 12 marga yang termasuk dalam 9 suku. Tan et al. (2006) juga pernah
melakukan penelitian lumut di perkebunan teh Nirmala, tetapi penelitian tersebut
METODE PENELITIAN Lokasi Penelitian
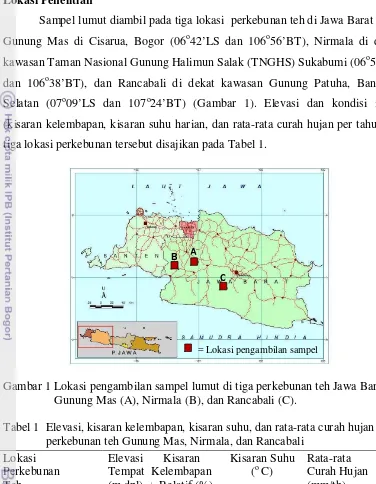
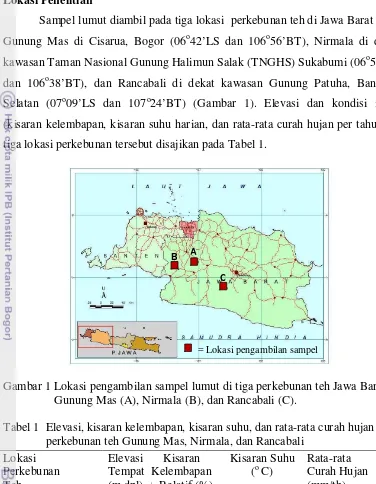
Sampel lumut diambil pada tiga lokasi perkebunan teh di Jawa Barat yaitu Gunung Mas di Cisarua, Bogor (06o42’LS dan 106o56’BT), Nirmala di dekat
kawasan Taman Nasional Gunung Halimun Salak (TNGHS) Sukabumi (06o51’LS
dan 106o38’BT), dan Rancabali di dekat kawasan Gunung Patuha, Bandung Selatan (07o09’LS dan 107o24’BT) (Gambar 1). Elevasi dan kondisi iklim (kisaran kelembapan, kisaran suhu harian, dan rata-rata curah hujan per tahun) di tiga lokasi perkebunan tersebut disajikan pada Tabel 1.
Gambar 1 Lokasi pengambilan sampel lumut di tiga perkebunan teh Jawa Barat: Gunung Mas (A), Nirmala (B), dan Rancabali (C).
Tabel 1 Elevasi, kisaran kelembapan, kisaran suhu, dan rata-rata curah hujan di perkebunan teh Gunung Mas, Nirmala, dan Rancabali
Sumber Informasi:
1)Badan Meteorologi Klimatologi dan Geofisika Balai BesarWilayah II Bogor 2)
ht t p:/ / w w w .dephut .go.id/ INFORM ASI/ TN%20INDO-ENGLISH/ t nhalim un.ht m l
Pengambilan Sampel Lumut
Pengambilan sampel lumut dilakukan dengan metode transek dan petak contoh menurut Mueller-Dombois dan Ellenberg (1974) dengan modifikasi. Di setiap lokasi perkebunan teh dibuat tiga plot penelitian masing-masing berukuran 30 m x 20 m, jarak antar plot minimal 500 m. Pada setiap plot dibuat tiga transek, jarak antar transek 5 m. Pada setiap transek ditentukan 5 sampel tanaman teh, satu sama lain berjarak 5 m. Di setiap plot ada 15 sampel tanaman teh sehingga di setiap lokasi perkebunan ada 45 sampel tanaman teh (Gambar 2). Dicatat dan
diperkirakan persentase penutupan oleh setiap jenis lumut dan penutupan oleh total lumut pada setiap sampel tanaman teh. Selain itu, jenis-jenis lumut yang ditemukan diambil untuk dibuat spesimen herbarium (Gambar 3) dan diidentifikasi lebih lanjut di laboratorium (Gambar 4). Sebagai data pendukung
dicatat pula diameter tajuk tanaman teh, persentase penutupan batang dan cabang primer tanaman teh oleh epifit selain lumut (liken dan tumbuhan paku).
= tanaman teh
Gambar 2 Skema plot dan penentuan sampel tanaman teh dalam setiap plot di perkebunan teh.
20 m
30 m
10
(a) (b)
Gambar 3 Sampel tanaman teh dengan jenis-jenis lumut epifit (a) dan sampel lumut yang dikoleksi sebagai spesimen herbarium (b).
Gambar 4 Alur identifikasi spesimen lumut di laboratorium.
IDENTIFIKASI
M IKROSKOP STEREO M IKROSKOP BINOKULER
HASIL FOTO
HASIL FOTO SPESIM EN
HERBARIUM
Identifikasi Sampel Lumut
Sampel lumut yang diambil dari perkebunan teh selanjutnya dibawa ke laboratorium, diamati ciri-ciri spesifik gametofit maupun sporofit dari masing-masing jenis dengan menggunakan mikroskop untuk keperluan identifikasi, dan dilanjutkan dengan dokumentasi. Spesimen lumut yang dikoleksi dibuat herbarium dan disimpan di Herbarium Bogoriense (BO) dan herbarium BIOTROP (BIOT). Masing-masing koleksi lumut epifit diberi kode kolektor Akmal. Identifikasi lumut hati dilakukan dengan menggunakan kunci identifikasi dalam Yamada (1979), Zhu dan So (2001), Zhu dan Gradstein (2005), dan Gradstein
(2011). Identifikasi lumut sejati menggunakan kunci identifikasi menurut Bartram (1939), Eddy (1988, 1990, 1996), dan Pollatwan (2008).
Analisis Data
Hasil identifikasi berupa nama jenis lumut epifit disusun dalam checklist
yang menunjukkan keanekaragaman jenis lumut epifit di tiga lokasi perkebunan teh yang diteliti. Diversitas lumut epifit di setiap lokasi perkebunan teh dibandingkan berdasarkan kekayaan jenis lumut. Kekayaan jenis lumut ditunjukkan dari rata-rata jumlah jenis lumut per tanaman, rata-rata jumlah jenis lumut per plot dan total jenis lumut per lokasi perkebunan. Perbedaan jumlah jenis lumut per plot dan per tanaman teh di tiga lokasi perkebunan teh ditunjukkan dari nilai rata-rata dan standar deviasi. Tingkat kemiripan komposisi jenis lumut epifit antar perkebunan teh dibandingkan dengan indeks Sorensen (Rumus 1). Selain itu, diversitas komunitas lumut epifit di tiga perkebunan teh juga dinyatakan berdasarkan indeks Shannon (Rumus 2) yang dihitung berdasarkan kemelimpahan proporsional jenis-jenis lumut penyusunnya. Dominansi jenis pada tiga perkebunan teh ditunjukkan dengan indeks nilai penting (INP) (Rumus 3). Selanjutnya dibandingkan jenis-jenis lumut yang kemelimpahannya relatif tinggi (INP lebih dari 10%) dan jenis dominan (INP paling tinggi) di masing-masing lokasi perkebunan. Kemelimpahan total lumut epifit di masing-masing lokasi perkebunan ditunjukkan dari rata-rata persentase penutupan oleh total lumut
12
Rumus 1
Indeks Sorensen (CS)
CS = 2 j/(a+b)
Keterangan :
CS = indeks similaritas Sorensen berdasarkan data
kualitatif
jN = ∑ jenis yang ada di dua lokasi (lokasi 1 dan 2)
a = ∑ jenis di lokasi 1 b = ∑ jenis di lokasi 2
Rumus 2
Indeks Shannon H’ = - ∑ p i ln p i
Keterangan : p i = kemelimpahan proporsional jenis ke – i = n/N
N = total jenis
n = jumlah jenis ke - i
Rumus 3
INP = {FR (%) + PR (%)} Keterangan;
FR = Frekuensi Relatif PR = Penutupan Relatif
Checklist Lumut Epifit di Perkebunan Teh Jawa Barat: Gunung Mas, Nirmala, dan Rancabali
Total jenis lumut epifit yang diinventarisasi dari tiga perkebunan teh di Jawa
Barat (Gunung Mas, Nirmala, Rancabali) adalah 90 jenis yang termasuk dalam 48
marga dan 25 suku. Jumlah tersebut meliputi 42 jenis lumut hati (12 marga, 6 suku)
dan 48 jenis lumut sejati (36 marga, 19 suku). Daftar nama jenis lumut dan
persebarannya di tiga perkebunan teh tersebut disajikan pada Tabel 2. Suku dengan
keanekaragaman paling tinggi adalah Lejeuneaceae, dijumpai sebanyak 26 jenis di
ketiga perkebunan teh.
14
47 Mitthyridium obtusifolium (Lindb.)Robinson v v v Akmal 24, 60, 140
16
Diversitas Lumut Epifit di Tiga Perkebunan Teh pada Elevasi Berbeda
Kekayaan Jenis. Perbandingan kekayaan jenis lumut epifit pada tiga
perkebunan teh di Jawa Barat dapat dilihat pada Gambar 5a. Di Gunung Mas
ditemukan 27 jenis, terdiri atas 13 jenis lumut hati dan 14 jenis lumut sejati.
Kekayaan jenis lumut epifit di Nirmala hampir 1.5 kali lipat dari Gunung Mas,
meliputi 20 jenis lumut hati dan 20 jenis lumut sejati. Perkebunan teh Rancabali
memiliki kekayaan jenis paling tinggi (hampir 3 kali lipat kekayaan jenis di Gunung
Mas), meliputi 36 jenis lumut hati dan 38 jenis lumut sejati.
Rata-rata jumlah jenis lumut epifit per plot dan rata-rata jumlah jenis lumut
epifit per tanaman teh di Gunung Mas, Nirmala, dan Rancabali menunjukkan hasil
yang sebanding dengan total jenis di tiga perkebunan tersebut. Rata-rata jumlah jenis
lumut epifit per plot dan per tanaman teh paling tinggi dijumpai di Rancabali, dan
dan 15 jenis per tanaman teh ditemukan di Rancabali, sedangkan di Gunung Mas
hanya dijumpai rata-rata 19 jenis lumut epifit per plot dan 7 jenis per tanaman teh.
Gambar 5 Total jenis lumut di perkebunan teh Gunung Mas, Nirmala, dan Rancabali (a), rata-rata jumlah jenis lumut per tanaman dan rata-rata jumlah jenis lumut per plot (b) di masing-masing perkebunan teh.
Kesamaan Komposisi Jenis. Beberapa jenis lumut epifit dijumpai hanya di
satu lokasi perkebunan saja, jenis lainnya dijumpai di dua, atau tiga perkebunan teh
sekaligus. Berdasarkan indeks similaritas Sorensen (ISS), komposisi jenis lumut
epifit di Gunung Mas lebih mirip dengan komposisi jenis lumut epifit di Nirmala
(54%) dibandingkan dengan komposisi jenis lumut yang terdapat di Rancabali (38%).
Kemiripan komposisi jenis antara lumut epifit di Nirmala dan Rancabali sebesar 51%
(Gambar 6).
(b)
Jumlah jenis Jumlah jenis
18
Gambar 6 Total jumlah jenis lumut epifit dan indeks similaritas Sorensen (ISS) pada dua perkebunan teh yang dibandingkan: Gunung Mas (GM) vs. Nirmala (NR), Gunung Mas vs. Rancabali (RB), dan Nirmala vs Rancabali.
Diversitas Komunitas Lumut Berdasarkan Indeks Shannon. Berdasarkan
indeks diversitas Shannon (H’), komunitas lumut epifit yang ada di tiga lokasi
perkebunan teh menunjukkan diversitas terendah terdapat di Gunung Mas (H’=2.67), diikuti oleh Nirmala (H’= 3.19), dan yang tertinggi terdapat di Rancabali (H’=3.82).
