Rr SRI CATUR SETYAWATININGSIH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa disertasi berjudul Karakteristik Biologi Biawak Air (Varanus salvator) Asal Wilayah Sumatera: Tinjauan Morfologi, Molekuler , dan Potensi Reproduksi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2016
RINGKASAN
SRI CATUR SETYAWATININGISIH. Karakteristik Biawak Air (Varanus salvator) Asal Wilayah Sumatera: Tinjauan Morfologi, Molekuler, dan Potensi Reproduksi. Dibimbing oleh DEDY DURYADI SOLIHIN, ARIEF BOEDIONO, WASMEN MANALU, dan EVY AYU ARIDA.
Biawak air (Varanus salvator) merupakan spesies kompleks, terdistribusi luas, dan sebagai komoditas ekspor reptil terbesar dari Indonesia. Spesies ini menghuni daratan utama (pulau) dan pulau satelitnya. Pulau satelit yang terisolasi diduga mendorong terjadinya spesiasi alopatrik yang dapat mengakibatkan perubahan taksonomi biawak air, sebagaimana terjadi di Filipina. Kami tertarik untuk melakukan karakterisasi aspek biologi V. s. macromaculatus yang berasal dari wilayah Sumatera karena beberapa hal. Pertama, V. s. macromaculatus adalah subspesies tersamar. Kedua, wilayah Sumatera terdiri atas daratan utama (Pulau Sumatera) dan pulau-pulau satelitnya. Beberapa pulau-pulau satelit dari Pulau Sumatera, berdasarkan sejarah geologi, merupakan pulau yang terisolasi, misalnya Pulau Simeulue. Kami menduga spesiasi alopatrik juga terjadi pada V. s. macromaculatus di pulau-pulau satelit yang terisolasi dari Pulau Sumatera.
Ketiga, wilayah Sumatera merupakan lokasi sumber komoditas V. salvator yang menyumbangkan lebih 50% dari total kuota tangkap (Dir Jend KSDAE 2015a). Penelitian ini bertujuan mengidentifikasi V. s. macromaculatus yang berasal dari Pulau Sumatera dan pulau-pulau satelitnya secara morfologi dan molekuler. Selain itu, karakteristik reproduksi V. s. macromaculatus betina yang berasal dari wilayah Riau dilakukan dengan mendeskripsikan anatomi oviduk dan ovarinya untuk menentukan strategi dan potensi reproduksinya.
Kajian morfologi menggunakan 58 ekor V. salvator yang diperoleh dari 12 lokasi, yaitu Pulau Simeulue, Meulaboh, Serdang Bedagai, Pekanbaru, Siak, Pulau Bengkalis, Pulau Mendol, Pulau Kundur, Pulau Combol, Pulau Batam, Pulau Bangka, dan Pulau Jawa (sebagai outgroup). Kajian ini menentukan karakter morfologi diagnostik antara V. s. macromaculatus yang berasal dari Pulau Sumatera dan pulau-pulau satelitnya. Kajian molekuler berhasil mengamplifikasi sebanyak 50 sekuens V. s. macromaculatus yang diperoleh dari 12 lokasi yang sama dengan kajian morfologi. Keragaman haplotipe, dan keragaman genetik gen COI dihitung menggunakan DNA Sequence Polymorphism
V. salvator yang berasal dari Pulau Simeulue dan Pulau Bangka berhasil diidentifikasi sebagai morfo-spesies yang berbeda dari V. s. macromaculatus yang berasal dari Pulau Sumatera (Meulaboh, Serdang Bedagai, Siak, dan Pekanbaru) dan Riau Kepulauan (Pulau Bengkalis, Pulau Mendol, Pulau Kundur, Pulau Combol, dan Pulau Batam). Karakter morfologi diagnostik berupa karakter morfometrik di bagian kepala (indeks 10 dan indeks 11), karakter hitungan sisik (Q dan S), dan pola warna tubuh. Penggunaan gen COI sebagai penanda genetik berhasil mengidentifikasi sebanyak 18 haplotipe. Keragaman genetik terbesar dan terkecil ditunjukkan oleh kelompok biawak air yang berasal dari Pulau Jawa dan Pulau Simeulue secara berturut-turut. Fenogram dan jaring haplotipe menunjukkan kelompok biawak air yang berasal dari Pulau Bangka dan Pulau Simeulue terpisah dengan kelompok yang berasal dari Pulau Sumatera dan Riau Kepulauan. Rerata jarak genetik bersih antarkelompok mengonfirmasi kelompok biawak air yang berasal dari Pulau Simeulue dan Pulau Bangka secara berturut-turut sebagai kandidat spesies dan subspesies baru, sedangkan biawak air yang berasal dari Pulau Sumatera dan Riau Kepulauan diidentifikasi tetap sebagai subspesies V. s. macromaculatus. Hasil yang diperoleh menunjukkan terjadinya spesiasi alopatrik pada biawak air yang berasal dari Pulau Simeuleu dan Pulau Bangka terkait dengan sejarah geologinya.
Berdasarkan anatomi reproduksinya, semua spesimen yang diuji mewakili semua tingkatan umur, yaitu dari juvenil hingga dewasa. Fenomena simetri oviduk V. s. macromaculatus ditunjukkan dalam jumlah dan panjangnya. Jumlah total folikel ovari pada individu dewasa minimal 200 butir sehingga tergolong memiliki potensi reproduksi tinggi. Subspesies ini memiliki strategi mulai bereproduksi pada ukuran relatif kecil, memiliki clutch size berukuran sedang dan tergolong multiple clutching. Strategi reproduksi subspesies ini sama dengan V. salvator kompleks melintasi kisaran geografiknya sehingga dua kandidat taksa baru diduga memiliki strategi dan potensi reproduksi yang sama. Dengan demikian V. salvator berpotensi untuk ditangkarkan mengingat potensi reproduksinya tinggi dan kisaran jenis pakan yang luas.
Penelitian ini secara keseluruhan memberikan fakta-fakta ilmiah bahwa karakterisasi morfologi, molekuler, dan reproduksi dapat digunakan dalam mengupayakan konservasi V. salvator. Meskipun demikian, studi lanjut tentang validasi taksa dan penentuan status perlindungan V. salvator yang berasal dari Pulau Simeulue dan Pulau Bangka perlu dilakukan. Eksplorasi dan survei lebih luas perlu dilakukan pada pulau terisolasi (contoh: Pulau Enggano) untuk menemukan kelompok V. salvator yang belum teridentifikasi.
SUMMARY
SRI CATUR SETYAWATININGISIH. Characteristics of Sumatran Water Monitor Lizard (Varanus salvator): An Overview of Morphology, Molecular and Reproductive Potency. Supervised by DEDY DURYADI SOLIHIN, ARIEF BOEDIONO, WASMEN MANALU, and EVY AYU ARIDA
Water monitor lizard (Varanus salvator) is a complex species, widely distributed and as the largest reptile export commodity from Indonesia. It inhabits the mainland (island) and satellite islands. Isolated satellite islands are predicted to boost the allopatric speciation which can result in taxonomy changes such as the one occurring in Philippines. We were interested in characterizing the biological aspects of the V. s. macromaculatus originating from Sumatra region due to several reasons. Firstly, V. s. macromaculatus is a cryptic species. Secondly, Sumatra region consists of mainland (Sumatra Island) and satellite islands. Based on the geological history, some satellite islands of Sumatra Island are considered as isolated islands such as Simeulue Island. We assumed that allopatric speciation also occurred in V. salvator satellite islands which were isolated from Sumatra Island. Thirdly, Sumatra region roles as the source area of
V. salvator commodities contributing more than 50% of the total capture quota (Directorate General of Natural Resource Conservation and Ecosystem, 2015). The objectives of this research were to identify V. salvator from Sumatra Island and its satellite islands morphologically and molecularly. In addition, the reproduction characteristics of female V. salvator from Riau were obtained by identifying its ovary and oviduct anatomy to determine the reproductive strategy and potential.
Morphological study used 58 samples that were obtained from 12 locations, namely: Simeulue Island, Meulaboh, Serdang Bedagai, Pekanbaru, Siak, Bengkalis Island, Mendol Island, Kundur Island, Combol Island, Batam Island, Bangka Island, and Java Island (as outgroup). This study determined the diagnostic morphological characters between V. salvator from Sumatra Island and its satellites islands. Molecular study successfully amplified 50 sequences of V. salvator that were obtained from 12 locations that were same with the morphological study. Haplotype diversity and genetic diversity of COI gene were calculated using DNA Sequence Polymorphism (DNA SP) version 5. Net haplotypes using Median Joining Network (MJ) and NETWORK version 5.0.0.0 were used to identify the development stages of phenogram group. The mean between net genetic distances was estimated using MEGA program 6 to determine the status of the taxa. If the mean between net genetic distances was higher than 2% then it was classified as a candidate for the new species, while if it was less than 2% then it was classified as a candidate for a new subspecies. Study of anatomy of the ovary and oviduct used 12 samples from 6 locations that were included in Riau region, namely: Bengkalis Island, Pekanbaru, Siak, Mendol Island, Kundur and Batam Island. In this study, the oviduct and ovary were described; the follicles and eggs were counted. Data were obtained to determine the reproductive strategy and potential.
(Meulaboh, Serdang Bedagai, Siak, and Pekanbaru) and Riau Islands (Bengkalis Island, Mendol Island, Kundur Island, Combol Island, and Batam Island) based on morphometric characters in the head (index 10 and index 11), scale counts around the tail and midbody (Q and S), and color pattern of the body. The use of COI genes as genetic markers successfully identified 18 haplotypes. The largest and smallest genetic diversities were found at V. salvator from the Java Island and Simeulue Island, respectively. The phenogram and nets haplotype showed that V. salvator from Bangka Island and Simeulue Island were separated with the other groups from Sumatra Island and Riau Islands. The mean of net genetic distance among groups confirmed that V. salvator group from Simeulue and Bangka Island were candidate for new species and subspecies, respectively. Meanwhile, water monitors from Sumatra Island and Riau Islands were still identified as V. s. macromaculatus sub species. The results indicated that allopatric speciation on water monitors from Simeulue and Bangka Island was associated with geological history.
Conservation efforts on V. salvator also need reproduction information. V. s. macromaculatus oviduct symmetry phenomenon was shown by the number and length. The ovarian follicles of mature V. salvator shows high quantities (minimal 200). The high reproductive potency is reflected in their reproductive strategy. This subspecies had strategy to start reproduction at a relatively small size, a medium clutch size and multiple clutching. Therefore, the reproductive potential of this species was high. The reproductive strategy of this subspecies was similar to V. salvator complex across its geographic range so that two new taxa candidates were thought to have same reproductive strategy and potential. These results indicated that there were potency to keep V. salvator in captivity because of its reproductive potential and a broad range of food.
The overall results of this research provided scientific fact that the morphological, molecular and reproduction characterization could be used for the conservation of V. salvator. Further study on validation of taxa and population density of V. salvator from Simeulue and Bangka Island is required to determine the protection status for the two new taxa candidates. Broader exploration and survey are necessary to be conducted on isolated islands (e.g. Enggano) to find an unidentified group of V. salvator.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB.
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Biosains Hewan
KARAKTERISTIK BIAWAK AIR (
Varanus salvator
) ASAL
WILAYAH SUMATERA: TINJAUAN MORFOLOGI,
MOLEKULER, DAN POTENSI REPRODUKSI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Penguji Luar Komisi pada Ujian Tertutup:
Prof Dr Ir Cece Sumantri, MSc
(Staf Pengajar pada Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, IPB)
Dr Ir Mirza Dikari Kusrini, MSi
(Staf Pengajar pada Departemen Konservasi Sumberdaya Hutan dan Ekowisata, Fakultas Kehutanan, IPB)
Penguji Luar Komisis pada Sidang Promosi:
Prof Dr Adel Zamri, MS, DEA
(Dekan Fakultas Matematika dan Ilmu Pengetahuan, Universitas Riau)
Prof Dr Ir Cece Sumantri, MSc
Judul Disertasi : Karakteristik Biawak Air (Varanus salvator) asal Wilayah Sumatera: Tinjauan Morfologi, Molekuler, dan Potensi Reproduksi
Nama : Rr Sri Catur Setyawatiningsih
NIM : G 362110021
Disetujui oleh Komisi Pembimbing
Dr Dedy Duryadi Solihin, DEA
Ketua Prof drh Arief Boediono, PhD, PAVet(K) Anggota
Prof Ir Wasmen Manalu, PhD
Anggota Dr rer nat Evy Ayu Arida Anggota
Diketahui oleh
Ketua Program Studi Biosains Hewan
Dr Ir Raden Roro Dyah Perwitasari, MSc
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian Tertutup : 15 Juli 2016
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian ini ialah biawak air, dengan judul Karakteristik Biawak Air (Varanus salvator) asal Wilayah Sumatera: Tinjauan Morfologi, Molekuler, dan Potensi Reproduksi.
Saya menghaturkan banyak terima kasih atas supervisi dan bimbingan Dr Dedy Duryadi Solihin, DEA sepanjang perjalanan studi ini. Saya juga mengucapkan banyak terima kasih kepada anggota komisi pembimbing, yaitu Dr rer nat Evy Ayu Arida atas ide, dukungan, dan bimbingannya selama ini; Prof drh Arief Boediono, PhD, PAVet(K) atas semua saran, bimbingan khususnya tentang etika terhadap hewan; serta Prof Ir Wasmen Manalu, PhD atas tata cara penulisan artikel ilmiah dan cerita-cerita menarik yang menimbulkan ide penelitian ke depannya.
Saya mengucapkan banyak terima kasih kepada: semua staf pengajar di Biosains Hewan yang telah memberikan bekal ilmu selama kuliah; Dr Drh Chairun Nisa‟, MSi PAVet, Dr Drh Nurhidayat MS, PhD yang telah menularkan ilmunya tentang cara pengambilan darah dan membedah biawak; Drh Adi Winarto, PhD PAVet, untuk diskusi dan bantuannya mengenal anatomi dan histologi reproduksi biawak air.
Banyak terima kasih saya ucapkan kepada Agus Widodo, SSi, Rohilen, Juliadi, Rita Octavia, MSi, Dedek, Ogek Komarudin, Maria Ulfa (Fauna & Flora International/ FFI Marine), Aini Qamariah, Anti, MSi, Ajeng Pramita, Sukoco, Juliman, Ravki Adiyandra, Bapak Evan, Ibu Solihin dan Dedy Rivandi, M.Si, Bapak Salim, Bapak Edun, Bapak Samingan, Kel. Mak Nga, Yani, Misro Heryanto, Melianasari, Indra, Asy‟ari atas bantuannya selama di lapangan. Saya berterima kasih kepada Bapak Heri Djumhaeri, Ibu Retno Untari, Ibu Tini Wahyuni, dan Pak Iwan Rochmana yang telah menularkan ilmunya tentang penggunaan peralatan di laboratorium. Saya mengucapkan terima kasih kepada Bapak Harlan Suharlan (staf KSDAE) yang telah membantu saya dalam mendapatkan data realisasi penangkapan biawak air.
Saya juga berterima kasih kepada teman-teman seperjuangan saya di Biosains Hewan, yaitu Dr Andy Darmawan, Dr Yuliadi Zamroni, I Dr Maria Ulfa, Widadi Padmarsari, MSi, Dr Jusmaldi, Dr Keliopas Krey yang telah memberikan ide, saran dan kebersamaan. Saya juga berterima kasih kepada teman-teman di Laboratorium Biologi Molekuler, yaitu Yuli Wahyu Tri Mulyani, MSi, Robba Fahrisy Darus, MSi, Syamsul Bahri, M.Si, dan teman-teman lainnya yang tidak dapat saya sebutkan satu per satu atas pertemanannya, diskusi, kebersamaan dan dukungannya. Saya berterima kasih kepada Dr Pieter Agusthinus Riupassa dan Andy Khaerul, MSi atas program Endnote sehingga mempermudah dalam penulisan ini.
kepada Safi‟i (suami), Syifaa‟ Ardiyanti Safitri dan Anisah Ardiningrum untuk cinta, pengertian, kerja sama, serta kesabarannya.
Disertasi ini didanai oleh Kementerian Riset Teknologi dan Pendidikan Tinggi Republik Indonesia, Universitas Riau dan bantuan pendidikan Pemerintah Provinsi Riau.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2016
DAFTAR ISI
DAFTAR TABEL xiv
DAFTAR GAMBAR xv
DAFTAR LAMPIRAN xvi
1 PENDAHULUAN UMUM 1
Latar Belakang 1
Perumusan Masalah 4
Tujuan dan Manfaat 4
Kebaharuan 5
2 VARIASI MORFOLOGI BIAWAK AIR (Varanus salvator) ASAL
WILAYAH SUMATERA 7
Pendahuluan 7
Metode Penelitian 8
Hasil 11
Pembahasan 18
Simpulan 20
3 KARAKTERISTIK GENETIKA MOLEKULER BIAWAK AIR (Varanus salvator) ASAL WILAYAH SUMATERA BERDASARKAN GEN COI
21
Pendahuluan 21
Bahan dan Metode 21
Hasil 24
Pembahasan 28
Simpulan 30
4 KARAKTERISTIK REPRODUKSI BIAWAK AIR (Varanus salvator)
ASAL WILAYAH RIAU 31
Pendahuluan 31
Metode Penelitian 31
Hasil 34
Pembahasan 39
Simpulan 41
5 PEMBAHASAN UMUM 42
6 SIMPULAN DAN SARAN 45
Simpulan 45
Saran 45
DAFTAR PUSTAKA 46
LAMPIRAN 52
DAFTAR TABEL
Tabel 2.1 Senarai, singkatan, dan takrif karakter morfologi yang digunakan dalam studi taksonomi V. salvator (Koch et al.
2007)
9
Tabel 2.2 Data rentang, rerata, simpangan baku, dan hasil uji Duncan pada karakter morfometrik tubuh dan kepala (indeks 1, indeks 2, indeks 10, dan indeks 11) V. salvator yang diuji
12
Tabel 2.3 Nilai akar ciri, proporsi variansi, dan proporsi kumulatif karakter morfometrik tubuh dan kepala V. salvator yang diuji
12
Tabel 2.4 Hubungan karakter morfometrik tubuh dan kepala V. salvator yang diuji dengan unit taksonomi operasional berdasarkan analisis AKU
12
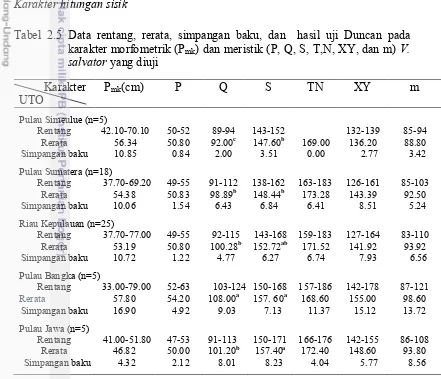
Tabel 2.5 Data rentang, rerata, simpangan baku, dan hasil uji Duncan pada karakter morfometrik (Pmk) dan meristik (P, Q, S, T,N,
XY, dan m) V. salvator yang diuji
14
Tabel 2.6 Nilai akar ciri, proporsi variansi, dan proporsi kumulatif karakter hitungan sisik V. salvator XY, dan m) V. salvator
yang diuji
15
Tabel 2.7 Hubungan karakter meristik dengan unit taksonomi
operasional (UTO) berdasarkan analisis AKU 15 Tabel 2.8 Persentase V. salvator yang diuji berdasarkan pola warna 17 Tabel 3.1 Haplotipe 52 individu V. salvator yang diuji 25 Tabel 3.2 Situs substitusi nukleotida khas sebagai penciri
kelompok-kelompok pada V. salvator yang diuji (huruf yang ditebalkan dan diarsir)
25
Tabel 3.3 Rentang (angka tanpa tanda kurung), rerata (angka dalam tanda kurung) jarak genetik dalam kelompok (pada posisi diagonal) dan jarak genetik antarkelompok (di bagian bawah diagonal) pada V. salvator yang diuji menggunakan metode
p distance
28
Tabel 3.4 Rerata (angka tanpa tanda kurung) jarak genetik bersih antarkelompok (di bagian bawah diagonal) pada V. salvator
yang diuji menggunakan metode p distance dan faktor koreksi (angka dengan tanda kurung)
28
Tabel 4.1 Pengelompokan ukuran folikel ovari dan telur dalam oviduk
DAFTAR GAMBAR
Gambar 1.1 Peta persebaran V. salvator kompleks 2
Gambar 1.2 Diagram alur penelitian 5
Gambar 2.1 Peta lokasi penangkapan V. salvator yang diuji 8 Gambar 2.2 Pengamatan morfometrik dan meristik V.salvator yang diuji 10 Gambar 2.3 AKU karakter morfometrik tubuh dan kepala V. salvator
yang diuji 13
Gambar 2.4 Dendrogram karakter morfometrik tubuh dan kepala V. salvator yang diuji dengan jarak menggunakan metode Euclidean dan hierarchial clustering (hclust)
13
Gambar 2.5 AKU karakter meristik V. salvator yang diuji 16 Gambar 2.6 Dendrogram karakter hitung sisik dengan V. salvator yang
diuji dengan jarak menggunakan metode Euclidean dan
hierarchial clustering (hclust)
16
Gambar 2.7 Pola warna ventral dan dorsal tubuh V. salvator yang diuji 18 Gambar 3.1 Skema posisi penempelan primer VS-COI_F dan VS-COI_R
dan produk amplifikasi 23
Gambar 3.2 Fenogram dan jaring haplotipe 52 individu V. salvator yang
diuji 27
Gambar 4.1 Pengelompokan status reproduksi V. s. macromaculatus
(sensu Zug et al. 1979 dan Fitzgerald et al. 1993) 33 Gambar 4.2 Situs viscerum organ reproduksi V. s. macromaculatus betina 34 Gambar 4.3 Tipe oviduk V. s. macromaculatus pada sisi ventral 35 Gambar 4.4 Bentuk-bentuk ovarium dan tahap perkembangan folikel V. s.
macromaculatus 36
Gambar 4.5 Histogram jumlah folikel dan telur berdasarkan kelas pada
DAFTAR LAMPIRAN
Lampiran 1 Persentase kuota tangkap V. salvator di Indonesia periode
2016 (Dir Jend KSDAE 2015a) 55
Lampiran 2 Senarai V. salvator yang diuji 56
Lampiran 3 Beberapa alat tangkap V. salvator 58 Lampiran 4 Hasil analisis uji Duncan karakter morfometrik V. salvator
yang diuji 59
Lampiran 5 Uji korelasi Pearson antara Pmk dengan karakter merisitk V.
salvator yang diuji 60
Lampiran 6 Hasil analisis uji Duncan karakter meristik V. salvator yang
diuji 61
Lampiran 7 Hasil pengamatan pola warna pada V. salvator yang diuji 62 Lampiran 8 Komposisi empat basa nukleotida gen COI (516 nt)
V. salvator yang diuji 63
1
PENDAHULUAN
Latar Belakang
Indonesia memiliki keanekaragaman spesies biawak tinggi, yaitu 40% dari total biawak (73 spesies) di dunia (Koch et al. 2010a, Arida 2014). Biawak air (Varanus salvator) merupakan salah satu spesies yang paling umum terdapat di Indonesia. Status spesies ini tergolong tidak dilindungi dalam Peraturan Pemerintah Indonesia No. 7 Tahun 1999, “least concern” dalam Lembaga Konservasi Alam Internasional (International Union Conservation Nature/IUCN). Hewan ini juga tergolong dalam Appendiks II dalam Perjanjian Perdagangan Internasional Spesies Flora dan Fauna yang Terancam Punah atau Convention on International Trade in Endangered Species of Wild Fauna and Flora/CITES,yang berarti V. salvator dapat diperdagangkan berdasarkan kuota (Bennett et al. 2010, Arida 2014).
Biawak air merupakan komoditas ekspor kulit reptil terbesar di Indonesia sejak tahun 1980-an hingga saat ini. Rerata penangkapan V. salvator mencapai sekitar 500 ribu ekor/tahun dan diperkirakan lebih dari 95% diperoleh dari alam (Mardiastuti dan Soehartono 2003, Pernetta 2009, CITES 2016). Di berbagai tempat di Indonesia, penduduk menangkap biawak air tidak hanya untuk diambil kulitnya, tetapi juga daging (sebagai sumber protein), lemak (sebagai bahan obat) dan atau sebagai hewan peliharaan (Mardiastuti dan Soehartono 2003, Subasli 2012, Uyeda et al. 2014). Selain itu, hewan ini juga ditangkap karena dianggap sebagai hama (sensu Soehartono dan Mardiastuti 2003). Lima puluh persen lebih dari kuota biawak air Indonesia berasal dari wilayah Sumatera, khususnya Sumatera Utara, Sumatera Selatan, Bengkulu, Jambi, Lampung, Riau, dan Sumatera Barat (Lampiran 1) (Dir Jend KSDAE 2015a).
V. salvator merupakan species complex atau kompleks spesies (Koch et al.
2007, Koch et al. 2010b). Kompleks spesies adalah kelompok-kelompok spesies yang terisolasi secara reproduksi, berkerabat dekat, dan mirip secara morfologis (Fegan dan Prior 2005). Kompleks spesies dapat terjadi karena persebaran yang luas dan banyak dipicu oleh peristiwa geologi sehingga terjadi isolasi geografi. Isolasi geografi menghambat aliran gen (gene flow) dan memunculkan hanyutan gen atau genetic drift. Akibatnya, terjadilah spesiasi alopatrik. Spesiasi alopatrik diduga terjadi pada V. salvator kompleks sehingga saat ini V. salvator kompleks terdiri atas 10 spesies, yaitu: 8 spesies biawak air berada di Filipina (V. palawanensis, V. bangonorum, V. marmoratus, V. dalubhasa, V. samarensis, V. nuchalis, V. cumingi,dan V. ramusseni); ada satu spesies berada endemik di Pulau Togean, Indonesia (V. togianus); dan satu spesies, yaitu V. salvator terdapat di India, Burma, Thailand, China bagian Selatan, Malaysia, dan Indonesia (Koch et al. 2007, Koch et al. 2010b, Koch et al. 2013, Welton et al. 2013b, Welton et al.
2014a).
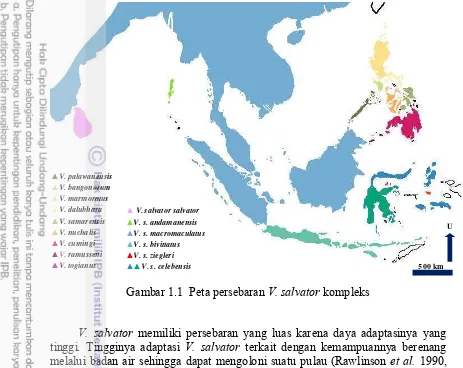
V. salvator merupakan spesies politipik yaitu satu spesies yang memiliki banyak subspesies. Menurut Koch et al. (2013), V. salvator terdiri dari enam subspesies yaitu V. salvator salvator, V. s. andamanensis, V. s. macromaculatus,
Gambar 1.1 Peta persebaran V. salvator kompleks
V. salvator memiliki persebaran yang luas karena daya adaptasinya yang tinggi. Tingginya adaptasi V. salvator terkait dengan kemampuannya berenang melalui badan air sehingga dapat mengoloni suatu pulau (Rawlinson et al. 1990, De Lisle 2007). Spesies ini juga dikenal memiliki kesintasan yang tinggi karena rentang jenis pakannya yang lebar dan hidup pada habitat yang beragam (Bennett 1995, Bundhitwongrut et al. 2008, Uyeda 2009, Cota 2011a, Chatterjee dan Bhattacharyya 2015).
Biawak air merupakan cryptic species atau spesies tersamar. Spesies tersamar adalah dua atau lebih spesies yang tersamar dalam satu nama spesies (Bickford et al. 2007). V. s. macromaculatus yang berasal dari wilayah Sumatera merupakan subspesies tersamar (sensu Welton et al. 2013b). Artinya kelompok individu yang berada di Pulau Sumatera dan pulau–pulau satelitnya berkemungkinan menjadi spesies dan atau subspesies baru jika deskripsi morfologi maupun genetik molekulernya dapat diidentifikasi dengan jelas, seperti yang telah terjadi pada biawak air di Filipina (Welton et al. 2013b, Welton et al.
2014a). ▲V. palawanensis
▲V. bangonorum ▲V. marmoratus ▲V. dalubhasa
▲V. samarensis
▲V. nuchalis
▲V. cumingi ▲V. ramusseni ▲V. togianus
▲V. salvator salvator ▲V. s. andamanensis
▲V. s. macromaculatus
▲V. s. bivittatus ▲V. s. ziegleri
▲▲V. s. celebensis 500 km
Identifikasi biawak air dapat dilakukan menggunakan berbagai karakter. Karakter morfologi yang umum digunakan dalam mengidentifikasi V. salvator
adalah karakter morfometrik, meristik (hitungan sisik), dan pola warna (Koch et al. 2007). Identifikasi menggunakan kromosom menunjukkan bahwa V. s. macromaculatus memiliki 16 makrokromosom, terdiri atas: 2 pasang metasentrik berukuran besar (pada kromosom ke-1 dan ke-2); 2 pasang metasentrik berukuran medium; satu pasang akrosentris (no 5) berukuran kecil; 3 pasang submetasentrik berukuran kecil (pada kromosom ke-6-8); dan 12 pasang mikrokromosom (King dan King 1975, Chaiprasertsri et al. 2013). Morfologi kariotipe kromosom 6-8 merupakan karakter diagnostik antara V. s. macromaculatus dengan V. exanthematicus (Srikulnath et al. 2013). Studi elektroforesis menggunakan protein darah (lactat dehydrogenase atau LDH) pada V. s. macromaculatus menunjukkan bahwa subspesies ini berada dalam kelompok yang sama dengan V. rudicollis, V. nebulosus, V. bengalensis (Holmes et al. 1975).
Seiring dengan kemajuan teknologi, identifikasi dapat dilakukan dengan menggunakan DNA, baik DNA bukan gen, DNA inti, dan DNA mitokondria. Berdasarkan DNA bukan gen, yaitu: mikrosatelit, berhasil diisolasi VSAREP1 dan VSAREP2 pada V. s. macromaculatus (Chaiprasertsri et al. 2013), tetapi tiga lokus Short Interspersed Retrotransposable Elements atau SINE, yaitu: VIN1, VIN2, dan VIN6 tidak terdapat pada V. salvator (Piskurek et al. 2006). Gen-gen pada DNA inti, yaitu gen diacylglyceral lipase-alpha GL-α dan gen prolactin receptor atau PRLR berhasil dirunut dari V. salvator kompleks (Welton et al.
2013b). Ast (2001) berhasil merunut tiga gen mitokondria, yaitu gen NADH dehydrogenase subunit 1 dan 2 atau ND1, ND2, dan gen cytochrome c oxidase subunit 1 atau COI pada V. salvator.
Gen COI telah digunakan sebagai pengidentifikasi V. salvator ssp. di Indonesia (Arida 2014). Fragmen COI, yang sebagian besar bersifat konservatif di tingkat spesies dan hanya beberapa bagian saja bersifat variabel sehingga dapat menentukan identitas spesies secara akurat, tetapi masih berbeda dalam genus yang sama. Avise et al. (1987) menyatakan bahwa gen COI yang terdapat dalam DNA mitokondria memiliki polimorfisme intraspesies yang ekstensif. Data variabilitas dalam DNA mitokondria pada intraspesies dapat dijadikan sebagai dasar hubungan filogenetik antarkelompok dalam satu wilayah dan distribusi geografik dari kelompok-kelompok filogenetik. Gabungan antara hubungan filogenetik antarkelompok dan distribusi geografi dari kelompok filogenetik disebut filogeografi intraspesies (Avise et al. 1987). Studi filogeografi berimplikasi pada diketahuinya asal-usul suatu taksa (spesies atau subspesies) dari daerah alaminya sehingga jika kelak spesies tersebut dilepasliarkan sebaiknya ke daerah asalnya. Dengan demikian, filogeografi merupakan satu upaya konservasi spesies yang dijadikan komoditas.
Pemanenan V. salvator dalam jumlah yang tinggi dalam periode waktu lama akan memengaruhi ukuran populasi. Jika peremajaan lebih sedikit dibandingkan pemanenan maka ukuran populasi akan menurun. Penurunan populasi V. salvator
2013). Jika penurunan populasi tidak segera ditangani dan eksploitasi berjalan terus, tidak menutup kemungkinan terjadinya kepunahan V. salvator lokal di masa mendatang. Oleh karena itu, pengetahuan dasar mengenai biologi reproduksi hewan ini akan dapat menyelesaikan persoalan seberapa jauh kontribusi peremajaan populasi didasarkan pada kemampuan mandiri spesies ini dalam memperbanyak anggota populasinya setiap kurun waktu tertentu. Dengan demikian akan diketahui pemulihan populasi antara pemanenan dengan peremajaan didasarkan pada kemampuan intrinsiknya. Informasi status reproduksi tersebut dapat berupa hubungan ukuran tubuh dengan status reproduksi, jumlah telur yang dihasilkan (clutch size), berapa kali bertelur dalam satu musim reproduksi (clutch number). Informasi potensi reproduksi V. salvator adalah merupakan langkah awal dalam konservasi spesies terkait dengan estimasi ukuran populasi di masa mendatang dan kuota tangkap tahunan sehingga pemanfaatan spesies dapat berkelanjutan.
Perumusan Masalah
Berdasarkan uraian di atas timbullah pertanyaan-pertanyaan sebagai berikut: (i) adakah perbedaan karakter morfologi dan molekuler di antara V. s. macromaculatus yang berasal dari Pulau Sumatera dan pulau-pulau satelitnya? (ii) bagaimana potensi reproduksi subspesies yang sama yang berasal dari wilayah Riau? Dalam menjawab pertanyaan tersebut, akan dilakukan 3 pendekatan, yaitu (1) pemeriksaan karakter morfologi untuk melihat variasi morfologi di antara V. s. macromaculatus yang berasal dari wilayah Sumatera; (ii). penggunaan penanda genetik DNA mitokondria (gen COI) pada V. s. macromaculatus yang berasal dari wilayah Sumatera untuk mendapatkan karakteristik genetika molekulernya; (iii). pengamatan anatomi organ reproduksi V. s. macromaculatus betina untuk menentukan strategi dan potensi reproduksinya. Secara skematis, diagram alur pemikiran disajikan pada (Gambar 1.2).
Tujuan dan Manfaat
Gambar 1.2 Diagram alur penelitian
Kebaharuan
Kebaharuan penelitian ini adalah: (i) pengujian hubungan V. s. macromaculatus yang berasal dari wilayah Sumatera berdasarkan karakter morfologi (morfometrik, hitungan sisik, dan pola warna) berhasil mengidentifikasi kelompok biawak air yang berasal dari Pulau Simeulue dan Pulau Bangka sebagai morfospesies yang berbeda dari kelompok yang berasal dari Pulau Sumatera dan Riau Kepulauan. (ii) Penggunaan penanda molekuler genetik (gen COI) pada subspesies yang sama menghasilkan dua taksa baru, yaitu kelompok biawak air Informasi data dasar biologi berupa data morfologi, molekuler, dan potensi reproduksi
V. s. macromaculatus. Kajian molekuler:
Penggunaan gen COI untuk mengonfirmasi status taksa V. s. macromaculatus yang berasal dari wilayah Sumatera.
Karakterisasi aspek biologi
Varanus salvator
Informasi dasar biologi masih terbatas
(khususnya taksonomi dan reproduksinya). Indonesia (Dir Jend KSDAE 2015a). Komoditas ekspor reptil terbesar di
Mengapa V. s. macromaculatus wilayah Sumatera?
V. s. macromaculatus merupakan subspesies tersamar (sensu Welton et al. 2013b).
Wilayahnya terdiri atas Pulau Sumatera dan pulau-pulau satelitnya. Terkait sejarah geologi, pulau satelit ini ada yang terisolasi sejak terbentuknya hingga saat ini sehingga memungkinkan terjadinya spesiasi alopatrik (Voris 2000).
Sumatera merupakan lokasi sumber komoditas V. salvator.
Kajian morfologi V. s. macromaculatus:
Data morfometrik dan meristik untuk menentukan ada tidaknya variasi antara V. s. macromaculatus yang berasal dari Pulau Sumatera dan pulau satelitnya.
Pola warna untuk mengidentifikasi variasi secara visual V. s. macromaculatus yang
2
VARIASI MORFOLOGI BIAWAK AIR (
Varanus salvator
)
ASAL WILAYAH SUMATERA
Pendahuluan
Pencandraan spesies biawak, termasuk biawak air (Varanus salvator) dapat menggunakan karakter meristik (Koch et al. 2007). Telaah ulang taksonomi V. salvator berdasarkan karakter meristik berupa hitungan sisik dengan analisis klaster metode hirarki mengelompokkan V. salvator yang berasal dari Pulau Sumatera dan pulau-pulau satelit di sekitarnya menjadi satu subspesies yang sama, yaitu V. s. macromaculatus dengan asumsi rentang hitungan sisik pada V. salvator saling tumpang tindih (Koch et al. 2007). Ketumpangtindihan tersebut mengakibatkan hitungan sisik bukan sebagai karakter pendiagnosis yang mandiri (Aplin et al. 2006) sehingga diperlukan karakter lainnya.
Karakter morfologi lainnya yang digunakan untuk pencandraan biawak adalah karakter morfometrik dan pola warna. Karakter morfometrik berupa variasi ukuran tubuh umumnya digunakan untuk mencandra biawak yang berasal dari Australia bagian Barat (Pianka 1995, Thompson dan Withers 1997). Karakter morfometrik yang digunakan adalah panjang moncong hingga kloaka (Pmk),
panjang ekor (PE), panjang kepala (PK), Lebar kepala (LK), tinggi kepala (TK); panjang thoraks hingga abdomen (Pta); panjang kaki depan bagian atas (Pkda),
panjang kaki depan bagian bawah (Pkdb), panjang kaki belakang bagian atas (Pkba),
dan panjang kaki belakang bagian bawah (Pkbb). Berdasarkan proporsi karakter
tersebut, V. mertensi memiliki kepala dengan lebar yang paling sempit dan pipih, anggota tubuh dan ekor yang paling pendek dari 17 spesies yang diteliti. Pola warna tubuh biawak juga dapat digunakan untuk pencandraannya (Koch et al.
2007, Welton et al. 2014a). V. s. ziegleri memiliki supraokular berwarna gelap yang dikelilingi oleh garis terang, sedangkan organ pineal dikelilingi oleh area cokelat gelap sehingga membentuk banyak pola simetris di kepala (Koch dan
merupakan subspesies yang memiliki persebaran terluas di antara subspesies V. salvator yang ada (Gambar 1.1). Subspesies ini selain terdapat di wilayah Sumatera (Pulau Sumatera dan pulau-pulau satelitnya, misalnya Nias, Siberut, Simeulue, Bangka, dan Belitung) dan Kalimantan, juga tersebar di India, China bagian selatan (Hainan), Myanmar, Laos, Vietnam, Kamboja, Thailand, Semenanjung Malaysia, dan Borneo (de Rooij 1915, Koch et al. 2007, Koch et al.
2013). Oleh karena sebaran geografiknya yang begitu luas, V. s. macromaculatus
Metode Penelitian
Waktu dan Tempat
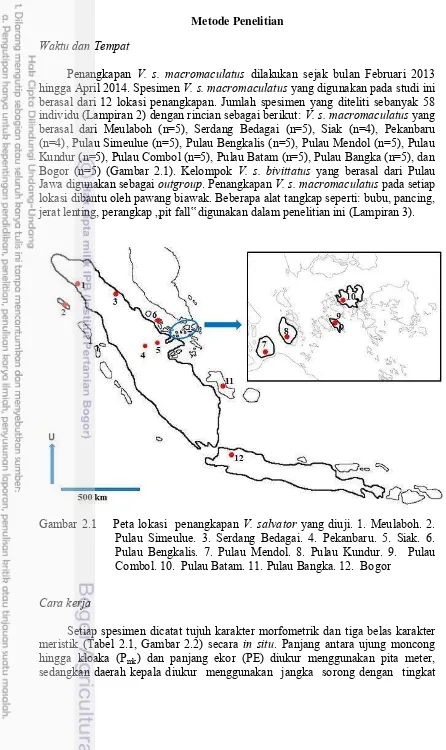
Penangkapan V. s. macromaculatus dilakukan sejak bulan Februari 2013 hingga April 2014. Spesimen V. s. macromaculatus yang digunakan pada studi ini berasal dari 12 lokasi penangkapan. Jumlah spesimen yang diteliti sebanyak 58 individu (Lampiran 2) dengan rincian sebagai berikut: V. s. macromaculatus yang berasal dari Meulaboh (n=5), Serdang Bedagai (n=5), Siak (n=4), Pekanbaru (n=4), Pulau Simeulue (n=5), Pulau Bengkalis (n=5), Pulau Mendol (n=5), Pulau Kundur (n=5), Pulau Combol (n=5), Pulau Batam (n=5), Pulau Bangka (n=5), dan Bogor (n=5) (Gambar 2.1). Kelompok V. s. bivittatus yang berasal dari Pulau Jawa digunakansebagai outgroup.Penangkapan V. s. macromaculatus pada setiap lokasi dibantu oleh pawang biawak. Beberapa alat tangkap seperti: bubu, pancing, jerat lenting, perangkap „pit fall‟ digunakan dalam penelitian ini (Lampiran 3).
Gambar 2.1 Peta lokasi penangkapan V. salvator yang diuji. 1. Meulaboh. 2. Pulau Simeulue. 3. Serdang Bedagai. 4. Pekanbaru. 5. Siak. 6. Pulau Bengkalis. 7. Pulau Mendol. 8. Pulau Kundur. 9. Pulau Combol. 10. Pulau Batam. 11. Pulau Bangka. 12. Bogor
Cara kerja
Setiap spesimen dicatat tujuh karakter morfometrik dan tiga belas karakter meristik (Tabel 2.1, Gambar 2.2) secara in situ. Panjang antara ujung moncong hingga kloaka (Pmk) dan panjang ekor (PE) diukur menggunakan pita meter,
ketelitian 0.01 cm. Jumlah sisik dihitung menggunakan pensil tumpul. Setiap spesimen (bagian dorsal, ventral, dan kepala) didokumentasikan dengan menggunakan foto digital. Hasilnya digunakan untuk mengidentifikasi pola warna
V. s. macromaculatus yang berasal dari wilayah Sumatera.
Tabel 2.1 Senarai, singkatan, dan takrif karakter morfologi yang digunakan dalam studi taksonomi V. salvator (Koch et al. 2007)
No. Singkatan Takrif karakter morfologi Karakter morfometrik
1. Pmk Panjang moncong-kloaka, yaitu jarak antara ujung moncong
dan bagian tengah kloaka.
2. PE Panjang ekor, jarak antara bagian tengah kloaka dan ujung ekor.
3. PK Panjang kepala, jarak antara ujung moncong dan tepi anterior telinga.
4. LK Lebar kepala, lebar maksimum antara dua mata dan dua telinga, yang diukur melewati kepala.
5. TK Tinggi kepala, jarak antara rahang bawah dan bagian atas mata.
6. Jmtn Jarak mata-nostril, jarak antara tepi anterior mata dan tengah
nostril.
7. Jnm Jarak nostril-moncong, jarak antara bagian tengah nostril dan
ujung moncong. Karakter meristik (hitungan sisik)
8. P Sisik melintasi kepala bagian dorsal dari ujung mulut ke ujung mulut yang lain.
9. Q Sisik kontinyu pertama mengelilingi pangkal ekor. 10. R Sisik yang mengelilingi ±1/3 bagian pangkal ekor.
11. S Sisik yang mengelilingi bagian tengah tubuh (bagian antara dua ekstremitas).
12. T Baris sisik ventral dari lipatan gular ke sisipan kaki belakang.
13. N Baris sisik ventral dari ujung moncong ke lipatan gular. 14. TN Baris sisik ventral dari ujung moncong ke sisipan kaki
belakang.
15. X Baris sisik dorsal melintang dari tepi belakang timpanum ke lipatan gular.
16. Y Baris sisik dorsal melintang dari lipatan gular ke sisipan kaki belakang.
17. XY Baris sisik dorsal melintang dari tepi timpanum belakang ke sisipan kaki belakang.
18. c Sisik supralabial kecuali satu sisik bagian tengah yang paling besar (rostral).
Gambar 2.2 Pengamatan morfometrik dan meristik V.salvator yang diuji. A.
Bagian dorsal. B. Bagian ventral. C. Bagian kepala. Singkatan karakter mengikuti Tabel 2.1
Analisis data
Karakter morfologi yang dipilih pada perlakuan pranalisis dalam mencandra biawak, yaitu: karakter morfometrik tubuh (Pmk, PE), kepala (PK, LK, TK, Jmtn,
Jnm) dan beberapa karakter meristik (P, Q, S, TN, XY, dan m). Karakter
morfometrik dianalisis dengan menggunakan rasio. Rasio yang digunakan adalah rasio antara PE dan Pmk (indeks 1), rasio antara Jmtn, dan Jnm (indeks 2), rasio
antara PK dan LK (indeks 10), rasio antara PK dan TK (indeks 11). Karakter yang tidak dianalisis adalah R karena karakter tersebut tidak selalu terukur. Karakter c dan U tidak dianalisis karena rentangnya hampir sama dan sempit. Karakter yang menyambung (kontinyu), yaitu T dan N, serta X dan Y dijumlahkan untuk
A
B
memperlebar rentang hitungan sisik. Analisis data dalam penelitian ini tidak membedakan data dari individu jantan dan betina karena di alam sulit dibedakan jenis kelamin secara langsung. Spesimen V. s. macromaculatus dikelompokkan ke dalam lima unit taksonomi operasional (UTO), yaitu: Pulau Simeulue (n=5); Pulau Sumatera (n=18) terdiri atas Meulaboh, