OPTIMALISASI
SISTEM FERMENTAS1
RUMEN
MELALUX
, SUPLEMENTASISULFUR,
DEFAUNASI, REDUKSI EMISI
METAN DAN
STIMULASI PERTUMBUHAN MIKROBA
PADA
TERNAK
RUMINANSIA
PROGRAM
PASCASARJ
ANA
INSTITUT PERTANIAN
BOGOR
ERWANTO. Optimalisasi Sistem Fermentasi Rumen melalui Suplementasi Sulfur, Defaunasi, Reduksi Emisi Metan dan Stimulasi Pertumbuhan Mikroba pada Terdak Ruminansia (di bawah bimbingan Toha Sutardi (ketua), Djokowoerjo Sastradipradja, Narsum, Muhilal dan Suryahadi).
Limbah serat mem il iki kecernaan rendah, karena itu penggunaannya sebagai pakan memerlukan sentuhan teknologi. Teknologi pengolahan, misalnya amoniasi, dapat meningkatkan fermentabil itas. Peningkatan fermentabilitas harus disertai penye- diaan nutrien prekursor sintesis biomassa mikroba. Keterbatasan nutrien menurunkan pertumbuhan mikroba.
Kecernaan dalam rumen ditentukan populasi mikroba. Peningkatan populasi mi kroba dapat d idekati dari segi ekologi dan kecukupan pasokan nutriennya. Pada ternak yang mendapat pakan serat, kehadiran protozoa dalam rumen kurang mengun- tungkan, karena menekan perkembangan bakteri. Defaunasi rumen ditujukan untuk menghilangkan atau mengurangi protozoa, agar tercipta kondisi ekologi yang kondusif bagi pertumbuhan bakteri. Beberapa ha1 perlu diperhatikan pada pelaksanaan defauna- si, misalnya kecukupan pasokan nitrogen (N) dan kerangka karbon bercabang dalam rumen.
Pakan serat rendah kadar N, sehingga pemakaiannya perlu disertai penambahan N
.
Penggunaan ransum mengandung nitrogen bukan protein (NPN) akan berhasil apabi- la disertai suplementasi asam lemak rantai cabang (BCFA). BCFA dalam rumen berasal dari fermentasi protein pakan dan bakteri yang lisis. Apabila ransum rendah protein dan ada penerapan defaunasi, maka jumlah BCFA dalam rumen sangat terbatas.Pada ransum menganglung NPN, selain suplementasi karbon bercabang, perlu pula suplementasi sulfur (S). Mineral tersebut diperlukan untuk sintesis de novo asam amino mengandung S. Mikroba rumen dapat memanfaatkan S anorganik dengan baik. S terinkorporasi ke dalam sistein dalam bentuk sul fida.
Penelitian ini ditujukan untuk memadukan teknologi defaunasi, suplementasi sulfur, asam lemak tidak jenuh dan asam amino rantai cabang, sebagai upaya memacu b ioproses pakan serat dalam rumen dan meningkatkan pertumbuhan ternak.
Percobaan dilaksanakan di Fakultas Peternakan IPB selama 4 bulan. Ternak yang digunakan 15 ekor sapi perah jantan, yang dibagi menjadi
3
kelompok berdasar- kan bobot tubuhnya. Bobot tubuh pada awal penelitian 215 f 26 kg. Percobaan pemberian makanan selama 10 minggu, sedangkan percobaan mikrobiologi rumen selama 10 hari. Suplementasi sulfur diberikan berupa amonium sulfat. Defaunasi rumen menggunakan minyak kelapa. Sumber asam lemak tidak jenuh untuk mengalih- kan hidrogen dalam rangka menekan metan digunakan minyak ikan. Kerangka karbon bercabang diberikan berupa asam amino, yaitu L-Valin dan L-Leusin.Semun bahan suplementasi dicampurkan merata ke dalam ransum. Ransum berbahan dasar rumput raja yang diamoniasi dengan urea. Ransum basal (Rl) disusun dengan kadar protein kasar 14 % dan kandungan energi metabolis (ME) 10.5 MJ per kg bahan kering. Ada 5 macam ransum perlakuan yang dicobakan, dan masing-masing perlakuan terdiri atas 3 ulangan, yaitu: R1 = Ransum basal, R2 = R1
+
Amonium sulfat (sumber sulfur), R3 = R2+
Minyak kelapa (agensia defaunasi), R4 = R3+
Minyak ikan (sumber asam lemak tidak jenuh), dan R5 = R4+
Asam amino berca- bang (L-Val in dan L-Leusin).Percobaan disusun dalam rancangan acak kelompok (RAK) dengan 3 ulangan. Ransum diberikan 2 kali sehari dalam bentuk pelet, sedangkan air minum disediakan sepanjang hari. Peubah yang diamati adalah: (1) pertambahan bobot tubuh, (2) kompo- sisi tubuh (3) kecernaan zat-zat makanan, (4) populasi bakteri dan protozoa, (5) kadar VFA individcal cairan rumen, (6) pH dan konsentrasi amonia cairan rumen, (7) kadar asam-asam amino plasma darqh, dan (8) kadar allantoin dalam urin.
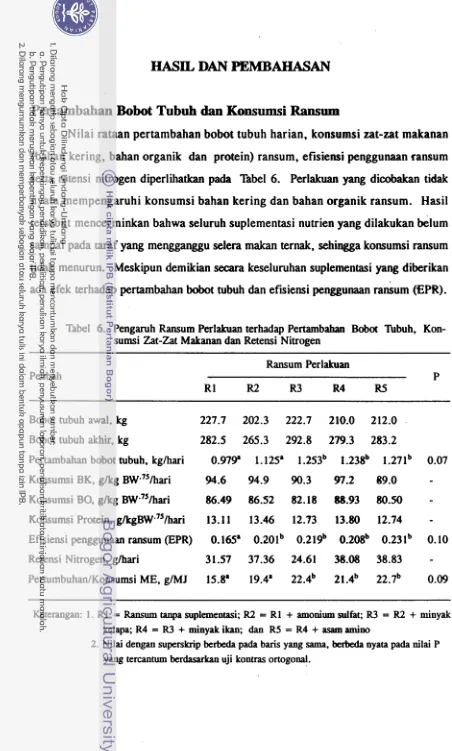
Pertambahan Bobot Tubuh dan Konsumsi Ransum
Penambahan sulfur memperbaiki pertambahan bobot tubuh dan berpengaruh positif pada pencernaan fermentatif di rumen. Penambahan sulfur memacu pertumbu- han mikroba, sehingga meningkatkan produk fermentasi rumen (VFA) dan pasokan nutrien untuk ternak induk semang. Hal tersebut dapat dilihat pada peningkatan popu- lasi b'akteri dnn p r o t o m rumen.
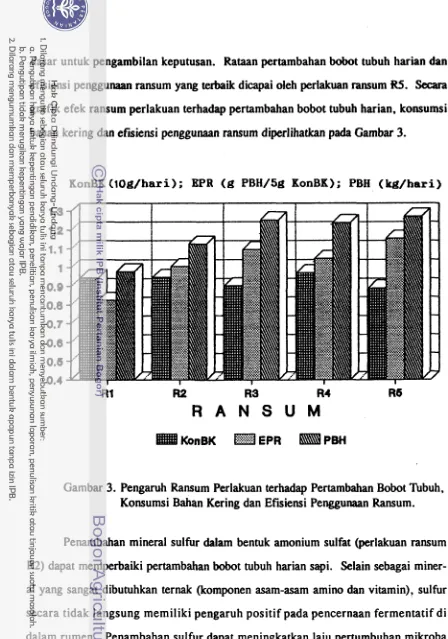
Pertambahan bobot tubuh dapat ditingkatkan dengan defaunasi menggunakan minyak kelapa. Penambahan minyak kelapa menurunkan populasi protozoa sebesar
23 %. Pada perlakuan R3 turunnya populasi protozoa diiringi peningkatan populasi bakteri.. Peningkatan populasi bakteri berpengaruh baik terhadap kecernaan, terutama fraksi serat. Kecernaan serat deterjen asam meningkat pada perlakuan R3. lmplikasi dari semua perbaikan tersebut adalah pertambahan bobot tubuh meningkat.
