PERTUMBUHAN UDANG VANAME
(
Litopenaeus vannamei
, Boone) DI MEDIA
BERSALINITAS RENDAH
ERLY YOSEF KALIGIS
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Peningkatan Sintasan dan Kinerja Pertumbuhan Udang Vaname (Litopenaeus vannamei, Boone) di Media Bersalinitas Rendah adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Mei 2010
Erly Y. Kaligis NIM. C161050051
Performance of the Pacific White Shrimp (Litopenaeus vannamei, Boone) Rare on Low Salinity Medium. Under direction of DANIEL DJOKOSETIYANTO, RIDWAN AFFANDI, ING MOKOGINTA, and KADARWAN SOEWARDI.
The research was carried out in three phases. The first phase was to study the effect of adding calcium on the short-term survival of Pacific white shrimp postlarvae (Litopenaeus vannamei, Boone). Preliminary experiment consisted of
16 treatments in combination between the level of calcium (0 ppm, 50 ppm, 100 ppm, 150 ppm) and the medium salinity endpoint (0 ppt, 2 ppt, 4 ppt, 6 ppt).
The second experiment consist of different Ca2+ level namely A (0 ppm Ca2+), B (50 ppm Ca2+), C (100 ppm Ca2+), D (150 ppm Ca2+), and D (25 ppt salinity /control). Several biological responses, including the survival rate in first trial, osmoregulatory capacity and oxygen consumption in the main trial, were discussed. Result of the first experiment showed that maximum percentage of mean survival rate was 92.2 % in B treatment (50 ppm Ca2+). No significant effect between calcium level observed on both osmoregulatory capacity and oxygen consumption, but it occurs lower in salinity 25 ppt and control. The second phase was conducted to determine the effect of potassium concentration medium on the metabolism, potassium whole body, survival rate, growth, and feed eficiency of L. vannamei postlarvae. Treatments consisted of different concentration of K+ added to distilled water. Four treatments were set, namely A, B, C, and D, where K2CO3 levels were 0, 30, 60, and 90 ppm, respectively. After
the 42-days feeding trial, survival rate of the shrimp under the four treatments were not significantly affected by different K+ concentration in low salinity water. Solutions with K+ without or added K+ exhibited an average high survival rate above 83.33%. The final wet weight of the shrimp under treatment D (90 ppm K2CO3) were significantly higher, while the value of osmoregulatory capacity and
oxygen consumption in treatment A (0 ppm K2CO3) significantly higher than
those under the other four treatments.The aim of the third phase was to study the effect of protein and calcium level in the diet on the growth performance of postlarvae. Three level of dietary protein and three level of calcium was used in this experiment. Fifteen shrimp (PL25) were placed in triplicate 45-L glasses aquarium. The shrimp fed on the experimental diets four times daily, 8% of body weight. Feeding trial was conducted for 28 days. Growth performance was evaluated by RNA/DNA ratio, calcium and protein retention, frequency of moulting, daily growth rate, and feed efficiency. The last experiment showed that the survival rate was the same between treatments. Shrimp fed on diet 45% protein and 2% calcium produce higher RNA/DNA ratio, protein retention, daily growth rate, and feed efficiency compare to the other treatments. It was concluded that postlarvae L. vannamei require 45% protein and 2% calcium in low salinity condition.
vaname (Litopenaeus vannamei, Boone) di media bersalinitas rendah. Dibimbing oleh DANIEL DJOKOSETIYANTO, RIDWAN AFFANDI, ING MOKOGINTA, dan KADARWAN SOEWARDI.
Penelitian dilakukan dalam tiga tahap. Penelitian tahap pertama terdiri dari dua seri percobaan. Percobaan pertama bertujuan mengkaji pengaruh penambahan berbagai kadar kalsium pada berbagai salinitas media terhadap sintasan postlarva vaname (Litopenaeus vannamei, Boone). Rancangan yang digunakan adalah model faktorial terdiri enam belas perlakuan dengan 3 ulangan dibedakan berdasarkan penurunan salinitas (0 ppt, 2 ppt, 4 ppt, 6 ppt) dengan penambahan kalsium (0 ppm, 50 ppm, 100 ppm, 150 ppm) dalam media pengencer air tawar. Percobaan kedua merupakan pengulangan dari percobaan pertama pada salinitas akhir 2 ppt melalui evaluasi osmolaritas dan laju metabolisme. Rancangan percobaan menggunakan Rancangan Acak Lengkap terdiri 5 perlakuan berdasarkan penambahan kalsium dalam media yaitu: (A) 0 ppm, (B) 50 ppm, (C) 100 ppm, (D) 150 ppm, dan kontrol (salinitas 25 ppt) dengan 3 ulangan. Benih vaname (PL 20) yang digunakan diperoleh dari balai pembenihan (hatchery) komersial hasil pemijahan dari satu induk dalam upaya meminimalkan variasi unit percobaan. Setiap wadah diisi air bersalinitas 25 ppt yang kemudian diturunkan secara gradual selama 96 jam hingga salinitas perlakuan akhir 0 ppt, 2 ppt, 4 ppt, dan 6 ppt. Sedangkan pada percobaan kedua salinitas akhir yang dituju 2 ppt. Pemberian pakan Artemia dilakukan 4 kali perhari.
Beberapa respon biologi yang diamati dalam percobaan pertama yaitu sintasan, kemudian variabel gradien osmotik dan konsumsi oksigen diamati selanjutnya pada percobaan kedua. Hasil penelitian menunjukkan bahwa rata-rata sintasan tertinggi di salinitas 0 ppt dicapai dengan perlakuan B (0 ppt; 50 ppm kalsium) yaitu sekitar 92,2%. Pada percobaan kedua, seluruh perlakuan penambahan Ca2+ menunjukkan sintasan yang tinggi. Berdasarkan pengamatan juga tidak terdapat perbedaan gradien osmotik dan tingkat konsumsi oksigen pada perlakuan kalsium di salinitas 2 ppt, akan tetapi gradien osmotik dan tingkat konsumsi oksigen lebih tinggi pada udang di media salinitas 2 ppt daripada kontrol (salinitas 25 ppt). Disarankan bahwa penambahan kalsium 50 ppm sebagai perlakuan terbaik untuk meningkatkan sintasan postlarva vaname selama aklimasi ke salinitas rendah.
Penelitian tahap kedua bertujuan menentukan kadar potasium optimal bagi kandungan potasium tubuh, gradien osmotik, tingkat konsumsi oksigen, sintasan, laju pertumbuhan bobot rerata harian, dan efisiensi pemanfaatan pakan pada media bersalinitas 2 ppt. Perlakuan merupakan perbedaan kadar potasium dalam air bersalinitas rendah. Ada 4 perlakuan yang digunakan yaitu A (0 ppm K2CO3),
B (30 ppm K2CO3), C (60 ppm K2CO3), dan D (90 ppm K2CO3). Sebelum
tertinggi dicapai pada perlakuan D (90 ppm K2CO3), sementara gradien osmotik
dan tingkat konsumsi oksigen secara nyata lebih tinggi di perlakuan A (0 ppm K2CO3) dan B (30 ppm K2CO3).
Tujuan dari penelitian tahap ketiga adalah mengkaji pengaruh kadar protein dan kalsium pakan terhadap kinerja pertumbuhan postlarva vaname.
Perlakuan terdiri atas perbedaan kadar protein (25, 35, 45 %) dengan kalsium (0, 2, 4 %) dalam pakan. Hewan uji yang digunakan sebelumnya telah
diaklimatisasi dalam kondisi laboratorium kemudian dilanjutkan tahapan aklimasi secara gradual dari salinitas 25 ppt ke 2 ppt. Saat stadia PL 25, postlarva diseleksi berdasarkan ukuran yang sama dengan bobot awal rata-rata sekitar 0,0279 g. Setiap akuarium ditebar udang sebanyak 15 individu. Selanjutnya udang uji diberikan pakan perlakuan 4 kali perhari. Waktu pemeliharaan untuk mengkaji efek pemberian pakan terhadap postlarva sekitar 4 minggu. Beberapa variabel yang diamati pada penelitian tahap ketiga adalah rasio RNA/DNA, retensi protein, retensi kalsium, frekuensi ganti kulit, laju pertumbuhan bobot rerata harian, efisiensi pemanfaatan pakan, dan sintasan.
Hasil pengujian memperlihatkan respons nyata terhadap kinerja pertumbuhan. Perlakuan terbaik untuk mempercepat peningkatan pertumbuhan postlarva adalah kadar protein 45% dengan kadar kalsium 2% pakan (perlakuan H). Dengan kadar protein dan kalsium optimal, terjadi peningkatan rasio RNA/DNA, retensi protein, retensi kalsium, dan efisiensi pakan, sehingga menghasilkan laju pertumbuhan yang tinggi pada pemeliharaan di media salinitas 2 ppt.
PERTUMBUHAN UDANG VANAME
(
Litopenaeus vannamei
, Boone) DI MEDIA
BERSALINITAS RENDAH
ERLY YOSEF KALIGIS
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PENGUJI PADA UJIAN TERTUTUP Dr. Ir.Agus Oman Sudrajat, M.Sc.
Staf Pengajar Departemen Budidaya Perairan, FPIK, IPB Dr. Ir. Tatag Budiardi, M.Si.
Staf Pengajar Departemen Budidaya Perairan, FPIK, IPB
PENGUJI PADA UJIAN TERBUKA Dr. Ir. Zafril Imran Azwar, M.S.
Peneliti di Balai Riset Perikanan Air Tawar, Bogor Prof. Dr. Ir. Enang Harris, M.S.
Nama : Erly Yosef Kaligis
NIM : C161050051
Disetujui
Komisi Pembimbing
Prof. Dr. Ir. Daniel Djokosetiyanto, DEA Dr. Ir. Ridwan Affandi, DEA Ketua Anggota
Prof. Dr. Ir. Ing Mokoginta Prof. Dr. Ir. Kadarwan Soewardi Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB Ilmu Perairan
Prof. Dr. Ir. Enang Harris, M.S. Prof. Dr. Ir. Khairil Anwar Notodiputro, M.S.
