ALEL MONOMORFIK GEN
OSTEOPONTIN
(OPN|
Bsr
I)
PADA SAPI BALI DENGAN METODE PCR-RFLP
KOMANG ALIT PARAMITASARI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Alel Monomorfik Gen Osteopontin (OPN|BsrI) pada Sapi Bali dengan Metode PCR-RFLP adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Mei 2013
ABSTRAK
KOMANG ALIT PARAMITASARI. Alel Monomorfik Gen Osteopontin (OPN|BsrI) pada Sapi Bali dengan Metode PCR-RFLP. Dibimbing oleh JAKARIA dan CECE SUMANTRI.
Bangsa sapi bali sebagai sumberdaya genetik ternak asli Indonesia memiliki keunggulan, salah satunya adalah daya reproduksi yang tinggi. Gen osteopontin (OPN) merupakan anggota dari alur POU class 1 homeobox 1 (POU1F1) yang berkaitan dengan sifat reproduksi. Penelitian ini bertujuan untuk mengidentifikasi ada tidaknya polimorfisme gen osteopontin (OPN) sebagai pengontrol sifat reproduksi pada sapi bali di Pulau Bali dengan menggunakan metode PCR-RFLP. Sampel sapi bali sebanyak 100 sampel yang berasal dari BPTU sapi bali dideteksi keberadaan keragaman genetiknya pada gen OPN menggunakan enzim restriksi BsrI yang memotong sekuens basa ACTGG|N. Amplifikasi gen OPN menghasilkan fragmen dengan panjang 290 bp yang terletak pada intron 4 dengan suhu annealing 60 oC selama 20 detik. Hasil analisis PCR-RFLP menunjukkan sapi bali yang dianalisis memiliki alel monomorfik T adalah 100%. Hasil sekuens fragmen gen OPN intron 4 menunjukkan terdapat mutasi antara basa sitosin (C) dengan timin (T).
Kata kunci: gen osteopontin (OPN), PCR-RFLP, sapi bali
ABSTRACT
KOMANG ALIT PARAMITASARI. Monomorphic Allele Osteopontin Gene (OPN|BsrI) in Bali Cattle Using PCR-RFLP Method. Supervised by JAKARIA polymorphisms in bali cattle at Bali Island using the PCR-RFLP method. A total of 100 bali cattle from BPTU bali cattle were identified using BsrI restriction enzyme that cuts the sequence of ACTGG|N bases. OPN gene amplification was performed with annealing temperature of 60 oC for 20 seconds resulting in a fragment with the length of 290 bp which is located at intron 4. The results of PCR-RFLP analysis indicated that the analyzed bali cattles which have monomorphic T allele was 100%. OPN gene fragment intron 4 sequences showed that there was a mutation between cytosine (C) base with thymine (T).
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Peternakan
pada
Departemen Ilmu Produksi dan Teknologi Peternakan
ALEL MONOMORFIK GEN
OSTEOPONTIN
(OPN|
Bsr
I)
PADA SAPI BALI DENGAN METODE PCR-RFLP
KOMANG ALIT PARAMITASARI
DEPARTEMEN ILMU PRODUKSI DAN TEKNOLOGI PETERNAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Alel Monomorfik Gen Osteopontin (OPN|BsrI) pada Sapi Bali dengan Metode PCR-RFLP
Nama : Komang Alit Paramitasari NIM : D14090089
Disetujui oleh
Dr Jakaria, SPt MSi Pembimbing I
Prof Dr Ir Cece Sumantri, MAgr Sc Pembimbing II
Diketahui oleh
Prof Dr Ir Cece Sumantri, MAgr Sc Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan pada bulan Desember 2012 sampai Maret 2013 ini adalah gen reproduksi pada sapi bali, dengan judul Alel Monomorfik Gen Osteopontin (OPN|BsrI) pada Sapi Bali dengan Metode PCR-RFLP.