Dominansi Jenis. Dominansi jenis lumut epifit ditunjukkan oleh frekuensi
kehadiran masing-masing jenis lumut epifit dan persentase penutupannya pada
semua tanaman teh yang diamati. Nilai dominansi tersebut ditunjukkan oleh indeks
nilai penting (INP). Kebanyakan jenis-jenis lumut yang dijumpai memiliki INP
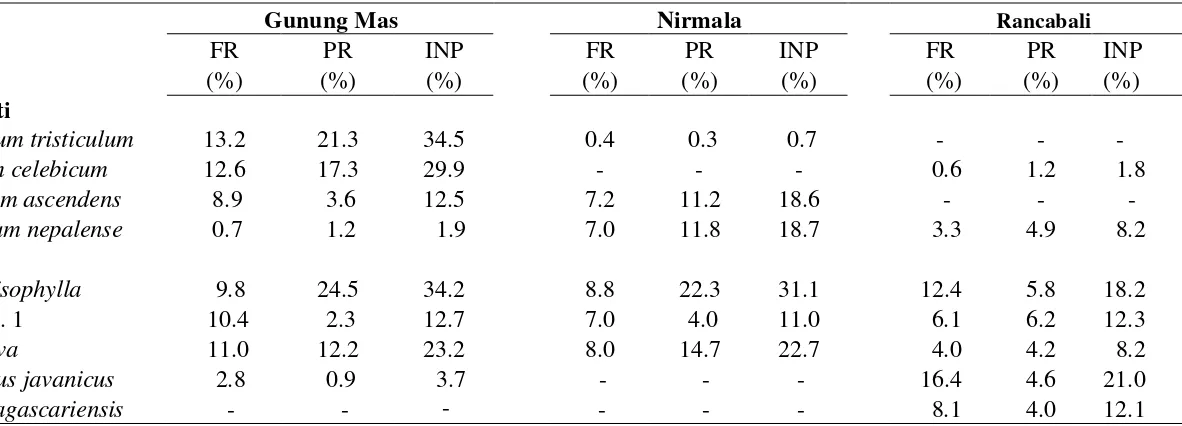
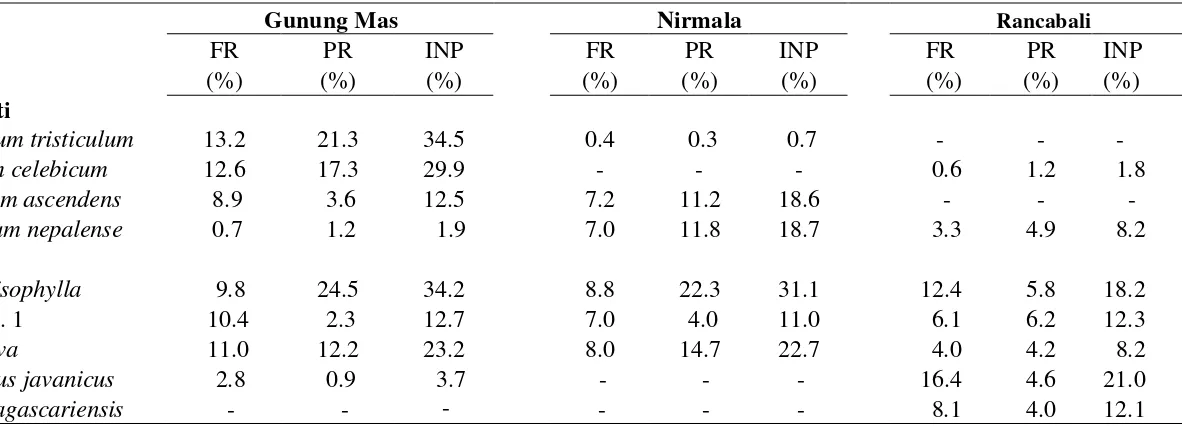
kurang dari 10%. Pada ketiga perkebunan hanya ada empat jenis lumut sejati dan
lima jenis lumut hati dengan INP lebih dari 10%. Daftar jenis lumut hati dan lumut
sejati epifit dengan INP lebih dari 10% ditunjukkan pada Tabel 3.
ISS = 0.51
ISS = 0.38
ISS = 0.54
Keterangan: FR= Frekuensi Relatif, PR= Penutupan Relatif, INP= Indeks Nilai Penting.
Gunung Mas Nirmala Rancabali
No FR PR INP FR PR INP FR PR INP
(%) (%) (%) (%) (%) (%) (%) (%) (%)
Lumut Sejati
1 Sematophyllum tristiculum 13.2 21.3 34.5 0.4 0.3 0.7 - - -
2 Eurhynchium celebicum 12.6 17.3 29.9 - - - 0.6 1.2 1.8
3 Actinodontium ascendens 8.9 3.6 12.5 7.2 11.2 18.6 - - -
4 Brachymenium nepalense 0.7 1.2 1.9 7.0 11.8 18.7 3.3 4.9 8.2
Lumut Hati
1 Lejeunea anisophylla 9.8 24.5 34.2 8.8 22.3 31.1 12.4 5.8 18.2
2 Metzgeria sp. 1 10.4 2.3 12.7 7.0 4.0 11.0 6.1 6.2 12.3
3 Lejeunea flava 11.0 12.2 23.2 8.0 14.7 22.7 4.0 4.2 8.2
4 Acanthocoleus javanicus 2.8 0.9 3.7 - - - 16.4 4.6 21.0
5 Radula madagascariensis - - - - - - 8.1 4.0 12.1
20
Di Rancabali empat jenis lumut dengan INP di atas 10% merupakan lumut
hati, tidak ada jenis lumut sejati yang memiliki INP lebih dari 10%. Jenis lumut
dominan di Rancabali adalah Acanthocoleus javanicus dengan INP 21%. Jenis lumut
tersebut dijumpai juga di Gunung Mas tetapi sangat jarang dan sedikit (frekuensi relatif 2.8%, penutupan relatif 0.9%, INP 3.7%). Acanthocoleus javanicus tidak
dijumpai di Nirmala. Lumut hati Radula madagascariensis sering dijumpai di
Rancabali (frekuensi relatif 8.1 %, penutupan relatif 4.0 %, INP 12.1%), tetapi tidak
dijumpai di dua perkebunan teh lainnya.
Jenis lumut epifit dominan di Nirmala adalah Lejeunea anisophylla dengan
INP lebih dari 30%. Jenis lain yang memiliki nilai INP lebih dari 10% di Nirmala
adalah lumut hati Lejeunea flava dan Metzgeria sp.1, lumut sejati Actinodontium
ascendens dan Brachymenium nepalense. Jenis terakhir juga dijumpai di dua
perkebunan teh lainnya tetapi dengan INP kurang dari 10%. Actinodontium
ascendens juga sering dijumpai di Gunung Mas tetapi tidak dijumpai di Rancabali.
Lumut sejati Sematophyllum tristiculum dengan INP 34.5% dominan di
Gunung Mas. Jenis ini sangat jarang dijumpai di Nirmala dan tidak ditemukan di
Rancabali. Lumut sejati Eurhynchium celebicum (dengan INP 29.9% di Gunung
Mas), tidak dijumpai di Nirmala dan sangat jarang ditemukan di Rancabali. Empat
jenis lainnya dengan INP lebih dari 10% di Nirmala adalah A. ascendens, L.
anisophylla, L. flava, dan Metzgeria sp.1.
Kemelimpahan Total Lumut dan Diameter Tajuk Tanaman Teh.
Kemelimpahan total lumut epifit ditunjukkan dari persentase penutupan lumut
terhadap substrat yang tersedia (batang dan cabang primer tanaman teh).
Kemelimpahan total lumut terendah dijumpai di Nirmala, meningkat di Gunung Mas,
dan tertinggi di Rancabali (Gambar 7). Penutupan oleh lumut rata-rata lebih dari 80%
pada tanaman teh di Rancabali, sedangkan pada tanaman teh di Nirmala penutupan
oleh lumut kurang dari 70%. Berbeda dengan penutupan oleh lumut, penutupan
Gambar 7 Rata-rata persentase penutupan oleh total lumut ( ), liken ( ), dan tumbuhan paku ( ), terhadap batang dan cabang primer tanaman teh di perkebunan teh Gunung Mas, Nirmala, dan Rancabali.