@ Hak Cipta milik IPB, tahun 2010 Hak Cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencamtumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian,
penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
atas segala berkatNya sehingga penulisan hasil penelitian disertasi dengan judul “Peningkatan sintasan dan kinerja pertumbuhan udang vaname (Litopenaeus vannamei, Boone) di media bersalinitas rendah” dapat terselesaikan. Disertasi ini merupakan salah satu syarat untuk menyelesaikan pendidikan Program Doktor di Program Studi Ilmu Perairan, Sekolah Pascasarjana, Institut Pertanian Bogor.
Penulis mengucapkan terima kasih yang mendalam kepada Bapak Prof. Dr. Ir. Daniel Djokosetiyanto, DEA, Bapak Dr. Ir. Ridwan Affandi,
DEA, Ibu Prof. Dr. Ir. Ing Mokoginta, dan Bapak Prof. Dr. Ir. Kadarwan Soewardi selaku komisi pembimbing atas semua arahan, masukan, dan nasihat selama penelitian dan penulisan disertasi. Ucapan terima kasih juga kepada Bapak Prof. Dr. Ir. Enang Harris, M.S., Bapak Dr. Ir. Zafril Imran Azwar, M.S., Bapak Dr. Ir. Agus Oman Sudrajat, M.Sc., Bapak Dr.Ir. Tatag Budiardi, M.Si. yang telah memberikan banyak saran dan masukan selama Ujian Terbuka dan Ujian Tertutup, juga kepada mantan ketua Program Studi Ilmu Perairan, Bapak Dr. Chairul Muluk, M.Sc. yang telah memberi masukan serta memperluas wawasan keilmuan penulis selama pendidikan.
Banyak pihak telah membantu selama studi, penelitian, hingga ujian akhir. Untuk itu, penulis menghaturkan terima kasih kepada pimpinan Universitas Sam Ratulangi dan Fakultas Perikanan dan Ilmu Kelautan, Manado atas kesempatan kepada penulis melanjutkan pendidikan, Departemen Pendidikan Nasional Republik Indonesia sebagai sponsor beasiswa pendidikan pascasarjana (BPPS), yayasan Mandiri, yayasan Toyota & Astra, Pemerintah Kabupaten Minahasa Selatan atas bantuan dana penelitian disertasi, Pemerintah Daerah Sulawesi Utara yang telah memberikan bantuan dana pendidikan dan fasilitas tempat tinggal selama di Bogor, serta kepada PT. Central Proteinaprima, Tbk. dan PT. BAA Anyer yang telah memberi bantuan hewan uji bagi terlaksananya penelitian.
Lolowang, Debby Rayer, Susan Rumengan, Donata Pandin, Jack Mamangkey, Hasnawaty, Hengky Sinjal, Agung Windarto, Alfret Luasunaung; Rekan-rekan di
Bogor: Deny Karwur, Broerie Pojoh, Ridwan Lasabuda, Joy Kumaat, Sukma Gedoan, Only Rembet, Altien Rindengan, dan Yosep Karamoy.
Secara khusus, penulis menyampaikan terima kasih kepada Ayahanda Deki Kaligis dan Ibu Marrien Sekeon, kakak-kakak: Jou, Denny, Ito, dan seluruh keluarga atas bantuan doa dan dukungan moril selama penyelesaian studi.
Penulis menyadari bahwa apa yang sudah dicapai melalui penulisan disertasi ini masih belum sempurna. Oleh karena itu, saran dan kritik diperlukan bagi penyempurnaan tulisan ini. Semoga tulisan ini bermanfaat dan memberikan terobosan baru dalam budidaya udang di Indonesia.
Bogor, Mei 2010
7 September 1973 dari pasangan Bapak Deki Kaligis dan Ibu Fredrika Putong sebagai anak ke-dua dari dua bersaudara. Pendidikan sarjana ditempuh pada Fakultas Perikanan Program Studi Ilmu Kelautan, Jurusan Manajemen Sumber Daya Perairan, Universitas Sam Ratulangi, Manado, dan lulus pada tahun 1998. Selanjutnya, penulis diangkat menjadi staf pengajar Fakultas Perikanan sejak tahun 2000 pada Program Studi Ilmu Kelautan, Jurusan Manajemen Sumber Daya Perairan. Pada tahun 2002 penulis melanjutkan studi ke program Magister sains (S2) di Program Studi Ilmu Perairan, Sekolah Pascasarjana IPB, dan lulus pada tahun 2005. Selanjutnya, pada tahun 2005 penulis memperoleh kesempatan untuk melanjutkan studi program doktor (S3)
x
Peranan Mineral Kalsium dan Potasium ... 17
Kebutuhan Nutrien Udang ... 23
BAHAN DAN METODE ... 28
Ruang Lingkup Penelitian ... 28
Penelitian Tahap Pertama ... 29
Waktu dan Tempat ... 29
Hasil Penelitian Tahap Pertama ... 50
Percobaan Pertama ... 50
xi
Pembahasan ... 53
Hasil Penelitian Tahap Kedua ... 57
Kandungan Potasium Tubuh ... 59
Gradien Osmotik ... 60
Tingkat Konsumsi Oksigen ... 61
Sintasan ... 62
Laju Pertumbuhan Bobot Rerata Harian ... 62
Efisiensi Pemanfaatan Pakan ... 63
Pembahasan ... 65
Hasil Penelitian Tahap Ketiga ... 70
Rasio RNA/DNA ... 70
Retensi Protein ... 71
Retensi Kalsium ... 71
Frekuensi Ganti Kulit ... 71
Laju Pertumbuhan Bobot Rerata Harian ... 72
Efisiensi Pemanfaatan Pakan ... 73
Sintasan ... 74
Pembahasan ... 74
SIMPULAN DAN SARAN ... 80
DAFTAR PUSTAKA ... 81
xii
DAFTAR TABEL
Halaman
1 Waktu serta karakteristik setiap stadia pada larva udang vaname saat berkembang pada kondisi normal (Wyban &
Sweeney 1991) ………... 11
2 Komposisi bahan pakan percobaan ………... 42
3 Komposisi proksimat dan kandungan kalsium pakan percobaan ... 43
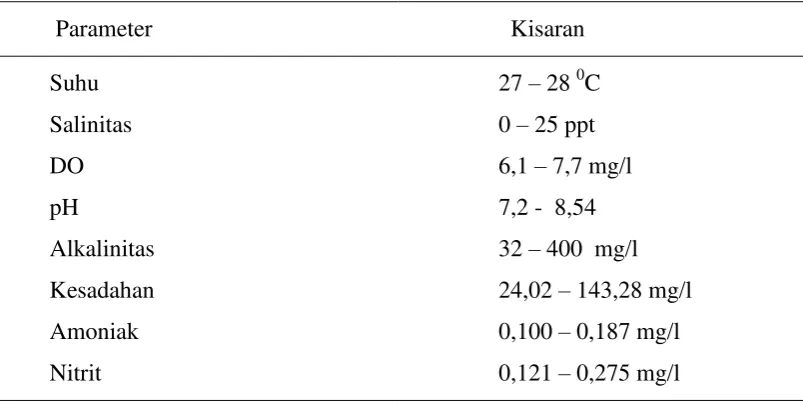
4 Parameter fisika kimia media selama percobaan ………... 47
5 Nilai gradien osmotik (GO), tingkat konsumsi oksigen (OC), pembelanjaan energi pada metabolisme basal (EB), dan sintasan (S) postlarva vaname (L. vannamei) yang terukur dalam percobaan kedua pada salinitas 2 ppt ………... 51
6 Nilai kadar potasium tubuh (K+), gradien osmotik (GO), tingkat konsumsi oksigen (OC), pembelanjaan energi pada metabolisme basal (EB), sintasan (S), laju pertumbuhan bobot rerata harian (LPRH), dan efisiensi pemanfaatan pakan (EP) postlarva vaname (L. vannamei) pada penelitian tahap kedua ……….... 58
7 Konsentrasi mineral utama setiap perlakuan pada air bersalinitas 2 ppt yang digunakan dalam penelitian tahap kedua ………. 59
xiii
DAFTAR GAMBAR
Halaman
1 Bagan alir perumusan masalah pengaruh penambahan Ca2+ selama aklimasi, penambahan mineral K+ dalam media, serta pengkayaan protein dan Ca2+ dalam pakan bagi kualitas dan kuantitas postlarva
udang vaname ……… 8
2 Anatomi lengkap L. vannamei (Wyban & Sweeney 1991) ……... 10 3 Reaksi-reaksi metabolisme dalam mineralisasi tulang (Georgievskii
VI 1982) ………. 19
4 Skema urutan kegiatan penelitian ………... 28 5 Skema pengaturan wadah pada percobaan ke-2 dalam penelitian
penelitian tahap pertama melalui tampak atas ……….... 31 6 Tata letak penempatan wadah penelitian tahap ke-2 ... 37 7 Tata letak penempatan wadah penelitian tahap ke-3 ……….. 45 8 Sintasan (S) postlarva L. vannamei setelah 4 hari percobaan pada
berbagai kombinasi salinitas dan penambahan kalsium media, yaitu 0 ppt; 0 ppm Ca2+ (A), 0 ppt; 50 ppm Ca2+ (B), 0 ppt;100 ppm Ca2+
tingkat konsumsi oksigen (OC) postlarva vaname …………...……. 62 12 Bobot rata-rata individu postlarva vaname selama enam minggu
penelitian tahap kedua dengan perlakuan K2CO3 media berbeda
(A:0 ppm K2CO3, B:30 ppm K2CO3, C:60 ppm K2CO3, D:90 ppm
K2CO3) ………... 63
13 Hubungan antara kadar potasium karbonat media dengan nilai laju
xiv 14 Hubungan antara kadar potasium karbonat media dengan nilai
efisiensi pemanfaatan pakan (EP) postlarva vaname ………. 64 15 Bobot rata-rata individu postlarva vaname selama empat minggu
penelitian tahap ketiga dengan perlakuan pakan kadar protein dan kalsium berbeda (A: 25% protein;0% kalsium, B: 25% protein;2% kalsium, C: 25% protein;4% kalsium, D: 35% protein;0% kalsium, E: 35% protein;2% kalsium, F: 35% protein;4% kalsium, G: 45% protein;0% kalsium, H: 45% protein;2% kalsium, I: 45%
xv
DAFTAR LAMPIRAN
Halaman
1 Pengukuran osmolaritas dengan osmometer (SOP OSMOMAT
030) ………. 91
2 Prosedur preparasi sampel untuk analisa mineral ... 92
3 Pengukuran kandungan mineral dengan Atomic Absorption Spectrofotometer (AAS, Shimadzu AA-680) ... 93