Terima kasih penulis ucapkan kepada Dr Jakaria, SPt MSi selaku pembimbing akademik dan pembimbing skripsi, Prof Dr Ir Cece Sumantri, MAgrSc selaku pembimbing skripsi, Ir Rini H. Mulyono, MSi dan Dr Iwan Prihantoro, SPt MSi selaku penguji ujian akhir, dan Dr Ir Sri Darwati, MSi selaku perwakilan departemen dalam ujian akhir sarjana penulis. Ungkapan terima kasih juga disampaikan kepada kedua orang tua, I Nyoman Arnaya dan Putu Agustini Eliyati, kedua kakak, Putu Indira Pradnyawati dan Kadek Noni Lokasari, serta seluruh keluarga, atas segala doa dan kasih sayangnya. Di samping itu, penulis sampaikan terima kasih kepada I Putu Arimbawa Pande, Kak Eryk, Kak Ferdy, Kak Irine, Kak Isyana, Rany Pratiwi, Roaslein Putri, Opak, Adit, Eja, Lusiana, Tasya, dan teman-teman laboratorium genetika molekuler ternak, serta keluarga besar Golden Ranch IPTP 46 atas segala dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Mei 2013
DAFTAR ISI
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 1
Ruang Lingkup Penelitian 1
METODE 2
Lokasi dan Waktu Penelitian 2
Bahan 2
Alat 2
Prosedur 3
Ekstraksi DNA 3
Amplifikasi DNA 3
Genotyping Gen OPN 4
Sekuens Fragmen Gen OPN Intron 4 4
Analisis Data 4
Frekuensi Alel Gen OPN 4
Sekuens Fragmen Gen OPN Intron 4 5
HASIL DAN PEMBAHASAN 5
Amplifikasi Gen Osteopontin (OPN) 5
Identifikasi Genotipe Gen Osteopontin (OPN) 6
Frekuensi Alel Gen Osteopontin (OPN) 6
Homologi dan Deteksi Mutasi Gen Osteopontin (OPN) 7
SIMPULAN DAN SARAN 9
DAFTAR PUSTAKA 9
DAFTAR GAMBAR
1 Posisi penempelan primer pada fragmen gen OPN dan situs
pemotongan enzim BsrI 3
2 Hasil amplifikasi gen osteopontin pada gel agarose 1.5% 5 3 Hasil PCR-RFLP fragmen gen osteopontin (OPN|BsrI) pada gel
agarose 2% 6
4 Hasil alignment nukleotida sekuens gen OPN|BsrI intron 4 sapi bali dengan GenBank Bos taurus (kode akses GU143824) 8 5 Rekonstruksi struktur gen OPN pada sapi dengan SNP (single nuclotide
polymorphism) pada intron 4 8
DAFTAR LAMPIRAN
1 Sekuens gen osteopontin (OPN) yang diakses di GenBank kode akses
GU143824 11
2 Hasil BLAST nukleotida sekuens gen OPN sapi bali dengan sekuens
GenBank kode akses GU143824 12
PENDAHULUAN
Latar Belakang
Sapi bali (Bos javanicus) merupakan salah satu ternak asli Indonesia hasil domestikasi banteng (Bibos banteng) (Talib 2002) yang merupakan sumberdaya genetik ternak yang tidak ternilai harganya dan telah diakui oleh FAO sebagai salah satu bangsa ternak di dunia (DGLS 2003). Sapi bali memiliki beberapa keunggulan yaitu mampu beradaptasi terhadap lingkungan marginal, selain itu memiliki persentase karkas tinggi (56%-57%) serta angka kelahiran tinggi (69%-83%) (Talib 2002). Sapi bali memiliki daya reproduksi yang tinggi dibandingkan dengan bangsa-bangsa sapi lain yang dikembangkan di Indonesia. Oleh karena itu, sapi bali merupakan bangsa sapi yang potensial untuk dikembangkan dan dimanfaatkan untuk memenuhi kebutuhan daging secara nasional.
Upaya peningkatan mutu genetik ternak sapi bali dapat dilakukan melalui seleksi yaitu mempertahankan sifat-sifat unggul yang terdapat pada sapi bali seperti kemampuan sifat reproduksi tinggi. Seleksi sifat reproduksi sapi bali pada tingkat molekuler DNA dapat dilakukan dengan cara menganalisis gen-gen utama pengontrol sifat reproduksi.