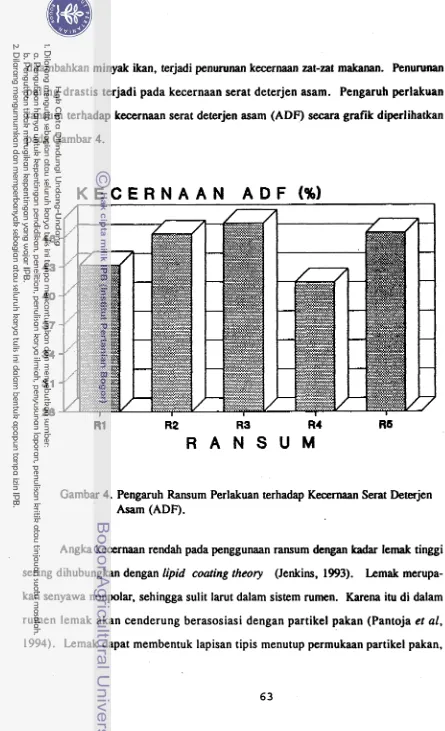
Penambahan minyak ikan (akseptor hidrogen dalam rangka menekan metan) pada perlakuan R4, tidak memberikan perubahan pada pertumbuhan, meskipun berda- sarkan profil VFA individual dan data produksi metan terlihat usaha menekan produk- si metan berhasil. Kejadian tersebut diduga akibat tingginya kadar lemak pada perla- kuan R4. Pada perlakuan tersebut selain suplementasi minyak kelapa juga ada suple- mentasi m in::ak ikan. Kadar lemak yang tinggi dapat menekan kecernaan ransum. Penurunan kecernaan yang terjadi pada perlakuan R4 terlihat jelas pada kecernaan
ADF. Gangguan kecernaan menyebabkan pasokan nutrien, sehingga tidak mendukung peningkatan produksi.
Penambahan asam amino bercabang memperbaiki pertalnbahan bobot tubuh. Seperti halnya perlakuan R4, pada perlakuan
R5
sebenarnya juga ada gangguan kecer- naan oleh lemak. Tetapi gangguan tersebut dapat dikonvensasi oleh manfaat positif suplementasi asam amino bercabang. Pada perlakuanR5
juga ada suplementasi sulfur dan defaunasi. Defaunasi rnenyebabkan berkurangnya bakteri yang lisis (pemasok karbon bercabang). Karena itu pada ternak didefaunasi karbon bercabang mungkin sekali menjadi faktor pembatas pertumbuhan mikroba, sehingga suplementasi nutrien tersebut dalam ransum sangat bermanfaat.Kecernaan Zat-Zat Makanan
pertumbuhan mikroba rumen menyebabkan pertumbuhan ternak membaik.
Kecernaan protein meningkat setelah suplementasi sulfur. Perbaikan kecernaan protein tersebut mengindikasikan terjadinya peningkatan efisiensi penggunaan
N
ran- sum. Data retensi N memperkuat indikasi tersebut, bahwa sulfur dapat meningkatkan retensi N. Penambahan sulfur dapat memacu pertumbuhan mikroba rumen, sehingga meningkatkan laju asimilasi amonia (N).Pada perlakuan R4 cenderung terjadi penurunan kecernaan. Penurunan paling drastis terjadi pada kecernaan ADF. Kecernaan rendah pada ransum kadar lemak tinggi sering dihubungkan dengan lipid coating theory. Sebagai senyawa nonpolar lemak sul it larut da! am sistem rumen, sehingga lemak cenderung berasosiasi dengan partikel pakan. Lemak membentuk lapisan tipis menutup permukaan partikel pakan, sehingga menghalangi kontak langsung antara mikroba serta enzimnya dengan partikel pakan.
Penambahan lemak berpengaruh positif terhadap kecernaan lemak ransum. Perlakuan R4 dan R5, yang mendapat tambahan minyak kelapa dan minyak ikan, memiliki kecernaan lemak lebih tinggi dibandingkan ransum lainnya. Minyak yang ditambahkan menjadi komponen lemak ransum di luar struktur partikel pakan, sehing- ga langsung terdedah kepada mikroba rumen dan enzim-enzim pengurai lemak.
Pada perlakuan R5 populasi bakteri tertinggi. Populasi bakteri yang tinggi tersebut berimplikasi pada meningkatnya kecernaan ADF yang semula turun drastis (pada perlakuan R4). Hasil tersebut menambah bukti bahwa bakteri yang sangat re- spansif terhadap asam amino bercabang adalah bakteri selulolitik. Data pengeluaran alantoin urin juga mencerminkan bahwa pada perlakuan R5 sumbangan protein asal mikroba rumen untuk ternak tertinggi (P
<
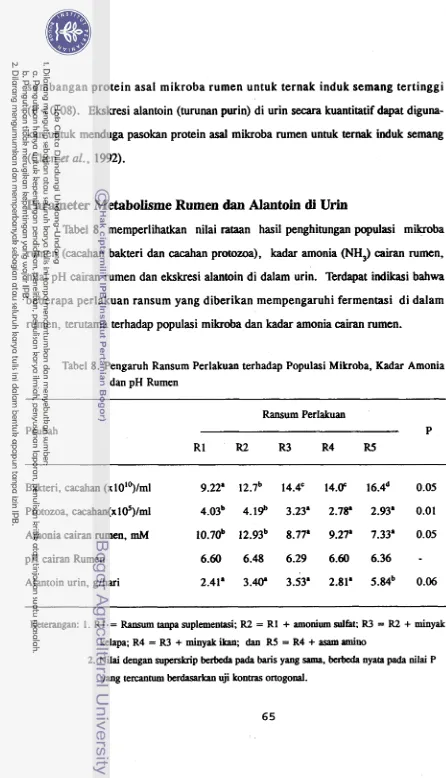
0.06).Parameter Metabolismq Rumen dan Alantoin di Urin
Perlakuan mempengarulii fermentasi dalam rumen, terutama terhadap populasi mikroba dan kadar amonia rumen. Ransum penelitian menggunakan pakan serat diamo- niasi dengan urea. Tampak bahwa semua ransum perlakuan dapat dikatakan mampu menyediakan amonia rumen dalam kadar yang cukup untuk pertumbuhan mikroba. Hasil tersebut membuktikan bahwa amoniasi selain meningkatkan degradasi pakan, juga dapat menjaniin kecukupan pasokan amonia pada kondisi in vivo.
(tahap inisiasi) sintesis protein dalam
sel
mikroba. Karena fungsi metionin yang strate- gis tersebut, maka penambahan sulfur meningkatkan pertumbuhan bakteri.Minyak kelapa sebagai agensia defaunasi berpengaruh positif terhadap populasi bakteri. Pada perlakuan R3 populasi protozoa lebih rendah daripada perlakuan R2.
Terhdmbatnya pertumbuhan protozoa memungkinkan kondisi ekologi yang baik untuk pertumbuhan bakteri. Di dalam rumen protozoa dikenal sebagai predator bakteri.
Asam amino bercabang memacu pertumbuhan mikroba. Pada perlakuan R5 populasi bakteri lebih tinggi daripada perlakuan R4. Pada perlakuan
R5
ada defaunasi, yang dapat menurunkan pasokan asam lemak bercabang. Menurunnya pasokan karbon bercabang menyebabkan metabolit tersebut menjadi faktor pembatas pertumbuhan mikroba dan rumen menjadi sangat tanggap terhadap suplementasi karbon bercabang. Peningkatan pertumbuhan bakteri tercermin pula dari data alantoin urin. Perlakuan R5 yang mendapat suplementasi lengkap memberikan angka ekskresi alantoin tertingg i.
Minyak kelapa sebagai agensia defaunasi menurunkan populasi protozoa. Beberapa teori menjelaskan mekanisme efek anti protozoa tersebu t, tetapi teori physical couting lebih mantap. Asam lemak cenderung membalut sel mikroba rumen. Protozoa tidak memiliki aktivitas lipolitik sebaik bakteri. Akibatnya apabila dalam rumen banyak asam lemak maka protozoa sulit bertahan hidup, sehingga populasinya berkurang.
Penurunan populasi protozoa diikuti penurunan kadar amonia rumen. Fenomena ini mengingatkan peranan protozoa dalam daur ulang N. Protozoa berperan dalam daur ulang N dengan mencerna N bakteri. Protein bakteri yang dimangsa protozoa sebagian dikeluarkan kembali ke dalam rumen berupa peptida, asam amino dan amonia. Saat defaunasi peranan protozoa tidak berjalan dan daur ulang N dalam rumen terganggu.