4 Prosedur ekstraksi DNA (Puregene) ……….. 94
5 Prosedur ektraksi RNA ………... 95
6 Prosedur pengukuran konsentrasi DNA dan RNA ………. 96
7 Prosedur analisis proksimat pakan dan udang ……….... 97
8 Hasil perhitungan sintasan pada percobaan pertama dalam penelitian tahap pertama ………... 101
9 Nilai osmolaritas cairan tubuh, osmolaritas media, gradien osmotik (GO), tingkat konsumsi oksigen (OC), pembelanjaan energi pada metabolisme basal (EB), dan sintasan (S) postlarva vaname (L. vannamei) dari setiap perlakuan dan ulangan pada akhir percobaan kedua penelitian tahap pertama ……… 102
10 Nilai kandungan potasium tubuh (K+), gradien osmotik (GO), tingkat konsumsi oksigen (OC), pembelanjaan energi pada metabolisme basal (EB), sintasan (S), laju pertumbuhan bobot rerata harian (LPRH), efisiensi pemanfaatan pakan (EP), pertumbuhan bobot (W), dan konsumsi pakan (KP) postlarva vaname (L. vannamei) dari setiap perlakuan dan ulangan pada akhir penelitian tahap kedua ……... 103
11 Peningkatan pertumbuhan bobot selama 4 minggu penelitian tahap ketiga ……… 104
12 Nilai total RNA, total DNA, rasio RNA/DNA, retensi protein (RP), retensi kalsium (RCa), frekuensi ganti kulit (FGK), laju pertumbuhan bobot rerata harian (LPRH), efisiensi pemanfaatan pakan (EP), dan sintasan (S) postlarva vaname (L. vannamei) dari setiap perlakuan dan ulangan pada akhir penelitian tahap ketiga ….. 105
xvi 14 Analisis ragam dan uji lanjut Tukey data gradien osmotik (GO)
postlarva vaname (L. vannamei) pada percobaan ke-2 penelitian
tahap pertama ... 108 15 Analisis ragam dan uji lanjut Tukey data tingkat konsumsi oksigen
(OC) postlarva vaname (L.vannamei) pada percobaan ke-2
penelitian tahap pertama ... 109 16 Analisis ragam dan uji lanjut Tukey data pembelanjaan energi pada
metabolisme basal (EB) postlarva vaname (L. vannamei) pada
percobaan ke-2 penelitian tahap pertama ... 110 17 Analisis ragam dan uji lanjut Tukey data sintasan (S) postlarva 19 Hasil analisis regresi data kandungan potasium tubuh (K+) postlarva
vaname (L. vannamei) pada penelitian tahap kedua ... 113 20 Analisis ragam dan uji lanjut Tukey data gradien osmotik (GO)
postlarva vaname (L. vannamei) pada penelitian tahap kedua ... 114 21 Hasil analisis regresi data gradien osmotik (GO) postlarva vaname
(L. vannamei) pada penelitian tahap kedua ... 115 22 Analisis ragam dan uji lanjut Tukey data tingkat konsumsi oksigen
(OC) postlarva vaname (L. vannamei) pada penelitian tahap kedua ... 116 23 Hasil analisis regresi data tingkat konsumsi oksigen (OC) postlarva
vaname (L. vannamei) pada penelitian tahap kedua ... 117 24 Analisis ragam dan uji lanjut Tukey data pembelanjaan energi pada
metabolisme basal (EB) postlarva vaname (L. vannamei) pada
penelitian tahap kedua ... 118 25 Analisis ragam dan uji lanjut Tukey data sintasan (S) postlarva
vaname (L. vannamei) pada penelitian tahap kedua ... 119 26 Analisis ragam dan uji lanjut Tukey data laju pertumbuhan bobot
rerata harian (LPRH) postlarva vaname (L. vannamei) pada
penelitian tahap kedua ... 120 27 Hasil analisis regresi data laju pertumbuhan bobot rerata harian
(LPRH) postlarva vaname (L. vannamei) pada penelitian tahap
xvii 28 Analisis ragam dan uji lanjut Tukey data efisiensi pemanfaatan
pakan (EP) postlarva vaname (L. vannamei) pada penelitian tahap
kedua ... 122 29 Hasil analisis regresi data efisiensi pemanfaatan pakan (EP)
postlarva vaname (L. vannamei) pada penelitian tahap kedua ... 123 30 Analisis ragam dan uji lanjut Tukey data konsumsi pakan (KP)
postlarva vaname (L. vannamei) pada penelitian tahap kedua ... 124 31 Analisis ragam dan uji lanjut Tukey data rasio RNA/DNA postlarva
vaname (L. vannamei) pada penelitian tahap ketiga ... 125 32 Analisis ragam dan uji lanjut Tukey data retensi (RP) protein
postlarva vaname (L. vannamei) pada penelitian tahap ketiga ... 126 33 Analisis ragam dan uji lanjut Tukey data retensi kalsium (RCa)
postlarva vaname (L. vannamei) pada penelitian tahap ketiga ... 127 34 Analisis ragam dan uji lanjut Tukey data frekuensi ganti kulit
(FGK) postlarva vaname (L. vannamei) pada penelitian tahap
ketiga ... 128 35 Analisis ragam dan uji lanjut Tukey data laju pertumbuhan bobot
rerata harian (LPRH) postlarva vaname (L. vannamei) pada
penelitian tahap ketiga ... 129 36 Analisis ragam dan uji lanjut Tukey data efisiensi pemanfaatan
pakan (EP) postlarva vaname (L. vannamei) pada penelitian tahap
PENDAHULUAN
Latar BelakangPerairan nasional Indonesia menyimpan potensi perikanan yang besar untuk dikembangkan. Dalam upaya memenuhi kebutuhan pangan masyarakat yang terus meningkat, maka sektor perikanan memikul tanggung jawab dan peran penting sebagai penyedia protein ikan. Perlu kearifan dalam mengelola kekayaan alam perairan yang dimiliki yaitu dengan mengedepankan aspek-aspek efisiensi produksi serta penerapan teknologi yang handal dan ramah lingkungan. Melalui pengembangan produksi perikanan nasional khususnya dari sektor budidaya, diharapkan dapat mengurangi ketergantungan dari alam sekaligus meningkatkan produktivitas, menjamin ketersediaan pangan, dan memberikan kesejahteraan bagi masyarakat.
Udang vaname (Litopenaeus vannamei) adalah salah satu produk perikanan yang sedang berkembang saat ini. Sejak tahun 2001, jenis udang ini telah ditetapkan pemerintah sebagai unggulan sektor perikanan budidaya di Indonesia. Salah satu tujuan diintroduksinya udang vaname adalah untuk memacu produksi udang nasional yang selama beberapa tahun sebelumnya mengalami penurunan. Kontribusi udang vaname tercermin dari produksi udang ini yang terus mengalami peningkatan setiap tahunnya, dengan total tahun 2009 mencapai 244.650 ton atau sekitar 70,2% dari produksi udang nasional (Nurdjana 2010).
Penerapan skala teknologi sederhana hingga intensif dalam produksi udang di wilayah tropis telah menunjukkan bahwa udang vaname memiliki beberapa kelebihan dibanding udang yang lain. Udang vaname memiliki sintasan
tinggi sekitar 90% walaupun dengan densitas pemeliharaan yang padat, 150 ekor/m2 (SEAFDEC 2005), pertumbuhan yang cepat (size 60-80 dalam 60 hari pertama), kandungan daging yang lebih banyak (66-68%) dibanding udang windu (62%) (Anonim 2005), serta relatif toleransi terhadap serangan penyakit viral seperti WSSV (White Spot Syndrome Virus), TSV (Taura Syndrome Virus), dan IHHNV (Infectious Hypodermal and Hematopoietic
Necrosis Virus) (Taukhid et al. 2006). Berbagai keunggulan tersebut
Selain itu, udang vaname ternyata memiliki sifat euryhalin (Velasco et al. 1999). Bray et al.(1994) menyatakan bahwa udang vaname dapat dipelihara di daerah perairan pantai (coastal) dengan kisaran salinitas 0,5 ppt - 40 ppt. Kemampuan ini memberi peluang bagi petambak udang untuk mengembangkan komoditas ini di perairan daratan (inland water). Selama ini, budidaya udang vaname dilakukan di daerah perairan bersalinitas tinggi di tambak-tambak estuari, sementara potensi lahan untuk pengembangan budidaya di air bersalinitas rendah sangat besar, mencapai 2,072 juta hektar lahan air tawar (kolam dan sawah) dengan belum termanfaatkan sekitar 89,9% (DKP 2005). Budidaya udang vaname di air bersalinitas rendah juga dapat merupakan pilihan alternatif mengingat mulai munculnya berbagai penyakit infeksi pada udang yang dipelihara di tambak air asin. Dengan penerapan teknologi pemeliharaan di lingkungan salinitas rendah, maka terbuka peluang untuk lebih memperluas produksi budidaya udang vaname.