Khatib et al. (2009) menyatakan bahwa sifat reproduksi dikontrol oleh beberapa gen, salah satunya adalah gen osteopontin (OPN). Gen osteopontin (OPN) atau sering disebut sebagai secreted phospoprotein 1 (SPP1) pada sapi terletak pada kromosom 6 (Leonard et al. 2005) yang terdiri atas tujuh exon dan enam intron dengan ukuran sekitar 7 000 basa dari total genom (Pareek et al. 2008). Keragaman gen OPN di intron 4 telah ditemukan oleh beberapa hasil penelitian seperti sifat produksi susu pada sapi FH (Leonard et al. 2005), sifat pertumbuhan pasca sapih pada sapi pedaging (White et al. 2007), sifat fertilitas pada embrio sapi (Khatib et al. 2009), dan sifat kembar pada sapi FH (Anggraeni et al. 2012). Keragaman atau polimorfisme gen OPN pada sapi bali dapat diketahui dengan menggunakan teknik Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP).
Tujuan Penelitian
Penelitian ini bertujuan untuk mengidentifikasi ada tidaknya polimorfisme gen osteopontin (OPN) pada sapi bali di Pulau Bali dengan menggunakan metode PCR-RFLP. Selain itu, tujuan penelitian ini adalah untuk mendeteksi ada tidaknya mutasi pada situs enzim pemotong BsrI gen OPN intron 4.
Ruang Lingkup Penelitian
2
gen OPN intron 4 dianalisis melalui teknik sequencer. Analisis data dilakukan melalui pendekatan frekuensi alel dan analisis Basic Local Alignment Search Tools (BLAST) nukleotida.
METODE
Lokasi dan Waktu Penelitian
Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler Ternak, Bagian Pemuliaan dan Genetika, Fakultas Peternakan, Institut Pertanian Bogor. Penelitian ini berlangsung dari bulan Desember 2012 sampai dengan Maret 2013.
Bahan
Sampel DNA yang dianalisis berasal dari sampel darah 100 ekor sapi bali di BPTU Bali yang merupakan koleksi Laboratorium Genetika Molekuler Ternak. Bahan-bahan yang digunakan untuk ekstraksi DNA adalah NaCl 0.2%, Sodium Dodecyl Sulfat (SDS) 10%, enzim Proteinase-K (5 mg/ml), 1 x STE (NaCl 5 M, tris HCl 2 M, EDTA 0.2 M), chlorofom isoamil alkohol (CIAA), phenol, EtOH 96%, EtOH 70%, dan buffer Tris EDTA (TE) 80%. Bahan yang digunakan untuk amplifikasi, elektroforesis dan analisis RFLP adalah sampel DNA hasil ekstraksi, destilation water (DW), 10 x buffer, MgCl2, pasangan primer forward dan reverse fragmen gen OPN, enzim Taq polymerase (Fermentas), deoxy Nucleotide Triphospat (dNTPs), produk PCR, serbuk agarose, 0.5 x Tris-Borat EDTA (TBE), Ethidium Bromide (EtBr), loading dye (0.01% Xylene Cyanol, 0.01% Bromthymol blue dan 50% gliserol), marker 100 bp, enzim restriksi BsrI, dan buffer B.
Bahan yang digunakan untuk analisis sekuensing adalah produk PCR dan primer forward fragmen gen OPN. Primer yang digunakan untuk mengamplifikasi gen osteopontin (OPN|BsrI) berdasarkan Leonard et al. (2005) adalah forward: 5´-GCAAAT-CAGAAGTGTGATAGAC-3´ dan reverse: 5´ CCAAGCCAAACGTATGAGTT-3´.
Alat
3
Prosedur
Ekstraksi DNA
Ekstraksi DNA dilakukan berdasarkan metode Sambrook et al. (1989) yang dimodifikasi. Sampel darah diambil sebanyak 200 µl dan ditambahkan dengan 1 000 µl NaCl 0.2%, kemudian dihomogenkan menggunakan vortex dan didiamkan selama lima menit. Sampel disentrifugasi pada kecepatan 8 000 rpm selama lima menit dan bagian supernatan dibuang. Endapan sampel ditambahkan dengan 10 µl Proteinase-K (5mg/ml), 350 µl 1 x STE dan 40 µl SDS 10%, kemudian diinkubasi pada suhu 55 oC selama dua jam. Larutan yang telah diinkubasi ditambahkan 40 µl NaCl 5 M, 400 µl phenol dan 400 µl CIAA kemudian dikocok perlahan pada suhu ruang selama satu jam. Tahap selanjutnya adalah larutan disentrifugasi dengan kecepatan 12 000 rpm selama lima menit sehingga terbentuk fase DNA. Fase DNA yang terbentuk diambil sebanyak 40 µl dan dipindahkan ke tabung 1.5 ml baru, ditambahkan dengan 40 µl NaCl 5 M dan 800 µl EtOH 96%, dihomogenkan, kemudian diistirahatkan overnight pada suhu -20 oC. Sampel DNA disentrifugasi pada kecepatan 12 000 rpm selama lima menit dan supernatan yang terbentuk dibuang, kemudian endapan ditiriskan sampai kering dan dilarutkan dalam 100 µl buffer TE 80%.
