KEANEKARAGAMAN CENDAWAN PADA RIZOSFER
KELAPA SAWIT DAN PALEM LIAR
KAVY SHOBAH
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Keanekaragaman Cendawan pada Rizosfer Kelapa Sawit dan Palem Liar adalah benar karya saya dengan arahan dari dosen pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2015
Kavy Shobah
KAVY SHOBAH. Keanekaragaman Cendawan pada Rizosfer Kelapa Sawit dan Palem Liar. Dibimbing oleh EFI TODING TONDOK.
Kelapa sawit merupakan salah satu komoditi utama perkebunan di Indonesia. Penyakit Busuk Pangkal Batang (Ganoderma boninense) merupakan penyakit utama pada kelapa sawit yang dapat menyebabkan kerugian sebesar 80%. Rizosfer adalah tanah di sekitar perakaran tanaman dan merupakan tempat yang ideal bagi tumbuh dan berkembangnya mikroba tanah termasuk di dalamnya agens antagonis. Penelitian ini bertujuan mengetahui keanekaragaman cendawan pada rizosfer kelapa sawit dan palem liar dan mengevaluasi kemampuannya dalam menekan perkembangan G. boninense secara in vitro. Sampel tanah berasal dari Jambi, yaitu perkebunan kelapa sawit di Bukit Harapan (H04 a, b, c), perkebunan kelapa sawit di Bukit Duabelas (B04 a, b, c), palem liar di dalam hutan karet (PLK), di dekat area perkebunan karet (PLdK) dan di perkebunan kelapa sawit (PLS). Keanekaragaman dan kelimpahan cendawan dianalisis dengan metode pengenceran dan pencawanan. Kelimpahan cendawan tanah tertinggi diperoleh dari sampel kelapa sawit B04 a sebesar 4.9 x 106 cfu/g sedangkan kelimpahan cendawan terendah diperoleh dari sampel kelapa sawit B04 b sebesar 0.3 x 105 cfu/g. Keanekaragaman cendawan dari yang tertinggi ke terendah adalah
palem liar (8 genus dan 13 UF), H04 (6 genus dan 9 UF), dan B04 (4 genus dan 6 UF). Isolat yang berpotensi sebagai agens antagonis adalah isolat Trichoderma sp. 2 dengan persentase penghambatan terhadap cendawan patogen G. boninense
dengan persentase 76%. Isolat ini ditemukan pada rizosfer palem liar (PLS dan PLdK) dan kelapa sawit (H04 a dan H04 c), selain itu didapatkan beberapa isolat
(Chrysosporium sp. dan Penicillium sp. 13) yang dapat menghasilkan antibiosis.
KAVY SHOBAH. Fungal diversity in the Rhizosphere of Oil Palm and Wild Palm. Guided by EFI TODING TONDOK.
Oil palm is a major estate crops in Indonesia. Basal Stem Rot (Ganoderma
boninense) is an important disease of oil palm which may cause 80% yield losses.
Rhizosphere is an ideal habitat for soil microorganisms to live, including antagonist agents. The aim of this research were to explore fungal diversity from rhizosphere of oil palms and wild palms and to evaluate their ability to suppress the development of G. boninense in-vitro. Soil samples were taken from Jambi province in two groups of oil palm, Bukit Harapan (H04 a,b,c) and Bukit Duabelas (B04 a,b,c) for oil palm plantations, forest rubber (PLK), forest near rubber plantation (PLdK), in oil palm plantation (PLS) for wild palms. Diversity and abundance of fungal were observed by serial dilution and plating method. The highest abundance of soil fungi was found from oil palm B04 a with total of 4.9 x 106 cfu/g and the lowest was found from oil palm plantation B04 b with total of 0.3 x 105 cfu/g. The diversity of soil fungi from the highest to the lowest was as
follows, wild palm (8 genus and 13 UF), H04 (6 genus and 9 UF), and B04 (4 genus and 6 UF). Trichoderma sp. 2 isolate was the most potentially applied as biocontrol agents with 76% growth inhibition of G. boninense. This isolate was found at wild palms (PLS and PLdK) and oil palms (H04 a dan H04 c) rizhosphere. Some isolated fungi (Chrysosporium sp. and Penicillium sp. 13) also have antibiosis activities.
©Hak Cipta milik IPB, tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
KEANEKARAGAMAN CENDAWAN PADA RIZOSFER
KELAPA SAWIT DAN PALEM LIAR
KAVY SHOBAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
Judul Skripsi : Keanekaragaman Cendawan pada Rizosfer Kelapa Sawit dan Palem Liar
Nama Mahasiswa : Kavy Shobah
NIM : A34090075
Disetujui oleh
Dr. Efi Toding Tondok, SP. M.Sc. Dosen Pembimbing
Diketahui oleh
Dr. Ir. Abdjad Asih Nawangsih, M.Si. Ketua Departemen Proteksi Tanaman
PRAKATA
Alhamdulillah puji dan syukur penulis panjatkan ke hadirat Allah SWT yang telah melimpahkan segala rahmat dan karunia-Nya sehingga penulis mampu
menyelesaikan tugas akhir yang berjudul “Keanekaragaman Cendawan pada Rizosfer Kelapa Sawit dan Palem Liar”. Penulisan tugas akhir penelitian ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Pertanian di Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor.
Dengan terselesaikannya penulisan skripsi ini, penulis mengucapkan terima kasih yang sebesar-besarnya kepada Dr. Efi Toding Tondok, SP. M.Sc. sebagai dosen pembimbing skripsi yang selalu memberikan bimbingan, pengetahuan, saran, arahan, dan masukan kepada penulis. Ucapan terima kasih juga diucapkan kepada Prof. Dr. Ir Aunu Rauf M.Sc. sebagai dosen pembimbing akademik yang telah membimbing penulis selama ini. Terima kasih kepada Dr. Ir. Nina Maryana, M.Si. sebagai dosen penguji tamu dan Dr. Ir. Giyanto, M.Si. sebagai dosen moderator pada seminar tugas akhir saya yang telah memberikan kritik dan saran untuk menyempurnakan penulisan tugas akhir ini serta Dr. Ir. Titiek Siti Yuliani, S.U yang telah banyak membantu dan memberikan semangat kepada penulis ketika penulis sedang sakit.
Terima kasih kepada orangtua penulis Bapak Mustaman dan Ma’e Zahroh serta kakak-kakak dan adik yang selalu memberikan doa semangat dan dukungan dalam belajar. Ucapan terima kasih juga ditujukan kepada teman-teman Laboratorium Mikologi Tumbuhan, teman-teman Proteksi Tanaman 46, Proteksi Tanaman 47 khususnya Ridho, Fahmi, Iwan, Nazir, Doni, Nce serta kakak tingkat dan juga teman-teman lainnya di Departemen Proteksi Tanaman yang tidak bisa disebutkan satu per satu dalam mendukung terlaksananya tugas akhir penelitian penulis, serta pihak lain yang turut mambantu dalam penyusunan tugas akhir ini.
Pada penulisan tugas akhir ini penulis menyadari masih banyak kekurangan. Oleh karena itu, penulis berharap ada masukan, kritik dan saran yang bersifat membangun dan memotivasi penulis agar dapat menuliskan karya tulis yang lebih baik. Semoga Skripsi ini dapat bermanfaat bagi penulis maupun pembaca.