Produksi benih berkualitas merupakan salah satu aspek penting dalam keberhasilan budidaya vaname di perairan bersalinitas rendah. Kesehatan dan vitalitas postlarva yang dihasilkan selama pendederan (nursery stage) akan menentukan potensi pertumbuhan dan sintasan di lingkungan kolam pembesaran. Hingga saat ini, telah dikembangkan berbagai metode aklimasi ke air bersalinitas rendah, namun masih dihadapkan dengan masalah sintasan yang rendah. Hana (2007) melaporkan bahwa sintasan postlarva vaname hanya sekitar 48,33% saat diaklimasi ke salinitas 2 ppt selama 96 jam. Sementara di tempat lain, dilaporkan udang ini mampu diaklimasi hingga salinitas 1 ppt selama 48 jam dengan sintasan sekitar 97% (McGraw et al. 2002; Davis et al. 2002). Teknik aklimasi yang diterapkan perlu diperbaiki dan disesuaikan dengan karakteristik lingkungan budidaya. Oleh karena itu, untuk keberhasilan budidaya udang vaname di air bersalinitas rendah, dibutuhkan pengembangan teknik aklimasi yang baru.
untuk peningkatan pH media dan pembentukan eksoskeleton juga esensial dalam proses osmoregulasi (Larvor 1983; Boyd 1988; Cheng et al. 2006). Fungsi kalsium menurut Wood (2000) yaitu menentukan permeabilitas sel untuk keseimbangan osmotis, sehingga kondisi stres dapat berkurang. Informasi tentang konsentrasi minimum kalsium media untuk sintasan udang vaname masih terbatas, sedangkan pada jenis krustasea lain telah diuji. Peran kalsium telah dilaporkan pada Penaeus monodon (Irawan 1988; Edi 1990), Callinectes sapidus (Neufeld & Cameron 1994; Perry et al. 2001), Gammarus lacustris dan Astacus astacus (Rukke 2002), Cherax quadricarinatus (Kaligis 2005), serta Paranephrops zealandicus (Hammond et al. 2006).
Setelah diadaptasikan pada air bersalinitas rendah, sintasan postlarva vaname yang dihasilkan dalam pemeliharaan selanjutnya masih rendah sehingga dibutuhkan penambahan mineral lain. Secara umum, faktor pembatas terhadap sintasan dan pertumbuhan udang di perairan salinitas rendah adalah komposisi mineral yang kurang dibandingkan di perairan salinitas normal. Proses-proses fisiologis dapat berlangsung secara normal bergantung dari ketersediaan anion (bikarbonat, karbonat, klorida, dan sulfat) serta kation tertentu (kalsium, magnesium, potasium, dan natrium) (Roy 2006). Berbagai penelitian telah mengungkapkan bahwa mineral yang krusial berpengaruh terhadap sintasan udang vaname di air bersalinitas rendah adalah potasium (K+) (Davis et al. 2002; Saoud et al. 2003; McGraw & Scarpa 2003; Davis et al. 2005). Potasium berperan
penting karena dalam metabolisme krustasea, mineral ini terhubungkan dengan aktivitas enzim osmoregulasi, Na+K+-ATPase (Larvor 1983; McGraw &
Scarpa 2003).
Udang vaname termasuk jenis penaeid yang melakukan regulasi hiperosmotik ketika terjadi perubahan ke media bersalinitas rendah. Adanya penurunan salinitas dapat menyebabkan kondisi stres sehingga udang akan berusaha mempertahankan tekanan osmotik tubuh dengan mengekstrak elektrolit dari lingkungan kemudian mempertahankan agar tidak terjadi difusi ion ke luar tubuh. Untuk menghadapi kondisi seperti ini, diperlukan energi ekstra yang dapat membantu postlarva mempertahankan vitalitas.
Beberapa penelitian telah membuktikan peran nutrien khusus terhadap sintasan udang saat kondisi stres lingkungan. McGraw et al. (2002) telah memperlihatkan bahwa sintasan postlarva vaname dapat meningkat apabila pakan Artemia tetap diberikan selama aklimasi ke air bersalinitas rendah. Telah diketahui sintasan postlarva dapat dipertahankan tetap tinggi dengan penambahan nutrien seperti HUFA. Ketika dilakukan penurunan mendadak salinitas dalam jangka pendek (24 jam), pengkayaan Artemia dengan asam lemak (HUFA) mampu meningkatkan sintasan udang windu (P.monodon) (Lavens & Sorgelos
2000). Hasil yang sama dilaporkan dalam pemeliharaan udang vaname (L. vannamei) melalui pengkayaan rotifer dengan HUFA (Wahyudin 2005).
Perumusan Masalah
Permasalahan yang dihadapi yaitu masih rendahnya sintasan postlarva ketika aklimasi ke air bersalinitas rendah, serta belum maksimalnya pertumbuhan selama pemeliharaan selanjutnya. Adanya penerapan aklimasi salinitas yang belum sesuai menyebabkan beban osmotik tetap tinggi saat mencapai salinitas rendah. Beban osmotik yang terus meningkat disertai kehilangan ion-ion penting dalam tubuh akan menurunkan sintasan postlarva vaname. Rendahnya pertumbuhan disebabkan metabolisme dan ganti kulit yang tidak berlangsung lancar. Penyebabnya antara lain kualitas pakan rendah sehingga nutrien yang dibutuhkan tidak memadai. Berbagai masalah tersebut akan menyebabkan ketidakcukupan benih vaname berkualitas untuk budidaya di air salinitas rendah.
Kemampuan udang untuk melakukan proses-proses fisiologis secara normal ditentukan oleh tersedianya mineral penting dalam media. Penambahan kalsium karbonat (CaCO3) dan potasium karbonat (K2CO3) dalam media dapat
mempertahankan kualitas air (buffer pH) serta untuk fungsi osmoregulasi udang. Kalsium (Ca2+) berperan penting dalam menjaga keseimbangan asam basa dan potensial membran bagi transport ion Na+ dan Cl-, sedangkan potasium (K+) diperlukan untuk mempertahankan konsentrasi K+ dalam sel yang mempengaruhi aktivitas enzim Na+K+-ATPase sehingga proses metabolisme tetap berjalan normal saat terjadi fluktuasi salinitas lingkungan. Bila kondisi media optimal dapat dicapai, akan terjadi sinergi dalam mekanisme osmoregulasi yang menurunkan beban osmotik. Dengan demikian, ketersediaan energi makin banyak teralokasi untuk mempertahankan sintasan dan meningkatkan pertumbuhan udang selama pemeliharaan di air bersalinitas rendah.
dipengaruhi jumlah asupan protein, sehingga keseimbangan kedua komponen ini dibutuhkan dalam pakan. Secara skematis, bagan alir perumusan masalah disajikan dalam Gambar 1.
Tujuan dan Manfaat Penelitian
Tujuan dari penelitian adalah :
1. menentukan kadar kalsium optimal bagi sintasan postlarva melalui evaluasi osmolaritas dan laju metabolisme selama aklimasi ke salinitas rendah
2. menentukan kadar potasium optimal bagi kandungan potasium tubuh, gradien osmotik, tingkat konsumsi oksigen, sintasan, laju pertumbuhan bobot rerata
harian, dan efisiensi pemanfaatan pakan selama pemeliharaan di salinitas 2 ppt
3. mengkaji pengaruh pakan yang berkadar protein dan kalsium berbeda terhadap kinerja pertumbuhan postlarva vaname di media salinitas 2 ppt.
Manfaat dari penelitian ini adalah mendapatkan informasi yang dijadikan landasan dalam pengembangan teknik pemeliharaan vaname pada lingkungan air bersalinitas rendah.
Hipotesis
Berdasarkan tujuan penelitian, maka hipotesis yang diajukan sebagai berikut:
1. apabila kadar kalsium dalam media mampu secara efektif mempertahankan keseimbangan mineral dalam tubuh maka beban osmotik semakin rendah sehingga sintasan tetap tinggi pada saat aklimasi ke media salinitas rendah 2. apabila kadar potasium media mampu meningkatkan efisiensi osmotik maka
ketersediaan energi semakin banyak teralokasi untuk menunjang sintasan dan pertumbuhan udang vaname selama pemeliharaan di salinitas rendah
Novelty
+ +
Gambar 1 Bagan alir perumusan masalah pengaruh penambahan Ca2+ selama aklimasi, penambahan mineral K+ dalam media, serta pengkayaan protein dan Ca2+ dalam pakan bagi kualitas dan kuantitas postlarva udang vaname.
TINJAUAN PUSTAKA
Biologi Udang Vaname (L. vannamei)Udang vaname termasuk dalam ordo dekapoda yang memiliki berbagai ciri. Ordo ini mempunyai 10 kaki. Karapas berkembang dengan baik sehingga menutupi seluruh kepala dan thoraks. Panjang tubuh vaname dapat mencapai 230 mm atau 9 inci. Bobot maksimal udang betina sekitar 120 g, sedangkan jantan relatif lebih ringan. Keseluruhan tubuh berwarna putih, termasuk kaki berwarna
putih atau pucat. Udang vaname agak sulit dibedakan dari udang biru (L. stylirostris) walaupun diketahui memiliki penyebaran geografis yang sama.
Udang vaname banyak ditemukan di perairan lebih dalam (Bailey-Brock & Moss 1992; Eloovara 2001; Wyban & Sweeney 1991).
Genus Penaeus memiliki ciri umum adanya gigi pada rostrum serta antena yang panjang (Gambar 2). Pada bagian ventral dari rostrum terdapat 2 gigi, serta 8 – 9 gigi di bagian dorsal. Antena beserta sepasang kaki pertama berfungsi untuk mengambil makanan. Sub genus Litopenaeus digambarkan dengan adanya telikum terbuka tanpa tempat penyimpanan sperma pada udang betina (Wyban & Sweeney 1991) .