Amplifikasi DNA
Sampel DNA hasil ekstraksi sebanyak 1 µl dan pereaksi amplifikasi DNA yang terdiri dari 10.95 µl DW, 0.2 µl primer, 0.05 µl Taq polymerase, 1.5 µl buffer, 0.3 µl dNTPs dan 1 µl MgCl2 dimasukkan ke dalam tabung PCR kemudian dihomogenkan. Amplifikasi DNA berlangsung di dalam mesin PCR Applied Biosystems dengan kondisi suhu predenaturasi 95 oC selama lima menit, 35 siklus untuk tahapan denaturasi 95 oC selama 10 detik, annealing pada suhu 60 oC selama 20 detik, dan elongasi pada suhu 72 oC selama 30 detik dan elongasi akhir pada suhu 72 oC selama lima menit dalam satu siklus. Produk PCR dielektroforesis menggunakan agarose 1.5%. Posisi penempelan primer pada amplifikasi DNA dapat dilihat pada Gambar 1.
4
Genotyping Gen OPN
Produk PCR sebanyak 5 μl ditambahkan dengan 1 μl DW, 0.3 μl enzim restriksi BsrI, dan 0.7 μl buffer B, kemudian diinkubasi di dalam inkubator pada suhu 65 °C selama 16 jam (overnight). Elektroforesis produk PCR-RFLP diawali dengan membuat gel agarose 2% yang dibuat dengan cara melarutkan 0.6 g agarose dalam larutan 0.5 x TBE sebanyak 30 ml kemudian dipanaskan dalam microwave selama lima menit, dikocok menggunakan magnetic stirrer dan ditambahkan 2.5 μl EtBr. Larutan dalam bentuk cair dituangkan ke dalam pencetak gel, sisir ditempatkan di dekat tepian gel dan dibiarkan hingga mengeras. Sisir dicabut setelah gel mengeras sehingga terbentuk sumur-sumur. Tahap selanjutnya gel ditempatkan ke dalam gel tray elektroforesis yang sudah berisi larutan buffer. Produk PCR-RFLP sebanyak 5 μl dicampur dengan 1 μl loading dye kemudian dimasukkan ke dalam sumur-sumur gel. Marker DNA 100 bp sebanyak 2 μl ditaruh ke dalam sumur paling kiri sebagai penanda. Gel dialiri listrik 100 volt selama 30-45 menit. Molekul DNA yang bermuatan negatif pada pH netral akan bergerak ke arah positif (anode). Setelah elektroforesis selesai, gel agarose diambil untuk dilihat panjang pita DNA dengan menggunakan sinar ultraviolet dalam mesin UV Transiluminator.
Pita-pita DNA yang muncul dibandingkan dengan marker untuk diketahui panjang fragmennya dan jumlah pita DNA dari setiap sampel dibandingkan untuk menentukan genotipe pita DNA. Alel T tidak memiliki titik potong dan hanya menunjukkan satu fragmen yang panjangnya sama dengan produk PCR yaitu 290 bp. Alel C memiliki titik potong enzim BsrI (ACTGG|N) dan menunjukkan adanya dua fragmen yang masing-masing memiliki panjang 200 bp dan 90 bp.
Sekuens Fragmen Gen OPN Intron 4
Sekuens fragmen gen OPN intron 4 dilakukan pada individu sapi bali yang bergenotipe TT dengan jumlah dua sampel. Primer yang digunakan untuk sekuens fragmen gen OPN hanya primer forward. Sekuensing gen OPN dilakukan dengan menggunakan mesin sekuenser (ABI Prims 3100-Avant Genetic Analyzer) di perusahaan sekuensing 1st Base, Selangor, Malaysia.