Pada penelitian ini dilakukan pengukuran diameter tajuk tanaman teh yang
diambil lumutnya, karena diduga luas tajuk tanaman teh dapat mempengaruhi
kemelimpahan lumut pada tanaman teh tersebut. Rata-rata diameter tajuk tanaman
teh paling rendah dijumpai di Nirmala (111.5 cm). sedangkan paling tinggi (rata-rata
158.7 cm) dijumpai di Rancabali (Gambar 8).
Gambar 8 Rata-rata diameter tajuk sampel tanaman teh di perkebunan teh Gunung Mas, Nirmala, dan Rancabali.
126
111
158
%
PEMBAHASAN
Checklist Lumut Epifit di Perkebunan Teh Jawa Barat: Gunung Mas, Nirmala, dan Rancabali
Sebanyak 42 jenis lumut hati dan 48 jenis lumut sejati ditemukan di tiga
perkebunan teh di Jawa Barat, yaitu Gunung Mas, Nirmala, dan Rancabali. Jumlah
lumut hati yang ditemukan tersebut hanya mewakili 7.6 % dari total 554 jenis lumut
hati yang pernah dilaporkan ada di Jawa (SÖderstrÖm et al. 2010), dan lumut sejati
yang ditemukan juga hanya mewakili 7.6 % dari total 628 jenis lumut sejati di Jawa
(Tan & Iwatsuki 1999). Lumut epifit yang dilaporkan pada penelitian ini termasuk
dalam 48 marga dan 25 suku. Lejeuneaceae merupakan suku dengan jumlah jenis
tertinggi (26 jenis), diikuti Sematophyllaceae (9 jenis), dan Frullaniaceae (7 jenis).
Menurut Gradstein et al. (2001) suku Lejeuneaceae adalah suku terbesar dalam divisi
Marchantiophyta, memiliki sekitar 90 marga dan lebih dari 1600 jenis. Suku
Lejeuneaceae, Sematophyllaceae, Frullaniaceae dan Plagiochilaceae, merupakan suku
yang umum di kawasan hutan hujan tropik (Gradstein & PÖcs 1989), seperti
dilaporkan juga oleh Gradstein dan Culmsee (2010), Sporn et al. (2010) di hutan
alam Sulawesi, dan oleh Chantanaorrapint (2010) di hutan alam Thailand.
Penelitian lumut di Jawa telah dimulai sekitar 200 tahun lalu, yaitu pada masa
penjajahan Belanda, namun tidak berlanjut dalam waktu yang cukup lama. Penelitian
dan publikasi tentang lumut baru muncul kembali sesudah tahun 2000-an.
Akhir-akhir ini dari penelitian yang dilakukan, masih sering dilaporkan catatan baru untuk
flora lumut di Jawa (Tan et al. 2006; Haerida et al. 2010; Gradstein et al. 2010). Tan
et al. (2006) melaporkan lumut sejati Daltonia armata yang ditemukan di Taman
Nasional Gunung Halimun Salak sebagai jenis catatan baru untuk Jawa. Jenis lumut
tersebut ditemukan di perkebunan teh Nirmala pada penelitian ini, tetapi tidak
ditemukan di dua perkebunan teh lainnya. Adanya catatan baru menunjukkan data
flora yang belum lengkap dan perlu dilakukan penelitian-penelitian untuk melengkapi
daftar jenis lumut yang ada. Daftar jenis yang lengkap diperlukan untuk memonitor
Selain itu, daftar jenis dapat menunjukkan persebaran jenis yang diperlukan bagi
studi taksonomi yang lebih komprehensif.
Diversitas Lumut Epifit di Tiga Perkebunan Teh pada Elevasi yang Berbeda
Perkebunan teh Rancabali dibandingkan dengan dua perkebunan teh lainnya
memiliki kekayaan dan kemelimpahan jenis lumut epifit yang paling tinggi.
Rancabali terletak pada lokasi tertinggi dengan elevasi 1628 m dpl, lebih tinggi dari
Gunung Mas dan Nirmala. Sebaliknya perkebunan teh Gunung Mas dengan elevasi
paling rendah (600 m dpl) memiliki kekayaan jenis lumut epifit paling rendah.
Peningkatan kekayaan jenis lumut epifit seiring dengan peningkatan elevasi juga
dilaporkan oleh Bruun et al. (2006), Grau et al. (2007), Gradstein dan Culmsee
(2010), Chantanaorrapint (2010), serta Ariyanti dan Sulistijorini (2011). Namun
demikian pada penelitian yang lain menunjukkan adanya penurunan kekayaan jenis
lumut pada elevasi lebih dari 2300 m dpl (Enroth 1990).
Persebaran lumut dapat dipengaruhi oleh suhu lingkungan. Menurut Glime
(2007) lumut tumbuh optimum pada kisaran suhu 15-25OC. Peningkatan elevasi dapat
berhubungan dengan penurunan suhu lingkungan. Menurut Richards (1984) setiap
kenaikan elevasi 100 m umumnya menyebabkan penurunan suhu 0.4-0.7OC.
Rata-rata suhu harian di perkebunan teh Rancabali ada pada kisaran 7-27OC, lebih rendah
dari Gunung Mas dan Nirmala.
Selain suhu, persebaran lumut juga dapat dipengaruhi oleh kelembapan
(Richards 1984; Glime 2007). Kelembapan penting bagi lumut karena sel-sel lumut
umumnya memiliki lapisan kutikula yang sangat tipis atau sama sekali tidak memiliki
kutikula (Gradstein et al. 2001). Perkebunan teh Rancabali memiliki kisaran
kelembapan harian 75-93%, lebih tinggi dari dua perkebunan teh lainnya.
Chantanaorrapint (2010) melaporkan meningkatnya kekayaan jenis lumut di hutan
tropik Thailand menyertai peningkatan kelembapan dan intensitas cahaya, serta
penurunan suhu seiring peningkatan elevasi pada kisaran 400-1300 m dpl.
Gunung Mas dengan kekayaan jenis lumut epifit paling rendah dibandingkan
dua perkebunan teh lainnya, terletak pada elevasi paling rendah. Perkebunan teh
24
lalu lintas kendaraan bermotornya padat. Harahap (2004) melaporkan terdapat
akumulasi timbal pada organ vegetatif tanaman teh di Gunung Mas, baik pada daun,
batang, maupun pada akar. Timbal merupakan polutan berupa partikel yang berasal
dari gas buangan kendaraan bermotor. Gas buangan yang berasal dari kendaraan bermotor ataupun dari kawasan industri dapat terakumulasi dalam sel-sel lumut.