Walaupun tergolong jenis omnivora atau pemakan detritus, dari penelitian terakhir berdasarkan kandungan perut, udang penaeid termasuk jenis karnivora yang memakan krustasea kecil, amphipods, dan polikaeta di alam. Jenis makanan ini tidak tersedia di kolam budidaya intensif sehingga udang hanya makan pakan buatan dan detritus kolam (Wyban & Sweeney 1991).
Udang vaname termasuk katadromos yaitu udang dewasa hidup di laut sedangkan udang muda akan berpindah ke daerah pantai. Udang ini di lingkungan alam mendiami dasar perairan berlumpur dari garis pantai hingga kedalaman sekitar 72 meter (235 kaki) dan menghuni daerah mangrove yang terlindung dari gangguan luar. Di habitat aslinya perairan Amerika Selatan, Tengah, dan Utara, udang akan matang kelamin, kawin, dan bertelur di kolom air laut terbuka
Gambar 2 Anatomi lengkap L. vannamei (Wyban & Sweeney 1991).
di dasar perairan dangkal. Laut perairan dangkal terdapat nutrien melimpah, serta salinitas dan suhu air lebih bervariasi dibandingkan laut terbuka. Setelah beberapa bulan di daerah estuari, udang dewasa akan kembali ke lingkungan laut dalam, dan mengalami kematangan seksual, kawin, dan bertelur (Eloovara 2001; Wyban & Sweeney 1991).
Perkembangan Larva
Perkembangan larva merupakan perubahan stadia saat larva hingga postlarva (Tabel 1). Menurut Wyban dan Sweeney (1991), setelah fertilisasi, telur vaname menetas sekitar 14-16 jam kemudian. Memasuki tahapan larva, perkembangan melalui 3 stadia, yaitu nauplius, zoea, dan mysis. Setelah itu berlangsung tahapan postlarva. Pergantian stadia terjadi setelah larva mengalami molting. Lamanya waktu untuk setiap stadia berbeda, yaitu stadia nauplius sekitar 5 hari, zoea dan mysis sekitar 3 hari, selanjutnya tahapan postlarva yang berlangsung lebih lama. Eloovara (2001) menyatakan bahwa postlarva (PL) mengalami perubahan dari PL 1 - PL 10 (sering sampai PL 20).
Tabel 1 Waktu serta karakteristik setiap stadia larva udang vaname saat berkembang pada kondisi normal (Wyban & Sweeney 1991)
Hari
ke Stadia Karakteristik
1 Naupli - 1 badan berbentuk bulat dengan panjang 0,40 mm, antena pertama terdiri 3 setae, antena kedua terdiri 5 setae
2 Naupli - 2 ujung antena pertama terdapat setae yang satu panjang, dan dua buah yang pendek, antena kedua terdiri 6 setae 3 Naupli - 3 dua buah furcal masing-masing memiliki 3 duri, antena
kedua terdapat 7 setae
4 Naupli - 4 masing-masing furcal terdapat 5 duri, antena kedua terdiri 8 setae, terjadi segmentasi di antena
5 Naupli - 5 terlihat jelas setation antara antena pertama dan kedua, tubuh lebih pipih
6 Zoea - 1 panjang tubuh sekitar 1,0 mm, tubuh terpisah dua bagian: thoraks (kepala) dan abdomen (ekor), makanan dapat terlihat bila larva sedang makan
8 Zoea - 2 mata telah bertangkai, rostrum dan duri di atas mata 9 Zoea - 3 sepasang uropoda biramus mulai berkembang, duri pada
ruas-ruas tubuh mulai tampak
10 Mysis - 1 badan seperti udang dewasa dengan panjang sekitar 3,4 mm, kuncup pleopoda, telson dan uropoda mulai berkembang
11 Mysis - 2 kuncup pleopoda telah ada tapi belum bersegmen
12 Mysis - 3 pleopoda bertambah panjang dan beruas-ruas, tampak duri pertama dorsal di rostrum
13 Postlarva panjang tubuh sekitar 3,4 mm, telah ada setae renang, udang telah mampu berenang ke depan
lakunya. Saat larva, udang vaname bersifat planktonik, kemudian berubah menjadi bentik saat postlarva. Udang tahap larva akan berpindah tempat dari laut terbuka ke arah pantai dan estuari sampai menjadi dewasa (Wyban & Sweeney 1991; Elovaara 2001).
Pertumbuhan dan Ganti Kulit
merupakan pertambahan jaringan akibat dari pembelahan sel secara mitosis yang terjadi bila ada kelebihan input energi dan asam amino (protein) yang berasal dari makanan (Effendie 2002). Pada krustasea, pertumbuhan terjadi secara berkala setelah pergantian kulit. Pertambahan panjang dan bobot tubuh akan terhambat bila tidak didahului oleh ganti kulit. Saat kondisi nutrisi memadai dan lingkungan mendukung, pertumbuhan terjadi secara optimal (Wyban & Sweeney 1991; Wickins & Lee 2002).
Pertumbuhan sebenarnya merupakan ekspresi dari proses sintesis protein. Proses ini terjadi pada kelompok-kelompok sitoplasmik yang sangat kecil yang disebut ribosom. Madigan et al. (2009) menyatakan bahwa proses sintesis protein diawali kode genetik dalam urutan nukleotida tertentu dalam DNA yang ditranskripsikan ke dalam urutan nukleotida dalam RNA. Kode genetik ditulis dalam bentuk messenger RNA (mRNA) yang selanjutnya ditranslasi melalui penggabungan kelompok RNA lain (tRNA, rRNA) dalam ribosom untuk mengarahkan ke sintesis protein. Dalam proses ini, dibutuhkan bahan dasar asam amino sitoplasma, katalisator enzim RNA-polimerase saat transkripsi dan aminoacyl-tRNA synthetases saat translasi, serta sumber energi dari ATP.
Asam ribonukleat (RNA) memegang peranan penting dalam proses sintesa protein. Busacker et al. (1990) menyatakan bahwa konsentrasi RNA dapat digunakan sebagai pengukur laju pertumbuhan. Awalnya dari hipotesis bahwa laju input pakan menentukan konsentrasi RNA pada jaringan sebagaimana laju input pakan menentukan laju sintesis protein. Ini teramati dari hubungan
langsung antara konsentrasi RNA dengan sintesis protein, kemudian terjadi pertumbuhan jaringan. Konsentrasi RNA akan berfluktuasi bergantung dari aktivitas sintesis protein, sebaliknya konsentrasi DNA relatif konstan dalam sel (Houlihan et al. 1993).
sehingga salinan yang banyak dapat membuat jutaan ribosom yang dibutuhkan untuk sintesis protein yang aktif.
Untuk mengestimasi pertumbuhan, penggunaan asam nukleat dalam bentuk rasio RNA/DNA merupakan metode yang cukup akurat, selain juga dapat menjadi indikator kondisi nutrisi ikan (Rooker & Holt 1996). Parameter ini telah diuji pada beberapa jenis ikan. Raae et al. (1988) melaporkan bahwa larva ikan cod (Gadus morhua) yang dipelihara tanpa pemberian pakan memperlihatkan penurunan nilai ratio RNA/DNA. Sebaliknya, pada ikan yang diberi pakan, nilai rasio RNA/DNA relatif konstan walau terjadi sedikit fluktuasi. Tarkii et al. (1994) juga dengan sukses menggunakan rasio RNA/DNA dalam memprediksi pertumbuhan pada ikan striped jack (Caranx delicatissimus). Hasil penelitian selama 42 hari pada larva menunjukkan adanya peningkatan rasio RNA/DNA dalam tubuh seiring dengan meningkatnya pertumbuhan ikan. Selanjutnya, Nandeesha et al. (2000) melaporkan bahwa penggantian sumber protein tepung ikan ke silkworm pupae mampu mempengaruhi rasio RNA/DNA, pertumbuhan, rasio konversi pakan, dan retensi protein ikan mas (Cyprinus carpio). Peningkatan ratio RNA/DNA terjadi pada kadar pupae 50% (8,29±0,51) dibandingkan kadar pupae 30% (7,15±0,28) dan 40% (6,69±0,05) yang mengindikasikan peningkatan efisiensi minyak pupae sebagai sumber energi bagi pertumbuhan.
Seperti halnya arthropoda lain, pertumbuhan udang ditentukan oleh frekuensi molting dan peningkatan pertumbuhan. Frekuensi molting adalah fungsi dari ukuran udang yaitu waktu antara peningkatan molting, sedangkan peningkatan pertumbuhan adalah seberapa besar pertumbuhan terjadi setiap molting baru. Untuk tumbuh, tubuh udang yang tertutup karapas keras harus melepaskan karapas tua dan mengembangkan karapas baru yang lebih ringan dan besar (Wyban & Sweeney 1991). Kecepatan pertumbuhan akan meningkat apabila kondisi lingkungan dan nutrisi cocok (Wickins & Lee 2002).
air. Lapisan membran yang merupakan lapisan terdalam endokutikula tidak mengalami kalsifikasi (Dennell 1960). Saat ganti kulit, akan terjadi pembagian pada kutikula (Lowery 1988).
Daur ulang kalsium terjadi pada keseluruhan tahapan ganti kulit. Menurut Perry et al. (2001), proses mineralisasi kulit (eksoskeleton) secara kimia merupakan pembentukan kalsium karbonat dan ion hidrogen dari ion kalsium dan bikarbonat dalam kutikula. Persamaan reaksi sebagai berikut :
Ca2+ + HCO3¯ CaCO3 + H+
Ion hidrogen yang diproduksi selama reaksi ini secara aktif ditransportasi dari kutikula ke darah (hemolimp) serta dari darah ke media eksternal melewati insang. Jumlah kalsium untuk mineralisasi berasal dari air laut dengan penambahan minimal dari kalsium yang tersimpan di tubuh. Sedangkan bikarbonat selain dibawa langsung dari air laut juga dari CO2 yang dihasilkan
selama respirasi. Kecepatan pengambilan kalsium dan bikarbonat bergantung pada konsentrasi ion-ion ini dalam air laut. Pada ketam biru (C. sapidus), jumlah bersih kalsium yang masuk ke tubuh saat tahap postmolt mencapai 26% dari kadar kalsium di air laut.