DAFTAR ISI
Penghitungan Keanekaragaman dan Kelimpahan Cendawan 3
Uji Antagonisme in vitro 4
Identifikasi 4
Rancangan Percobaan dan Analisis Data 4
HASIL DAN PEMBAHASAN 5
Keanekaragaman dan Kelimpahan Cendawan 5
Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Palem Liar 6 Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Kelapa Sawit di
Bukit Harapan 8
Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Kelapa Sawit di
Bukit Duabelas 8
Uji Antagonisme Cendawan Rizosfer terhadap G. boninense in vitro 9
Unidentified fungi 9 15
KESIMPULAN DAN SARAN 22
Kesimpulan 22
Saran 22
DAFTAR PUSTAKA 23
LAMPIRAN 25
1 Kelimpahan dan keanekaragaman cendawan pada setiap sampel 6
2 Keberadaan cendawan tanah di setiap sampel 7
3 Persentase penghambatan cendawan uji terhadap G. boninense pada 5
hsp 11
4 Hasil identifikasi cendawan rizosfer 15
DAFTAR GAMBAR
1 Hasil pencawanan pada media MA dari pengenceran 10-3 setiap sampel: (a) PLK, (b) PLS, (c) PLdK, (d) H04 a, (e) H04 b, (f) H04 c, (g) B04 a,
(h) B04 b, (i) B04 c 5
2 Kelimpahan cendawan pada rizosfer palem liar 8
3 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Harapan 9 4 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Duabelas 9 5 Morfologi G. boninense: (a) makroskopi, (b) clamp connection 10 6 Hasil uji antagonisme cendawan uji dengan G. boninense pada 5 hsp: (a)
kontrol, (b) kompetisi (Trichoderma sp.2), (c) antibiosis (Penicillium sp.
13), (d) seperti zona bening (Uf 7) 11
DAFTAR LAMPIRAN
1 Beberapa isolat cendawan yang berhasil diisolasi dari tanah tetapi
terkontaminasi sehingga tidak dapat diidentifikasi lebih lanjut 26 2 Kelimpahan cendawan pada rizosfer palem liar 26 3 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Harapan 27 4 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Duabelas 27 5 Daftar sidik ragam penghambatan G. boninense oleh cendawan rizosfer 27
Latar Belakang
Kelapa sawit merupakan salah satu komoditi utama perkebunan di Indonesia. Produksi minyak sawit (crude palm oil/ CPO) di Indonesia terus mengalami peningkatan dari tahun ke tahun. Pada tahun 2010 produksi CPO sebesar 21.9 juta ton, tahun 2011 sebesar 23 juta ton, tahun 2012 sebesar 23.5 juta ton dan pada tahun 2013 diperkirakan meningkat produksinya sebesar 24.4 juta ton (Ditjenbun 2013b). Salah satu faktor yang mempengaruhi peningkatan produksi CPO yaitu semakin meluasnya areal perkebunan kelapa sawit di Indonesia. Pada tahun 2010 luas area perkebunan kelapa sawit mencapai 8.3 juta hektar dan diperkirakan meningkat menjadi 9.1 juta hektar pada tahun 2013 (Ditjenbun 2013a).
Peningkatan luas area perkebunan monokultur kelapa sawit dapat menyebabkan pengaruh buruk pada ekosistem tersebut. Salah satunya adalah banyaknya serangan patogen pada perkebunan kelapa sawit yang dapat menyebabkan penurunan produktivitas perkebunan tersebut. Patogen utama yang menyerang kelapa sawit di Indonesia dan Malaysia adalah cendawan Ganoderma
boninense yang menyebabkan penyakit Busuk Pangkal Batang (BPB). Menurut
Paterson (2007), tanaman yang terinfeksi oleh G. boninense akan terdegradasi ligninnya karena cendawan bersifat lignolitik dan lama kelamaan akan mengalami kematian. Penyakit ini telah menyebabkan kematian kelapa sawit hingga 80% di beberapa perkebunan sawit di Indonesia (Susanto 2002). Keparahan yang diakibatkan oleh cendawan ini akan terus meningkat dengan banyaknya perkebunan monokultur kelapa sawit (Turner 1981). Menurut Magurran (1987), keanekaragaman dalam suatu ekosistem sangat penting karena dapat dijadikan sebagai indikator kesehatan suatu ekosistem tersebut, termasuk di dalamnya adalah keanekaragaman mikroba tanah dalam ekosistem tersebut.
Adanya serangan cendawan patogen pada perkebunan kelapa sawit diduga oleh rendahnya keanekaragaman mikroba tanah yang dapat berkompetisi dengan patogen tersebut. Salah satu habitat mikroba tanah adalah rizosfer. Rizosfer adalah tanah di sekitar perakaran tumbuhan yang banyak mengandung nutrisi dan mikroba. Rizosfer juga merupakan daerah yang dapat mengubah keanekaragaman mikroba, meningkatkan aktivitas dan jumlah organisme, serta terdapat interaksi yang kompleks antara mikroba dengan akar. Keanekaragaman suatu mikroba dalam rizosfer sangat tinggi karena kandungan bahan organiknya melimpah yang berasal dari eksudat akar tumbuhan (Kennedy 2005).
2
Keseimbangan ekosistem tersebut disebabkan oleh banyaknya interaksi antara organisme di dalam ekosistem tersebut.
Hasil penelitian Izzati dan Abdullah (2008), serangan patogen G. boninense
pada kelapa sawit dapat ditekan dengan cendawan Trichoderma harzianum dari 70% menjadi 5%. Hasil penelitian lain menunjukkan bahwa aplikasi bakteri kitinolitik (isolat TB41 atau AL11) yang dikombinasikan dengan Fungi Mikoriza Arbuskular (FMA) hasil dari eksplorasi rizoser dapat meningkatkan kolonisasi FMA pada akar bibit sawit dan dapat menekan keparahan penyakit G. boninense
(Nildayanti 2011). Oleh karena itu, informasi mengenai keanekaragaman cendawan tanah pada rizosfer kelapa sawit dan palem liar dapat menjadi sebuah bahan untuk mengendalikan patogen pada kelapa sawit.
Tujuan Penelitian
Penelitian ini bertujuan menganalisis keanekaragaman dan kelimpahan cendawan pada rizosfer kelapa sawit dan palem liar serta potensinya sebagai agens hayati.
Manfaat Penelitian
Penelitian tentang cendawan pada rizosfer kelapa sawit dan beberapa palem liar akan memberikan informasi tentang keanekaragaman dan kelimpahan cendawan rizosfer untuk menemukan peranannya sebagai salah satu agens hayati yang dapat bermanfaat dalam penelitian selanjutnya.
Tempat dan Waktu
Penelitian ini dilaksanakan di Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman, Institut Pertanian Bogor (IPB). Sampel tanah berasal dari Bukit Harapan dan Bukit Duabelas di Provinsi Jambi. Penelitian dilaksanakan dari bulan Maret 2013 sampai Desember 2014.
Bahan dan Alat
Bahan-bahan yang digunakan dalam penelitian ini di antaranya adalah sampel tanah dari rizosfer, media Martin Agar (MA), media Potato Dextrose Agar
(PDA). Alat-alat yang digunakan antara lain cawan petri, tabung reaksi, laminar
air flow, dan mikroskop cahaya.