Saxena et al. (2008) menemukan akumulasi beberapa jenis logam berat pada sel-sel
lumut yang tumbuh di lokasi dengan aktivitas kendaraan bermotor yang tinggi.
Hallingback (2008) melaporkan, adanya akumulasi gas buangan di udara
menyebabkan banyak jenis lumut epifit yang menghilang dari berbagai kota besar di
beberapa negara industri.
Rata-rata penutupan tanaman teh oleh lumut di Nirmala paling rendah
dibandingkan dua perkebunan teh lainnya. Hal ini diduga berhubungan dengan
ukuran tajuk tanaman teh. Kemungkinan kondisi kerapatan tajuk antar tanaman dapat
mempengaruhi iklim mikro (suhu, kelembapan, dan intensitas cahaya) di bawah
tajuk. Kondisi seperti ini dapat dibandingkan dengan kondisi di hutan alam. Menurut
Hallingback (2008) persentase penutupan kanopi mempengaruhi kondisi iklim mikro
di bawahnya. Sporn et al. (2010) dan Ariyanti et al. (2008) melaporkan adanya
perbedaan komunitas lumut epifit pada beberapa tipe habitat dengan persentase
penutupan kanopi berbeda.
Komunitas lumut epifit di Rancabali dan Gunung Mas memiliki kesamaan
komposisi lebih rendah (38%) dibandingkan antara Rancabali dan Nirmala (51%),
maupun antara Nirmala dan Gunung Mas (54%). Hal ini menunjukkan bahwa
semakin besar perbedaan elevasi antar lokasi semakin kecil kesamaan komunitas
lumut antar keduanya. Hasil yang sama dilaporkan oleh Chantanaorrapint (2010)
pada komunitas hutan alam di sepanjang gradasi elevasi di Thailand. Rancabali
terletak pada elevasi di atas 1600 m dpl, sedangkan Nirmala di 1150 m dpl, dan
Gunung Mas di 600 m dpl. Faktor yang diduga mempengaruhi perbedaan komposisi
jenis antar komunitas di lokasi dengan perbedaan elevasi yang besar adalah adanya
perbedaan suhu, kelembapan, dan intensitas cahaya yang menghasilkan perbedaan
elevasi terbatas sementara ada pula jenis-jenis lumut yang dapat dijumpai pada
kisaran elevasi yang lebih luas. Selain itu, jenis lumut yang sama dapat dijumpai
dengan kemelimpahan berbeda di habitat dengan elevasi berbeda.
Sebanyak 74 jenis lumut epifit dijumpai di Rancabali, 41 jenis di antaranya tidak ditemukan di lokasi perkebunan teh lainnya, seperti lumut hati Radula
madagascariensis dan Lejeunea discreta. Beberapa peneliti melaporkan bahwa kedua
jenis lumut tersebut merupakan jenis yang persebaran alaminya di vegetasi hutan
pegunungan (di atas 1500 m dpl). Yamada (1979) melaporkan persebaran R.
madagascariensis di Asia pada elevasi 1400 m dpl sampai 2600 m dpl. Zhu dan So
(2001) melaporkan di China L. discreta dijumpai pada elevasi 1050 m dpl dan 2200
m dpl. Sementara itu, Acanthocoleus javanicus dijumpai di Gunung Mas dan
Rancabali, namun dengan kemelimpahan berbeda. Di Rancabali A. javanicus
merupakan jenis lumut hati paling dominan dengan nilai INP di atas 20%, sedangkan
di Gunung Mas jenis tersebut memiliki INP kurang dari 4%. Sematophyllum
tristiculum merupakan jenis lumut sejati paling dominan di Gunung Mas dengan INP
34%. Jenis tersebut juga dijumpai di Nirmala tetapi dengan INP kurang dari 1%, dan
tidak dijumpai di Rancabali. Hal ini menunjukkan persebaran altitudinal jenis
tersebut yang terbatas pada elevasi rendah (kurang dari 1000 m dpl). Pollatwan
(2008) melaporkan persebaran altitudinal S. tristiculum di Thailand yakni pada
700-900 m dpl.
Penelitian keanekaragaman lumut di hutan alam di sepanjang gradien
altitudinal telah digunakan untuk membuat zonasi ketinggian berdasarkan beberapa
jenis indikator yang dijumpai. Chantanaorrapint (2010) mengelompokkan komunitas
lumut pada elevasi yang berbeda berdasarkan kehadiran jenis-jenis tertentu.
Chantanaorrapint (2010) juga membagi vegetasi hutan tropik di Thailand ke dalam
zona hutan dataran rendah (0-600 m dpl), hutan subpegunungan (800-1000 m dpl),
dan hutan pegunungan (1200-1300 m dpl) berdasarkan diversitas jenis lumut dan data
iklim mikro.
Perkebunan Rancabali memiliki diversitas lumut paling tinggi dibandingkan
26
paling tinggi dan jenis dominan di perkebunan teh tersebut memiliki INP kurang dari
25%. Di Rancabali dijumpai 38 jenis lumut sejati dan 36 jenis lumut hati, namun
tidak ada jenis lumut sejati yang memiliki nilai INP lebih dari 10%. Nilai INP paling
tinggi (23%) di Rancabali dimiliki oleh jenis dari lumut hati. Kondisi lingkungan seperti suhu dan kelembapan yang ideal untuk pertumbuhan lumut diduga
menyebabkan lebih banyak jenis lumut yang dijumpai di lokasi tersebut, dan
masing-masing berkompetisi sehingga tidak ada jenis yang kemelimpahannya sangat berbeda
dari jenis lainnya.
Kekayaan lumut epifit di perkebunan teh lebih rendah dibandingkan hutan
alam. Pada penelitian ini ditemukan sebanyak 90 jenis lumut epifit dari tiga
perkebunan teh dan rata-rata dijumpai 7-15 jenis per tanaman teh. Sementara itu di
hutan alam Taman Nasional Gunung Halimun, lokasi yang sangat dekat dengan
perkebunan teh Nirmala, dijumpai 116 jenis lumut epifit dengan rata-rata 11-19 jenis
per pohon (Ariyanti & Sulistijorini 2011). Dari beberapa penelitian juga
menunjukkan kekayaan jenis lumut yang lebih rendah di tipe perkebunan monokultur
lainnya dibandingkan hutan alam yang heterogen, seperti di perkebunan karet dan
cengkeh (Daniels & Kariyappa 2007), perkebunan kelapa sawit (Suleiman et al.