Pena~nbahan minyak ikan meningkatkan asam propionat, menurunkan angka
NGR,
nisball AIP dan produksi metan dalam rumen. Dari segi efisiensi penggunaan energi ransum, fermentasi rumen yang mengarah kepada sintesis propionat lebi h menguntungkan, karena energi yang terbuang sebagai metan berkurang. NGR diperli- hatkan berkorelasi positif dengan produksi metan, sehingga turunnya NGR dengan masuknya minyak ikan ikut mencerminkan adanya penurunan produksi metan.Pada perlakuan
R4
konsentrasi isoacids sedikit menurun. Kecenderungan menurun mungkin disebabkan menurunnya pasokan asam amino bercabang yang berasal dari bakteri yang lisis. Data populasi protozoa mendukung dugaan tersebut. PopuIasi protozoa pada perlakuan R4adalah
terendah, sedangkan protozoa berperan ddam daur ulani isoacids dalam rumen.Penambahan asam amino bercabang
,
meskipun tidak nyata, meningkatkan kadar isoacids dalam rumen. Peningkatan pasokan kerangka karbon bercabang tersebut meningkatkrn populasi bakteri. Hasil ini mencerminkan bahwa kerangka karbon bercab'ang pada perlakuanR4
mulai menjadi faktor pembatas, dan suplementasinya pada perlakuanR5
dapat mengoreksi kekurangan tersebut.Asam
Amino
Plasma Darah clan
Komposisi
Tubuh
Perlakuan tidak banyak menimbulkan perbedaan berarti pada kadar asam amino plasma. -Suplementasi sulfur sedikit menaikkan konsentrasi metionin. lhjuan suplemen- tasi sulfur adalah mendukung sintesis de
mvo
asam amino mengandung sulfur. Sulfur tereduksi bersama 0-asetilserin membentuk sis tein, yang selanjutnya akan menjadi met ionin melalui proses transul furisasi.Pertumbuhan tinggi pada perlakuan
R5
tentunya diikuti pasokan asam amino dalam laju yang tinggi dari aliran darah ke dalam sel. Dengan kata lain pada perlakuan R5 nilai "fluksw asam amino darah lebih tinggi dibandingkan dengan perlakuan lainnya. Berarti meskipun kadar asam amino mengandung sulfur pada perlakuanR1
tidak berbeda nyata dengan empat ransum lainnya, dapat terjadi ada perbeciaan dalam angka fluks asam aminonya. Fenomena yang sama dapat pula terjadi pada kadar asam amino bercabang dalam darah antara sapi pada perlakuan R5 dengan empat perlakuan lainnya.Komposisi tubuh re ktif tidak berbeda antar perlakuan. Perbedaan yang nyata hanya pada kadar mineral tubuh. Setelah suplementasi sulfur kadar mineral tubuh meningkat. Hasif tersebut mencerminkan bahwa suplementasi sulfur pada ransum yang defisien sulfur, dapat meningkatkan retensi mineral dalam tubuh terna..
OPTIMALISAS1 SISTEM FERMENTMI
RUMEN MELALUI
SUPLEMENTASI
SULFUR,
DEFAUNASI, REDUKSI EMISI
.
METAN DAN S-AS1
PEKI'UMBUHAN MIKROBA
PADA
TERNAK
RUMINANSIA
oleh
ERWANTO,
NRP.
B510
Disertasi sebagai salah satu syarat untuk
memperoleh gelar Doktor pada Program Pascasarjana
h t i t u t
Pertanian Bogor
PROGRAM PASCASUANA
INSTITUT PERTANIAN BOGOR
Judul Penelitian : Optimalisasi Sistem Fermentasi Rumen melalui Suplementasi Sulfur, Defaunasi, Reduksi Emisi Metan dan Stimulasi Pertum- buhan Mikroba psda Ternak Ruminansia
Nama Mahasiswa : ERWANTO
Nomor Pokok :
92510/PTK
Menyetujui
1. Komisi Pembimbing,
/ Prof. Dr. Toha Sutardi, M.Sc.
Ketua
C-
Prof. Dr. D. Sastradi~radia Prof. Dr. Narsum
Anggota Anggota
Dr.
Ir. H. Survahadi. DEA Anggota ,2. Ketua Program Studi Ilmu Ternak,
Dr. H. Muhilal Anggota
Penulis dilahiriran di Pau8ragan (Imnpung Utara) pada t a w 25 P e M
1961, merupakan putra pertama dari tujuh bersaudam, d e w ibu
bernama
Nllwirohdan ayah Abdullah Sani. Pendidikan Sekolah Dasar diselesaikan tdm 1973 dari
SD
~ e ~ e r i Pansragao dan Pendidikan Sekolah Menengab Pertama diselesaikm
tahun
1976dsri
SMP Negeri Mensgala Pendidikan Sekolah Menen& Atas diselesaikan tahun1980 pada SMA N e ~ e r i Kotabumi.
-
Tahm 1980 permlis diterima sebagai mahasiswa Institut Pertanian Bogor lewatjalur Proyek Perintis I1 dan lulua sebagai Sarjana Peternakan tahrm 1984. Pendidikan
Magistet- Sains (M.S.) dalam bidang Ilmu Ternak diselesaikan tahun 1992 pada
Program Pascasarjana Iastitut Pertanian Bogor. Tahun 1992 penulis 1
-melanjutkan Pendidikan Doktor pada Program Pascasarjana Institut Pertanian Bogor,
dengan biaya dari Tim Manajemen Program Doktor Direktorat Jenderal Pendidikan
Tiaggi. Sejak tahun 1985 penulis menjadi Staf Pengajar pada Program Studi Produksi
Ternak Fakultas Pertanian Universitas Laqung.
Penulis menikah dengan Ir. Mucharomah Prayu Widayati. Sampai saat ini
KATA PENGANTAR
Disertasi berjudul Optimaliiasi Sistem Fermentasi Rumen melalui Suplementasi Sulfur, Defeunasi, Reduksi Emisi Metan dan Stimulasi Pertumbuhan Mikroba pada Ternak Ruminansia, disusun sebagai salah satu syarat untuk memperoleh gelar ~ o k r pada Program Pascasarjana Institut Pertanian Bogor. Penelitian dil-n di Laboratorium Ruminologi dan Mikrobiologi Nutrisi Fakultas Peternakan IPB.
Penelitian dan penulisan disertasi ini dapat terselesaikan atas pengarahan serta bimbingan dari Tim Komisi Pembimbing. Penulis menyampaikan terimakasih kepada prof.* Dr. Toha Sutardi sebagai ketua komisi, Prof. Dr. Djokowoerjo Sastradipradja, Prof. Dr. Narsum, Dr. H. Muhilal dan Dr. IT.
H.
Suryahadi, DEA atas pengarahan, bimbingan dan segala bentuk bantuan yang sangat berharga.Ucapan terima kasih disampaikan kepada Rektor IPB dan Pimpinan Program Pascasarjana IPB atas kesempatan mengikuti studi program Doktor. Kepada Rektor Universitas Lampung dan Dekan Fakultas Pertanian Unila disampaikan terima kasih
atas ijin melcnjutkan studi Doktor. Ucapan terimakasih juga disampaikan kepada Tim Manajemen Program Doktor Di tjen Dikti yang membiayai studi pascasarjana penul is di IPB. Kepada Ibu Yayah Koswara dari Dikti penulis menyampaikan ucapan terima kasih, karena melalui dukungan dana Proyek Hibah Bersaing 113 (a.n. Prof. Dr. Toha Sutardi) di bawah program beliau penelitian ini dapat dilaksanakan.
Kepada Gubernur Lampung Bapak Poedjono Pranyoto, Harian Kompas, Yaya- san Toyota Astra dan PT Indocement Tunggal Prakarsa disampaikan banyak terima kasih atas bantuan yang diberikan. Bantuan tersebut sangat membantu proses penye- lesaian studi penulis tepat pada waktunya.
Kepada isteri tercinta lr. Mucharomah Prayu Widayati, ananda tercinta Andre Purnama Alam, orang tua dan mertua tercinta, penulis menyampaikan terima kasih dan penghargaan atas pengertian dan doa restunya. Terima kasih juga disampaikan kepada Sdr. Despal, Siti Nurbaya, Jalaludin, Iyan dan Eko semua pihak yang telah memberikan berbagai bentuk bantuan sehingga penelitian dan penulisan disertasi ini dapat terwujud. Akhirnya, semoga hasil penelitian ini dapat bermanfaat untuk kesejahteraan manusia.