Analisis Data
Frekuensi Alel Gen OPN
Keragaman alel yang terdapat pada populasi sapi bali di BPTU sapi bali, Pulau Bali dianalisis dengan pendekatan frekuensi alel. Frekuensi alel adalah rasio suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi.
nii = jumlah individu bergenotipe ii
nij = jumlah individu bergenotipe ij
5
Sekuens Fragmen Gen OPN Intron 4
Hasil sekuens dianalisis dengan program BioEdit (Hall 1999). Selanjutnya untuk mengetahui kesamaan dengan gen OPN yang terdapat di GenBank dianalisis menggunakan metode BLAST (www.ncbi.nhl.nih.gov./BLAST). Ada tidaknya mutasi pada sekuens fragmen gen OPN intron 4 dilakukan analisis dengan menggunakan program Molecular Evolutionary Genetic Analysis (MEGA5) dengan metode ClustalW (Tamura et al. 2011).
HASIL DAN PEMBAHASAN
Amplifikasi Gen Osteopontin (OPN)
Amplifikasi fragmen gen OPN pada sapi bali dengan menggunakan mesin thermocycler pada suhu annealing 60 oC selama 20 detik menghasilkan panjang produk PCR 290 base pair (bp) (Gambar 2). Sebanyak 100 sampel sapi bali berhasil diamplifikasi dengan tingkat keberhasilan 100%.
Berdasarkan hasil amplifikasi yang dilakukan oleh Leonard et al. (2005) bahwa kondisi annealing fragmen gen OPN|BsrI berlangsung pada suhu touchdown 63 oC ke 50 oC selama 45 detik. Suhu annealing adalah suhu optimum untuk berlangsungnya penempelan primer sesuai dengan sekuens DNA komplementer yang akan diperbanyak selama proses amplifikasi DNA berlangsung. Penelitian ini menggunakan suhu penempelan pasangan primer yang berbeda dengan suhu yang disarankan oleh Leonard et al. (2005). Hal tersebut disebabkan oleh perbedaan kondisi mesin PCR dan campuran komponen pereaksi PCR. Viljoen et al. (2005) menyatakan bahwa suhu annealing berkisar antara 55-72 oC, selain itu suhu optimal annealing salah satunya bergantung pada konsentrasi MgCl2. Pelt-Verkuil et al. (2008) menyatakan bahwa waktu annealing yang dibutuhkan supaya primer dapat berkomplemen dan menempel dengan targetnya bergantung pada kapasitas pemanasan mesin thermocycler yang digunakan, volume campuran PCR serta konsentrasi primer dan gen target.
6
Identifikasi Genotipe Gen Osteopontin (OPN)
Hasil pemotongan dengan enzim BsrI terhadap fragmen gen OPN sapi bali diperoleh hanya satu macam genotipe yaitu genotipe TT (satu pita) dengan panjang 290 bp (Gambar 3). Individu-individu sapi bali yang tidak dapat dipotong fragmen gen OPN-nya berarti mengalami mutasi pada situs pemotong sekuens enzim BsrI.
Berdasarkan hasil penelitian gen OPN dengan situs polimorfik intron 4 pada sapi FH oleh Leonard et al. (2005) diperoleh tiga macam genotipe yaitu genotipe TT, genotipe CC (dua pita), dan genotipe CT (tiga pita). Ketiga genotipe tersebut juga diperoleh dari hasil penelitian Boleckova et al. (2012) pada bangsa sapi Czech Fleckvieh dengan proporsi genotipe CC, CT, dan TT masing-masing sebesar 0.04, 0.28, dan 0.68. Khatib et al. (2009) menyatakan bahwa genotipe TT memiliki angka fertilitas yang lebih tinggi (70%) dibandingkan dengan genotipe CC (62%) pada embrio in vitro fertilization (IVF) sapi.