2009), dan perkebunan kakao (Sporn et al. 2010).
Perkebunan teh merupakan tipe habitat yang terbuka (hanya ada beberapa
pohon sebagai wind breaker) dan bersifat homogen (jenis dan ukuran tanaman
seragam). Sedangkan hutan alam merupakan habitat yang heterogen dengan kanopi
yang lebih tertutup. Hutan alam kawasan Taman Nasional Gunung Halimun Salak
memiliki keanekaragaman pohon sebanyak 46 jenis per hektar (Polosakan 2011).
Hutan alam dengan keanekaragaman jenis yang dimiliki dan adanya stratifikasi
tegakan, menyediakan habitat yang lebih beragam. Selain itu, beberapa penelitian
melaporkan terdapat hubungan kekayaan dan komposisi jenis lumut dengan tekstur,
kelembapan dan pH kulit pohon, diameter pohon inang dan jenis pohon inang
(Gradstein et al. 2001; Gradstein et al. 2003; Mezaka & Znotina 2006; Gradstein &
Berdasarkan preferensi terhadap kondisi habitat yang terbuka atau terlindung,
jenis lumut dikelompokkan ke dalam shade epiphyte, sun epiphyte dan generalist
(Gradstein 1992). Lumut yang lebih menyukai habitat ternaung seperti di lantai hutan
dikelompokkan sebagai shade epiphyte. Lumut yang menyukai habitat dengan intensitas cahaya yang tinggi seperti di kanopi hutan dikelompokkan sebagai sun
epiphyte. Sedangkan lumut yang menyukai hidup di kedua tipe habitat tersebut
dikelompokkan sebagai generalist.
Jenis-jenis lumut yang hidup pada kondisi iklim mikro yang berbeda dapat
mengembangkan berbagai adaptasi struktural. Perkebunan teh dengan kondisi terbuka
tanpa ternaungi oleh kanopi menyebabkan lumut harus dapat beradaptasi dengan
intensitas cahaya tinggi. Beberapa jenis lumut yang dijumpai pada penelitian ini
dengan nilai INP lebih dari 20% memiliki beberapa karakter adaptasi sebagai lumut
xerofit yang sering dijumpai pada kelompok lumut sun epiphyte. Lumut sejati
Sematophyllum tristiculum dan Eurhynchium celebicum memiliki dinding sel yang
tebal. Menurut Richards (1984), beberapa karakter xerofit pada lumut sejati antara
lain memiliki sel berdinding tebal, permukaan daun berpapil. Sedangkan pada lumut
hati antara lain tepi daun hyaline seperti sering dijumpai pada anggota Lejeuneaceae.
Adanya sel alar pada S. tristiculum dan adanya kantong air pada daun L flava, L.
anisophylla, R. madagascariensis, dan A. javanicus, mungkin merupakan karakter
yang berhubungan dengan adaptasi untuk dapat memanfaatkan kelembapan
lingkungan dengan lebih baik. Selain itu, bentuk perawakan lumut (life-forms)
merupakan karakter morfologi yang juga penting dalam adaptasi (Richards 1984;
Chantanaorrapint 2010). Lejeunea flava, L. anisophylla merupakan lumut hati
berdaun yang berukuran kecil, tumbuh melekat kuat pada kulit batang atau di antara
lumut lain yang ukurannya lebih besar. Sementara itu, Metzgeria sp.1 merupakan
lumut hati bertalus, sering dijumpai membentuk bantalan di pangkal batang. Richards
(1984) menjelaskan bahwa ukuran yang kecil berupa helaian seperti rambut, dan
hidup melekat pada substrat, merupakan karakter adaptasi xerofit yang sering
SIMPULAN
Keanekaragaman lumut epifit yang ditemukan di tiga perkebunan teh di Jawa Barat sebanyak 90 jenis, meliputi 42 jenis lumut hati dan 48 jenis lumut sejati yang termasuk dalam 48 marga dan 25 suku. Lejeuneaceae merupakan suku dengan jumlah jenis tertinggi (26 jenis), diikuti Sematophyllaceae (9 jenis), dan Frullaniaceae (7 jenis). Perkebunan teh Rancabali memiliki diversitas lumut paling tinggi, diikuti Nirmala dan Gunung Mas. Diversitas lumut epifit meningkat dengan pertambahan elevasi. Komunitas lumut epifit di Rancabali dan Gunung Mas memiliki kesamaan komposisi jenis yang lebih rendah dibandingkan antara Rancabali dan Nirmala, maupun antara Nirmala dan Gunung Mas. Hal ini menunjukkan bahwa semakin besar perbedaan elevasi antar lokasi semakin kecil kesamaan komunitas lumut antar keduanya. Pada penelitian ini ditemukan lumut
sejati Sematophyllum tristiculum sebagai jenis lumut epifit yang dominan di Gunung Mas, lumut hati Lejeunea anisophylla dominan di Nirmala, dan lumut hati Acanthocoleus javanicus dominan di Rancabali. Lejeunea anisophylla, L.
flava, dan Metzgeria sp.1 merupakan jenis-jenis lumut epifit yang paling umum di
AprianaD. 2010. Keragaman dan Kelimpahan Lumut Hati Epifit di Kebun Raya Bogor [skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Ariyanti NS, Bos MM, Kartawinata K, Tjitrosoedirdjo SS, Guhardja E, Gradstein SR. 2008. Bryophytes on tree trunks in natural forests, selectively logged forests and cacao agroforests in Central Sulawesi, Indonesia. Biol
Conserv. 141: 2516-2527.
Ariyanti NS, Sulistijorini. 2011. Contrasting arboreal and terrestrial bryophytes communities of the Mount Halimun Salak National Park, West Java.
Biotropia 2: 81-93.
Asakawa Y. 2008. Marchantiophyta: A good source of biologically active compounds. Di dalam: Mohamed H, Baki BB, Nasrulhaq-Boyce A, Lee PKY, editor. Bryology in the New Millenium. Proceedings of the World
Bryology Conference 2007 Kuala Lumpur Malaysia; Petaling Jaya, 23-27
Jul 2007. Kuala Lumpur: University of Malaya. hlm367-394.
Bartram EB. 1939. Mosses of the Philippines. The Philip. J Sci. 68: 1-425.
Beike AK. 2010. Applied Bryology-Bryotechnology. Trop Bryol. 31: 22-32.
Bodade RG, Borkar PS, Arfeen S, Khobragade CN. 2008. Invitro screening of bryophytes for antimicrobial activity. J. of Med. Plants. 7: 23-28.
Bruun et al. 2006. Effects of altitude and topography on species richness of plants,
bryophytes and lichens in alpine communities. J. of Veget. Sci. 17: 37-46.