Bogor, April 1995
Pe~lulis
KATA PENGANTAR
DAFTAR IS1
DAFTAR TABEL
DAFTAR GAMBAR
PEN~AHULUAN Latar Belakang Tujuan Penelitian Kegunaan Penelitian
TINJAUAN PUSTAKA
Sistem Pencernaan pada Ternak Ruminansia
Pasokan Zat-Zat Makanan untuk Ternak Ruminansia Metabolisme Karbohidrat
Metabolisme Protein
Peranan Sulfur pada Nutrisi Ternak Ruminansia
Protozoa dan Penggunaan Lemak sebagai Agensia Defaunasi Peranan Isoacids dalam Pertumbuhan Mikroba Rumen
MATERI DAN METODE
Prosedur Pendugaan Komposisi Tubuh dengan Ruang Urea Pengukuran Kecernaan Zat-zat Makanan dan Retensi Nitrogen Prosedur Pencacahan Populasi Protozoa Rumen
Prosedur Pencacahan Populasi Bakteri Rumen Prosedur Anal isis VFA Individual
Prosedur Pengukuran Konsentrasi N-Amonia Prosedur Analisis Asam Amino Plasma Darah Prosedur Analisis Alantoin Urin
Analisis Data
HASIL DAN PEMBAHASAN
Pertambahan Bobot Tubuh dan Konsumsi Ransum Kecernaan Zat-Zat Makanan
Parameter Metabolisme Rumen dan Alantoin Urin Asam Amino Plasma dan Komposisi Tubuh
Tinjauan Komprehensif
iii
KESIMPULAN
DAFTAR PUSTAKA
Nomor JuduI 'ItrbeI .
Halaman
1. Komposisi Asam-Asam Amino Bakteri Rumen 14
2. Ketersediaan Relatif Berbagai Sumber Sulfur diukur Berdasarkan
Sintesis Protein In Vitro 30
3. Pengaruh Defaunasi Rumen terhadap Pertumbuhan Ternak,
Konsumsi dan Parameter Rumen 35
4. Pengaruh Suplementasi Asarn Lemak Bercabang terhadap Kecer-
naan Dinding Sel dan Konsentrasi Amonia 43
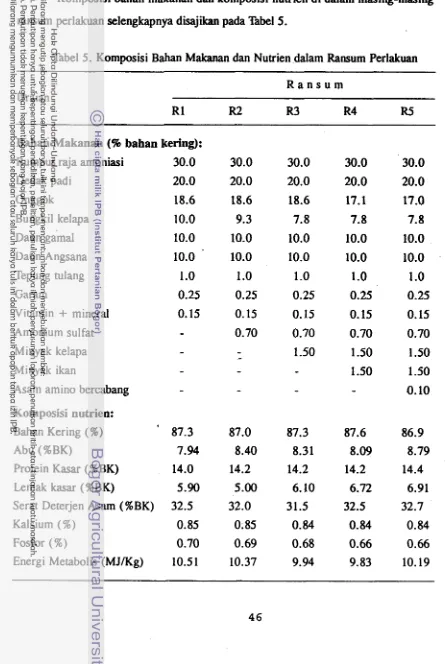
5. 'Komposisi Bahan Makanan dan Nutrien dalam Ransum Perlakuan 46
6. Pengaruh Perlakuan Ransum terhadap Pertambahan Bobot
'Ibbuh, Konsumsi BK, BO, PK dan Retensi N 56
7. Pengaruh Ransum Perlakuan terhadap Kecernaan Zat-zat Makanan 61
8. Pengaruh Perlakuan Ransum terhadap Populasi Mikroba, Kadar
Amonia dan pH Rumen 65
9. Pengaruh Perlakuan Ransum terhadap Konsentrasi Asam Lemak
Terbang Individual Cairan Rumen 72
10. Pengaruh Perlakuan Ransum terhadap Komposisi Asam Amino
Plasma Darah 79
Nomor Judul Gambar Halaman
1. Skema Fermentasi Karbohidrat dalam Rumen 20
2. Ikhtisar Degradasi Protein di dalam Rumen 25
3. Pengaruh Ransum Perlakuan terhadap Bobot lbbuh, Konsumsi
dan Efisiensi Penggunaan Ransum 58
4. Pengaruh Ransum Perlakuan terhadap Kecernaan Serat Deterjen
Asam 63
5. Pengaruh Ransum Perlakuan terhadap Ekskresi Alantoin di Urin 69
6 . Pengaruh Ransum Perlakuan terhadap Populasi Bakteri,
Protozoa dan Kadar Amonia Rumen 71
7. Pengaruh Ransum Perlakuan terhadap Estimat Produksi Gas
Metan 74
8. Pengaruh Ransum Perlakuan terhadap Propionat, Nisbah AIP
dan Angka
NGR
759. Pengaruh Ransum Perlakuan Terhadap Efis iensi Konversi
- Energi Heksosa menjadi VFA 75
Nomor Judul Lampiran
1. Data, Hasil Analisis Ragam
dan
Uji Kontras Ortogonal untuk Pertarnbahan Bobot Tubuh (kglhari)2. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk
.
Efisiensi Penggunaan Ransum (kg bobot tubuhkg rallsum)3. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal u~ituk Konsumsi Bahan Kering (gtkg bobot metaboliklhari)
4. Data, Hasil Analisis Ragam
dan
Uji Kontras Ortogonal untuk Konsumsi Bahan Organik (gkg bobot metabolikfhari)5 . Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kecernaan Bahan Kering (%)
6. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kecernaan Bahan Organik (%)
7. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kecernaan Protein
(96)
8. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kecernaan Lemak (%)
9. Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Serat Deterjen Asam (%)
10 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk
Kecernaan Energi ( %)
11 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Populasi Bakteri Rumen (skala logaritma)
12 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk
Populasi Protozoa Rumen (skala logaritma)
13 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk
Konsct~trasi Amonia Cairan Rumen (mM)
14 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk
pH Cairan Rumen
Halaman
95
Data, Hasil Analisis Ragam
dan
Uji Kontras Ortogonal untuk 102Ekskresi Alantoin Urin (glhari)
Data, Hasil Analisis Ragam
dan
Uji Kontras Cktogonal untuk 102Retensi Nitrogen (gfhari)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 103 Konsentrasi Asetat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 103
Konsentrasi Propionat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 104 Konsentrasi Butirat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 104 Konsentrasi Isobutirat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 105
Konsentrasi Valerat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 105 Konsentrasi Isovalerat Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 106 Konsentrasi Isoacids Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 106
Konsentrasi Total VFA Cairan Rumen (mM)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 107 Nisbah AsetatIPropionat Cairan Rumen
,
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonai untuk 107 Angka NGR Cairan Rumen
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 108 Konsentrasi Metionin Plasma ~rno11100ml)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 108 Sistein Plasma (pmo1/100ml)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk 109 Konsentrasi Isoleusin Plasma (pmo11100mI)
31 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Konsentrasi Valin Plasma (pmo1/100ml)
32 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Konsentrasi Asam Amino Sulfur (pmo1/100mI)i)
1
33 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kadar Asam Amino Rantai Cabang (AARC)
34 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kadar Air Tubuh Kosong (%)
35 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kadar Lemak Tubuh (%)
36 Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kadar Protein Tubuh ( %)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Kadar Mineral Tubuh (%)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Produksi
Gas
Metan (mMl100mM VFA)Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Efisiensi Konversi Energi Heksosa Menjadi VFA (%)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Pertumbuhan/Konsumsi ME
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Deposisi Protein (glhari)
1
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Energi Teretensi (MJthari)
Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Deposisi
Lemak
(glhari)Data, Hasil Analisis Ragam dan Uji Kontras Ortogonal untuk Konsumsi Protein (glkg BW."Ihari)
Data dan Rataan Bobot Badan Awal Sapi Percobaan
46 Data dan Rataan Bobot Badan Akhir Sapi Percobaan
Latar Belakang
4
Untuk mampu berproduksi sesuai dengan potensi genetiknya, ternak unggul
hasil pemuliaan dan bioteknologi memerlukan pakan berkualitas
baik.
Limbah seratmerupakan sumberdaya yang tersedia dalam jumlah besar dan murah di daerah
tropis. Untuk menekan biaya ransum, manfaat limbah serat sebagai pakan utama
ternak ruminansia perlu terus ditingkatkan. Selarna ini limbah serat dikenal sebagai
pakan berkualitas rendah. Karakteristik utama kelompok pakan tersebut adalah
tinggi kadar serat (fiber), sehingga memiliki nilai kecernaan
yang
rendah. Dengandemikian untuk penggunaannya sebagai pakan dalam jumlah besar, memerlukan
sentuhan teknologi.
