LAMPIRAN
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level DIAMETER LATERAL ROOT
* The mean difference at the 0,05 level
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level
PANJANG LATERAL ROOT
* The mean difference at the 0,05 level
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level
JUMLAH LATERAL ROOT
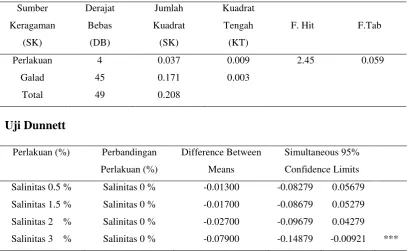
Perlakuan 4 59528.915 14882.228 18.57 <.0001
Galad 45 36060.836 801.351
Total 49 95589.751
Uji Dunnett
JUMLAH TAP ROOT
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level
JUMLAH DAUN
Perlakuan (%) Perbandingan Perlakuan (%)
RASIO AKAR DAN TAJUK
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level
TEBAL DAUN
Perlakuan (%) Perbandingan Perlakuan (%)
LUAS DAUN
Perlakuan (%) Perbandingan Perlakuan (%)
* The mean difference at the 0,05 level
KADAR AIR AKAR
Tabel Anova
Perlakuan (%) Perbandingan Perlakuan (%)
Penanaman Propagul 22 Mei 2015
Pemanenan Semai B. cylindrica 22 Oktober 2015
DAFTAR PUSTAKA
Azwar, S. 2009. Signifikan atau Sangat Signifikan?. Fakultas Psikologi. UGM
Basyuni, M., Lollie, A, P, P., Berliana, N dan Putri, E. 2014. Growth and Biomass in Response to Salinity and Subsequent Fresh Water in Mangrove
Seedlings Avicennia marina and Rhizophora stylosa. JMHT Vol XX
(1): 17-25.
Bohm. W., 1979. Methods of Studying Root Systems. Springer - Verlag Berlin
Heidelberg New York. Ecological Studies 33.
BPS. 2010. Statistik Indonesia. Sumatera Utara.
Davies, J. & G. Claridge. 1993. Wetland Benefits. The Potential for Wetlands toSupport and Maintain Development. Asian Wetland Bureau,
InternationalWaterfowl & Wetlands Research Bureau, Wetlands for the America’s, hal 45.
Easlon, Hsien Ming dan Arnold Bloom. 2014. Easy Leaf Area: Automated Digital
Image Analysis For Rapid And Accurate Measurement of Leaf Area.
Application in Plant Sciences. Botanical Society of America.
Fofonoff, N.P., Lewis, E.L.1979. A practical salinity scale. J. Oceanografi. 35, 63–64
Giesen, W. & B. van Balen. 1991. Several Short Surveys of Sumatran
Wetlands.Notes and Observations. Laporan Proyek PHPA/AWB Sumatra
Wetlands No.26, hal 98.
Giri C, E.Ochieng, L.L. Tieszen, Z. Zhu, A. Singh, T. Loveland, J. Masek dan Duke. 2011. Status ang Distribution of Mangrove Forests of The World Using Earth Observation Satellite Data. Global Ecology and
Biogeography (2011) 20. 154 - 159.
Harahab, N. 2009. Pengaruh Ekosistem Hutan Mangrove Terhadap Produksi Perikanan Tangkap (Studi Kasus Di Kabupaten Pasuruan). Jawa Timur. Jurnal Perikanan. Universitas Brawijaya. Malang.
Heddy, S. 2001. Ekofisiologi Tanaman. PT Raja Grafindo Persada. Jakarta. Hutahean, Eben., Cecep Kusmana., Helmy Ratna Dewi. 1999. Studi Kemampuan
Tumbuh Anakan Mangrove Jenis Rhizopora mucronata, Bruguiera gymnorrhiza, dan Avicennia marina Pada Berbagai Tingkat Salinitas. Jurnal Manajemen Hutan Tropika Vol. V, No. 1 : 77 – 85 (1999)
Hutching, P. dan P. Saenger, 1987. Ecology of Mangrove. University of Queensland Press, St. Lucia. Australia.
Kantor Menteri Negara Lingkungan Hidup. 1993. Pengelolaan Ekosistem Hutan Mangrove. Prosiding Lokakarya Pemantapan Strategi Pengelolaan LingkunganWilayah Pesisir dan Lautan dalam Pembangunan Jangka Panjang Tahap Kedua.Kapal Kerinci, 11-13 September 1993, hal 47.
Keliat, D A. 2016. Pertumbuhan dan Perkembangan Akar Semai Rhizopora Apoculata. Blume Pada Berbagai Salinitas. Skripsi. Universitas Sumatera Utara. Medan.
Klepper, B. 1991. Root-shoot relationships, p:265-286. In Waisel et al., 1991.
Plant roots the hidden half Marcel Dekker Inc. New York. 948.
Komiyama, A., H. Moriya, S. Prawiroatmodjo, T. Tomi & K. Ogino. 1988. Forest as an Ecosystem, Its Structure and Function; #1: Floristic Composition and Stand Structure. Dalam Biological System of Mangroves. Laporan Ekspedisi Mangrove Indonesia Timur tahun 1986, Ehime University, Japan. Hal. 85-96.
Kridalaksana, A. dan Suryanto, A. 2014. Pengelolaan Tambak dan Mangrove di Area Pertambakan di Desa Mororejo. Kecamatan Kaliwungu. Kabupaten Kendal. Diponegoro. Journal of Maquares. Management Of Aquatic Resources.
Lubis, M U. 2016. Pertumbuhan Dan Perkembangan Akar Semai Mangrove Sejati Ceriops Tagal (Perr.) C.B.Rob Pada Berbagai Salinitas. Skripsi. Univesitas Sumatera Utara. Medan.
Pasaribu, Gripsy Yati Handayani. Pertumbuhan Propagul Bakau Putih
(B.cylindrica) Setelah Proses Pemeraman. Skripsi. Universitas Sumatera Utara. Medan
Pessarakli, M. 1993. Handbook of Plan and Crop Stress. Marcel Dekker Inc. New York.
Pi. N,N.F.Y Tam, Y. Wu, M.H. Wong. 2009. Root Anatomy and Spatial Pattern
of Radial Oxygen Loss if Eight True Mangrove Species. Aquatic Botany
90 222-230.
Ramayani. 2012. Pengeruh Salinitas Terhadap Pertumbuhan Dan Biomassa Semai Dan Kandungan Lipida Pohon Non-Sekresi Ceriops tagal. Skripsi
Universitas Sumatera Utara
Rost, Thomas L. 1996. Root Architecture. Section of Plant Biology Division of Biological Sciences. University of California.
Departemen Kehutanan Royal. 1997. Growth Development of Bruguiera cylindrica BL. Seedlings. Bangkok
Salisbury, F.B dan Cleon W. Ross. 1995. Fisiologi Tumbuhan. Jilid I. ITB. Bandung. hal. 67-72.
Samingan, M.T. 1980. Notes on the Vegetation of the Tidal Areas of South Sumatra, Indonesia, with Special Reference to Karang Agung. Dalam International Social Tropical Ecology, Kuala Lumpur. Hal. 1107-1112.
Saparinto, C. 2007. Pendayagunaan Ekosistem mangrove. Dahara Prize. Semarang.
Schuurman, J.J., and M.A. J.Goedewaagen. 1971. Methods for the Examination of Root Systems and Roots. Wageningen. The Netherlands.
Simbolon, C R H. 2013. Pengaruh Intensitas Naungan Terhadap Pertumbuhan Bakau Putih (Bruguiera cylindrica) di Sicanang Medan Belawan. Skripsi. Universitas Sumatera Utara. Medan.
Sunyoto, D. 2012. Dasar-Dasar Statiska Ekonomi. CAPS. Yogyakarta
Van Steenis, C.G.G.J. 1958. Ecology of Mangroves. Introduction to account of the Rhizophoraceae by Ding Hou, Flora Malesiana, Ser. 1(5) : 431-441
Yusmaini, F. dan T.K.Suharsi. 2008. Pengaruh Jenis Bahan Stek dan
Penyungkupan terhadap Keberhasilan Stek Stevia (Stevia rebaudiana
Bertoni M.). Makalah Seminar Departemen Agronomi dan Hortikultura. IPB. Bogor.
Wong, Yee-Yen., Ho, Chai-ling., Nguyen, Phuoc Dang., Teo, Swee-Sen.,
Harikrishna, Jennifer Ann., Rahim, Raha Abdul., Wong, Michael C. V.L. 2007. Isolation of salinity tolerant genes from the mangrove plant,
METODOLOGI PENELITIAN
Waktu dan Tempat
Penanaman propagul B. cylindrica dengan perlakuan berbagai variasi salinitas selama 5 bulan dilakukan pada Mei 2015 sampai Oktober 2015 di rumah
kaca Fakultas Pertanian, Universitas Sumatera Utara.
Lokasi Pengambilan Sampel
Sampel penelitian berupa propagul B. cylindrica diambil dari hutan mangrove di Pulau Sembilan. Pulau Sembilan merupakan nama salah satu desa yang berada di gugusan pulau-pulau di Kabupaten Langkat. Desa Pulau Sembilan
berdekatan dengan Selat Malaka dan merupakan salah satu tujuan wisata utama di Kabupaten Langkat. Pulau Sembilan secara administrasi terletak di kecamatan
Pangkalan, Susu Kabupaten Langkat. Luas Pulau Sembilan 24,00 km2 atau 8.84% dari total luas kecamatan Pangkalan Susu. Di Pulau ini terdapat hutan mangrove yang mengelilingi pulau dan tumbuh ekosistem pesisir. Kondisi air tanah masih
cukup baik dimana tidak ditemukan adanya air sumur yang asin atau terkena intrusi air laut (BPS, 2010)
Bahan dan Alat
Bahan yang digunakan dalam penelitian adalah propagul B. cylindrica
yang sehat dan matang, bubuk garam komersial (marine salt), air tawar, pasir dari
botol plastik, cutter, jangka sorong, penggaris, ember, kamera, timbangan aplikasi
SAS 9.1, software image J, SPSS versi 22, dan alat tulis.
Prosedur Penelitian
1. Persiapan Media Tanam
Pasir disterilisasi untuk membunuh bakteri dan jamur yang hinggap kemudian diisikan pada botol plastik sesuai jumlah masing-masing perlakuan dan
ulangan. Propagul B. cylindrica ditanam dan diberi salinitas yang bervariasi 0%, 0,5%, 1,5%, 2% dan 3% (sama dengan tingkatan air laut yaitu 15%, 45%, 60%, 90%).
Di dalam penelitian ini, salinitas ditentukan dari perbandingan massa bubuk garam dengan massa larutan. Metode ini berdasarkan Fofonoff dan Lewis
(1979) dimana jenis garam yang dipakai adalah bubuk garam komersial (marine salt). Untuk membuat konsentrasi salinitas 0%, 0,5%, 1,5%, 2% dan 3% dibuat dengan melarutkan 5,66 g, 17 g, 22,6 g, dan 34 g bubuk garam komersial untuk 1
liter air. Salinitas adalah massa serbuk garam/massa larutan. Konsentrasi garam pada setiap perlakuan pot diperiksa seminggu sekali selama percobaan dengan
hand refraktometer.
2. Pemilihan Propagul
Propagul B. cylindrica yang digunakan berasal dari pohon induk yang
berumur 5 tahun atau lebih. Propagul yang dipilih sebaiknya telah matang secara fisiologi dengan warna propagul hijau kecoklatan dan sehat, tidak terserang oleh
hama dan penyakit.
Propagul B. cylindrica yang telah diseleksi ditanam ke dalam botol plastik
berisi media tanam yang telah disesuaikan dengan perlakuannya persentase salinitas masing-masing.
Analisis Data
Penelitian ini menggunakan metode analisis data Rancangan Acak Lengkap (RAL) dengan 5 perlakuan konsentrasi garam (salinitas) berdasarkan
salinitas yang ada di lapangan dengan masing-masing 5 ulangan : a. Salinitas 0 %
b. Salinitas 0,5 %
c. Salinitas 1,5 % d. Salinitas 2 %
e. Salinitas 3 %
Model linear RAL:
Yij = μ + τi + εij
Keterangan:
Yij = hasil pengamatan pada perlakuan ke-i dan ulangan ke-j
μ = nilai rataan umum (mean)
τi = pengaruh faktor perlakuan ke-i
εij = pengaruh galat perlakuan ke-i ulangan ke-j
Analisis data dilakukan dengan metode ANOVA (Analisis of Varians) satu
arah menggunakan uji Dunnett untuk perbandingan seluruh perlakuan salinitas yang diberikan terhadap kontrol. Nilai P < 0,05 dan P < 0,01 dipakai sebagai batas
untuk menunjukkan pengaruh perlakuan. Uji statistik dilakukan memakai software SAS versi 9.1. Korelasi parameter menggunakan SPSS versi 22.
Parameter Pengamatan
Pengamatan dilakukan 5 bulan setelah tanam dan parameter yang diamati adalah:
1. Persentase Hidup (%)
Persentase hidup dihitung dengan membandingkan antara jumlah semai yang hidup dan jumlah biibit yang ditanam pada awal penelitian. Pengambilan
data dilakukan pada akhir pengamatan (Yusmaini dan Suharsi, 2008)
Persen Hidup (%) = x 100%
2. Mortalitas (%)
Kematian semai dihitung dengan membandingkan antara jumlah semai yang mati dan jumlah semai yang ditanam pada awal penelitian. Pengambilan data
dilakukan pada akhir pengamatan setelah 5 bulan
Mortalitas (%) = x 100%
3. Tinggi semai (cm)
Pengukuran tinggi semai menurut dilakukan dengan menggunakan penggaris. Pengukuran dilakukan mulai dari bagian plumula sampai titik tumbuh tertinggi semai B. cylindrica. Pengukuran tinggi dilakukan setelah pemanenan.
4. Diameter semai (cm)
Jumlah semai yang hidup Jumlah propagul ditanam
Pengukuran diameter batang semai berdasarkan dilakukan dengan
menggunakan jangka sorong. Pengukuran dilakukan setelah pemanenan. Diameter diukur tiga kali mulai dari bagian plumula, pertengahan batang, sampai titik
tumbuh tertinggi kemudian dihitung rata – ratanya B.cylindrica
5. Jumlah akar (cm)
Perhitungan jumlah akar dilakukan secara manual setelah pemanenan
semai B. cylindrica umur 5 bulan. Jumlah akar dihitung berdasarkan kedudukan akar pada sistem perakaran (tingkat percabangan) menurut klasifikasi Pi dkk
(2009), yang terdiri dari tap root dan lateral root. 6. Panjang akar
Pengukuran panjang akar dilakukan secara manual dengan menggunakan
mistar dan benang. Pengukuran panjang dilakukan setelah pemanenan semai
B. cylindrica pada 5 bulan. Panjang akar diukur berdasarkan kedudukan akar pada
sistem perakaran (tingkat percabangan) menurut klasifikasi Pi dkk (2009).
7. Diameter akar (cm)
Pengukuran diameter dilakukan setelah pemanenan semai B. cylindrica
pada 5 bulan. Hasil dari diameter akar dapat memberikan informasi penting hubungannya dengan ukuran pori tanah dan potensial penetrasi akar (Bohm, 1979). Pengukuran diameter akar dilakukan pada setiap tipe percabangan dengan
menggunakan jangka sorong Pi dkk (2009). 8. Jumlah Daun
Berat Kering Tajuk Berat Kering Akar 9. Luas Daun (cm2)
Pengukuran luas daun dilakukan pada akhir pengamatan data. Perhitungan luas daun menggunakan program komputer. Untuk melakukan perhitungan
terlebih dahulu daun digambar di kertas millimeter blok yang selanjutnya dilakukan scanning pada gambar tersebut. Setelah di pindai maka gambar tersebut dihitung dengan program image J sesuai penelitian oleh Easlon dan Bloom (2014)
bahwa hasil penghitungan perangkat ini akurat, software dapat diunduh secara bebas, dan menghasilkan data dalam waktu singkat. Jika dibandingkan dengan
beberapa metode yang ada maka Software Image J merupakan perangkat terbaik dalam melakukan penghitungan jumlah luas daun.
10. Tebal Daun (mm)
Pengukuran tebal daun dilakukan di akhir pengamatan dengan menggunakan mikrometer sekrup digital.
11. Kadar Air Tajuk dan Kadar Air Akar
Perhiungan persentase kadar air tajuk dan kadar air akar menggunakan rumus sebagai berikut:
Kadar Air (%) = x 100%
12.Rasio Tajuk dan Akar
Perhitungan rasio tajuk dan akar dilakukan pada akhir pengamatan. dengan menggunakan rumus sebagai berikut:
Rasio =
HASIL DAN PEMBAHASAN
Pengaruh Salinitas Terhadap Pertumbuhan Semai B. cylindrica
Karakteristik dan pertumbuhan akar semai B. cyilindrica yang berumur 5
bulan pada berbagai variasi salinitas di rumah kaca disajikan pada gambar di bawah ini:
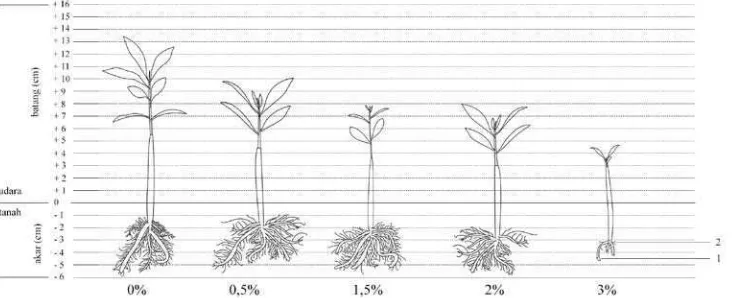
Gambar 1. Karakteristik dan Pertumbuhan Akar Semai B. cylindrica Umur 5 bulan di Rumah Kaca. Keterangan : 1, tap root; 2, lateral root
Berdasarkan Gambar 1 terlihat perbedaan pertumbuhan semai
B. cylindrica dari berbagai konsentrasi salinitas. Hasil yang diperoleh pada
Gambar 1 menunjukkan bahwa persentase salinitas terhadap pertumbuhan dan
perkembangan akar semai mangrove B. cylindrica menunjukkan perbedaan yang nyata pada setiap pemberian salinitas pada umur 5 bulan. Rost (1996) menyatakan bahwa sistem perakaran taproot pada flora umumnya ditemukan pada dikotil,
yang tersusun dari pusat perakaran dengan ukuran lebih besar disebut dengan tap root atau akar utama. Diameter tap root lebih besar daripada lateral root atau akar
kemudian bercabang dari tap root. Akar B. cylindrica diklasifikasikan menjadi 2
bagian, yaitu tap root dan lateral root (Pi dkk, 2009).
Persentase hidup dan mortalitas semai B. cylindrica di sajikan pada Tabel 1. Tabel 1. Persentase hidup dan mortalitas semai B. cylindrica
No. Perlakuan Persentase hidup (%) Mortalitas (%)
1. Salinitas 0 % 100 -
2. Salinitas 0,5% 100 -
3. Salinitas 1,5% 100 -
4. Salinitas 2 % 100 -
5. Salinitas 3 % 100 -
Tabel 1 menunjukkan persentase hidup semai B. cylindrica pada setiap
salinitas adalah 100 %. Sementara tidak ada semai yang mati pada penelitian yang
dilakukan. Sesuai dengan pernyataan Noor (2006) bahwa kadar salinitas optimum
untuk Bruguiera adalah < 2,5%. Jenis B. parviflora tumbuh optimum pada kadar
salinitas 2,0% sementara B. gymnorrhiza pada salinitas 1,0 – 2,5%. Hasil pada
penelitian ini menunjukkan bahwa B. cylindrica dapat tumbuh pada kadar
salinitas > 2,5% hanya saja pertumbuhannya agak lambat dan kurang optimal
yang disebabkan oleh berbagai faktor lingkungan. Faktanya mangrove memiliki
kemampuan dalam beradaptasi pada lingkungan dengan kadar salinitas tinggi
maupun rendah. B. cylindrica dapat tumbuh dalam salinitas rendah ataupun tinggi.
Pertumbuhan semai B. cylindrica mencakup tinggi dan diameter.
Parameter ini merupakan visual yang terlihat jelas pada percobaan yang dilakukan
seperti tersaji pada gambar 2
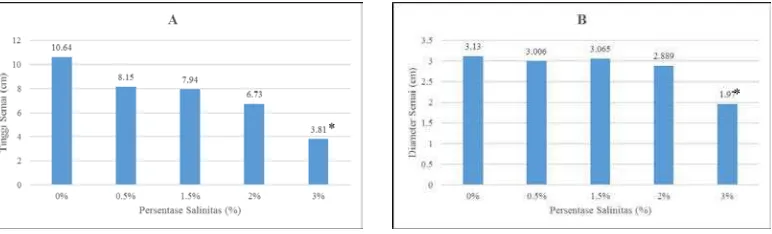
Gambar 2. Respons tinggi semai (cm) B. cylindrica terhadap salinitas (A); Respons
diameter semai (mm) B. cylindrica terhadap salinitas (B). Tanda (*)
mengindikasikan secara statistik signifikan dari kontrol (0%) sampai (3%) pada P<0,01 dan P>0,05 dengan Uji Dunnet
Respons pertumbuhan tinggi dan diameter semai B. cylindrica umur 5
bulan terhadap salinitas terlihat pada gambar 2A. Pertumbuhan semai tertinggi
adalah pada salinitas 0% yaitu sebesar 10,64 cm sementara yang terendah adalah
sebesar 3,81 pada salinitas 3%. Hasil ini menunjukkan bahwa semai B. cylindrica
yang ditanam selama 5 bulan di rumah kaca tumbuh dengan baik. Perbedaan
konsentrasi salinitas jelas memberikan pengaruh yang berbeda terkait adaptasi
mangrove. Data ini merupakan penemuan baru untuk melengkapi data
petumbuhan tinggi semai jenis-jenis mangrove sehingga menjadi acuan untuk
penelitian yang lain. Penelitian serupa yang dilakukan Ramayani (2012) dengan
sampel mangrove C. tagal menunjukkan hasil semai tertinggi berada pada kadar
salinitas 0,5%. Pemberian salinitas berpengaruh signifikan secara statistik
dibandingkan dengan kontrol pada P<0,05 dengan uji Dunnet pada perlakuan
salinitas 3%. Diameter batang semai seperti terlihat pada gambar 2B yang paling
besar pertumbuhannya adalah 3,13 mm pada salinitas 0% dan diameter pada
salinitas 3% adalah yang terendah yaitu sebesar 1,97 mm. Uji Dunnet
menunjukkan bahwa pemberian salinitas berpengaruh signifikan dengan kontrol
pada salinitas 3%. Fenomena diakibatkan karena konsentrasi garam yang yang tinggi. Kondisi seperti ini dapat menyebabkan semai menjadi stress sehingga
terhambat pertumbuhannya.
Tingkat salinitas terhadap pertumbuhan tinggi dan diameter semai B.
dilakukan Departemen Kehutanan Royal (1997) bahwa pertumbuhan diameter B.
cyindrica tidak berbeda jauh dan tinggi yang mana akan berkorelasi dengan
perakaran. Pertumbuhan tinggi dan diameter semai B. cyindrica terhambat akibat
tingginya konsentrasi garam sehingga tidak mampu untuk mentoleransi garam yang diserap. Hal ini sesuai dengan pernyataan Salisbury (1995) bahwa setiap jenis organisme mempunyai tingkat toleransi yang berbeda terhadap faktor
lingkungan termasuk terhadap konsentrasi salinitas garam yang tinggi.
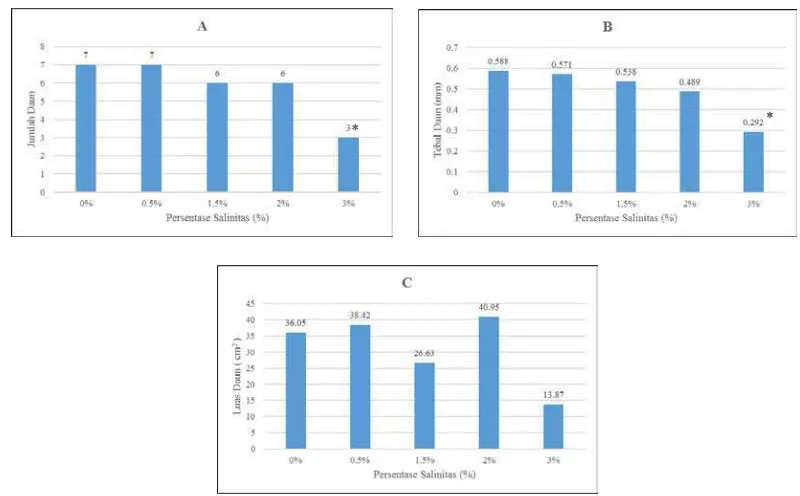
Gambar 3. Respons jumlah daun semai B. cylindrica terhadap salinitas(A); Respons
tebal daun semai B. cylindrica terhadap salinitas (B); Respons luas daun semai B. cylindrica terhadap salinitas (C). Tanda (*) mengindikasikan secara statistik signifikan dari kontrol (0%) sampai (3%) pada P<0,01 dan P>0,05 dengan Uji Dunnet
Jumlah daun paling banyak sesuai gambar 3A pada penelitian ini adalah
pada salinitas 0% dan 0,5% sebanyak 7 helai daun. Jumlah daun dengan
pemberian salinitas 3% adalah yang paling sedikit dengan hasil hanya 3 daun.
Pemberian salinitas berpengaruh signifikan secara statistik dianding salinitas 0%
pada P<0,05 dengan uji Dunnet pada perlakuan salinitas 3%. Nilai daun yang
merupakan hasil dari perlakuan salinitas 0% sementara daun yang paling tipis
bernilai 0,292 mm yaitu hasil perlakuan salinitas 3%. Pemberian salinitas
berpengaruh signifikan secara statistik dianding salinitas 0% pada P<0,05 dengan
uji Dunnet pada perlakuan salinitas 3%. Perhitungan luas daun menggunakan software imageJ menghasilkan daun yang paling luas adalah pemberian salinitas dengan kadar salinitas 2 % yaitu 40,95 cm2. Daun yang luasnya paling kecil
adalah hasil perlakuan salinitas dengan kadar 3% yaitu 13,87 cm2. Pemberian salinitas tidak berpengaruh signifikan secara statistik dianding salinitas 0% pada
P<0,05 dengan uji Dunnet.
Perhitungan jumlah daun, tebal daun, dan luas daun menunjukkan bahwa
semakin tinggi tingkat salinitas semakin rendah pula jumlah, tebal, dan luas daun
yang diperoleh. Struktur dari suatu tanaman yaitu ukuran daun yang lebih kecil dipengaruhi oleh salinitas, sehingga kurangnya penyerapan hara dan air yang akan menghambat laju fotosintesis. Pada akhirnya tanaman tersebut akan tumbuh
menjadi abnormal. Hal ini sesuai dengan penelitian Basyuni dkk (2014) yang menyatakan bahwa tingkat salinitas akan mempengaruhi jumlah daun dari semai
R. stylosa dan A. marina dimana semakin tinggi salinitas maka semakin sedikit jumlah daun. Harjadi dan Yahya (1988) menyatakan pengaruh salinitas terhadap pertumbuhan dan perubahan struktur tanaman yaitu lebih kecilnya ukuran daun.
Sehingga penyerapan hara dan air yang berkurang akan menghambat laju fotosintesis yang pada akhirnya akan menghambat pertumbuhan tanaman.
Pertumbuhan akar merupakan pertumbuhan tanaman pada bagian di dalam
tanah. Penelitian yang dilakukan terbukti bahwa salinitas berpengaruh terhadap
akar B.cylindrica. Pertumbuhan akar semai B. cylindrica mencakup jumlah tap
root jumlah lateral root, panjang tap root, panjang lateral root, diameter tap root
dan diameter lateral root seperti yang dapat dilihat pada gambar 4 di bawah ini:
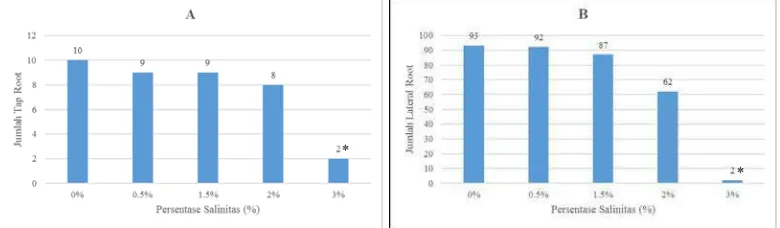
Gambar 4. Respons pertumbuhan jumlah tap root semai B. cylindrica terhadap salinitas
(A); Respons pertumbuhan jumlah lateral root semai B. cylindrica terhadap
salinitas (B). Tanda (*) mengindikasikan secara statistik signifikan dari 0%
sampai 3% pada P<0,01 dan P>0,05 dengan uji Dunnet
Jumlah tap root semai yang paling tinggi seperti yang terlihat pada gambar 4A ditunjukkan pada salinitas 0%, yaitu 10 tap root dan jumlah tap root semai
yang paling sedikit atau terdapat pada perlakuan salinitas 3% yaitu hanya 1 tap root. Pemberian salinitas berpengaruh signifikan secara statistik dibandingkan
salinitas 0% (kontrol) pada P<0,05 dengan uji Dunnet pada salinitas 3%. Sedangkan hasil lain yang diperoleh untuk jumlah lateral root dengan jumlah tertinggi adalah pada tingkat salinitas 0% yaitu sebanyak 93 akar lateral dan
perlakuan salinitas 3% menghasilkan akar lateral yang paling sedikit yaitu 2 akar lateral. Pemberian salinitas berpengaruh signifikan secara statistik dibandingkan
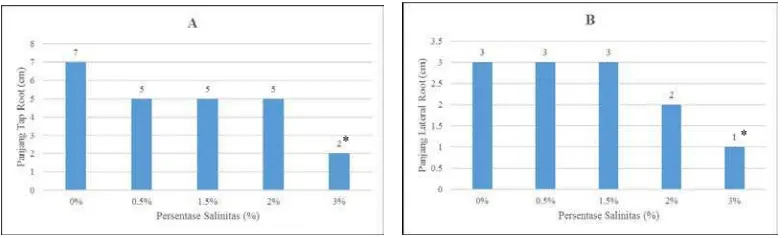
Gambar 5. Respons pertumbuhan panjang tap root semai B. cylindrica terhadap salinitas
(A); Respons pertumbuhan panjang lateral root semai B. cylindrica terhadap
salinitas (B). Tanda (*) mengindikasikan secara statistik signifikan dari 0%
sampai 3% pada P<0,01 dan P>0,05 dengan uji Dunnet
Panjang tap root semai tertinggi pada gambar 5A di tunjukkan oleh salinitas 0%, yaitu 7 cm dan yang terendah pada salinitas 3% yaitu 2 cm.
Pemberian salinitas berpengaruh signifikan secara statistik dibandingkan salinitas 0% (kontrol) pada P<0,05 dengan uji Dunnet pada salinitas 3%. Sedangkan untuk lateral root yang paling panjang terdapat pada tingkat salinitas 0%; 0,5%; 1,5%
yaitu 3 cm dan terendah atau lateral root yang paling pendek adalah pada perlakuan salinitas 3% yaitu sepanjang 1 cm. Pemberian salinitas berpengaruh
signifikan secara statistik dibandingkan salinitas 0% pada P<0,05 dengan uji
Dunnet’s pada perlakuan salinitas 3%.
Gambar 6. Respons pertumbuhan diameter tap root semai B. cylindrica terhadap salinitas
(A); Respons pertumbuhan diameter lateral root semai B. cylindrica terhadap
salinitas (B). Tanda (*) mengindikasikan secara statistik signifikan dari 0%
sampai 3% pada P<0,01 dan P>0,05 dengan uji Dunnet
terendah pada salinitas 3% yaitu 1,5 mm. Pemberian salinitas berpengaruh
signifikan secara statistik dibandingkan salinitas 0% (kontrol) pada P<0,05 dengan uji Dunnet pada salinitas 3%. Sedangkan untuk lateral root yang paling
panjang terdapat pada tingkat salinitas 0% yaitu 0,206 mm dan terendah pada salinitas 3% yaitu 0,127 mm. Pemberian salinitas berpengaruh signifikan secara statistik dibandingkan salinitas 0% pada P<0,05 dengan uji Dunnet’s pada
perlakuan salinitas 3%.
Hasil perhitungan jumlah, panjang, dan diameter akar semai B. Cylindrica
menunjukkan hubungannya dengan salinitas berbanding terbalik. Artinya tingkat salinitas yang tinggi akan mengurangi dan menghambat pertumbuhan akar. Sebaliknya salinitas yang rendah akan mendukung pertumbuhan akar semai.
meskipun demikian pemberian salinitas juga memberikan dampak yang signifikan terhadap pertumbuhan akar. Sesuai dengan pernyataan Pessarakli (1993) bahwa cekaman salinitas menyebabkan jumlah air pada tanaman semakin berkurang.
Stress air terus-menerus dimungkinkan dapat meningkatkan produksi metabolit sekunder di daun dan akar B. cylindrica. Noor dkk (2006) juga menegaskan
untuk salinitas <2,5% petumbuhan B. cylindrica lebih optimal termasuk bagian akar. Faktor lain juga dapat mempengaruhi seperti penelitian Simbolon (2013) adalah intensitas naungan pada saat persemaian yang menyatakan intensitas
naungan terbaik untuk persemaian B. cylindrica adalah 50%.
Penelitian Keliat (2016) dengan spesies mangrove R. Apiculata
sedikit dan diameter akar yang besar begitu sebaliknya jika salinitasnya rendah.
Jumlah akar mangrove sangat dipengaruhi oleh lokasi tempat tumbuh serta dapat merupakan indikasi dari kesesuaian mangrove terhadap tempat tumbuhnya.
Respons Salinitas Terhadap Biomassa Semai B. cylindrica.
Penelitian yang dilakukan selama 5 bulan dengan lokasi persemaian di rumah kaca menunjukkan adanya pengaruh tingkat salinitas terhadap biomassa
semai B. Cylindrica. Hasil ini merupakan data yang diperoleh berdasakan perhitungan kadar air akar, kadar air tajuk, rasio tajuk dan akar dapat di lihat pada
Gambar 7 dan Gambar 8 yang tertera di bawah ini:
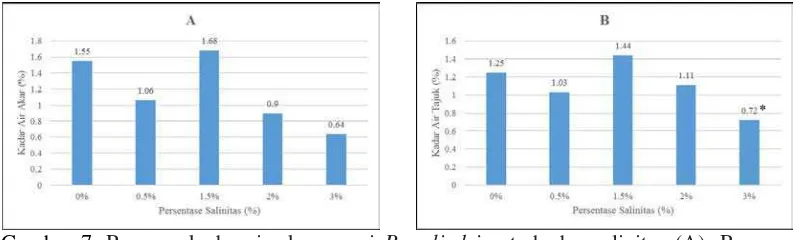
Gambar 7. Respons kadar air akar semai B. cylindrica terhadap salinitas (A); Respons
kadar air tajuk semai B. cylindrica terhadap salinitas (B). Tanda (*)
mengindikasikan secara statistik signifikan dari 0% sampai 3% pada P<0,01 dan P>0,05 dengan uji Dunnet
Gambar 7A menunjukkan kadar air akar semai B. cylindrica tertinggi adalah pada salinitas 1,5% yaitu 1,68% dan yang terendah pada salinitas 3% yaitu
0,64%. Pemberian salinitas tidak berpengaruh signifikan secara statistik dibandingkan salinitas 0% pada P<0,05 dengan uji Dunnet pada perlakuan. Pada Gambar 7B untuk kadar air tajuk tertinggi adalah pada tingkat salinitas 1,5%
Gambar 8. Respons rasio akar dan tajuk semai B. cylindrica terhadap salinitas. Tanda (*) mengindikasikan secara statistik signifikan dari 0% sampai 3% pada P<0,01 dan P>0,05 dengan uji Dunnet
Gambar 8 menunjukkan ratio tajuk dan akar semai B. cylindrica tertinggi di tunjukkan pada salinitas 3% yaitu 3,01 dan yang terendah pada salinitas 0,5% yaitu 1,41. Salinitas berpengaruh signifikan secara statistik dibandingkan salinitas
0% (kontrol) pada P<0,05 dengan uji Dunnet pada perlakuan 2% dan 3%.
Rasio tajuk dan akar menunjukkan adanya pengaruh yang sejalan yaitu
pertumbuhan semai di atas tanah, yaitu daun pada semai B. cylindrica sejalan dengan pertumbuhan organ di dalam tanah yaitu, akar. Penelitian serupa dilakukan oleh Lubis (2016) dengan sampel mangrove C. tagal menunjukkan
hasil bahwa perbandingan tajuk akar mempunyai pengertian bahwa pertumbuhan suatu tanaman diikuti dengan pertumbuhan bagian tanaman lainya, dimana tajuk
akan meningkat secara ratio tajuk akar mengikuti peningkatan berat akar.
Menurut Basyuni dkk (2014) dengan sampel semai mangrove R. stylosa
semakin tinggi salinitas maka rasio akar dan tajuk semakin kecil. Namun pada
penelitian ini dengan sampel B. cylindrica tingkat salinitas yang tinggi justru menghasilkan rasio terendah. Hal ini disebabkan kemampuan setiap tanaman yang
organ tanaman dalam mempertahankan keseimbangan fisiologis, sehingga
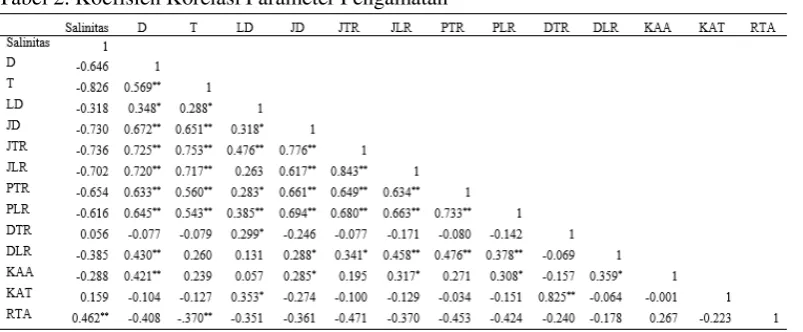
masing-masing organ tanaman dapat melakukan fungsinya secara normal. Tabel 2. Koefisien Korelasi Parameter Pengamatan
Keterangan: D: diameter, T: Tinggi Semai, LD: Luas Daun, JD: Jumlah Daun, JTR: Jumlah Tap Root, JLR: Julah Lateral Root, PTR: Panjang Tap Root, PLR: Panjang Lateral Root, DRT: Diameter Tap Root, DLR: Diameter Lateral Root, KAA: Kadar Air Akar, KAT: Kadar
Air Tajuk, RTA: Rasio Tajuk dan Akar. Rasio akar dan tajuk. Tanda (*) mengindikasikan secara statistik korelasi yang
signifikan pada P<0,05 dan tanda (**) mengindikasikan secara statistik sangat signifikan pada P<0,01.
Analisis korelasi digunakan untuk mengetahui keeratan hubungan antara
dua variabel (Harahab, 2009). Penggunaan analisis korelasi adalah untuk mengukur tinggi rendahnya derajat hubungan antara variabel yang diteliti. Tinggi rendahnya derajat hubungan antara variabel yang diteliti tersebut dapat dilihat dari
koefisien korelasi. Koefisien korelasi mendekati angka +1 mengindikasikan terjadi hubungan positif yang erat, namun apabila mendekati angka –1
mengindikasikan terjadi hubungan negatif yang erat. Koefisien korelasi mendekati angka 0 (nol) mengindikasikan bahwa hubungan kedua variabel adalah lemah atau tidak erat. Dengan demikian nilai koefisien korelasi berada pada–1 ≤ r ≤ +1
(Kridalaksana dan Suryanto, 2014). Analisis korelasi bertujuan untuk mengetahui nilai dari keeratan hubungan antara masing-masing parameter. Selanjutnya akan
diketahui keeratan hubungan antara parameter dengan parameter yang lain.
dan dianggap tidak ada, berapa besarnya pun nilai tersebut. Signifikan berarti nilai
statistik tidak dapat diabaikan dan harus dianggap ada, berapa kecilnya pun nilai statistik tersebut (Azwar, 2009).
Analisis koefisien korelasi menunjukkan perlakuan salinitas yang diberikan berkorelasi positif terhadap tinggi, jumlah taproot, jumlah lateral root, panjang tap root, panjang lateral root, diameter lateral root, dan kadar air akar semai
B.cylidrica. Hal ini dilihat dari koefisien korelasi yang bernilai positif. Perlakuan salinitas berkorelasi negatif terhadap rasio tajuk dan akar semai B.cylidrica. Nilai
koefisien korelasi menunjukkan kekuatan korelasi yang lemah negatif, sehingga hubungan variabel parameter rasio tajuk dan akar tidak terlalu sensitif terhadap perubahan yang terjadi pada variabel salinitas (Sunyoto, 2012).
Supriharyono (2000) menyatakan bahwa spesies mangrove dapat tumbuh pada salinitas yang ekstrim atau sangat tinggi, namun biasanya pertumbuhannya kurang baik atau pendek-pendek. Heddy (2001) melaporkan bahwa analisis
pertumbuhan tanaman hanya dapat memberikan sedikit informasi tentang proses-proses fisiologis yang mengatur reaksi tanaman terhadap faktor-faktor
lingkungan. Tabel 2 menunjukkan bahwa panjang lateral root berkorelasi positif dan signifikan pada taraf 0,01 terhadap diameter, tinggi, luas daun, jumlah daun, jumlah tap root, dan jumlah lateral root. Konteks ini menunjukkan bahwa
pertambahan panjang lateral root semai akan menyebabkan meningkatkannya diameter, tinggi, luas daun, jumlah daun, jumlah tap root, dan jumlah lateral root
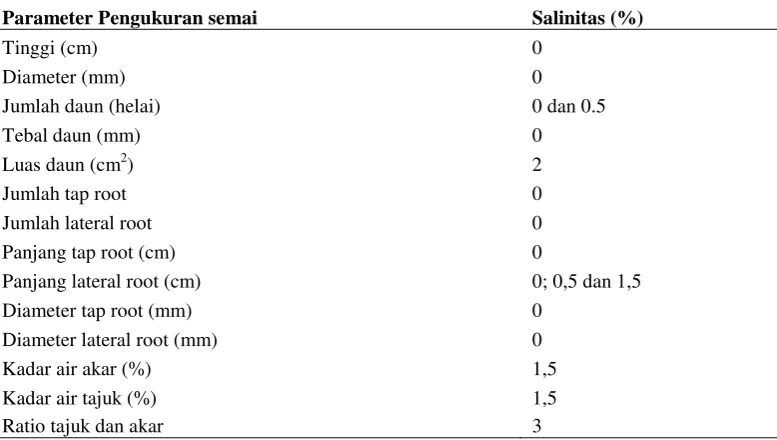
Berikut ringkasan pertumbuhan terbaik dari setiap parameter
Tabel 3. Ringkasan pertumbuhan terbaik parameter penelitian pada tingkat salinitas. Parameter Pengukuran semai Salinitas (%)
Tinggi (cm) 0
Jumlah lateral root 0
Panjang tap root (cm) 0
Panjang lateral root (cm) 0; 0,5 dan 1,5
Diameter tap root (mm) 0
Diameter lateral root (mm) 0
Kadar air akar (%) 1,5
Kadar air tajuk (%) 1,5
Ratio tajuk dan akar 3
Rata-rata pertumbuhan optimal dari setiap parameter untuk pertumbuhan
semai B. cylindrica terdapat pada salinitas 0%. Sesuai dengan pernyatan Noor dkk (2006) bahwa salinitas optimum untuk tumbuhnya Bruguiera adalah < 2,5%.
Hasil penelitian pada tabel 3 ini menunjukkan pertumbuhan optimum jika berada pada kadar salinitas 0% yang meliputi tinggi, diameter, jumlah daun, tebal daun, luas daun, jumlah, panjang, diameter tap root maupun lateral root, kadar air akar
dan tajuk, serta rasio tajuk dan akar. Namun perlu dilakukan pemberian naungan 50% sesuai penelitian simbolon (2013) untuk mendapatkan pertumbuhan yang
lebih baik. Sementara itu faktor lain yang perlu dipertimbangkan adalah perlunya dilakukan pemeraman propagul sebelum menuju tahap persemaian seperti pernyataan Pasaribu (2013) bahwa pemeraman propagul berpengaruh nyata
terhadap tinggi, diameter, jumlah daun, luas daun, rasio akar dan tajuk semai
Dalam penelitiannya, Wong dkk (2007) telah mengidentifikasi dan
mengisolasi 126 cDNA gen yang membawa sifat toleransi salinitas dari akar mangrove jenis B. cylindrica menggunakan suppression subtractive hybridization
(SSH) dan bacterial function screening. Sebanyak 75 rangkaian DNA pembawa sifat toleransi terhadap salinitas berasosiasi dengan bakteri Escherichia coli
sehingga berfungsi mengoptimalkan transportasi, meningkatkan metabolisme, dan
fungsi lainnya sehingga tidak masalah menerima tekanan salinitas 2 %. Hasil ini berhubungan dengan kemampuan B. cylindrica yang pada dasarnya tumbuh
KESIMPULAN DAN SARAN
Kesimpulan
Semai B. cyindrica menunjukkan respons pertumbuhan terbaik pada
salintas 0% karena 85% parameter mengarah kepada pertumbuhan yang optimal. Pemberian salinitas memberikan pengaruh yang nyata terhadap pertumbuhan semai B.cylindrica terutama pada bagian akar. Salinitas yang tinggi
mengakibatkan pertumbuhan semai B.cylindrica terhambat.
Saran
Sebaiknya semai B. cylindrica diberi naungan 50% untuk pertumbuhan yang lebih optimal dengan kadar salinitas 0%. Diperlukan penelitian lanjutan untuk mengidentifikasi gen pembawa sifat yang membuat B. cylindrica mampu
bertahan dalam tekanan salinitas yang tinggi. Selain itu penelitian di lapangan juga dibutuhkan untuk mengetahui kemampuan pertumbuhan akar semai
TINJAUAN PUSTAKA
Kondisi Umum Hutan Mangrove
Hutan mangrove umumnya tumbuh dalam 4 zona, yaitu pada daerah
terbuka, daerah tengah, daerah yang memiliki sungai berair payau sampai hampir
tawar, serta daerah ke arah daratan yang memiliki air tawar.
Samingan (1980) menemukan bahwa di Karang Agung, Sumatera Selatan,
di zona mangrove yang berhadapan dengan laut didominasi oleh S. alba yang
tumbuh pada areal yang betul-betul dipengaruhi oleh air laut. Van Steenis (1958)
melaporkan bahwa S. alba dan A. alba merupakan jenis-jenis ko-dominan pada
areal pantai yang sangat tergenang ini. Komiyama, dkk (1988) menemukan bahwa
di Halmahera, Maluku, di zona ini didominasi oleh S.alba. Komposisi floristik
dari komunitas di zona terbuka sangat bergantung pada substratnya. S. alba
cenderung untuk mendominasi daerah berpasir, sementara A. marina dan
R.mucronata cenderung untuk mendominasi daerah yang lebih berlumpur.
Meskipun demikian, Sonneratia akan berasosiasi dengan Avicennia jika tanah
lumpurnya kaya akan bahan organik.
Mangrove di zona ini terletak dibelakang mangrove zona terbuka biasanya
didominasi oleh jenis Rhizophora. Namun, Samingan (1980) menemukan di
Karang Agung didominasi oleh B.cylindrica. Jenis-jenis penting lainnya yang
ditemukan di Karang Agung adalah B. gymnorrhiza, E. agallocha, R. mucronata,
X. granatum dan X. moluccensis.
Pada zona mangrove di sepanjang sungai berair payau hingga hampir
Agung, komunitas Nypa fruticans terdapat pada jalur yang sempit di sepanjang
sebagian besar sungai. Di jalur-jalur tersebut sering sekali ditemukan tegakan
N. fruticans yang bersambung dengan vegetasi yang terdiri dari Cerbera sp,
Glutarenghas, S. palustris dan X. granatum. Ke arah pantai, campuran Sonneratia
- Nypalebih sering ditemukan. Di sebagian besar daerah lainnya, seperti di Pulau
Kaget dan Pulau Kembang di mulut Sungai Barito di Kalimantan Selatan atau di
mulut Sungai Singkil di Aceh, S. caseolaris lebih dominan terutama bagian
estuari yang berair hampir tawar (Giesen & van Balen, 1991).
Jenis-jenis yang umum ditemukan pada zona perairan payau atau hampir
tawar di belakang jalur hijau mangrove yang sebenarnya ini termasuk Ficus
microcarpus, Intsia bijuga, N. fruticans, Lumnitzera racemosa, Pandanus sp. dan
X. moluccensis. Zona ini meeiliki kekayaan jenis yang lebih tinggi dibandingkan
dengan zonasi lain (Kantor Menteri Negara Lingkungan Hidup, 1993).
Kemampuan mangrove untuk mengembangkan wilayahnya ke arah laut
merupakan satu dari peran penting mangrove dalam pembentukan lahan baru.
Akar mangrove mampu mengikat dan menstabilkan substrat lumpur, pohonnya
mengurangi energi gelombang dan memperlambat arus,
sementara vegetasi secara keseluruhan dapat memerangkap sedimen
(Davies dan Claridge, 1993).
Deskripsi Umum Bruguiera cylindrica
Nama lokalnya adalah burus, tanjang, tanjang putih, tanjang sukim, tanjang sukun, lengadai, bius, lindur. Pohon dengan ketinggian mencapai 23
meter. Kulit kayu abu-abu, relatif halus dan memiliki sejumlah lentisel kecil.
Biasanya tumbuh megelompok dalam jumlah besar di belakang zona Avicennia,
atau bagian tengah vegetasi mangrove kea rah laut. B.cylindrica mempunyai
kemampuan tumbuh pada substrat yang baru terbentuk dan tidak cocok untuk
jenis lainnya. Kemampuan tumbuhnya pada tanah liat membuat pohon ini sangat
bergantung kepada akar untuk memperoleh pasokan oksigen yang cukup, dan oleh
karena itu sangat responsif terhadap penggenangan yang berkepanjangan.
Memiliki buah yang ringan dan mengapung sehingga penyebarannya dapat
dibantu oleh arus air, tapi petumbuhannya lambat. Perbungaan terjadi sepanjang
tahun. Manfaat dari tumbuhan mangrove jenis B. cylindrica adalah untuk kayu
bakar (Noor dkk, 2006).
Salinitas dan Adaptasi Mangrove
Di Indonesia, areal yang selalu digenangi walaupun pada saat pasang rendah umumnya didominasi oleh A. alba atau S. alba. Areal yang digenangi oleh pasang sedang didominasi oleh jenis-jenis Rhizophora. Adapun areal yang
digenangi hanya pada saat pasang tinggi, yang mana areal ini lebih ke daratan, umumnya didominasi oleh jenis- jenis Bruguiera dan X. granatum, sedangkan
areal yang digenangi hanya pada saat pasang tertinggi (hanya beberapa hari dalam sebulan) umumnya didominasi oleh B. sexangula dan L. littorea.
Berdasarkan adaptasi terhadap kandungan garam, mangrove
dikelompokkan menjadi dua kelompok yakni, salt - excreting mangrove, seperti jenis Avicennia, Aegiceras, dan Aegialitis, dan non - secretor mangrove, seperti
flora mangrove menyerap air dengan salinitas tinggi kemudian mengekskresikan
garam dengan kelenjar garam yang terdapat pada daun. Kedua, mencegah masuknya garam (salt exclusion), flora mangrove menyerap air tetapi mencegah masuknya garam, melalui saringan yang terdapat pada akar. Ketiga, akumulasi
garam (salt accumulation), flora mangrove seringkali menyimpan Na dan Cl pada bagian kulit kayu, akar dan daun yang lebih tua.
B. cylindrica dapat mengatasi tekanan garam sampai kadar salinitas 2%
(Wong dkk, 2007). Hal ini disebabkan oleh bakteri E.coli yang berasoiasi dengan gen DNA pembawa sifat tahan terhadap salinitas oleh perakaran semai
B.cylindrica. Pertumbuhan B. cylindrica yang optimal adalah pada naungan 50
Simbolon (2013). Penelitian pengaruh salinitas terhadap pertumbuhan dan
biomassa semai non-sekresi C. tagal dan kandungan lipid pada tingkat pohon dilakukan untuk mengetahui tingkat pertumbuhan dan biomassa semai C.tagal
pada berbagai tingkat salinitas, mengetahui kandingan lipid pada daun dan akar
C.tagal pada tingkat pohon. Penelitian menggunakan 5 perlakuan, yaitu salinitas
0%, 0.5%, 1.5%, 2%, dan 3% dengan masing-masing 5 ulangan. Ekstraksi dan
analisis lipid dilakukan dengan mengekstraksi daun dan akar dari pohon dewasa yang diambil dari hutan mangrove.
Ramayani (2012) mengemukakan bahwa variasi salinitas memberikan
pengaruh yang nyata terhadap pertumbuhan tinggi, diameter, dan biomassa semai
C. tagal. Pertumbuhan semai paling baik adalah 0,5 % dengan pertumbuhan tinggi
PENDAHULUAN
Latar Belakang
Pada tahun 1982 hutan mangrove di Indonesia tercatat seluas 4,25 juta ha
kemudian pada 1993 berkurang menjadi 3,7 juta ha. Pada tahun 2011 menjadi 3,1
juta ha (Giri dkk, 2011). Mangrove Indonesia adalah yang terluas di dunia
namun kondisinya semakin menurun baik dari segi kualitas dan kuantitas dari tahun ke tahun. Luas hutan mangrove yang semakin menurun mengakibatkan dampak yang sangat mengkhawatirkan sehingga mengharuskan kegiatan
rehabilitasi dilakukan di seluruh kawasan mangrove yang rusak. Dalam rehabilitasi mangrove, faktor salinitas dan zona mangrove yang baik saat ini perlu
dipenuhi untuk membangun ekosistem yang dapat memberikan fungsinya secara optimal (Noor dkk, 2006).
Saat ini, kondisi mangrove berada dalam proses kemunduran. Tekanan
populasi yang tinggi, konversi mangrove ke pertanian, industri tambang, industrialisasi pesisir dan urbanisasi, serta konversi pesisir ke pertambakan
menjadi penyebab utama degradasi ekosistem mangrove. Degradasi ekosistem mangrove dapat berdampak pada fungsi fisik, ekologi dan ekonomi. Beberapa daerah wilayah pesisir di Indonesia sudah terlihat adanya degradasi atau
penurunan luas hutan mangrove yang disebabkan karena adanya tekanan akibat pemanfaatan dan pengelolaannya yang kurang memperhatikan aspek kelestarian.
Satu dari upaya konservatif adalah melakukan kegiatan rehabilitasi ekosistem mangrove untuk mengembalikan fungsi hutan mangrove yang mengalami degradasi agar kembali pada kondisi yang dianggap baik dan mampu mengemban
Dalam meningkatkan keberhasilan rehabilitasi, perlu dilakukan
perencanaan yang tepat untuk rehabilitasi, penggunaan semai yang baik dan adaptasi penanaman vegetasi mangrove yang sesuai dengan faktor pertumbuhannya seperti kualitas sifat fisik kimia tanah, salinitas dan pH tanah,
serta lama penggenangan yang dipengaruhi pasang surut air laut. Kondisi salinitas sangat mempengaruhi komposisi mangrove. Berbagai jenis mangrove mengatasi
kadar air dengan cara yang berbeda. Beberapa diantaranya secara selektif mampu menghindari penyerapan garam dari media tumbuhnya, sementara beberapa jenis lainnya mampu mengeluarkan garam dari kalenjar khusus pada daunnya.
B.cylindrica merupakan jenis mangrove dengan ciri unik berupa akar lutut. Spesies ini yang jarang digunakan tetapi diminati dalam rehabilitasi karena
kemampuannya dalam beradaptasi pada kondisi salinitas yang cukup tinggi dan sistem perakaran yang kuat. Kadar salinitas optimum untuk jenis Bruguiera adalah < 2,5 0/0. Noor dkk (2006) menyebutkan bahwa kadar salinitas optimum
untuk B. parviflora adalah 200/0 sementara B. gymnorrhiza adalah 1,0 – 2,50/0. Akar merupakan tempat masuknya unsur hara yang diserap oleh tanaman.
Akar menyerap hara dan menyalurkannya ke daun untuk diproses menjadi energi melalui fotosintesis. Data perkembangan akar dapat diaplikasikan pada penanaman di lapangan melalui kegiatan rehabilitasi. Dengan diketahuinya
perkembangan akar pada setiap kadar salinitas maka dapat dibuat jarak tanam yang baik di lapangan sehingga tidak terjadi perebutan unsur hara dan dapat
Spesies mangrove memiliki kemampuan adaptasi yang berbeda-beda
terhadap kondisi tanah, salinitas, temperatur, curah hujan dan pasang surut. Hal ini menyebabkan terjadinya struktur dan konposisi tumbuhan mangrove dengan batas-batas yang khas, mulai dari zona yang dekat daratan sampai dengan zona
yang dekat dengan lautan, serta menyebabkan terjadinya perbedaan struktur tumbuhan mangrove antar daerah. Informasi tentang kemampuan adaptasi
tumbuhan mangrove khususnya akar terhadap pengaruh lingkungan masih sangat terbatas. Salinitas merupakan salah satu faktor yang sangat menentukan perkembangan hutan mangrove. Oleh karena itu diperlukan penelitian
pertumbuhan akar mangrove jenis B. cylindrica pada berbagai variasi salinitas.
Tujuan Penelitian
Penelitian ini bertujuan untuk mengetahui kadar salinitas terbaik terhadap
pertumbuhan dan perkembangan akar semai B. cylindrica. Manfaat Penelitian
1. Memberikan data dan informasi kadar salinitas optimal pada persemaian
B. cylindrica berdasarkan pertumbuhan bagian atas dan perkembangan akar.
2. Menjadi referensi dalam menentukan jarak tanam terbaik untuk penanaman
jenis B.cylindrica.
Hipotesis Penelitian
Diduga kadar salinitas 0,5 % memberikan respons terbaik terhadap
ABSTRACT
ARIF SYUHADA. Root Growth of B. cylindrica in Salinity Variation. iSupercised by MOHAMMAD BASYUNI dan YUNASFI
ABSTRAK
ARIF SYUHADA. Pertumbuhan Akar Semai Bruguiera cylindrica Pada Berbagai Variasi Salinitas. Dibimbing oleh MOHAMMAD BASYUNI dan YUNASFI
Luas hutan mangrove terus mengalami penurunan sehingga mengharuskan kegiatan rehabilitasi pada seluruh kawasan hutan mangrove yang rusak.
B.cylindrica merupakan spesies mangrove dengan ciri unit berupa akar lutut yang
PERTUMBUHAN AKAR SEMAI Bruguiera cylindrica PADA
BERBAGAI VARIASI SALINITAS
SKRIPSI
Oleh: ARIF SYUHADA
121201118/ BUDIDAYA HUTAN
FAKULTAS KEHUTANAN UNIVERSITAS SUMATERA UTARA
PERTUMBUHAN AKAR SEMAI Bruguiera cylindrica PADA
BERBAGAI VARIASI SALINITAS
SKRIPSI
Oleh: ARIF SYUHADA
121201118/ BUDIDAYA HUTAN
Skripsi sebagai syarat untuk memperoleh gelar sarjana di Fakultas Kehutanan
Universitas Sumatera Utara
FAKULTAS KEHUTANAN UNIVERSITAS SUMATERA UTARA
LEMBAR PENGESAHAN
Judul Penelitian : Pertumbuhan Akar Semai Bruguiera cylindrica pada Berbagai Variasi Salinitas
Nama : Arif Syuhada NIM : 121201118 Minat : Budidaya Hutan
Menyetujui Komisi Pembimbing
Mohammad Basyuni, S.Hut, M.Si, Ph.D
Ketua Anggota Dr. Ir. Yunasfi, M.Si
Mengetahui,
Dekan Fakultas Kehutanan Siti Latifah, S.Hut, M.Si., Ph.D
ABSTRACT
ARIF SYUHADA. Root Growth of B. cylindrica in Salinity Variation. iSupercised by MOHAMMAD BASYUNI dan YUNASFI
ABSTRAK
ARIF SYUHADA. Pertumbuhan Akar Semai Bruguiera cylindrica Pada Berbagai Variasi Salinitas. Dibimbing oleh MOHAMMAD BASYUNI dan YUNASFI
Luas hutan mangrove terus mengalami penurunan sehingga mengharuskan kegiatan rehabilitasi pada seluruh kawasan hutan mangrove yang rusak.
B.cylindrica merupakan spesies mangrove dengan ciri unit berupa akar lutut yang
RIWAYAT HIDUP
Penulis dilahirkan di Kabanjahe pada tanggal 11 Desember 1993 dari pasangan Bapak Hamdani, S.Pd dan Ibu Nuraini, S.Pd. Penulis merupakan anak kedua dari tiga orang bersaudara.
Pada tahun 2006 Penulis lulus dari SD 048071 Kabanjahe, tahun 2009 lulus dari SMP Negeri 1 Kabanjahe dan tahun 2012 lulus dari SMA Negeri 1
Kabanjahe. Tahun 2012 Penulis melanjutkan kuliah di Universitas Sumatera Utara (USU) Medan sebagai Mahasiswa di Program studi Kehutanan Fakultas Pertanian melalui jalur Ujian Masuk Bersama (UMB) reguler.
Selama Mengikuti Perkuliahan Penulis Aktif dalam organisasi Rain Forest dari tahun 2012-2016 di Fakultas Kehutanan, Universitas Sumatera Utara dan
turut aktif dalam organisasi Kemangteer (Kesemat Mangrove Volunteer) Medan pada tahun 2014-2016. Penulis mengikuti Praktik Pengenalan Ekosistem Hutan di kawasan hutan mangrove Pulau Sembilan Kecamatan Pangkalan Berandan,
Kabupaten Langkat, Sumatera Utara pada tahun 2013. Penulis menjadi asisten laboratorium Ekologi Hutan pada tahun 2013 dan asisten laboratorium Dendrologi
KATA PENGANTAR
Alhamdulillahirobbil’alamin. Puji dan syukur kehadirat Allah SWT atas rahmat dan karunia-Nya Penulis dapat merampungkan skripsi yang berjudul: “Pertumbuhan Akar Semai Bruguiera cylindrica Pada Berbagai Variasi Salinitas”.
Penulisan skripsi ini merupakan tugas akir dalam pendidikan Strata-1 dan syarat untuk memperoleh gelar Sarjana pada Program Studi Kehutanan Fakultas
Kehutanan Universitas Sumatera Utara.
Penyusunan skripsi ini tidak terlepas dari bantuan, dukungan, dan bimbingan berbagai pihak. Oleh karena itu, penulis ingin mengucapkan terima
kasih kepada:
1. Kedua orang tua, Bapak Hamdani, S.Pd dan Ibu Nuraini, S.Pd atas dukungan
dari segi moriil maupun materiil serta kasih sayang tanpa batas. Setiap hal yang diberikan kedua orang tua kepada penulis merupakan semangat dalam perjuangan menyelesaikan skripsi ini. Skripsi ini adalah satu dari bukti cinta
penulis kepada kedua orangt tua.
2. Bapak Mohammad Basyuni, S.Hut., M.Si., Ph.D dan Bapak Dr. Ir. Yunasfi,
M.Si selaku Komisi Pembimbing yang senantiasa meluangkan waktu untuk membimbing, serta memberikan kritik dan saran terhadap penulisan skripsi ini.
3. Rekan tim peneliti (Desya Alvioinita Keliat, Nina B. Manalu, Marnida Uli Lubis, Muammar Syafwan, Taufik Fardiman C. Telaumbanua, Fuad Khalil
4. Terakhir, penulis hendak menyapa setiap nama yang tidak dapat penulis
cantumkan satu per satu, terima kasih atas doa dan dukungan yang senantiasa mengalir tanpa sepengetahuan penulis. Terimakasih kepada orang-orang yang turut bersuka cita atas keberhasilan penulis menyelesaikan skripsi ini.
Penulis menyadari bahwa pembuatan skripsi ini masih jauh dari kesempurnaan baik dari segi materi maupun teknik penulisan. Oleh karena itu,
penulis sangat mengharapkan kritik dan saran yang membangun dari para pembaca demi penyempurnaan skripsi ini. Semoga skripsi ini dapat bermanfaat dalam pengembangan ilmu pengetahuan, khususnya bidang kehutanan.
Medan, Agustus 2016 Penulis
DAFTAR ISI
Salinitas dan Adaptasi mangrove... ... 6
METODOLOGI PENELITIAN Waktu dan Tempat Penelitian ... 8
Lokasi Pengambilan Sampel ... 8
Bahan dan Alat Penelitian ... 8
Prosedur Pengamatan ... 9
Analisis Data ... 10
Parameter Pengamatan ... 11
HASIL DAN PEMBAHASAN Pengaruh Pertumbuhan Semai B. cylindrica ... 14
Pengaruh Pertumbuhan Akar Semai B. cylindrica ... 18
DAFTAR TABEL
No. Halaman
DAFTAR GAMBAR
No. Halaman
1. Karakteristik Akar Semai B. Cylindrica ... 14
2. Respons Tinggi Dan Diameter Semai B. cylindrica ... 15
3. Respons Jumlah, Diameter, dan Luas Daun B. cylindrica ... 17
4. Respons Jumlah Akar B. cylindrica ... 19
5. Respons Panjang Akar B. cylindrica ... 19
6. Respons Diameter Akar B. cylindrica ... 20
7. Respons Kadar Air Semai B. cylindrica ... 22