Buck WR, Goffinet B. 2002. Morphology and Classification of Mosses. Di dalam Shaw AJ, Goffinet B, editor. Bryophytes Biology. Cambridge: Cambridge
University Press. hlm 71-123.
Chantanaorrapint S. 2010. Ecological studies of epiphytic bryophytes along altitudinal gradient in Southern Thailand [disertasi]. Bonn: Naturnissenschaftlichen Fakultat der
Rheineschen-Friedrich-Wilhelms-Universitat Bonn.
Crandall-Stotler B, Stotler RE. 2002. Morphology and classification of the Marchantiophyta. Di dalam Shaw AJ, Goffinet B, editor. Bryophytes
Biology. Cambridge: Cambridge University Press. hlm 21-70.
30
Dey A, De JN. 2012. Antioxidative potential of bryophytes: Stress tolerance and commercial perspective: A review. Pharmacologia 3: 151-159.
Eddy A. 1988. A Handbook of Malesian Mosses. Vol.1. Sphagnales to Dicranales.
London: Natural History Museum Publication.
Eddy A. 1990. A Handbook of Malesian Mosses. Vol.2. Leucobryaceae to
Buxbaumiaceae. London: Natural History Museum Publication.
Eddy A. 1996. A Handbook of Malesian Mosses. E Vol.3. Splachnobryaceae to
Leptostomataceae. London: Natural History Museum Publication.
Enroth J. 1990. Altitudinal zonation of bryophytes on the Huon Peninsula, Papua New Guinea. A floristic approach, with phytogeographic considerations.
Trop Bryol. 2: 61-90.
Glime, JM. 2007. Bryophyte Ecology. Volume 1. Physiological Ecology.
Ebooksponsored by Michigan Technological University and the International Association of Bryologists. accessed on November 13rd, 2011 at <http://www.bryoecol.mtu.edu/>.
Goffinet B. 2002. Origin and phylogenetic relationships of bryophytes. Di dalam Shaw AJ, Goffinet B, editor. Bryophytes Biology. Cambridge: Cambridge
University Press. hlm 124-149
Gradstein SR. 1992. Threatened bryophytes of the neotropical rain forest: a status report. Tropic Bryol. 6: 83-93.
Gradstein SR, Culmsee H. 2010. Bryophytes diversity on tree trunks in montane forests of Central Java. Tropic Bryol 31: 95-105.
Gradstein SR et al. 2005. A catalogue of the bryophytes of Sulawesi, Indonesia.
J. Hattori Bot. Lab. 98: 213-257.
Gradstein SR et al. 2010. Bryophytes of Mount Patuha, West Java, Indonesia.
Reinwardtia 13: 107-123.
Gradstein SR, PÖcs T. 1989. Bryophytes. Di dalam: Lieth H, Werger MJA, editor.
Tropical rainforest ecosystems. Amsterdam: Elsevier Science Publishers.
hlm 311-325.
Grau O, Gitnes J, Birks HJB. 2007. A comparison of altitudinal species richness patterns of bryophytes with other plant groups in Nepal, Central Himalaya.
J. Biogeogr. 34: 1907-1915.
Hallingback T. 2008. Bryophytes and the Global Plant Conservation Strategy. Di dalam: Mohamed H, Baki BB, Nasrulhaq-Boyce A, Lee PKY, editor.
Bryology in the New Millenium. Proceedings of the World Bryology
Conference 2007 Kuala Lumpur Malaysia; Petaling Jaya, 23-27 Jul 2007.
Kuala Lumpur: University of Malaya. hlm405-424.
Harahap H. 2004. Pengaruh Pencemaran Timbal dari Kendaraan Bermotor dan Tanah Terhadap Tanaman dan Mutu Teh [disertasi]. Bogor: Program Pascasarjana, Institut Pertanian Bogor.
Haerida I, Gradstein SR, Tjitrosoedirdjo SS. 2010. Lejeuneaceae subfam. Ptychanthoideae (Hepaticeae) in West Java. Gard. Bull. Singapore 62:
59-111. .
Ho BC, Tan BC, Hernawati NS. 2006. A check list of mosses of Sumatera, Indonesia. J.Hattori Bot. Lab. 100: 143-190.
Junita N. 2010. Lumut Sejati Epifit pada Pangkal Pohon di Kebun Raya Bogor [Skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Magurran AE. 1987. Ecological Diversity and Its Measurement. Princeton:
Princeton University Press.
Mezaka A, Znotina V. 2006. Epiphytic bryophytes in old growth forests of slopes, screes and ravines in north-west Latvia. Acta Universitatis Latviensis 710:
13-116.
Mueller-Dumbois D, Ellenberg H. 1974. Aims and Methods of Vegetation
Ecology. New York : John Wiley & Sons.
Mues R. 2002. Chemical constituents and biochemistry. Di dalam: Shaw AJ, Goffinet B, editor. Bryophytes Biology. Cambridge: Cambridge University
Press. hlm. 150-181
Ohsawa M. 1982. Weeds of Tea plantations. Di dalam Holzner W, Numata M, editor. Biology and Ecology of Weeds. London: Dr.W.Junk Publisher. hlm
435-448.
32
Pollatwan R. 2008. Systematic treatment of Sematophyllaceae (Musci) in Thailand [Disertasi]. Bonn: Naturwissenschaftlichen Fakultat der Rheinischen- Friedrich-Willhems-Universitat Bonn.
Polosakan R. 2011. Keanekaragaman jenis pohon dan potensi pemanfaatan kayu oleh masyarakat di kawasan Taman Nasional Gunung Halimun Salak – Sukabumi. Berk.Penel.Hayati Edisi Khusus: 5A 67-72.
Renzaglia KS, Vaughn KC. 2002. Anatomy, development, and classification of hornworts. Di dalam Shaw AJ, Goffinet B. Bryophytes Biology.
Cambridge: Cambridge University Press. hlm 1-20.
Richards PW. 1984. The Ecology of Tropical Forest Bryophytes. Di dalam Schuster RM, editor. New Manual of Bryology. Tokyo: The Hattori
Botanical Laboratory 2: 1233-1270.
Saxena DK, Singh S, Srivastava K. 2008. Biomapping studies from India: Use of moss Isopterygium elegans for measuring seasonal metal precipitation in
Garhwal by transplant technique. Di dalam: Mohamed H, Baki BB, Nasrulhaq-Boyce A, Lee PKY, editor. Bryology in the New Millenium. Proceedings of the World Bryology Conference 2007 Kuala Lumpur
Malaysia; Petaling Jaya, 23-27 Jul 2007. Kuala Lumpur: University of
Malaya. hlm353-365.