Upaya-upaya yang telah dilakukan selama ini masih terpusat pada teknik-
teknik pengolahan pakan, misalnya pengolahan secara fisik, kimia dan biologis,
untuk meningkatkan kecernaan bahan tersebut di dalam rumen. Berbagai teknolog i
pengolahan pakan, misalnya perlakuan amoniasi dengan urea, perlakuan alkali dan
fermentasi mikologis telah berhasil meningkatkan fermentabilitas pakan serat di
dalam rumen. Tetapi berbagai teknolog i pengolahan tersebut masih memerlukan
kajian lebih lanjut untuk dapat diaplikasikan secara komersial. Teknologi tersebut
harus segera dipadukan dengan usaha memacu fermentasi dalam rumen.
Untuk memaksimumkan pasokan nutrien kepada ternak, keberhasilan men-
ingkatkan fermentabilitas pakan di dalam rumen harus diikuti dengan upaya penye-
diaan semua prekursor untuk sintesis biomassa mikroba rumen. Biomassa mikroba
rumen telah diketahui merupakan sumber protein berkualitas bagi ternak induk
semang. Keterbatasan pasokan salah satu nutrien akan menurunkan laju pertumbu-
han mikroba. Laju sintesis protein mikroba rumen sangat ditentukan oleh keterse-
Pada ternak ruminansia kecernaan pakan serat sangat ditentukan d e h popula-
si mikroba rumen. Proses perombakan pakan serat pada dasarnya adalah kerja
enzimenzim yang diproduksi oleh mikroba rumen. Keberhasilan usaha peningka-
tan populasi mikroba rumen akan meningkatkan konsentrasi enzim-enzim tersebut,
sehingga diharapkan dapat meningkatkan kecernaan pakan. Bagi ternak induk
semang, peningkatan populasi mikroba rumen tersebut selain akan meningkatkan
kecerriaan pakan, juga akan meningkatkan pasokan protein asal mikroba rumen yang
telah diketahui berkualitas tinggi.
Peningkatan populasi mikroba rumen pencerna serat dapat didekati dari segi
ekologi dan dari segi kecukupan pasokan nutrien (prekursor) untuk pertumbuhan
mikroba tersebut. Pada ransum dengan bahan utama pakan serat berkualitas rendah,
kehadiran protozoa di dalam rumen kurang menguntungkan, karena cenderung
menekan perkembangan populasi bakteri rumen. Pada kondisi ini hubungan yang
positif dalam interrelasi antara bakteri-protozoa tidak menonjol, bahkan kehadiran
protozoa akan banyak memboroskan energi untuk pemenuhan kebutuhan hidup
pokoknya.
Teknologi defaunasi yang dapat mengendalikan populasi protozoa pada situasi
seperti tersebut di atas diharapkan akan memberikan kondisi ekologi yang kondusif
bag i pertumbuhan bakteri rumen. Beberapa penel itian telah memperlihatkan indikasi
I
bahwa defaunasi dapat meningkatkan pertumbuhan bakteri rumen. Meskipun demi-
kian ada beberapa ha1 yang perlu diperhatikan pada pelaksanaan defaunasi, misalnya
kecukupan pasokan nitrogen dan kerangka karbon bercabang di dalam rumen.
Protozoa mempunyai andil yang berarti pada daur ulang nitrogen (nitrogen recy-
cling) dan penyediaan kerangka karbon bercabang di dalam rumen.
Pakan serat sangat rendah kandungan nitrogennya, sehingga penggunaannya
nitrogen untuk mendukung pertumbuhan mikroba rumen. Sumber nitrogen yang
murah dan dapat digunakan adalah nitrogen bukan protein (Non-Protein Nitrogen
= NPN). Mikroba rumen telah terbukti dapat menggunakan amonia
asal
urea (atausumber NPN-lainnya) dengan sangat baik. Sekitar 80% dari jenis mikroba rumen
diketahui dapat menggunakan amonia sebagai sumber nitrogen tunggal
(Bryant
danRobinson, 1963).
Penggunaan ransum yang mengandung NPN dalarn jumlah besar, akan lebih
berhasil apabila disertai dengan suplementasi asam lemak rantai cabang (Branched
Chain Fatty Acids = BCFA). Asam-asam lemak tersebut sangat diperlukan teruta- ma untuk pertumbuhan kelompok bakteri selulolitik. Asam-asam lemak rantai
cabang dalam rumen berasal dari hasil fermentasi protein pakan dan dari protein
bakteri yang mengalami lisis dalam rumen. Dengan dernikian apabila ransum rendah
kadar proteinnya, tentu saja jumlah asam lemak rantai cabang di dalam rumen akan
sangat terbatas. Keterbatasan asam lemak rantai cabang akan semakin kritis apabila
kondisi tersebut disertai dengan penerapan defaunasi dalam rumen. Defaunasi
menyebabkan aliran protein bakteri ke organ pascarumen meningkat dan bakteri
yang mengalami lisis oleh protozoa cenderung berkurang. Karena itu penerapan
defaunasi selayaknya juga memperhatikan aspek kecukupan pasokan kerangka
karbon bercabang.
Pada ransum yang mengandung NPN dalam jumlah besar, selain suplementa-
si kerangka karbon bercabang, perlu pula diikuti dengan suplementasi mineral sulfur
(S).
Tanpa suplementasi, maka mineral sulfur akan menjadi faktor pembatas per-tumbuhan mikroba rumen. Mineral tersebut sangat diperlukan untuk sintesis de
novo asam-asam amino mengandung sulfur (Sulphur Containing Amino Acids =
SCAA). Mikroba rumen telah dibuktikan dapat memanfaatkan sulfur anorganik
sul fida (sulfur tereduksi). Mikroba rumen, terutama fungi sangat responsif terhadap
penambahan sulfur dalam ransum. Akhir-akhir ini telah diketahui bahwa fungi
rumen ternyata memiliki peranan sangat berarti pada pencernaan pakan serat.
Karkna
itu suplementasi sulfur dapat diharapkan berdampak positip terhadap kecer- naan ransum yang banyak mengandung pakan serat.Pada ternak ruminansia sebagian energi pakan ada yang terbuang dalam
bentuk produksi gas metan (CHJ. Gas metan terbentuk dari reaksi antara gas CO,
dan gas H,. Proses tersebut terjadi untuk menyalurkan akumulasi hidrogen di dalam
rumen, sehingga potensial redoks dalam rumen tetap dapat dipertahankan pada
kisaran yang ideal. Potensial redoks dalam rumen sekitar -250 sampai
-
450 mV,yang mencerminkan kondisi tanpa oksigen dan daya reduksi tinggi (Van Soest,
1982). Pemborosan energi pakan yang muncul dalam bentuk produksi gas metan
ternyata cukup b a r , umumnya berkisar 12.4 %
.
Akhir-akhir ini muncul isu pemanasan global (global warming), yang men-
ganggap bahwa emisi gas metan merupakan salah satu faktor penyebab yang perlu
segera dikendalikan. Dengan demikian perlu segera diupayakan metode manipulasi
proses nutrisi yang mengarah kepada penurunan produksi gas metan di dalam rumen.
Hal tersebut dapat dicapai misalnya dengan mengarahkan fermentasi menuju sintesis
asam propionat, atau dengan mengalihkan akumulasi hidrogen dalam rumen.
I
Pemberian asam lemak tidak jenuh (polyunsaturated fatty acids = PUFA) ke dalam
ransum di harapkan dapat mengalihkan penggunaan gas hidrogen untuk menjenuhi
asam lemak tak jenuh (biohidrogenasi). Dengan demikian produksi gas metan diha-
rapkan dapat ditekan.
Optimasi bioproses di dalam rumen, menuntut penciptaan kondisi ekologi
sistem fermentasi yang mendukung dan kecukupan pasokan nutrien untuk pertumbu-
bioproses yang melibatkan secara bersamaan teknologi defaunasi (segi ekologi),
suplementasi sulfur, kerangka karbon bercabang serta penggunaan
asam
lemak tidakjenuh perlu dikaji rnanfaatnya. Optimasi bioproses di dalarn rumen diharapkan dapat
memhcu fermentasi dan pertumbuhan mikroba rumen, sehingga dapat meningkatkan
produksi ternak ruminansia.