Pengerasan kutikula berlangsung 2 – 3 jam setelah ecdysis melalui pembentukan protein kulit serta deposisi kalsium dan bikarbonat, sedangkan
eksoskeleton baru akan kembali normal (ke tahap intermolt) setelah sekitar 7 sampai 10 hari postmolt. Total kandungan mineral dalam kulit saat mineralisasi
adalah setengah hingga satu kali dari konsentrasi saat intermolt (Perry et al. 2001).
Passano (1960) mengemukakan bahwa ganti kulit pada dekapoda merupakan proses kompleks yang terdiri atas tahapan premolt, molt, postmolt, dan intermolt. Urutan utama siklus molting adalah sebagai berikut (Wickins & Lee 2002):
(1) akumulasi cadangan mineral dan bahan organik
(2) pemindahan material dari kulit lama dan pembentukan eksoskeleton baru (3) ecdysis (molting) diiringi penyerapan air
(5) pergantian cairan melalui pertumbuhan jaringan tubuh.
Siklus ganti kulit merupakan proses yang dipengaruhi berbagai kelenjar dan organ. Permulaan ganti kulit dikontrol oleh MIH (Molt Inhibiting Hormone) yang dibentuk sel neurosekretori tertentu dalam organ X. Sebelum dilepas ke hemolimp, MIH disimpan di kelenjar sinus yang akan dilepas dari akhiran axon lebih besar pada struktur ini (Passano 1960). Organ Y merupakan sumber dari hormon molting yang disekresikan sebagai suatu prekursor (ecdysone), selanjutnya di hemolimp berubah menjadi hormon aktif, 20-OH-ecdysone. MIH dapat menghambat proses molting melalui penghambatan sekresi ecdysteroid dari organ Y (Huberman 2000).
Jika udang tumbuh, frekuensi molting juga akan meningkat. Frekuensi molting secara alamiah bergantung pada suhu dan spesies. Udang penaeid saat stadia larva mengalami molting 2-3 kali dalam sehari, kemudian saat juvenil akan molting setiap 3-25 hari (Wickins & Lee 2002). Saat suhu 28oC, udang vaname tahap larva mengalami molting setiap 30-40 jam, tahap juvenil yang berukuran 1-5 g setiap 4-6 hari, dan udang dewasa ukuran 15 g akan molting dengan interval 2 minggu (Wyban & Sweeney 1991).
Osmoregulasi
Osmoregulasi merupakan suatu bentuk sistem homeostasis yang berperan penting dalam mengatur keadaan stabil atau mengontrol fluktuasi lingkungan internalnya (Affandi & Tang 2002). Pengaturan osmotik cairan tubuh juga diartikan sebagai pengaturan total konsentrasi partikel dari cairan pada tingkatan berbeda terhadap media eksternal (Robertson 1960). Selama osmoregulasi, hewan air membutuhkan keseimbangan osmotik antara cairan tubuh dan media yang sangat krusial bagi kelangsungan kehidupannya. Spesies air tawar dapat kehilangan ion-ion saat berpindah ke lingkungan hipotonik atau ketika bertahan dari hidrasi (Davis & Gatlin III 1991).
tertentu walaupun terjadi fluktuasi salinitas lingkungan. Dekapoda yang secara khusus berasal dari air payau termasuk pada poikilosmotik pada kisaran salinitas normal atau tinggi, dan homoiosmotik pada salinitas rendah (Robertson 1960).
Jenis dekapoda yang darahnya isoosmotik dengan air laut normal menunjukkan jenis pengaturan hiperosmotik di media pengenceran air laut. Krustasea ini melakukan pengambilan aktif garam dari medium eksternal dalam upaya mengimbangi kehilangan garam karena difusi keluar atau sekresi antennal. Karena ion hilang selama sekresi isoosmotik, pengambilan air bersama ion-ion dari medium harus dikontrol untuk mempertahankan keseimbangan air yaitu lewat adanya permukaan-permukaan sel yang permeabel air. Hasil bersih ion-ion akan diserap dari cairan sehingga konsentrasi osmotik sama antara darah dan urin (Robertson 1960).
Dalam sel-sel klorida insang atau terletak pada bagian perpanjangan sistem tubular sitoplasma, terdapat suatu Na+K+-activated ATPase yang berfungsi sebagai pompa ion. Enzim ini secara ekstensif dan aktif melakukan transport Na+ keluar dari sel untuk bertukar dengan K+ ke dalam sel (Imsland et al. 2003). Kurangnya kebutuhan energi untuk pompa ion pada salinitas optimal dapat mengarahkan pada pengurangan aktivitas sel-sel klorida yang kemudian akan mengurangi aktivitas enzim Na+K+-ATPase. Hasil penelitian menunjukkan bahwa pada postlarva vaname aktivitas enzim ini sekitar 46,8±13,2 mol fosfat/g.j di media salinitas 30 ppt, sedangkan di media salinitas 10 ppt sekitar 79,9±19,6 mol
fosfat/g.j (Palacios et al. 2004). Kebutuhan energi untuk aktivitas enzim Na+K+-ATPase secara umum lebih rendah di lingkungan yang isoosmotik,
sehingga energi yang disimpan cukup substansial untuk meningkatkan sintasan dan pertumbuhan (Imsland et al. 2003; Palacios et al. 2004).
mOsm/l H2O. Osmolaritas hemolimp kemudian meningkat menjadi 1102±50,27
mOsm/l H2O setelah 60 hari pemeliharaan ketika ukuran udang 4,27±0,76 gram.
Pada pengukuran hari ke-20, udang yang dipelihara pada salinitas 46 ppt menunjukkan osmolaritas hemolimp tertinggi sekitar 1001,00±41,6 mOsm/l H2O,
yang berbeda nyata dibanding perlakuan salinitas lainnya (10, 22, 34 ppt). Osmolaritas hemolimp terendah, 664,00±57,70 mOsm/l H2O dihasilkan pada
pemeliharaan di salinitas 10 ppt. Hasil pengukuran lainnya menunjukkan indeks hepatosomatik dan indeks otot ekor lebih rendah pada udang di salinitas 46 ppt yang mengindikasikan penggunaan energi lebih banyak dari hepatopankreas untuk aktivitas osmoregulasi pada kondisi salinitas tersebut.
Peranan Mineral Kalsium dan Potasium
Kalsium adalah mineral yang esensial untuk struktur jaringan keras, pembekuan darah, kontraksi otot, transmisi saraf, osmoregulasi, dan sebagai kofaktor proses-proses enzimatik (Larvor 1983; Cheng et al. 2006). Elemen ini ditemukan terbanyak kelima di kulit bumi, serta terbanyak dalam tubuh hewan yakni 1-2% dari total bobot tubuh. Kalsium tidak terdapat dalam bentuk bebas, namun berupa kation yang bermuatan dua ion positif (Piliang 2005).
Penambahan kalsium karbonat (CaCO3) pada kolam budidaya bertujuan
untuk: 1) menetralkan ion Al, Fe, Mn, H, dan 2) menambah unsur Ca2+ ke dalam perairan. Boyd (1988) menyatakan bahwa penetral utama dalam kapur yaitu karbonat (CO32-) yang menghasilkan OH-, sehingga akan merangsang
perombakan bahan organik yang mengandung Al, Fe, Mn menjadi dipercepat. Bahan kapur (CaCO3) di perairan juga akan bereaksi dengan CO2 yang kemudian
membentuk kalsium dan bikarbonat. Persamaan reaksi kesetimbangan yang terjadi antara calcite (CaCO3) dengan bikarbonat (HCO-3) sebagai berikut :
CaCO3 + CO2 + H2O Ca2+ + 2 HCO3-
Konsentrasi kalsium (Ca2+) akan meningkat setelah pengapuran. Calcite sebenarnya memiliki daya larut rendah, namun kelarutan bahan ini akan meningkat jika karbondioksida melimpah. Bikarbonat mengandung sifat basa karena tersusun atas karbonat (CO32-). Melalui reaksi hidrolisis selanjutnya
alkalinitasnya rendah akan menyebabkan peningkatan pH pada pagi hari karena adanya peningkatan pada sistem penyangga (peningkatan ion OH-) yang selanjutnya menyebabkan perubahan pH air harian tidak terlalu berfluktuatif (Boyd 1988).
Fungsi kalsium dalam proses osmoregulasi sehubungan dengan stabilitas
permeabilitas branchial insang dalam mengatur transport natrium (Na+) dan air (Potts 1984). Menurut Wood (2000), kalsium akan berikatan dengan komponen
protein membran untuk mengubah fluiditas dalam membran. Fungsi kalsium berhubungan dengan perkembangan chanel khusus dalam membran sel yang berpengaruh penting dalam menentukan permeabilitas sel dari ion Na+ dan K+. Efek kalsium melalui kandungan ion ini dalam pori sel yang dapat mempengaruhi transepithial menjadi positif yaitu mengurangi pengeluaran Na+ melebihi pengeluaran Cl- dalam sel (Potts 1984). Suatu penelitian telah melaporkan bahwa konsentrasi kalsium eksternal berpengaruh terhadap keseimbangan osmotik pada ikan pinfish air laut (Lagodon rhomboides) selama pemeliharaan di air bersalinitas rendah (Carrier & Evans 1976). Konsentrasi kalsium media sekitar 10 mmol dapat
mempengaruhi permeabilitas insang untuk ion Na+ sehingga kadar ion ini dalam tubuh tetap tinggi. Namun, peningkatan efflux ion Na+ ke luar tubuh
ditunjukkan pada media tanpa suplementasi Ca2+ yang meningkatkan mortalitas ikan hingga 96%.