Schiffner V. 1900. Die Hepaticae der Flora von Buitenzorg. Leiden: Erster Band. Brill.
SÖderstrÖm L, Gradstein SR, Hagborg A. 2010. Checklist of the hornworts and
liverworts of Java. Phytotaxa 9: 53-149.
Sporn SG, Bos MM, Gradstein SR. 2007. Is productivity of cacao impeded by
epiphytes? An experimental approach. Agriculture, Ecosystem and
Environment 122: 490-493.
Sporn SG, Bos MM, Kessler M, Gradstein SR. 2010. Vertical distribution of epiphytic bryophytes in an Indonesia rainforest. Biodivers.Conserv. 19:
745-760.
Tan BC, PÖcs T. 2002. Bryogeography and conservation of brophytes. Di dalam
Shaw AJ, Goffinet B, editor. Bryophytes Biology. Cambridge: Cambridge University Press. hlm. 403-448.
Verdoorn F.1930. Die Frullaniaceae der Indomalesischen Inseln. Annales
Bryologici Supplement I: 1-187.
Wahyuni I. 2010. Lumut Terestrial dan Ganggang Asosiasinya di Kebun Raya Bogor [Skripsi]. Bogor: Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Yamada K. 1979. A revision of asean taxa of Radula, Hepaticae. Journ.Hattori
Bot. Lab. 45: 201-322.
Zhu R-L, Gradstein SR. 2005. Monograph of Lopholejeunea (Lejeuneaceae, Hepaticae) in Asia. Syst. Bot. Monograph 74: 1-98.
ABSTRACT
HILDA AKMAL. Diversity of Epiphytic Bryophytes at Three Tea Plantations in West Java. Supervised by SRI SUDARMIYATI TJITROSOEDIRDJO, NUNIK SRI ARIYANTI and SULISTIJORINI.
Study on the epiphytic bryophytes was conducted at three tea plantations located at different elevation in West Java: Gunung Mas (600 m) Bogor, Nirmala (1150 m) Sukabumi, Rancabali (1628 m) Bandung. Three plots were established within each tea plantation, three transects of 30 m were lined in each plot. Samples were collected from the trunk and primary branches of five tea trees in each line transect. The aims of this study were to record the epiphytic bryophytes species of the plantations and to compare diversity of the bryophytes community among the plantations. Ninety species of epiphytic bryophytes were recorded. Checklist of those species were provided in this paper. The bryophytes species richness of the plantations increased with increasing of the elevation. Twenty seven species were found at Gunung Mas, 40 species at Nirmala, and 74 species at Rancabali. The plantations were dominated by different species. The dominant species were Sematophyllum tristiculum (mosses) at Gunung Mas, Lejeunea
anisophylla (liverworts) at Nirmala, and Acanthocoleus javanicus (liverworts) at
Rancabali. The species composition of the bryophytes communities of Gunung Mas was more similar to that of Nirmala compared to that of Rancabali.
RINGKASAN
HILDA AKMAL. Diversitas Lumut Epifit di Tiga Perkebunan Teh Jawa Barat. Dibawah bimbingan SRI SUDARMIYATI TJITROSOEDIRDJO, NUNIK SRI ARIYANTI dan SULISTIJORINI.
Lumut (bryophyte) merupakan kelompok tumbuhan terbesar kedua setelah Angiosperma. Hutan hujan di kawasan tropik merupakan salah satu habitat dengan keanekaragaman lumut yang tinggi. Penelitian tentang flora lumut di pulau Jawa telah berlangsung lama, yaitu sejak hampir dua ratus tahun yang lalu. Sesudah tahun 1960 hanya sedikit studi floristik tentang lumut yang dilakukan di pulau Jawa. Dari beberapa penelitian lumut akhir-akhir ini masih diperoleh catatan baru untuk flora lumut di Jawa. Hal ini menunjukkan bahwa data tentang flora lumut di pulau Jawa sampai saat ini masih belum lengkap. Penelitian dan eksplorasi yang ada umumnya dilakukan pada komunitas lumut di hutan alam, penelitian terhadap lumut di luar kawasan hutan alam masih sangat jarang.
Penelitian ini bertujuan untuk mencatat keanekaragaman jenis lumut epifit di tiga perkebunan teh pada elevasi berbeda di Jawa Barat, dan membandingkan diversitas lumut epifit pada perkebunan teh tersebut. Keanekaragaman jenis lumut disajikan dalam bentuk daftar jenis (checklist) lumut epifit di tiga perkebunan teh. Diversitas komunitas lumut epifit dibandingkan berdasarkan kekayaan, komposisi, dan dominansi jenis, serta kemelimpahan total lumut.
Pengambilan sampel lumut dilakukan di tiga perkebunan teh yaitu di Gunung Mas (600 m dpl), Nirmala (1150 m dpl), dan Rancabali (1628 m dpl). Di setiap perkebunan teh dibuat tiga plot penelitian masing-masing berukuran 30 m x 20 m, jarak antar plot minimal 500 m. Pada setiap plot dibuat tiga transek, jarak antar transek 5 m. Pada setiap transek ditentukan 5 sampel tanaman teh, berjarak 5 m. Persentase penutupan oleh setiap jenis lumut dan penutupan oleh total lumut pada setiap sampel tanaman teh dicatat. Sebagai data pendukung dicatat pula penutupan tanaman teh oleh liken dan tumbuhan paku. Sampel lumut diambil dari batang dan cabang primer pada setiap sampel tanaman teh. Sampel lumut dari lokasi diidentifikasi dengan mengamati ciri-ciri spesifik gametofit dan sporofitnya menggunakan mikroskop, dilanjutkan dengan dokumentasi.
kemelimpahan totalnya sangat berbeda dari jenis-jenis lainnya. Komunitas lumut epifit di Rancabali dan Gunung Mas memiliki kesamaan komposisi yang paling kecil (38%) dibandingkan antara Rancabali dan Nirmala (51%), maupun antara Nirmala dan Gunung Mas (54%). Hal ini menunjukkan bahwa semakin besar perbedaan elevasi antar lokasi semakin kecil kesamaan komunitas lumut antar keduanya. Di perkebunan teh Rancabali dijumpai 74 jenis lumut epifit, 41 jenis di antaranya tidak ditemukan di lokasi perkebunan lainnya. Acanthocoleus javanicus
merupakan jenis lumut hati yang dominan di Rancabali, Lejeunea anisophylla di Nirmala, dan Sematophyllum tristiculum dominan di Gunung Mas. Rata-rata
penutupan tanaman teh oleh lumut paling rendah di Nirmala. Hal ini diduga berhubungan dengan ukuran tajuk tanaman teh