Tujuan Penelitian
Berdasarkan pemikiran di atas, rangkaian percobaan dalam penelitian ini
ditujukan untuk memadukan teknologi defaunasi, suplementasi sulfur, asam lemak
tidak jenuh dan asam amino rantai cabang, sebagai upaya optimasi bioproses pakan
serat di -cialam rumen. Dengan pendekatan bioproses tersebut diharapkan porsi
pakan serat di dalam ransum dapat lebih dominan. Pertumbuhan mikroba rumen,
kecernaan zat-zat makanan dan pertumbuhan ternak merupakan tolok ukur utama
keberhasilan pendekatan bioproses yang dilakukan.
Kegunaan Penelitian
Hasil penel itian, di samping untuk pengembangan ilmu pengetahuan, juga
diharapkan membuka jalan untuk lebih efektif memanfaatkan limbah serat sebagai
pakan utama ternak ruminansia. Pemanfaatan limbah serat sebagai pakan ternak
,
akan memberikan nilai tambah yang berarti pada sumberdaya limbah serat tersebut.
Penggunaan limbah serat tersebut, selain menunjang pembangunan peternakan, juga
akan menjadi alternatif menarik dalam mencegah pencemaran lingkungan yang
semakin serius. Pengolahan limbah serat dalam sekala besar dapat melahirkan
cabang usaha
baru
dalam agroindustri, sehingga dapat menciptakan lapangan kerjabaru. Pemanfaatan sumberdaya dengan efisien akan meningkatkan keunggulan
TIN
JAUAN
PUSTAKA
Sistem Pencernaan pada Ternak Ruminansia
' Pencemaan adalah rangkaian proses perubahan fisik dan kimia yang dialami
oleh bahan makanan di dalam alat pencernaan. Proses pencernaan makanan pada
ternak ruminansia relatif lebih kompleks dibandingkan dengan proses pencernaan
pada jenis ternak lainnya. Menurut Sutardi (1980) proses pencernaan pada ternak
ruminansia te rjadi secara mekanis (di dalarn mulut), secara fermentatif (oleh enzim-
enzim yang berasal dari mikroba rumen) dan secara hidrolitis (oleh enzim-enzim
pencernaan hewan induk semang). Lokasi (posisi) proses pencernaan fermentatif
bervariasi antar jenis ternak. Posisi tersebut akan menentukan karakteristik pakan
yang sesuai untuk jenis ternak bersangkutan.
Organ pencernaan pada temak ruminansia terdiri atas 4 bagian penting, yaitu:
mulut, perut, usus halus dan organ pencernaan bagian belakang (hind gut). Perut
ternak ruminansia dibagi menjadi 4 bagian, yaitu retikulum (perut jala), rumen
(perut beludru), omasum (perut buku), dan abomasum (perut sejati). Dalam studi
fisiologi pencernaan ternak ruminansia, rumen dan retikulum sering dipandang
sebagai organ tunggal (single organ) dengan sebutan retikulorumen. Omasum dise-
but sebagai perut buku karena dipenuhi oleh lembaran jaringan (tissu leaves), yaitu
sekitar 100 lembar. Fungsi omasum belum terungkap dengan jelas, tetapi pada
organ tersebut ada penyerapan air, amonia, asam lemak terbang dan elektrolit, serta
ada produksi amonia dan mungkin asam lemak terbang (Forbes dan France, 1993).
Termasuk organ pencernaan bagian belakang adalah sekum (cecum), kolon (colon)
dan rektum (rectum). Pada organ pencernaan bagian belakang tersebut juga terjadi
aktivitas fermentasi. Namun, belum banyak informasi yang terungkap tentang pera-
Proses pencernaan fermentatif di dalam retikulo-rumen terjadi amat intensif
dan dalam kapasitas yang sangat besar. Proses pencernaan tersebut terletak sebelum
usus halus (organ penyerapan utama). Hal tersebut sangat menguntungkan, karena:
(1) {akan dapat diubah dan disajikan dalam bentuk produk fermentasi yang mudah
diserap, (2) ternak ruminansia menjadi mampu memanfaatkan pakan serat dalam
jumlah lebih banyak dan lebih efisien. Sebaliknya, pada ternak
kuda,
babi dan ayarnproses'pencernaan fermentatif terjadi setelah usus halus, yaitu di sekum (hind gut
fermenter). Karena itu kelompok ternak ini memerlukan pakan yang lebih bermutu
(lebih rendah kadar serat) dibandingkan dengan pakan ternak ruminansia.
Pada sistem pencernaan ternak ruminansia terdapat suatu proses yang disebut
memamah biak (ruminasi). Pakan berserat (hijauan) yang dimakan di tahan untuk
sementara di dalam rumen. Pada masa hewan tersebut beristirahat, pakan dari
dalam rumen lalu dikembalikan ke mulut (proses regurgitasi) untuk dikunyah kemba-
li (proses remastikasi), kemudian pakan ditelan kembali (proses redeglutasi). Selan-
jutnya pakan tersebut dicerna lagi oleh enzim-enzim mikroba rumen (microbial
attack). Kontraksi retikulorumen yang terkoordinasi dalam rangkaian proses tersebut
bermanfaat pula untuk pengadukan digesta, inokulasi digesta dan penyerapan nutri-
en. Selain itu kontraksi retikulorumen juga bermanfaat untuk pergerakan digesta
meninggal kan reti kulorumen vmelalui recikulo-omasal oriJice.
Pada proses pencernaan fermentatif di dalam rumen karbohidrat struktural
berupa serat (selulosa dan hemiselulosa) dan karbohidrat sederhana yang fermentabel
(gula, pati) mengalami fermentasi anerob oleh mikroba rumen menjadi asam-asam
lemak terbang (VFA), gas metan (CH,) dan gas karbon dioksida (CO,). Sebagian
VFA akan diserap melalui dinding rumen, lalu masuk ke dalam aliran darah dan
ruminansia asam lemak terbang meliputi sekitar 50% dari energi pakan yang tercer-
na. Meskipun kontribusi proses pencernaan fermentatif tersebut cukup
besar,
etapipada tahap tersebut sebagian energi pakan ada yang terbuang sebagai gas metan dan
4
panas fermentasi
.
Aktivitas pengunyahan dan ruminasi di dalam mulut membantu perombakan
pakan secara mekanis. Selain itu aktivitas tersebut juga merangsang proses sekresi
cairan 'saliva dari mulut. Pada sapi produksi saliva sangat banyak, dapat mencapai
180 literlhari pada ransum hijauan berkualitas baik (Church, 1988), dan kondisinya
lebih alkalis (pH 8.2). Saliva tersebut merupakan sumber cairan buffer utama untuk
stabilisasi ekosistem rumen. Selain berperan sebagai bufer, manfaat lain saliva
adalah bagian dari siklus nitrogen, sebagai lubrikan (pelicin) dan berperan dalam
ekonomi air. Sekitar 70% nitrogen dalam saliva ada dalam bentuk urea.
Ukuran rumen dan retikulum sangat besar, dapat mencapai 15-2296 dari
bobot tubuh ternak (Sutardi, 1981). Jumlah tersebut meliputi sekitar 75% dari
seluruh volume organ pencernaan ternak ruminansia (Van Soest, 1982). Retikulo-
rumen dihuni oleh macam-macam mikroba. Dari segi pencernaan zat-zat makanan,
peranan rumen memberi andil sekitar 40-70% dari angka kecemaan bahan organik
ransum (Hvelplund dan Madsen, 1985). Karena itu rumen
dan
retikulum (forestom-achs) merupakan bagian yang' sangat penting dari organ pencernaan ruminansia.
Rumen dihuni oleh tidak kurang dari 4 jenis mikroorganisme anerob, yaitu
bakteri, protozoa, fungi (jamur) dan virus (Preston dan Leng, 1987). Dua jenis
mikroorganisme yang disebut pertama telah lama dipelajari dan diungkap peranannya
dalam fermentasi rumen dan manfaatnya sebagai pemasok nutrien untuk hewan
induk sernang. Akhir-akhir ini studi tentang mikroba rumen mulai memperhatikan
monas communis dan Piromonas commutis. Peranan fungi rumem dalarn pencemaan
pakan diperkirakan cukup berarti, terutama pada kelompok temak yang mendapat
ransum tinggi kadar serat.
6
Kehadiran fungi di dalam rumen diakui sangat bermanfaat bagi kecernaan
fraksi serat dalam pakan. Fungi dapat membentuk koloni pada jaringan lignoselulo-
sa partikel pakan (Fonty et
al.,
1990). Dengan cara demikian fungi menjadi mikro-ba pelopor dalam aktivitas selulolitik di dalam rumen. Rhizoid fungi tumbuh jauh
menembus dinding sel serat tanaman, sehingga menciptakan akses bagi mikroba
(bakteri) dan enzim selulasenya untuk mencema serat. Namun, masih banyak aspek
yang perlu diungkap ten tang peranan fungi rumen, terutama menyangkut aspek
kuantitas dan kualitasnya sebagai sumber nutrien untuk hewan induk semang.