Frekuensi Alel Gen Osteopontin (OPN)
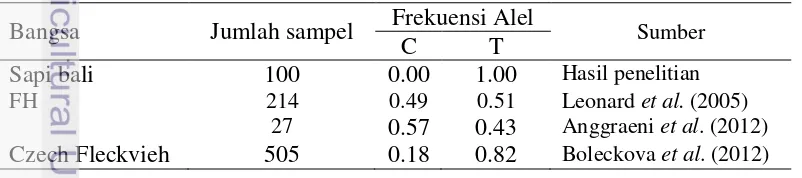
Hasil analisis frekuensi alel gen OPN pada sapi bali menunjukkan bahwa hanya ditemukan satu macam alel yaitu alel T (100%) dan tidak ditemukan alel C. Gen OPN pada sapi bali bersifat seragam atau monomorfik. Nei (1987) menyatakan bahwa suatu alel tergolong polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0.99. Frekuensi alel sapi bali dan beberapa bangsa sapi lainnya disajikan pada Tabel 1.
Tabel 1 Frekuensi alel gen OPN pada sapi bali dan beberapa bangsa sapi
Bangsa Jumlah sampel Frekuensi Alel Sumber
C T
Sapi bali 100 0.00 1.00 Hasil penelitian
FH 214 0.49 0.51 Leonard et al. (2005) 27 0.57 0.43 Anggraeni et al. (2012)
Czech Fleckvieh 505 0.18 0.82 Boleckova et al. (2012)
7 Keragaman genetik atau polimorfisme genetik adalah terdapatnya lebih dari satu bentuk atau macam genotipe di dalam populasi. Sumber keragaman genetik disebabkan oleh adanya pengulangan urutan sekuen, insersi, delesi dan rekombinasi di dalam runutan DNA antar individu, kelompok atau suatu populasi (Nei dan Kumar 2000). Hasil penelitian ini menunjukkan bahwa gen OPN|BsrI pada sapi bali bersifat monomorfik karena hanya ditemukan satu tipe alel yaitu alel T.
Fenomena monomorfisme pada sapi bali juga ditemukan pada beberapa hasil penelitian yaitu gen GHR|AluI (Zulkharnaim et al. 2010) dan gen GH MspI dan AluI (Jakaria et al. 2009; Jakaria dan Noor 2011). Program nasional pemurnian dan peningkatan mutu genetik ternak telah dilakukan pada sapi bali sejak tahun 1976 di Pulau Bali berdasarkan Surat Keputusan (SK) Menteri Pertanian nomor 776/Kpts/Um/12/1976. Kegiatan yang dilakukan oleh Proyek Pengembangan dan Pembibitan sapi bali (P3Bali) adalah pemuliaan sapi bali dengan melakukan seleksi dalam bangsa sehingga diperoleh bibit sapi bali yang bermutu baik (Soehadji 1990). Pemusatan dan pemurnian genetik sapi bali di daerah tertentu dalam jangka waktu yang cukup panjang menyebabkan terjadinya silang dalam. Noor (2008) menyatakan bahwa silang dalam adalah salah satu bentuk isolasi secara genetik karena pada suatu populasi yang terisolasi akan menyebabkan keterbatasan pilihan dalam proses perkawinan. Konservasi di BPTU sapi bali berkaitan dengan pewarisan sifat dari tetua yang menyumbangkan alel homozigot kepada keturunannya sehingga keragaman genotipe pada populasi bersifat monomorfik.
Homologi dan Deteksi Mutasi Gen Osteopontin (OPN)
Hasil analisis sekuens fragmen gen OPN|BsrI menunjukkan bahwa sapi bali memiliki kesamaan yang tinggi terhadap sekuens gen OPN Bos taurus yang terdapat di GenBank (kode akses GU143284) dengan tingkat kesamaan (homologi) yang tinggi yaitu sebesar 100%. Selain itu, ditemukan bahwa terjadi mutasi antara basa C dan T pada sekuens gen OPN intron 4 (Gambar 4, Lampiran 2).
Mutasi transisi basa sitosin (C) menjadi timin (T) menyebabkan perubahan sekuens situs pemotong enzim BsrI ACTGG|N menjadi ATTGG|N sehingga enzim BsrI tidak dapat mengenali situs pemotongnya dan menghasilkan satu pita pada 290 bp. Mutasi transisi adalah mutasi titik yang disebabkan oleh perubahan basa purin menjadi purin (AG, GA) dan basa pirimidin menjadi basa pirimidin (CT, TC) (Brown 2002). Mutasi antara basa pirimidin sitosin dengan timin pada sekuens gen OPN intron 4 Bos taurus berdasarkan GenBank (kode akses GU143824) sesuai dengan mutasi yang terjadi pada seluruh sapi bali di penelitian ini.