Metode Sampel Tanah
Sampel tanah yang digunakan berjumlah 9, yaitu 3 sampel berasal dari rizosfer perkebunan kelapa sawit di Bukit Harapan Jambi (H04 a, b, c), 3 sampel dari perkebunan kelapa sawit di Bukit Duabelas Jambi (B04 a, b, c) dan 3 sampel dari rizosfer palem liar dengan 3 ekosistem yang berbeda-beda, yaitu palem liar yang tumbuh di dalam hutan karet (PLK), palem liar yang tumbuh di hutan dekat area perkebunan karet (PLdK) dan palem liar yang tumbuh di area perkebunan sawit (PLS).
Isolasi Cendawan Tanah
Isolasi cendawan tanah dilakukan dengan metode pengenceran dan pencawanan. Sebanyak 10 g sampel tanah dilarutkan di dalam 90 ml air steril sehingga didapat suspensi tanah. Suspensi diguncang dengan menggunakan alat
orbital shaker selama 2 jam dengan kecepatan 150 rpm. Suspensi kemudian
diencerkan segera secara seri dengan cara mencampurkan 1 ml suspensi tanah dengan 9 ml air steril dalam tabung reaksi sehingga didapat pengenceran 10-2. Pengenceran terus dilakukan hingga tingkat pengenceran 10-4. Untuk pengenceran 10-3 sampai 10-4 diambil 1 ml kemudian dibiakkan dalam media MA dengan
masing-masing 3 ulangan. Hasil biakan diamati selama 7 hari setelah pencawanan (HSP). Setiap koloni cendawan yang tumbuh dicatat, dihitung jumlahnya dan dikelompokkan berdasarkan bentuk dan warna koloni kemudian dimurnikan pada media PDA.
Penghitungan Keanekaragaman dan Kelimpahan Cendawan
Keanekaragaman cendawan ditentukan dengan mengelompokkan koloni berdasarkan perbedaan bentuk koloni, warna permukaan atas dan bawah, serta tepiannya. Kelimpahan cendawan ditentukan dengan menghitung langsung koloni yang tumbuh pada media MA, kemudian dihitung per satuan colony forming unit
4
Kelimpahan cfu/g =
Uji Antagonisme in vitro
Uji antagonisme dilakukan untuk mengetahui potensi isolat cendawan yang didapatkan. Uji antagonisme dilakukan terhadap cendawan patogen G. boninense
yang diperoleh dari koleksi Laboratorium Mikologi Tumbuhan, Departemen Proteksi Tanaman IPB. Pengujian dilakukan pada media PDA dengan mengikuti metode Li et al. (2007), potongan isolat cendawan tanah berdiameter 5 mm diletakkan tepat di tengah cawan petri berdiameter 85 mm dan potongan cendawan G. boninense berdiameter 5 mm diletakkan pada media dengan jarak 21.25 mm di samping isolat cendawan rizosfer baik sisi vertikal atau horizontal, sedangkan untuk kontrol adalah media yang hanya berisi 4 potongan cendawan G.
boninense. Tiap pengujian dilakukan 3 kali ulangan. Pengamatan dilakukan
selama 7 hari dengan mengukur diameter koloni G. boninense pada kontrol (r1) dan diameter koloni G. boninense pada perlakuan (r2) dan untuk mengetahui persentase penghambatannya dihitung dengan rumus:
P = (r1˗r2) / r1 x 100%
keterangan:
P = persentase penghambatan (%)
r1 = diameter koloni cendawan patogen padakontrol r2 = diameter koloni cendawan patogen pada perlakuan Identifikasi
Koloni cendawan yang tumbuh pada media MA kemudian dimurnikan pada media PDA, lalu diidentifikasi berdasarkan morfologinya dengan bantuan kunci identifikasi (Watanabe 2002). Untuk memperoleh struktur cendawan yang lengkap, dibuat agar blok dari media PDA yang diinokulasi isolat cendawan dan diletakkan di atas kaca preparat dan diinkubasi selama 2-7 hari sebelum diamati di bawah mikroskop cahaya.
Rancangan Percobaan dan Analisis Data
Rancangan percobaan untuk uji penghambatan G. boninense secara in vitro
Keanekaragaman dan Kelimpahan Cendawan
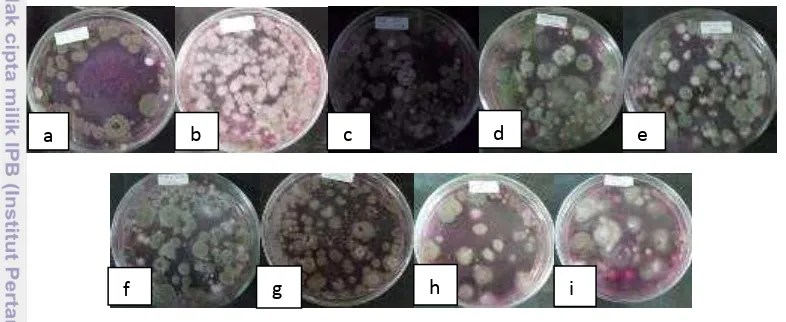
Keanekaragaman dan kelimpahan cendawan ditentukan dengan banyaknya koloni cendawan yang tumbuh pada media MA. Setiap sampel rizosfer yang diisolasi pada media MA menunjukkan hasil yang berbeda-beda. Rata-rata koloni yang tumbuh pada media MA dari pengenceran 10-3 masih cukup rapat dan banyak kecuali pada sampel B04 b, B04 c dan PLK (Gambar 1).
Gambar 1 Hasil pencawanan pada media MA dari pengenceran 10-3 setiap sampel: (a) PLK, (b) PLS, (c) PLdK, (d) H04 a, (e) H04 b, (f) H04 c, (g) B04 a, (h) B04 b, (i) B04 c
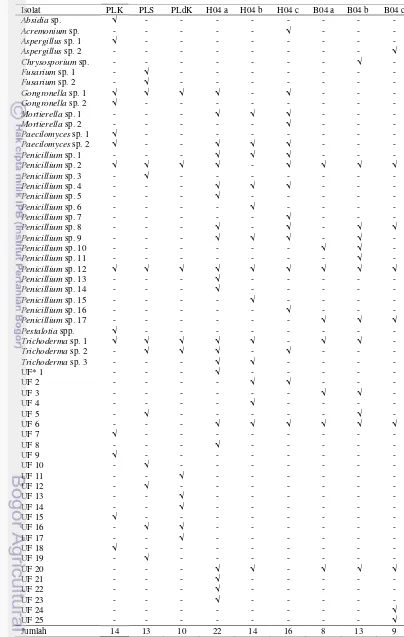
Dari 9 sampel yang diisolasi, kelimpahan cendawan tertinggi dihasilkan oleh sampel B04 a yang berasal dari rizosfer perkebunan kelapa sawit di Bukit Duabelas sebesar 4.9 x 106 cfu/g, sedangkan kelimpahan cendawan terendah berasal dari sampel B04 b sebesar 0.3 x 105 cfu/g yang juga berasal dari perkebunan kelapa sawit di Bukit Duabelas. Berdasarkan bentuk, warna dan tepian koloni, dari 9 sampel tersebut didapatkan 59 isolat cendawan. Keanekaragaman cendawan dari yang tertinggi adalah sampel dari rizosfer kelapa sawit di bukit Harapan dengan 30 macam isolat, kemudian sampel dari palem liar dengan 27 isolat, dan keanekaragaman terendah berasal dari rizosfer kelapa sawit di Bukit Duabelas dengan 16 isolat. Untuk keanekaragaman cendawan per sampel, keanekaragaman tertinggi berasal dari sampel H04 a dengan 22 isolat, sedangkan yang terendah berasal dari sampel B04 a dengan 8 isolat (Tabel 1).
Keanekaragaman mikroba tanah akan meningkat dengan beragamnya tumbuhan di ekosistem tersebut (Zak et al. 2003). Selain itu, keanekaragaman mikroba tanah juga lebih dipengaruhi oleh banyaknya senyawa organik yang dihasilkan oleh tanaman tersebut. Hasil eksplorasi cendawan tanah yang dilakukan Puangsombat et al. (2010) di 5 tipe lahan yang berbeda juga didapatkan bahwa indeks keanekaragaman cendawan di lahan nanas lebih tinggi daripada di hutan sekunder. Menurut Shi et al. (2013), keanekaragaman cendawan tanah pada berbagai tipe hutan dipengaruhi oleh suhu, garis lintang dan keanekaragaman tumbuhan. Keanekaragaman cendawan tertinggi didapatkan dari hutan dengan keanekaragaman tumbuhan terendah atau di hutan iklim sedang.
a b c d e
6
Tabel 1 Kelimpahan dan keanekaragaman cendawan pada setiap sampel
Sampel tanah Kelimpahan
Kelapa sawit Bukit Harapan 30
H04 a 3.9 x 106 22
H04 b 2.8 x 106 14
H04 c 5.8 x 105 16
Kelapa sawit Bukit Duabelas 16
B04 a 4.9 x 106 8
B04 b 0.3 x 105 13
B04 c 3.0 x 105 9
*angka estimasi. PLK: palem liar di hutan karet. PLS: palem liar di kebun sawit. PLdK: palem liar di hutan dekat kebun karet.
Semua isolat yang ditemukan, banyak yang ditemukan hanya dari satu sampel saja, dan hanya isolat Penicillium sp. 12 yang ditemukan di setiap sampel (Tabel 2). Cendawan Penicillium merupakan cendawan yang sering ditemukan di tanah sebagai saprofit. Hasil penelitian Makut dan Owolewa (2011), Penicillium
dan Aspergillus ditemukan hampir di setiap lokasi sampel. Unidentified fungi
(UF) 1-9 adalah kode untuk isolat cendawan yang belum diketahui taksonominya dan UF 10-25 adalah cendawan yang telah diisolasi tetapi terkontaminasi dan mati sebelum sempat diidentifikasi (Lampiran 1).
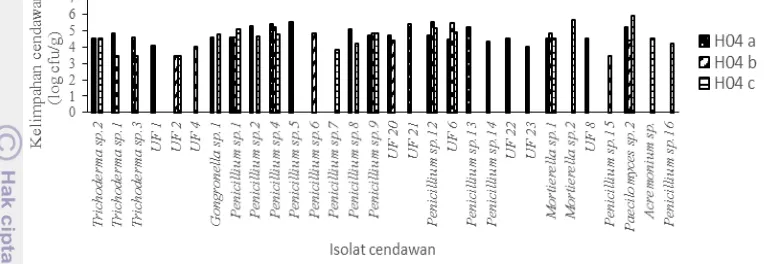
Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Palem Liar Sampel dari palem liar memiliki ekosistem yang berbeda-beda, yaitu palem liar yang tumbuh di dalam hutan karet (PLK), palem liar yang tumbuh di hutan dekat area perkebunan karet (PLdK) dan palem liar yang tumbuh di area perkebunan sawit (PLS). Kelimpahan cendawan tertinggi didapatkan dari sampel PLdK sebesar 4.4 x 106 cfu/g, dan kelimpahan terendah dari sampel PLK dengan 4.6 x 105 cfu/g. Pada sampel PLS didapatkan kelimpahan cendawan sebanyak 3.4 x 106 cfu/g (angka estimasi) (Tabel 1). Hasil kelimpahannya disebut sebagai angka estimasi karena jumlah koloni cendawan pada setiap cawan dari sampel tidak masuk dalam kategori jumlah koloni yang dapat yaitu antara 8-80 koloni (Tortora
et al. 2004; Sutton 2006). Total isolat yang didapatkan dari rizosfer palem liar berjumlah 27 macam. Keanekaragaman cendawan yang dihasilkan dari setiap sampel ini tidak terlalu berbeda, berikut urutan dari yang tertinggi, yaitu PLK (14 isolat), PLS (13 isolat), dan PLdK (10 isolat). Pada sampel PLK, isolat yang dominan adalah isolat UF 15 dengan kelimpahan 80 x 10⁴ cfu/g dan pada sampel PLS dan PLdK didominasi oleh isolat UF 16 dengan kelimpahan secara berturut- turut adalah 493 x 104 cfu/g dan 177 x 104 cfu/g (angka estimasi) (Lampiran 2). Isolat yang selalu ditemukan di ketiga sampel rizosfer palem liar adalah
Isolat PLK PLS PLdK H04 a H04 b H04 c B04 a B04 b B04 c
8
Gambar 2 Kelimpahan cendawan pada rizosfer palem liar, PLK: palem liar di hutan karet, PLS: palem liar di kebun sawit, PLdK: palem liar di hutan dekat kebun karet
Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Kelapa Sawit di Bukit Harapan
Isolat yang didapatkan dari Bukit Harapan ada 30 macam. Keanekaragaman cendawan dari yang tertinggi sampai yang terendah adalah H04 a (22 isolat), H04 c (16 isolat), dan H04 b (14 isolat) sedangkan kelimpahan cendawan dari yang tertinggi sampai terendah adalah H04 a (3.9 x 106 cfu/g), H04 b (2.8 x 106 cfu/g), dan H04 c (5.8 x 105 cfu/g). Isolat yang dominan pada sampel H04 a adalah
Penicillium sp. 5 dengan kelimpahan 33.3 x 104 cfu/g, pada sampel H04 b didominasi oleh isolat Penicillium sp. 12 dengan kelimpahan 34.3 x 104 cfu/g dan isolat yang dominan pada sampel H04 c adalah isolat Paecilomyces sp. 2 dengan kelimpahan 83.3 x 104 cfu/g (Lampiran 3). Isolat yang ditemukan pada ketiga sampel adalah Penicillium sp. 1, Penicillium sp. 4, Penicillium sp. 9, Penicillium
sp. 12, Mortierella sp. 1 Paecilomyces sp. 2 dan UF 6. Pada lokasi ini keanekaragaman genusnya lebih sedikit dari lokasi palem liar yaitu 6 genus dan 9 UF karena didominasi oleh cendawan dari genus Penicillum (Gambar 3).
Keanekaragaman dan Kelimpahan Cendawan pada Rizosfer Kelapa Sawit di Bukit Duabelas
Isolat yang diperoleh dari lokasi ini sebanyak 16 macam, jumlah ini merupakan jumlah terendah dari 2 lokasi lainnya. Sampel B04 a merupakan sampel dengan keanekaragaman terendah (8 macam isolat) dari seluruh sampel meskipun kelimpahannya adalah yang tertinggi, yaitu 4.9 x 106 cfu/g, sedangkan sampel B04 b memiliki kelimpahan terendah (0.3 x 105 cfu/g) dari seluruh sampel akan tetapi keanekaragamannya lebih tinggi dari sampel B04 a yaitu 13 macam isolat. Hal ini menunjukkan bahwa sampel dengan kelimpahan cendawan tertinggi belum tentu memiliki keanekaragaman yang tinggi juga. Ini bisa disebabkan karena adanya dominasi suatu cendawan terhadap cendawan lainnya. Sampel B04 a didominasi oleh isolat Penicillium sp. 2 dan Penicillium sp. 12 dengan kelimpahan masing-masing adalah 96.7 x 104 cfu/g dan 50.3 x 104 cfu/g sedangkan isolat lainnya kelimpahannya hanya berkisar antara 0.3-0.7 x 104 cfu/g
(Lampiran 4). Isolat yang ditemukan di setiap sampel adalah isolat Penicillium sp.
didominasi oleh genus Penicillium (Gambar 4).
Gambar 3 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Harapan
Gambar 4 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit Duabelas Uji Antagonisme Cendawan Rizosfer terhadap G. boninense in vitro
Uji antagonisme dilakukan untuk mengetahui kemampuan cendawan rizosfer dalam menghambat pertumbuhan cendawan patogen. Mekanisme penghambatan yang umum terjadi adalah antibiosis, kompetisi, mikoparasitisme, enzim pendegradasi dinding sel dan induksi resistensi. Mekanisme antagonisme dapat melibatkan semua mekanisme penghambatan atau hanya salah satu dari mekanisme tersebut. Salah satu cendawan antagonis yang dapat menghambat patogen dengan berbagai macam mekanisme antagonisme adalah cendawan dari genus Trichoderma yang dapat menghasilkan beberapa senyawa antibiotik (Gliovirin, Gliotoksin, Akyl pyrones dan Peptaibol antibioik), enzim pendegradasi dinding sel (enzim kitinolitik dan glukanase), kompetisi ruang dan nutrisi dan mikoparasitisme (Lo 1998).
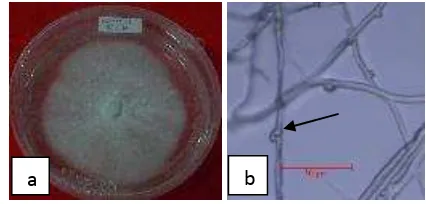
Uji antagonisme dilakukan dengan menguji isolat cendawan tanah (43 isolat) yang ditemukan dengan cendawan G. boninense. Pada media PDA, cendawan ini memiliki miselium berwarna putih, melekat di permukaan media dan perkembangannya cepat. Pada umur 9 hari, miselium cendawan ini sudah memenuhi permukaan media. Struktur hifa cendawan ini bersekat dan terdapat
10
Gambar 5 Morfologi G. boninense: (a) makroskopi, (b) clamp connection
Hasil pengujian antagonisme antara cendawan rizosfer dengan cendawan patogen G. boninense didapatkan beberapa mekanisme penghambatan. Mekanisme penghambatan yang banyak didapat adalah adanya kompetisi ruang dan nutrisi antara cendawan uji dengan G. boninense yang salah satunya ditunjukkan oleh perlakuan Trichoderma sp. 2. Pada perlakuan ini terlihat bahwa miselium dari Trichoderma sp. 2 menyebar ke seluruh permukaan media yang menyebabkan koloni dari G. boninense tidak dapat berkembang dan diduga terjadi mekanisme penghambatan yang lain karena koloni cendawan G. boninense
berubah warnanya dari putih menjadi kecoklatan (Gambar 6b). Isolat ini juga menghasilkan persentase penghambatan yang tertinggi terhadap cendawan G.
boninense yaitu sebesar 76.183% (Tabel 3).
Mekanisme penghambatan lainnya adalah mekanisme antibiosis yang ditandai dengan adanya zona bening di antara pertemuan koloni cendawan uji dengan koloni cendawan patogen. Isolat yang menunjukkan mekanisme ini yaitu isolat Penicillium sp. 13 dan Chrysosporium sp. dengan lebar zona bening sebesar 1.08 mm dan 3.42 mm (Gambar 6c). Selain kedua mekanisme tersebut didapatkan juga mekanisme penghambatan yang lain, yaitu terbentuknya suatu zona seperti zona bening. Zona tersebut dapat terbentuk diduga karena pengaruh antibiosisnya tidak bertahan lama, sehingga miselium yang awalnya terhambat, dapat tumbuh kembali yang lebih tipis dari miselium yang menjauhi isolat uji. Isolat yang menunjukkan mekanisme ini adalah Aspergillus sp. 1 dan UF 7 dengan lebar 2.42 mm dan 3 mm (Gambar 6d). Hasil penelitian Makut dan Owolewa (2011) menunjukkan bahwa cendawan Aspergillus flavus dan Penicillium sp. menghasilkan senyawa antibiotik yang ditunjukkan dengan adanya zona bening pada uji penghambatan terhadap Candida albicans sebesar 12 mm.
Pada perlakuan uji antagonisme yang lain, persentase penghambatan yang tinggi juga ditunjukkan oleh cendawan Penicillium sp. 2 dan Aspergillus sp. 2 sebesar 75.584% dan 74.990%. Hasil ini tidak berbeda nyata dengan hasil yang ditunjukkan oleh cendawan Trichoderma sp. 2 berdasarkan uji Duncan (α= 0.05) (Tabel 3 dan Lampiran 5). Hasil pengujian kedua cendawan tersebut memang tinggi akan tetapi perlu diperhatikan keamanannya bagi tumbuhan maupun manusia karena kedua cendawan tersebut juga dapat menghasilkan senyawa aflatoksin yang beracun dan sering mengontaminasi pada tanaman (Wilson dan Payne 1994). Pada perlakuan isolat Acremonium sp. dan UF 3 tidak terjadi penghambatan pada G. boninense, yang ditunjukkan nilai penghambatan yang negatif (Tabel 3). Hal ini dikarenakan pertumbuhan isolat cendawan Acremonium
sp. yang lebih lambat dari cendawan G. boninense.
Gambar 6 Hasil uji antagonisme cendawan uji dengan G. boninense pada 5 hsp: (a) kontrol, (b) kompetisi (Trichoderma sp. 2), (c) antibiosis (Penicillium sp. 13), (d) seperti zona bening (Uf 7)
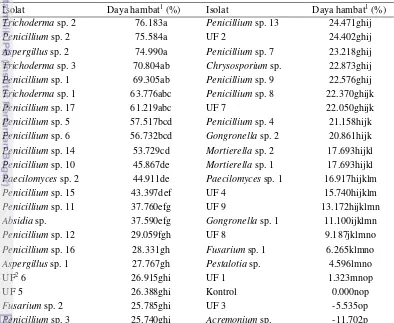
Tabel 3 Persentase penghambatan cendawan uji terhadap G. boninense pada 5 hsp
Isolat Daya hambat1 (%) Isolat Daya hambat1 (%)
Trichoderma sp. 2 76.183a Penicillium sp. 13 24.471ghij
Penicillium sp. 2 75.584a UF 2 24.402ghij
Aspergillus sp. 2 74.990a Penicillium sp. 7 23.218ghij
Trichoderma sp. 3 70.804ab Chrysosporium sp. 22.873ghij
Penicillium sp. 1 69.305ab Penicillium sp. 9 22.576ghij
Trichoderma sp. 1 63.776abc Penicillium sp. 8 22.370ghijk
Penicillium sp. 17 61.219abc UF 7 22.050ghijk
Penicillium sp. 5 57.517bcd Penicillium sp. 4 21.158hijk
Penicillium sp. 6 56.732bcd Gongronella sp. 2 20.861hijk
Penicillium sp. 14 53.729cd Mortierella sp. 2 17.693hijkl
Penicillium sp. 10 45.867de Mortierella sp. 1 17.693hijkl
Paecilomyces sp. 2 44.911de Paecilomyces sp. 1 16.917hijklm
Penicillium sp. 15 43.397def UF 4 15.740hijklm
Penicillium sp. 11 37.760efg UF 9 13.172hijklmn
Absidia sp. 37.590efg Gongronella sp. 1 11.100ijklmn
Penicillium sp. 12 29.059fgh UF 8 9.187jklmno
Penicillium sp. 16 28.331gh Fusarium sp. 1 6.265klmno
Aspergillus sp. 1 27.767gh Pestalotia sp. 4.596lmno
UF2 6 26.915ghi UF 1 1.323mnop
UF 5 26.388ghi Kontrol 0.000nop
Fusarium sp. 2 25.785ghi UF 3 -5.535op
Penicillium sp. 3 25.740ghi Acremonium sp. -11.702p
1Angka-angka pada kolom yang sama yang diikuti oleh huruf yang sama tidak berbeda nyata pada
taraf uji 5% (uji Duncan). 2UF (unidentified fungi).
Cendawan Trichoderma sp. 2 yang menghasilkan persentase penghambatan tertinggi terhadap cendawan patogen G. boninense didapatkan dari sampel palem liar (PLdK dan PLS) dan perkebunan kelapa sawit Bukit Harapan (H04 a dan H04 c), sedangkan pada sampel perkebunan kelapa sawit Bukit Duabelas tidak ditemukan cendawan ini. Cendawan ini paling banyak ditemukan pada sampel PLdK sebesar 8.7 x 104 cfu/g. Sampel ini berasal dari hutan di dekat area kebun
karet yang diduga ekosistemnya lebih seimbang karena keanekaragaman tumbuhan di dalamnya yang mempengaruhi keseimbangan mikroba tanah termasuk cendawan di dalamnya.
12
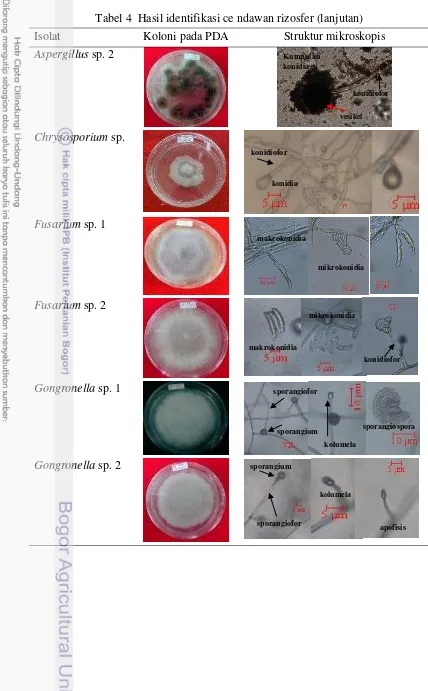
Identifikasi Cendawan
Isolat cendawan yang diidentifikasi berjumlah 43 dari 59 isolat. Hal ini disebabkan adanya isolat yang terkontaminasi dengan cendawan lain yang tidak dapat dimurnikan kembali selama proses penelitian. Identifikasi cendawan dilakukan dengan mengamati morfologi cendawan secara mikroskopis pada mikroskop cahaya dan morfologi koloni pada media PDA dengan mengacu kepada buku Pictorial Atlas of Soil and Seed Fungi (Watanabe 2002) (Tabel 4). Hasil identifikasi dari 43 isolat cendawan diperoleh 11 genus cendawan dan 9 cendawan yang belum teridentifikasi. Sebelas genus tersebut yaitu Absidia, Acremonium, Aspergillus, Chrysosporium, Fusarium, Gongronella, Mortierella,
Paecilomyces, Penicillium, Pestalotia dan Trichoderma.
Absidia sp.
Cendawan Absidia sp. memiliki miselium aerial, awalnya berwarna putih kemudian berubah menjadi kecoklatan dan pertumbuhannya cepat. Struktur mikroskopisnya berupa hifa aseptat, sporangiofornya bercabang atau tidak, lurus atau melengkung, kolumela berbentuk agak bulat, hialin dan berada diujung sporangiofor, sporangium bulat dan sporangiosporanya berbentuk elips dan hialin. Acremonium sp.
Cendawan ini memiliki morfologi berupa miselium berwarna putih pada awalnya kemudian muncul warna hitam, miselium padat, melekat di media dan permukaannya kasar. Struktur mikroskopis cendawan ini berupa hifa bersekat, konidiofor panjang atau pendek, bercabang atau tidak, dan terdapat kumpulan konidia di ujungnya. Konidia berbentuk lonjong dan agak gelap.
Aspergillus sp.
Cendawan Aspergillus sp. yang didapatkan memiliki miselium berwarna hitam (Aspergillus sp. 2) dan miselium berwarna coklat (Aspergillus sp. 1) serta permukaan koloni bertepung. Struktur mikroskopisnya berupa hifa bersekat, konidiofor panjang dan tidak bercabang, ujung konidiofor membengkak atau disebut vesikel dan merupakan tempat melekatnya kumpulan fialid, konidia bulat dan tersusun berantai.
Chrysosporium sp.
Cendawan ini memiliki miselium berwarna putih dan mengembang seperti kapas di bagian tengah sedangkan di bagian samping melekat di permukaan media dan berwarna kecoklatan. Struktur hifa cendawan ini bersekat, konidiofor pendek atau tidak ada, konidia melekat diujung konidiofor atau menempel langsung di hifa. Konidia berbentuk bulat agak lonjong seperti buah pir dan berwarna kecoklatan.
Fusarium sp.
mikrokonidia berbentuk elips atau oval. Gongronella sp.
Miselium cendawan ini berwarna putih, aerial seperti kapas dan pertumbuhannya cepat. Struktur hifanya tidak bersekat, sporangiofornya bercabang atau tidak, kolumela bulat dan hialin berada di ujung sporangiofor, sporangium bulat yang berisi sporangiospora di dalamnya. Sporangiospora berbentuk elips dan hialin.
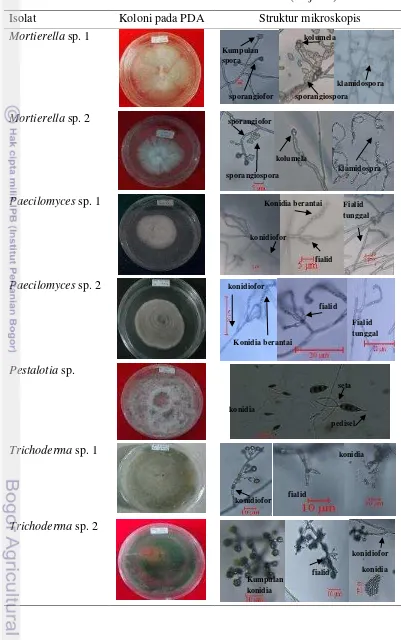
Mortierella sp.
Morfologi cendawan ini berupa miselium berwarna putih, tipis, melekat di media, permukaannya halus dan lama kelamaan muncul bintil-bintil warna putih
(Mortierella sp. 1) dan ada yang permukaannya seperti bertepung (Mortierella sp.
2). Cendawan ini memiliki struktur hifa aseptat, sporangiofor panjang atau pendek, bercabang atau tidak, terdapat kolumela diujung sporangiofor. Sporangiospora mengumpul di kolumela, berbentuk bulat dan hialin.
Paecilomyces sp.
Cendawan Paecilomyces yang didapat memiliki miselium berwarna putih, melekat di media dan bagian bawah berwarna merah muda (Paecilomyces sp. 1) dan miselium berwarna merah muda (Paecilomyces sp. 2). Struktur hifanya bersekat, konidiofor panjang, dan diujungnya terdapat kumpulan fialid, ada yang tunggal, konidia menempel pada ujung fialid, tersusun berantai, berbentuk bulat dan hialin.
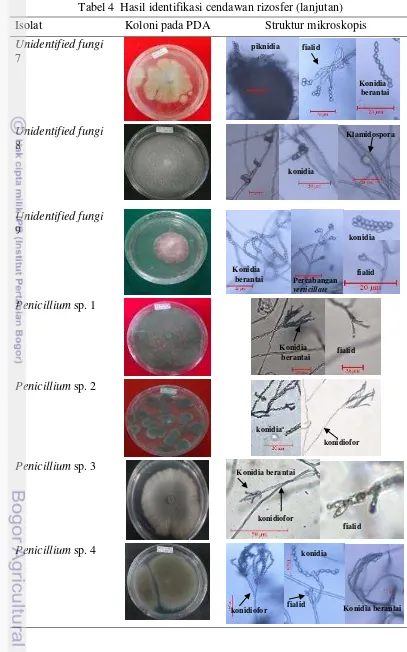
Penicillium sp.
Genus cendawan Penicillium adalah yang paling banyak ditemukan dan memiliki warna miselium yang bermacam-macam. Cendawan ini umumnya memiliki miselium berwarna putih dan lama kelamaan berubah menjadi hijau, hijau muda (Penicillium sp. 9, Penicillium sp. 13 dan Penicillium sp. 17), hijau tua (Penicillium sp. 7, Penicillium sp. 8 dan Penicillium sp. 11) atau abu-abu
(Penicillium sp. 4). Selain itu isolat dibedakan dari warna tepian dan permukaan
bawah. Warna tepian dan permukaan bawah isolat juga bermacam-macam, yaitu putih (Penicillium sp. 2), putih dan coklat ditengah (Penicillium sp. 16), jingga (Penicillium sp. 15), coklat (Penicillium sp. 14, Penicillium sp. 5 dan Penicillium
sp.6), kuning kecoklatan (Penicillium sp. 1 dan Penicillium sp. 10), kuning dan lama-kelamaan menjadi merah (Penicillium sp. 12 dan Penicillium sp. 3). Struktur hifanya bersekat, konidiofor panjang, dan diujungnya terdapat kumpulan fialid, konidia menempel pada ujung fialid, tersusun berantai, berbentuk bulat dan hialin. Pestalotia sp.
14
Trichoderma sp.
Cendawan Trichoderma sp. pertumbuhannya sangat cepat, memiliki miselium aerial, awalnya berwarna putih, kemudian berubah menjadi putih kehijauan atau kuning kehijauan (Trichoderma sp. 1), hijau tua dan menggumpal (Trichoderma sp. 2), dan ada yang bermiselium aerial pada awalnya kemudian melekat dan menggumpal seluruhnya pada media berwarna hijau tua (Trichoderma sp. 3). Struktur hifa cendawan ini bersekat, konidiofor bercabang dan terdapat fialid disetiap percabangannya. Percabangan fialid ada yang rapat ada yang renggang. Kumpulan konidia melekat diujung fialid, konidia berbentuk bulat atau oval dan hialin.
Unidentified Fungi 1
Cendawan ini memiliki miselium aerial di bagian tengah koloni, dan bagian sampingnya melekat di permukaan media. Miselium berwarna kuning kecoklatan dan hitam dibagian tengah dan lama kelamaan koloni akan berwarna hitam seluruhnya. Cendawan ini memiliki hifa bersekat, konidiofor panjang atau pendek, konidia menempel di ujung konidiofor atau melekat langsung di hifa, konidia berbentuk bulat atau seperti buah pir berwarna hitam.
Unidentified Fungi 2
Cendawan ini memiliki miselium aerial seperti kapas di bagian tengah dan melekat di bagian samping, miselium berwarna putih di tengah dan keabuan di bagian samping. Struktur hifa cendawan ini bersekat, konidiofor panjang atau pendek, konidia melekat diujung konidiofor atau menempel langsung di hifa. Konidia berbentuk bulat.
Unidentified Fungi 3
Cendawan ini memiliki miselium berwarna putih kecoklatan, melekat di permukaan media. Struktur hifa cendawan ini bersekat, konidiofor atau fialid panjang, konidia tersusun berantai atau tidak, hialin dan berbentuk elips.
Unidentified Fungi 4
Cendawan ini memiliki miselium berwarna kuning dan merah kecoklatan yang membentuk lingkaran konsentris, melekat di permukaan media. Struktur hifa cendawan ini tidak bersekat, sporangiofornya panjang, percabangannya didekat pangkal, kolumela bulat dan hialin berada di ujung sporangiofor, sporangium bulat yang berisi sporangiospora di dalamnya. Sporangiospora berbentuk elips dan hialin.
Unidentified Fungi 5
Cendawan ini memiliki miselium berwarna putih dan kuning di bagian tengah pada awalnya dan berubah kecoklatan, melekat di permukaan media. Struktur hifa cendawan ini bersekat, terdapar struktur seperti piknidia berwarna hitam, selain itu terdapat struktur konidiofor dan konidia bulat yang tersusun berantai pada fialid.
Unidentified Fungi 7
Cendawan ini memiliki miselium berwarna kecoklatan, melekat di permukaan media. Struktur hifa cendawan ini bersekat, terdapat struktur seperti piknidia berwarna hitam, selain itu terdapat struktur konidiofor dan konidia bulat yang tersusun berantai pada fialid.
Unidentified Fungi 8
Cendawan ini memiliki miselium aerial dan berwarna putih. Struktur hifa cendawan ini bersekat, terdapat struktur konidia berbentuk lonjong, berwarna gelap dan menempel pada konidiofor.
Unidentified Fungi 9
Cendawan ini memiliki miselium berwarna merah muda. Struktur hifa cendawan ini bersekat, konidiofor panjang, dan terdapat fialid dengan percabangan verticillate, konidia berbentuk oval, berwarna hialin dan tersusun berantai.
Tabel 4 Hasil identifikasi cendawan rizosfer
Isolat Koloni pada PDA Struktur mikroskopis
16
Tabel 4 Hasil identifikasi ce ndawan rizosfer (lanjutan)
Isolat Koloni pada PDA Struktur mikroskopis
18
Tabel 4 Hasil identifikasi cendawan rizosfer (lanjutan)
Isolat Koloni pada PDA Struktur mikroskopis
20
Tabel 4 Hasil identifikasi cendawan rizosfer (lanjutan)
Isolat Koloni pada PDA Struktur mikroskopis
Isolat Koloni pada PDA Struktur mikroskopis
Penicillium sp. 12
Penicillium sp. 13
Penicillium sp. 14
Penicillium sp. 15
Penicillium sp. 16
Penicillium sp. 17
fialid konidiofor
konidia
konidia
fialid
konidia fialid
konidiofor
konidia
fialid fialid
konidia
konidiofor
fialid konidiofor
22
KESIMPULAN DAN SARAN
Kesimpulan
Kelimpahan cendawan tanah tertinggi diperoleh dari sampel kelapa sawit B04 a sebesar 4.9 x 106 cfu/g sedangkan kelimpahan cendawan terendah diperoleh dari sampel kelapa sawit B04 b sebesar 0.3 x 105 cfu/g. Total urutan
keanekaragaman cendawan dari yang tertinggi ke terendah adalah H04, palem liar dan B04 (30, 27, 16 isolat) akan tetapi setelah diidentifikasi pada tingkat genus, keanekaragaman cendawan dari yang tertinggi adalah palem liar (8 genus dan 13 UF), H04 (6 genus dan 9 UF), dan B04 (4 genus dan 6 UF). Isolat yang ditemukan di setiap sampel adalah isolat Penicillium sp. 12 dan isolat yang berpotensi sebagai agens antagonis adalah isolat Trichoderma sp. 2 dengan persentase penghambatan terhadap G. boninense sebesar 76.183% yang didapatkan dari sampel PLS, PLdK, H04 a dan H04 c. Selain itu didapatkan isolat Penicillium sp. 13 dan Chrysosporium sp. yang dapat menghasilkan antibiosis.
Saran
[DITJENBUN] Direktorat Jenderal Perkebunan. 2013a. Perkembangan Luas Areal Perkebunan 2008–2013. Jakarta (ID): Direktorat Jenderal Perkebunan; [diunduh 2013 Januari 16]. Tersedia pada: http:// ditjenbun. pertanian. go.id/ statis-35-luasareal.html.
[DITJENBUN] Direktorat Jenderal Perkebunan. 2013b. Perkembangan Produksi Komoditi Perkebunan 2008–2013. Jakarta (ID): Direktorat Jenderal Perkebunan; [diunduh 2013 Januari 16]. Tersedia pada: http://ditjenbun. pertanian. go.id/statis-36-produksi.html.
Izzati ZMNA, Abdullah F. 2008. Disease suppression in Ganoderma-infected oil palm seedling treated with Trichoderma harzianum. J. Plant Protection Sci.
44(3):101-107.
Kennedy AC. 2005. Rhizosphere. Di dalam: Sylvia DM, Fuhrmann JJ, Hartel PG, Zuberer DA,editor. Principles and Aplications of Soil Microbiology. 2nd ed. New Jersey (US): Pearson Prentice Hall. hlm 242-262.
Li CJ, Gao JH, Nan ZB. 2007. Interactions of Neotyphodium gansuense,
Achnatherum inebrians, and plant-pathogenic fungi. Mycol Res. 3:
1220-1227.
Lo CT. 1998. General mechanisms of action of microbial biocontrol agents. Plant Pathol Bull. 7: 155-166.
Magurran AE. 1987. Ecologycal Diversity and Its Measurement. New Jersey (US): Princeton University Press.
Makut MD, Owolewa OA. 2011. Antibiotic-producing fungi present in the soil environment of Keffi Metropolis, Nasarawa State, Nigeria. Trakia J Sci.
9(2): 33-39.
Mattjik AA, Sumertajaya M. 2006. Perancangan Percobaan dengan Aplikasi SAS
dan Minitab. Bogor (ID): Institut Pertanian Bogor Press.
Nildayanti. 2011. Peran bakteri kitinolitik dan fungi mikoriza arbuskular dalam pengendalian busuk pangkal batang kelapa sawit [tesis]. Bogor (ID): Sekolah Pascasarjana Institut Pertanian Bogor.
Paterson RRM. 2007. Ganoderma disease of oil palm-a white rot perspective necessary for integrated control. Crop Protection. 26:1369-1376. doi:10.1016/j.cropro.2006.11.009.
Puangsombat P, Sangwanit U, Marod D. 2010. Diversity of soil fungi in different land use type in Tha Kum-Huai Raeng forest reserve, Trat province.
Kasetsart J. (Nat. Sci.). 44(6): 1162-1175.
Shi LL, Mortimer PE, Slik JWF, Zou XM, Xu J, Feng WT, Qiao L. 2013. Variation in forest soil fungal diversity along a latitudinal gradient. Fungal
Diversity. doi: 10.1007/s13225-013-0270-5.
Susanto A. 2002. Kajian pengendalian hayati Ganoderma boninense Pat. penyebab penyakit busuk pangkal batang kelapa sawit [disertasi]. Bogor (ID): Institut Pertanian Bogor.
24
Tortora GJ, Funke BR, Case CL. 2004. Microbiology an Introduction. 8th ed. San
Francisco (US): Benjamin Cummings.
Turner PD. 1981. Oil Palm Disease and Disorders. Oxford (US): Oxford University Press.
Watanabe T. 2002. Pictorial Atlas of Soil and Seed Fungi: Morphologies of
Cultured Fungi and Key to Species. Ed ke-2. Boca Raton (US): CRC Press.
Wilson DM, Payne GA. 1994. Factors affecting Aspergillus flavus group infection and aflatoxin contamination of crops. Di dalam: Eaton DL and Groopman JD, editor. The Toxicology of Aflatoxins. Human Health, Veterinary, and
Agricultural Significance. San Diego (US): Academic Press. hlm 383-406.
Zak DR, Holmes WE, White DC, Peacock AD, Tilman D. 2003. Plant diversity, soil microbial communities, and ecosystem function: are there any links?.
26
Lampiran 1 Beberapa isolat cendawan yang berhasil diisolasi dari tanah tetapi terkontaminasi sehingga tidak dapat diidentifikasi lebih lanjut
Lampiran 2 Kelimpahan setiap cendawan pada rizosfer palem liar
Isolat Kelimpahan (10⁴ cfu/g) Isolat Kelimpahan (10⁴ cfu/g)
PLK PLS PLdK PLK PLS PLdK
Absidia sp. 0.7* 0 0 Paecilomyces sp.1 0.3* 0 0
Trichoderma sp.2 0 1.3* 8.7 Aspergillus sp.1 1.7* 0 0
Trichoderma sp.1 6 3* 3.7 UF 14 0 0 13.3*
Gongronella sp.1 2* 1* 6 UF 15 80 0 0
UF** 10 0 0.3* 0 Gongronella sp.2 1.7* 0 0
UF 11 0 0 1.3* UF 16 0 493 177*
Penicillium sp.2 27 39 39 UF 17 0 0 3.3*
UF 12 0 3.3* 0 Fusarium sp.1 0 1* 0
Penicillium sp.3 0 0.3* 0 Fusarium sp.2 0 0.7* 0
UF 5 0 3.3* 0 UF 18 0.3* 0 0
Penicillium sp.12 2.3* 1.7* 0.7* Paecilomyces sp.2 2.7 0 0
UF 7 1* 0 0 UF 19 0 3.3* 0
Pestalotia sp. 3.3* 0 0 UF 9 2.7 0 0
UF 13 0 0 3.3*
Isolat Kelimpahan (10⁴ cfu/g) Isolat Kelimpahan (10⁴ cfu/g)
*angka estimasi. ** UF (unidentified fungi).
Lampiran 4 Kelimpahan cendawan pada rizosfer kelapa sawit di Bukit 12
Isolat Kelimpahan (10⁴ cfu/g) Isolat Kelimpahan (10⁴ cfu/g)
*angka estimasi. ** UF (unidentified fungi).
28
RIWAYAT HIDUP
Penulis dilahirkan di Lamongan Jawa Timur pada tanggal 20 November 1990, sebagai anak kesembilan dari 10 bersaudara dari Bapak Mustaman dan Ibu Zahroh. Tahun 2009 penulis lulus dari MA Al-Ishlah Sendangaagung Paciran Lamongan Jawa timur dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Beasiswa Utusan Daerah Kementerian Agama dan diterima di Departemen Proteksi Tanaman, Fakultas Pertanian.