Bakteri merupakan mikroba rumen yang paling banyak jenisnya dan lebih
beragam macam substratnya. Selain itu populasinya sangat tinggi, yaitu 10''
-
10"cacahan sel per gram isi rumen (Yokoyama dan Johnson, 1988). Berdasarkan
macam substrat yang disukain ya bakteri rumen dapat dikelompokkan sebagai bakteri
pencerna selutosa (misalnya Rurninococcus. albus), pencema hemiselulosa (Buryn'vi-
brio jibrisolvens), pencerna pati (Bacteroides amylophilus), pencerna gula (Lactoba-
cillus ruminus) dan bakteri pengguna produk sekunder (pemakai laktat). Sekitar
38 % bakteri rumen memilikf aktivitas proteolitik.
Protozoa rumen lebih sedikit populasinya, yaitu l d
-
106 cacahan sel per mlisi rumen (Yokoyama dan Johnson, 1988), tetapi dari segi jumlah biomassa temyata
cukup besar. Produk fermentasi yang dihasilkan protozoa termasuk asam asetat,
asam butirat, asam iaktat, gas karbon dioksida dan gas hidrogen (Russell dan Hes-
pell, 1981). Protozoa lebih menggemari substrat yang fermentabel (pati, gula dan
(ditelan) oleh protozoa. Proses tersebut ternyata ada manfaatnya, yaitu memperlam-
bat proses konversi karbohidrat menjadi asam laktat oleh bakteri rumen, sehingga
rumen terhindar dari penurunan pH secara drastis.
#
Akumulasi asam laktat
dan
penurunan pH rumenakan
mengganggu keselunt-han sistem rumen. Pada keadaan tersebut populasi Streptococcus bovis dan Lactoba-
cilli meningkat pesat. Russell dan Dombrowski (1980) memperlihatkan melalui
kuItivasi kultur murni daIam kemostat, bahwa S. bovis dan Lactobacillus vitulinus
lebih toleran terhadap kondisi pH yang rendah. Secara kontinyu sesungguhnya
sistem rumen mendapat pasokan bufer bikarbonat dan fosfat yang berasal dari saliva.
Meskipun demikian pada keadaan tertentu seperti tersebut di atas kapasitas bufer
(buffering capacity) dapat terlampaui.
Banyaknya jenis mikroorganisme rumen dan masing-masing mikroorganisme
memiliki produk fermentasi intermedier dan produk fermentasi akhir yang berma-
cam-macam, menyebabkan kehidupan di dalam rumen menjadi sangat kompleks.
Terdapat interaksi atau interrelasi yang luas antar mikroorganisme di dalam rumen.
Bentuk interelasi tersebut dapat berupa ketergantungan akan substrat, saling mengun-
tungkan, kompetisi substrat atau berupa hubungan yang merugikan. Studi tentang
ekologi mikroba rumen umumnya masih dipusatkan pada dua aspek utama, yaitu
pengendalian populasi mikroba rumen dan peningkatan peranan mikroba rumen
dalam mencerna pakan
.
Pasokan Zat-Zat Makanan untuk Ternak Rurninansia
Pakan ternak ruminansia biasanya dikelompokkan menjadi pakan hijauan dan
pakan konsentrat. Sampai saat ini belum ada definisi yang mantap tentang batasan
kelompok pakan hijauan (roughages) adalah jenis rumput-rumputan, tanaman le-
guminosa, aneka jerami, silase dan limbah serat tanaman. Pakan hijauan biasanya
berciri amba (bulky) dan tinggi kadar serat (fibrous). Karena itu pakan hijauan
cenderung memiliki nilai kecernaan energi dan kepadatan energi yang rendah.
Termasuk kelompok pakan konsentrat meliputi produk biji-bijian dan limbah
pengolahannya serta jenis bungkil-bungkilan. Pakan konsentrat umumnya berciri
rendah kadar serat dan lebih kaya fraksi isi sel. Keadaan tersebut menyebabkan nilai
kecernaan energi pakan konsentrat cenderung lebih tinggi dibandingkan dengan
kecernaan energi pakan hijauan. Karena perbedaan yang kontras tersebut maka
nisbah pakan hijauan dan konsentrat dalam ransum erat hubungannya dengan paso-
kan energi untuk proses-proses biologis ternak.
Pada tabel-tabel komposisi pakan informasi kandungan energi pakan dinyata-
kan dalam bentuk energi tercerna (DE), energi termetabolisasi (ME), energi netto
(NE) atau dalani satuan TDN (total digestible nutrients). Namun, informasi tersebut
biasanya tidak dilengkapi dengan data konsumsi bahan keringnya. Akibatnya sering
terjadi kesulitan dalam penyusunan ransum, terutama menyelaraskan kebutuhan
energi ternak dengan kemampuan konsumsi bahan keringnya. ~ e s u l i t a n menjadi
semakin besar pada daerah tropik, karena umumnya pakan hijauan yang tersedia
berkualitas rendah.
Fraksi karbohidrat dalam ransum ternak ruminansia sangat dominan, dapat
berkisar 60
-
75 % dari bahan kering ransum (Sutardi, 1980). Komponen karbohi-drat tersebut dapat berasal dari isi sel (misalnya gula dan pati) dan dinding sel
(misalnya selulosa dan hemiselulosa). Ternak ruminansia mampu memanfaatkan
fraksi serat pakan (selulosa dan hemiselulosa) karena sistem pencernaannya dileng-
anerob oleh mikroba rumen. Di dalarn retikulo-rumen karbohidrat struktwal men-
galami perombakan yang sangat intensif menjadi produk yang dapat diserap dan
dicerna di usus halus.
4
Kebutuhan zat-zat makanan pada ternak ruminansia dipenuhi dari
3
sumber,yaitu: (1) zat-zat makanan hasil fermentasi dalam rumen,
(2)
zat-zat makanan asalpakan yang 1010s degradasi rumen, dan (3) dari mikroba mmen yang tercerna di
dalam'organ pencernaan pascarumen. Pada ternak yang mendapat ransum dengan
bahan utama pakan serat berkualitas rendah (low quality roughages), maka pasokan
zat-zat makanan lebih diharapkan berasal dari mikroba rumen dan produk fermentasi
(asam lemak terbang). Pada kondisi tersebut sebagian
besar
asam-asam amino yangsampai hi usus halus berasal dari mikroba rumen. Menurut Sniffen dan Robinson
(1987) sumbangan protein asal mikroba rumen terhadap kebutuhan asam-asam amino
ternak ruminansia dapat mencapai 40
-
80%. Selain untuk deposisi protein tubuh,asam amino juga merupakan bahan baku sintesis glukosa melalui proses glukoneo-
genesis. Sumbangan asam amino terhadap kebutuhan glukosa dapat mencapai 36%
(Bergman, 1983).
Asam lemak terbang (VFA) hasil proses fermentasi di dalam rumen telah
diketahui merupakan sumber energi utama bagi ternak ruminansia. Menurut En-
sminger et ul. (1990) sumbangan energi yang berasal dari asam lemak terbang dapat
mencapai 60
-
80% dari kebutuhan energi pada ternak ruminansia. Hampir semuaVFA yang diproduksi diserap di mmen, retikulum dan omasum. Hanya sedikit VFA
yang sampai di abomasum (Fahey dan Berger, 1988). Karena itu pada ternak yang
mendapat ransum pakan serat berkualitas rendah ada 2 aspek penting yang perlu
ditingkatkan, yaitu aspek fermentabilitas pakan dan aspek pertumbuhan mikroba
Asam asetat yang diserap melalui dinding rumen sebagian kecil dikonversi
menjadi badan keton (ketone bodies). Tetapi sebagian besar asam asetat (80%) bisa
sampai dihati dan masuk peredaran darah tanpa mengalami oksidasi. Selanjutnya
4
sebagian asam asetat dioksidasi lewat siklus TCA (tricarboxylic acid) atau digunakan
untuk proses lipogenesis (Fahey dan Berger, 1988). Selama proses penyerapan
melalui epitel rumen asam propionat
(2
-
5
96) dikonversi menjadi asam laktat, dan
sisanya masuk ke aliran darah portal (Elliot, 1980). Sebagian besar asam propionat
yang sampai di hati dikonversi menjadi glukosa melalui proses glukoneogenesis.