8
Gambar 5 Rekonstruksi struktur gen OPN pada sapi dengan SNP (single nuclotide polymorphism) pada intron 4
dijadikan penciri sifat bobot badan dan produksi susu pada sapi Polish HF dan Polish Red, White et al. (2007) yang menyatakan bahwa terdapat alel fungsional pada gen OPN yang mempengaruhi bobot sapih dan bobot lahir sapi, dan Leonard et al. (2005) menyatakan bahwa alel C berhubungan dengan peningkatan kandungan protein dan lemak dalam susu pada sapi FH.
Osteopontin pertama kali dilaporkan terdapat pada jaringan tulang termineralisasi dan protein OPN yang disintesis oleh sel sertoli pada tubulus seminiferus berpotensi terlibat pada adhesi dan migrasi sel (Moura 2005). Khatib et al. (2009) menyatakan bahwa OPN berperan dalam proses binding antara sperma dengan sel telur dan tahap awal perkembangan embrionik, selain itu OPN berpengaruh terhadap kapasitasi sperma in vitro (Monaco et al. 2009). Williams et al. (2011) menyatakan bahwa SNPs pada gen OPN berpotensi sebagai penciri fertilitas sapi pejantan. Struktur gen OPN yang telah dimodifikasi berdasarkan Pareek et al. (2008) dapat dilihat pada Gambar 5.
9
SIMPULAN DAN SARAN
Simpulan
Gen osteopontin (OPN|BsrI) intron 4 pada sapi bali diperoleh hanya satu macam genotipe dan alel yaitu genotipe TT dan alel T (100%). Mutasi yang terjadi pada sapi bali adalah mutasi antara basa sitosin (C) dengan timin (T). Gen OPN intron 4 pada sapi bali yang terdapat di BPTU sapi bali, Pulau Bali bersifat monomorfik.
Saran
Perlu dilakukan penelitian pada sampel sapi bali yang terdapat di daerah pengembangan sapi bali lainnya di Indonesia. Selain itu, diperlukan analisis gen-gen lain yang terkait dengan sifat reproduksi.
DAFTAR PUSTAKA
Anggraeni A, Hasinah H, Arta SA, Tiesnamurti B, Misrianti R, Andreas E. 2012. Genetic variation of the IGF1 and OPN genes in Holstein-Friesian dairy cattle of historical and non-historical twins. [Editor tidak diketahui]. 2nd International Seminar on Animal Industry[Internet]; 2012 Jan 5-6; Jakarta, Indonesia. Jakarta (ID): [penerbit tidak diketahui] hlm 91-96; [diunduh 2013 Apr 5]. Tersedia pada: http//www.repository.ipb.ac.id.
Boleckova J, Matejickova J, Stipkova M, Kyselova J, Barton L. 2012. The association of five polymorphisms with milk production traits in Czech Fleckvieh cattle. Czech J Anim Sci. 57(2): 45–53.
Brown TA. 2002. Genomes 3. 3rd ed. New York (US): Garland Sci.
[DGLS] Directorate Generale of Livestock Services. 2003. National Report on Animal Genetic Resources in Indonesia. Directorate Generale of Livestock Services, Directorate of Livestock Breeding (ID).
Hall, TA. 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp. Ser. 41: 95-98.
Jakaria, Noor RR, Martojo H, Duryadi D, Tappa B. 2009. Identification of growth hormone (Gh) gene MspI and AluI loci polymorphism in beef cattles. The 1st International Seminar on Animal Industry. Bogor, Indonesia.
Jakaria, Noor RR. 2011. Analysis on Alu-I growth hormone (GH Alu-I) gene in bali cattle. J. Indones Trop Anim Agric. 1: 1-6.
10
Leonard S, Khatib H, Schutzkus V, Chang YM, Maltecca C. 2005. Effects of the osteopontin gene variants on milk production traits in dairy cattle. J Dairy Sci. 88: 4083-4086.
Monaco E, Gasparrini B, Boccia L, De Rosa A, Attanasio L, Zicarelli L, Killian G. 2009. Effect of osteopontin (OPN) on in vitro embryo development in cattle. J Theriogenology. 71: 450-457. doi:10.1016/j.theriogenology.2008.08.012. Moura AA. 2005. Seminal plasma proteins and fertility indexes in the bull: the
case for osteopontin. Anim Reprod. 2(1): 3-10.
Nei M. 1987. Molecular Evolutionary Genetics. New York (US): Columbia Univ Pr.
Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. New York (US): Oxford Univ Pr.
Noor RR. 2008. Genetika Ternak. Jakarta (ID): Penebar Swadaya.
Pareek CS, Czarnik U, Pierzchala M, Zwierzchowski L. 2008. An association between the C>T single nucleotide polymorphism within intron IV of osteopontin encoding gene (SPP1) and body weight of growing Polish Holstein-Friesian cattle. J Anim Sci. 26(4): 251-257.
Pelt-Verkuil, van E, Belkum van A, Hays JP. 2008. Principles and Technical Aspects of PCR Amplification. Netherlands (NL): Springer.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular Cloning: A Laboratory Manual. USA (US): CSH Laboratory Pr.
Soehadji. 1990. Kebijaksanaan pemuliaan ternak (breeding policy) khususnya sapi bali, dalam pembangunan peternakan. Seminar Nasional Sapi Bali; 1990 Sep 20-22; Denpasar, Indonesia. hlm A1-A9.
Talib C. 2002. Sapi bali di daerah sumber bibit dan peluang pengembangannya. Wartazoa 12(3): 100-107.
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: Molecular Evolutionary Genetics Analysis using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Molecular Biology and Evolution.
Viljoen GJ, Nel LH, Crowther JR. 2005. Molecular Diagnosis PCR Handbook. Netherlands (NL): Springer.
White SN, Casas E, Allan MF, Keele JW, Snelling WM, Wheeler TL,Shackelford SD, Koohmaraie M, Smith TPL. 2007. Evaluation in beef cattle of six deoxyribonucleic acid markers developed for dairy traits reveals an osteopontin polymorphism associated with postweaning growth. J Anim Sci. 85: 1-10. Williams CL, Lester TD, Rowe MP, Rorie RW. 2011. Effects of osteopontin
single nucleotide polymorphisms on bull semen quality. Ark Anim Sci. 17-21. Zulkharnaim, Jakaria, Noor RR. 2010. Identifikasi keragaman genetik gen
11
LAMPIRAN
Lampiran 1 Sekuens gen osteopontin (OPN) yang diakses di GenBank kode akses GU143824
GenBank: GU143824.1
LOCUS GU143824 291 bp DNA linear MAM 08-NOV-2010 DEFINITION Bos taurus osteopontin gene, intron 4.
ACCESSION GU143824
VERSION GU143824.1 GI:310947851 KEYWORDS .
SOURCE Bos taurus (cattle)
ORGANISM Bos taurus </Taxonomy/Browser/wwwtax.cgi?id=9913> Eukaryota; Metazoa; Chordata; Craniata; Vertebrata; Euteleostomi; Mammalia; Eutheria; Laurasiatheria; Cetartiodactyla; Ruminantia; Pecora; Bovidae; Bovinae; Bos. Technology, Sichuan Agricultural University, Road XinKang 46#, Ya'an, Sichuan 625014, China
variation </nuccore/310947851?from=198&to=198> 198 /gene="osteopontin"
12
ORIGIN
1 gcaaatcaga agtgtgatag acattaactg agctatagtt tctacacatg gataagagag 61 tcaccttttg attatccagg ctaataggga ggtgatttta gttttggggg tgtgcattaa 121 tacatggatt ctctgatccc ctgagaattt tcatttcaaa tagaaaaggt agtctcacaa 181 ttatgtatct gtatttattg gatcattgaa atttggtaaa ttagtgttta ttatgaacaa 241 ggaaaaacag tgtcattgat acaaatatta taactcatac gtttggcttg g //
Lampiran 2 Hasil BLAST nukleotida sekuens gen OPN sapi bali dengan sekuens GenBank kode akses GU143824
13
RIWAYAT HIDUP
Penulis dilahirkan di Bogor, 25 Februari 1991. Penulis adalah anak ketiga dari pasangan I Nyoman Arnaya dan Putu Agustini Eliyati. Penulis menamatkan pendidikan di SMA Negeri 1 Bogor pada tahun 2009 dan pada tahun yang sama penulis diterima menjadi mahasiswa Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor melalui jalur UTMI.