Fungsi kalsium lainnya adalah dalam pembentukan jaringan keras dari tulang, eksoskeleton, sisik, dan gigi. Selama proses pembentukan jaringan tubuh ini, kalsium berperan utama karena 99% kalsium dalam tubuh terdapat dalam jaringan terutama tulang atau eksoskeleton (Piliang 2005). Secara singkat, Larvor (1983) menyebutkan bahwa proses pembentukan eksoskeleton meliputi produksi awal osteoid dari osteoblasts, pematangan osteoid, permulaan mineralisasi, kemudian terjadi mineralisasi.
pusat kristalisasi. Mukopolisakarida sendiri memiliki afinitas tinggi ion kalsium dan fosfor. Sebelum mineral-mineral mulai terdeposit dalam zona mineralisasi, oksigen disalurkan dalam aliran darah. Kemudian, proses oksidasi dan produksi ATP terjadi secara intensif. Peningkatan aktivitas osteoblast melalui enzim alkaline phosphatase mempengaruhi produksi substansi dasar dan kompleks protein karbohidrat. Adanya hidrolisis kolagen ini melepas sejumlah besar hidroksiprolin yang akan terbuang ke urin. Kelompok reaktif yaitu sisi rantai asam amino yang memiliki kemampuan spesifik sebagai pusat kristalisasi dilepaskan membentuk jaringan kolagen. Kelompok-kelompok reaktif kolagen selanjutnya bereaksi dengan fosfat dan kalsium yang sebelumnya mengelilingi mukopolisakarida, dan membentuk pusat kristalisasi. Komposisi penyusun jaringan tulang terdiri dari bahan dasar organik/kolagen (38%), anorganik (32%), dan air (20%) yang dapat bervariasi menurut umur, makanan, dan tempat hidup. Gambar 3 Reaksi-reaksi metabolisme dalam mineralisasi tulang
(Georgievskii VI 1982).
Sintesis protein Metabolisme lipid dan
Kebutuhan kalsium untuk udang laut secara individual dan interaksinya telah terkandung dalam alkalinitas, pH, dan karbondioksida dari air kultur sehingga udang tidak membutuhkan suplemen kalsium pakan. Kandungan dalam media dapat mempengaruhi pengambilan kalsium oleh udang serta metabolisme yang terjadi saat mineralisasi kulit (eksoskeleton). Sedangkan udang air tawar yang hidup di perairan dengan kesadahan rendah membutuhkan suplementasi kalsium pakan. Sebagai contoh, alkalinitas tinggi sampai di atas 150 mg/l akan mengurangi toleransi kalsium pakan pada udang galah (M. rosenbergii). Namun, jika alkalinitas di bawah 100 mg/l dengan kesadahan berkisar antara 25-100 mg/l, penambahan kalsium dalam pakan akan meningkatkan sintasan dan
pertumbuhan juvenil udang air tawar ini. Sedangkan untuk golongan penaeid, alkalinitas yang diperlukan lebih tinggi sekitar 150-200 mg/l CaCO3 (Wickins &
Lee 2002).
penambahan kapur sebanyak 75 mg/l dengan biji bungkil teh dalam media (Edi 1990).
Efek kalsium juga telah diinvestigasi dari beberapa penelitian di media air tawar. Rukke (2002) melaporkan bahwa dalam pemeliharaan di danau perairan
lunak, G. lacustris dan A. astacus membutuhkan kandungan kalsium antara 5-20 mg/l dalam media untuk meningkatkan sintasan, bobot kering kalsium, dan
persentase kalsium tubuh, sedangkan konsentrasi kalsium di bawah 5 mg/l dapat membatasi produksi kedua jenis krustasea ini. Kaligis (2005) menyimpulkan
bahwa alkalinitas minimum 110 ppm yang dicapai dengan penambahan
kalsium karbonat (CaCO3) dibutuhkan pada pemeliharaan cherax red claw
(C. quadricarinatus). Adanya peningkatan efisiensi pakan dan frekuensi ganti kulit berkontribusi langsung ke pertumbuhan lobster air tawar tersebut. Berdasarkan penelitian pada crayfish air tawar P. zealandicus, Hammond et al.
(2006) melaporkan bahwa perlakuan konsentrasi kalsium minimum sekitar 10 mg/l mampu meningkatkan sintasan, namun perlakuan kalsium (0, 5, 10, 20,
80 mg/l) tidak berpengaruh pada pertumbuhan dan ganti kulit jenis krustasea ini. Potasium (K+) adalah suatu elemen intraselluler yang penting. Ion ini sangat berpengaruh dalam metabolisme ketika terjadi pengeluaran energi dibutuhkan untuk menjaga konsentrasi konstan gradien melewati dinding sel. Berbagai jenis bahan yang dibutuhkan sel dibawa melalui transpor aktif natrium (Na+) yang terhubungkan dengan transpor potasium (K+) di bagian dalam sel melalui sepasang pompa ion. Sistem ini menggunakan energi dari ATP yang termanfaatkan untuk aktivitas Na+K+-ATPase (Larvor 1983). Pada perairan air payau dan tawar, potasium merupakan unsur pokok yang ditemukan sedikit. Konsentrasi potasium mempengaruhi aktivitas enzim Na+K+ATP-ase pada krustasea yaitu dalam mempertahankan aktivitas enzim ini tetap konstan ketika terjadi fluktuasi salinitas lingkungan perairan (McGraw & Scarpa 2003).
dengan mineral lain menunjukkan bahwa sintasan postlarva vaname tetap tinggi ketika dilakukan penambahan K+ sebanyak 10 ppm dalam media air tawar. Nilai sintasan dicapai sekitar 83-97% pada pemeliharaan selama 24 jam dan 77-83% pada pemeliharaan 48 jam. Sedangkan penggunaan mineral lain seperti Ca2+, Mg2+, dan SO4 menunjukkan kecenderungan penurunan sintasan.
Penambahan K+ juga berpengaruh terhadap sintasan dan pertumbuhan udang vaname dalam jangka waktu pemeliharaan lebih lama. Davis et al. (2005) melaporkan bahwa penambahan K+ (bentuk larutan KCl dan KCl-MgCl) dalam media mampu mempertahankan sintasan benih vaname relatif tinggi (76,5-89,0%) setelah pemeliharaan 21 hari di media salinitas 4 ppt. Selanjutnya, dari hasil penerapan di tambak bersalinitas rendah, McNevin et al. (2004) melaporkan terjadi peningkatan produksi udang vaname mencapai 4.068 kg/hektar (awalnya 595 kg/hektar) ketika kandungan K+ dan Mg2+ ditingkatkan dari 6,2 ppm K+ ke 40 ppm K+ dan 4,6 ppm Mg2+ ke 20 ppm Mg2+. Sumber K+ dan Mg2+ yang digunakan adalah potasium klorida (KCl) dan potasium magnesium sulfat (Kmag: K2SO4.2MgSO4).
Roy et al. (2007) melaporkan bahwa konsentrasi K+ media berbeda berpengaruh terhadap sintasan dan pertumbuhan udang vaname. Hasil penelitian selama pemeliharaan 14 hari dengan menggunakan postlarva menunjukkan bahwa media dengan konsentrasi K+ 10, 20, 40 ppm dalam media buatan 4 ppt mampu mempertahankan sintasan sekitar 46,3-55,0%, sedangkan pada konsentrasi K+ terendah (5 ppm), nilai sintasan yang dicapai hanya 21,25%. Nilai sintasan dan pertumbuhan individu tertinggi ditunjukkan pada media pengenceran air laut 4 ppt (kontrol) yaitu sekitar 78% dan 0,132 g. Hasil uji selanjutnya dengan menggunakan juvenil vaname selama pemeliharaan 49 hari menunjukkan
peningkatan nilai sintasan pada perlakuan K+ 10-40 ppm serta kontrol (93,3-96,7%). Seperti hasil penelitian pertama, sintasan terendah (23,3%)
Kebutuhan Nutrien Udang
Protein merupakan komponen pakan yang dibutuhkan sebagai sumber energi maupun dalam pembentuk jaringan tubuh. Apabila kebutuhan energi dari sumber karbohidrat dan lemak tidak mencukupi, konsumsi protein berlebih dari yang dibutuhkan, ataupun kualitas protein rendah, maka protein akan digunakan sebagai sumber energi dan terbatas untuk pertumbuhan (Phillips Jr 1969). Berbeda dengan hewan mamalia, ikan dan hewan air umumnya memperoleh energi dari protein (Brett & Groves 1979).
Susunan asam amino yang terkandung dalam bahan sumber akan menentukan kualitas protein. National Research Council (1983) menyatakan bahwa udang membutuhkan 10 jenis asam amino esensial untuk pertumbuhan, yaitu : arginin, histidin, isoleusin, leusin, lisin, valin, metionin, fenilalanin, treonin dan triptofan. Untuk kebutuhan energi, asam amino dideaminasi sehingga menghasilkan amoniak dan rantai karbon. Selanjutnya terjadi oksidasi menghasilkan energi. Bentuk sampingan diekskresikan melalui insang dan ginjal. Ekskresi nitrogen akan lebih rendah dari ikan yang diberi pakan protein kualitas tinggi dibanding protein kualitas rendah. Kualitas protein tergantung dari kecernaan dan nilai biologis protein yang ditentukan oleh kualitas dan kuantitas asam amino penyusun protein (Furuichi 1988).
Kebutuhan protein didefinisikan sebagai jumlah protein yang dibutuhkan setiap hewan air atau setiap biomassa perhari yang disesuaikan kecernaan pakan (Kureshy & Davis 2002). Beberapa faktor biotik yang mempengaruhi kebutuhan protein yaitu spesies, keadaan fisiologis, ukuran, dan karakteristik pakan (kualitas protein dan ratio energi protein), sedangkan faktor abiotik adalah suhu dan salinitas.