Untuk mencapai pasokan asam-asam amino yang seimbang ke usus halus
maka komposisi asam-asam amino asal pakan yang 1010s degradasi haruslah saling
melengkapi dengan asam-asam amino
asal
mikroba rumen. Dengan demikian makainformasi komposisi asam-asam amino mikroba rumen perlu diketahui. Pada kenya-
taannya data komposisi asam-asam amino mikroba rumen cukup bervariasi. Namun,
Clark et al. (1992) berhasil menyajikan hasil review komposisi bakteri rumen dari
35 percobaan yang melibatkan 441 sampel bakteri rumen. Hasil tersebut diperlihat-
kan pada Tabel 1.
Peningkatan fermentabilitas pakan berserat tinggi dapat diupayakan dengan
memberikan beberapa perlakuan pendahuluan (pretreatment) pada pakan. Sebagai
contoh, misaln ya dengan pe!rlakuan kimia (perlakuan alkali, amoniasi), perlakuan
biologi (fermentasi oleh berbagai jenis mikroorganisme erob atau anerob), dan
perlakuan fisik (penggilingan, pembuatan pellet, dan pemanasan pada tekanan a
tinggi). Metode-metode tersebut sudah banyak dikaji dan telah memperlihatkan hasil
yang cukup baik. Namun, penerapan metode-metode tersebut secara komersial
masih menghadapi ban yak kendala dan harus segera dipadukan dengan usaha men-
nbel 1. Komposisi Asam-Asarn Amino
W r i
RumenAsam Amino Minimum Maksimum Rataan
4 Arginin Histidin Isoleusin Leusin Lisin Metionin Fenilalanin Treonin Valin
-
AlaninAsam aspartat Asam Glutamat Glisin
Prolin Serin Tirosin
(g1100g asarn amino)
3.8 6.8 5.1
1.2 3.6
2.0
4.6 6.7 5.7
5.3 9.7 8.1
4.9 9.5 7.9
1.1 4.9 2.6
4.4 6.3 5.1
5.0 7.8 5.8
4.7 7.6 6.2
5 -0 8.6 7.5
10.9 13.5 12.2
11.6 14.4 13.1
5.0 7.6 5.8
2.4 5.3 3.7
3.4 5.4 4.6
3.9 7.7 4.9
Al-Ani dan Smith (1988) melaporkan bahwa perlakuan kimia yang disertai
dengan autoclave terhadap bagas tebu dapat meningkatkan nilai kecernaan in vitro
sampai 75 %
.
Hasil serupa diperlihatkan Sutardi (1994) bahwa teknologi amoniasidapat meningkatkan manfaat pakan. Selain cara kimia, fisik dan biologi, fermentabi-
litas pakan ada kemungkinan dapat diperbaiki melalui optimasi bioproses dalam
rumen, dalam arti menciptakan kondisi ekologi yang mendukung bioproses dan
pertumbuhan mikroba rumen yang optimal.
Untuk pertumbuhan mikroorganisme yang optimal, semua nutrien prekursor
harus tersedia dalam konsentrasi yang optimum di dalam rumen. Nutrien tersebut
termasuk energi (dalam bentuk ATP), nitrogen, asam-asam amino, mineral dan
vitamin. Huber dan Kung (1981) menyatakan bahwa efisiensi fermentasi dan sintesis
protein mikroba rumen dapat dimaksimumkan bila semua prekursor tersebut terse-
dia dalam jumlah yang cukup. Hal ini berarti bahwa suplementasi suatu nutrien
harus diselaraskan dengan ketersediaan nutrien lainnya.
Kecernaan pakan serat dalam rumen pada dasarnya adalah kerja enzim-enzim
pencerna serat yang diproduksi oleh mikroba rumen. Untuk mencerna fraksi serat
dalam pakan, ternak ruminansia sepenuhnya tergantung kepada peranan mikroba
rumen. Sebagai kelompok ternak mamalia, ternak ruminansia tidak memproduksi
enzim-enzim yang dapat menghidrolisis selulosa atau hemiselulosa. Mengingat
peranan strategis enzim-enzim mikroba rumen tersebut, maka optimasi biofermentasi
rumen melalui peningkatan konsentrasi enzim relatif terhadap substrat harus menjadi
sasaran utama.
Peningkatan konsentrasi enzim di dalam rumen reIatif terhadap jumlah sub-
strat (serat) di harapkan dapat meningkatkan laj
u
kecernaan atau perombakan sub-strat. Dapat diupayakan 2 pendekatan untuk meningkatkan produksi atau konsentra-
si enzim di dalam rumen, yaitu dengan meningkatkan populasi mikroba rumen
penghasil enzim (pendekatan kuantitatif) dan melalui rekayasa genetik mikroba
rumen (pendekatan kualitatif) untuk mendapatkan strain mikroba rumen yang mampu
menghasilkan enzim lebih banyak. Selama lima tahun terakhir rekayasa genetik
mikroba rumen mulai memperlihatkan hasil menggembirakan, meskipun masih
Metabolisme Karbohidrat daiam Rumen
Pakan ternak ruminansia sebagian besar berupa kompleks polimer, misalnya
selulosa, hemiselulosa, pati, pektin dan protein. Bentuk yang demikian kompleks
e
menyebabkan pakan ruminansia memiliki nilai kelarutan (solubility) yang rendah.
Pada sisi lain, mekanisme transport nutrien pada bakteri rumen ternyata hanya
mampu mengangkut nutrien-nutrien yang terlarut dengan bobot molekul yang rendah
re us sell
dan Hespell, 1981). Karena itu proses pemanfaatan pakan di dalam rumen terlebih dahulu diawali kerja enzim-enzim ekstraseluler yang merombak partikelpakan (kompleks polimer) menjadi bentuk monomernya atau sen yawaan-senyawaan
yang berbobot molekul rendah. Enzim ekstraseluler bekerja pada permukaan parti-
kel pakan. Dengan demikian luas permukaan partikel pakan yang terbuka terhadap
akses enzim-enzim pencernaan menjadi faktor yang sangat penting.
Berlandaskan pada fenomena tersebut di atas, dapat dipahami bahwa sifat-
sifat fisik dan sifat-sifat kimia pakan
akan
sangat erat kaitannya dengan aspek potensidegradasi pakan di dalam rumen. Sifat-sifat fisik pakan meliputi aspek keambaan
(bulkiness), daya menyerap air (water regain capacity) dan aspek kelarutan (solubili-
ty). Ketiga sifat-sifat fisik tersebut erat kaitannya dengan aspek degradasi pakan
(Sutardi er al., 1994). Sifat-sifat kimia pakan meliputi komposisi kimia nutrien, kapasitas tukar kation (cation kxchange capacity) dan kapasitas penyangga (buffering
capacity )
.
Laju pertumbuhan mikroba dalam rumen sangat tergantung kepada keterse-
diaan karbohidrat
.
Laj u pencemaan karbohidrat merupakan salah satu faktor penen-tu produksi protein mikroba rumen. Selain sebagai sumber kerangka karbon, karbo-
hidrat adalah sumber energi untuk mikroba, dalam bentuk ATP (adenosine tryphos-
dalam rumen dilepaskan energi dalam bentuk ATP, CH,
dan panas
fermentasi. Duabentuk energi yang disebut terakhir tidak bermanfaat bagi ternak. Pertumbuhan
mikroba rumen proporsional terhadap jumlah ATP yang dihasilkan
dari
katabolisme *sumber energi.
Nilai Y, diperkenalkan oleh Bauchop dan Elsden (1960), untuk menyata-
kan gram bahan kering biomassa sel yang terbentuk per mol ATP yang digunakan.
Nilai
,
,
Y
mencapai sekitar 29-30 untuk mikroba rumen yang dikultur pada substratyang berupa monomer (Hespell and Bryant, 1979). Namun, untuk media sederhana
dengan substrat karbohidrat dan garam organik nilai
,
Y
berkisar 20-29. Selain itunilai YATp tersebut temyata dipengaruhi juga oleh laju dilusi (dilution rate) cairan
rumen. Semakin tinggi laju dilusi nilai Y,, cenderung semakin tinggi (Isaacson et
al.
,
1975).Karbohidrat dalam pakan dapat dikelompokkan menjadi karbohidrat struktur-
a1 (fraksi serat) dan karbohidrat nonstruktural (fraksi yang mudah tersedia). Propor-
si