Berbagai penelitian telah melaporkan akan kebutuhan protein vaname di air bersalinitas tinggi. Colvin dan Brand (1977), diacu dalam Cuzon et al.
pertumbuhan juvenil dan pradewasa vaname lebih tinggi dengan pemberian protein pakan 32% dibandingkan 15% dan 48%. Akan tetapi, pemberian pakan berkadar protein 48% menghasilkan nilai efisiensi pakan lebih tinggi dibandingkan kadar protein 32% yang mengindikasikan kadar protein optimum yang dibutuhkan kemungkinan di atas 32%. Pascual et al. (2004) kemudian melakukan evaluasi pengaruh pakan dengan kadar protein 5%, 15%, dan 40% terhadap pertumbuhan, efisiensi asimilasi, dan kapasitas imun udang. Laju pertumbuhan tertinggi juvenil dicapai dengan pemberian pakan berkadar protein 40%, sedangkan terendah dengan pakan berkadar protein 5%. Penurunan laju pertumbuhan pada perlakuan protein 5% berkaitan dengan terjadinya peningkatan metabolisme dan penurunan kapasitas imun lewat penurunan OxyHC (hemosianin), konsentrasi hemosit, dan kapasitas phagositosis. Berdasarkan hasil penelitian tersebut, disarankan pemberian protein dalam pakan untuk juvenil vaname di atas 35%.
Sehubungan dengan terjadinya stres fisiologis unik yang diiringi peningkatan ongkos energetik saat budidaya udang di perairan salinitas rendah, maka secara potensial kebutuhan protein dapat berbeda dengan kebutuhan protein di air salinitas tinggi (Roy 2006). Untuk kajian di salinitas rendah, perlu diketahui kebutuhan protein pada vaname karena berpotensi mempengaruhi kemampuan udang untuk osmoregulasi pada kondisi demikian. Asam amino bebas yang terkandung dalam protein memainkan peran penting dalam osmoregulasi dan dapat merupakan sumber energi umum yang dibutuhkan untuk mempertahankan pompa ion bagi pertukaran ion dan ongkos energi lainnya terkait dengan osmoregulasi (Davis et al. 2002).
Lemak pakan merupakan sumber energi dan sumber asam lemak esensial. Ikan membutuhkan lemak untuk kebutuhan energi, struktur sel, dan ketahanan integritas biomembran. Lemak dalam bentuk trigliserida akan dihidrolisis secara lengkap dalam saluran pencernaan menjadi gliserol dan asam lemak. Ada dua macam asam lemak yaitu asam lemak non esensial dan asam lemak esensial yang harus diperoleh dari luar tubuh (Furuichi 1988).
peranan penting untuk kegiatan metabolisme, komponen membran (fosfolipid dan kolesterol), sebagai senyawa awal prostaglandin, tromboksan, prostasiklin, dan leukotrin (BNF 1992), serta berfungsi dalam pengangkutan nutrien bukan lemak terutama vitamin A, D, E, dan K (Furuichi 1988). Beberapa asam lemak yang esensial untuk krustasea adalah 18μ2ω-6 (asam linoleat), 18μ3ω-3 (asam linolenat), 20μ5ω-3 (asam eikosapentaenoat, EPA), dan 22μ6ω-3 (asam dokosahexaenoat, DHA) (Gonzalez-Felix et al. 2003).
EPA dan DHA bermanfaat dalam mendukung sintasan dan pertumbuhan pada berbagai jenis krustasea. Hasil penelitian Gonzalez-Felix et al. (2002), diacu dalam Gonzalez-Felix et al. (2003) menunjukkan bahwa n-3 HUFA (High Unsaturated Fatty Acid) seperti EPA dan DHA lebih penting daripada PUFA
(Poly Unsaturated Fatty Acid) untuk pertumbuhan udang vaname (L. vannamei). Selanjutnya, Wahyudin (2005) mencatat bahwa terjadi peningkatan sintasan udang vaname sekitar 70,8-84,2% dengan pemberian rotifer yang diperkaya sumber lemak minyak ikan, minyak cumi, dan A1 DHA Selco, sedangkan dengan perlakuan rotifer tanpa pengkayaan menunjukkan sintasan lebih rendah (45,5%). Peran HUFA juga telah diuji pada udang windu (P. monodon). Kontara et al. (1995), diacu dalam Lavens dan Sorgeloos (2000) melaporkan bahwa pengkayaan Artemia dengan HUFA konsentrasi tinggi ternyata mampu meningkatkan sintasan
hingga 87% saat dilakukan penurunan mendadak salinitas media. Hasil yang sama dilaporkan Rees et al. (1994), diacu dalam Lavens dan Sorgeloos (2000) bahwa terjadi penurunan mortalitas udang windu sekitar 10% ketika pakan alami Artemia diperkaya HUFA dosis tinggi, 31 mg/g (rasio DHA/EPA: 0,5).
Kebutuhan kalsium dan fosfor dalam pakan adalah penting sehubungan dengan peran dalam struktur tulang dan metabolisme. Bentuk kristalin utama kalsium dan fosfor dalam tulang adalah hydroxyapatite, Ca10(PO4)6(OH)2. Pada
vertebrata, vitamin D (1,25 dihydroxycholecalsiferol) berfungsi sebagai hormon dalam mempertahankan konsentrasi serum kalsium dan fosfor yaitu melalui perubahan laju absorbsi pada usus (lewat ikatan Ca2+-protein), penyerapan di ginjal, dan mobilisasi dari tulang (Larvor 1983; Davis & Gatlin III 1991).
peran dalam metabolisme energi. Selain berperan dalam struktur tulang, fosfor merupakan bagian dari RNA dan DNA serta berfungsi esensial dalam fosforilasi metabolisme energi (Larvor 1983). Kecepatan pengambilan fosfor dari badan air hanya sekitar 0,001 ppm (Hilton 1989), sementara kadar ion ini di air laut sangat rendah, sekitar 0,003 ppm (Adey & Loveland 1991). Oleh karena itu, fosfor merupakan nutrien terbatas paling utama di lingkungan. Fakta ini menunjukkan kurangnya tingkat optimum rasio Ca/P dalam pakan bagi kebanyakan spesies ikan (Hilton 1989).
Interaksi kalsium dengan fosfor dalam pakan dilaporkan dapat meningkatkan pertumbuhan udang vaname. Davis et al. (1992) melaporkan bahwa pakan dengan kandungan mineral berbeda tidak mempengaruhi sintasan dan pertumbuhan juvenil vaname. Hasil penelitian Davis et al. (1993) kemudian menunjukkan bahwa keseimbangan kalsium dan fosfor diperlukan dalam pakan selama pemeliharaan juvenil di media salinitas 28 ppt. Pemberian kalsium tanpa fosfor dalam pakan tidak mempengaruhi pertumbuhan. Namun, pertumbuhan yang tinggi teramati pada perlakuan kandungan kalsium 2% dengan fosfor 1% (rasio Ca/P : 2:1) dan kalsium 1% dengan fosfor 1% (rasio Ca/P 1:1). Peningkatan kalsium 3-4% dalam pakan justru menurunkan laju pertumbuhan.
Berdasarkan penelitian di salinitas rendah oleh Cheng et al. (2006), rasio Ca/P pakan memperlihatkan respon pertumbuhan berbeda pada juvenil vaname dibandingkan di salinitas normal. Perlakuan merupakan kombinasi kalsium (0, 1, 2%) dan fosfor (0; 0,5; 1; 1,5; 2%) dalam pakan. Dari hasil penelitian tersebut, pertumbuhan, sintasan, dan mineralisasi jaringan tubuh secara nyata dipengaruhi kadar kalsium, fosfor, dan interaksi antara kalsium dan fosfor. Pemberian fosfor dalam pakan tergantung dari kadar kalsium pakan. Kesimpulan yang didapat bahwa pakan dengan kadar 1% kalsium dan 1,5 - 2% fosfor (Rasio Ca/P : 1:1,5 atau 1:2) adalah cocok untuk pertumbuhan maksimal.
Bentuk persenyawaan kalsium dan fosfor telah diuji pada ikan rainbow trout (Oncorhynchus mykiss) oleh Satoh et al.(1993). Perlakuan merupakan
fosfat: Ca3(PO4)2). Hasil penelitian menunjukkan bahwa pertumbuhan lebih tinggi
pada pakan yang mengandung dikalsium fosfat (CaHPO4) dibanding
monokalsium fosfat (Ca(H2PO4)2 dan trikalsium fosfat (Ca3(PO4)2. Proporsi dari
kalsium terhadap fosfor juga mempengaruhi penyerapan dari Zn. Perlakuan pakan dengan dikalsium fosfat (CaHPO4) mampu meningkatkan penyerapan Zn
BAHAN DAN METODE
Ruang Lingkup Penelitian
Kajian penelitian ini meliputi: (1) ketahanan hidup postlarva vaname selama tahapan aklimatisasi ke salinitas rendah, (2) perkembangan dan ketahanan hidup postlarva setelah tahapan aklimasi dengan adanya penambahan mineral pada media, serta (3) peningkatan pertumbuhan postlarva lewat pengkayaan pakan buatan selama pemeliharaan di air bersalinitas rendah.
Sebagai input yang dirancang untuk perlakuan yaitu penambahan mineral kalsium (Ca2+) selama aklimasi ke salinitas rendah berbeda dan salinitas 2 ppt pada penelitian tahap-1, penambahan mineral potasium (K+) dalam media pada penelitian tahap-2, serta pemberian protein dan kalsium pakan dalam penelitian tahap-3. Seluruh kegiatan penelitian disajikan dalam Gambar 4.
Setiap tahapan penelitian merupakan rangkaian saling berkaitan. Data yang diukur pada udang umur PL 20 hingga juvenil. Pengambilan data dilaksanakan selama kira-kira 6 minggu penelitian berlangsung. Hasil percobaan tahapan aklimasi salinitas pada penelitian tahap-1 merupakan bentuk keberhasilan dari proses adaptasi yang terukur dari nilai gradien osmotik, tingkat konsumsi oksigen, dan sintasan. Hasil penelitian awal dijadikan pedoman untuk penelitian tahap kedua yang berlanjut pada penelitian tahap ketiga.
Gambar 4 Skema urutan kegiatan penelitian.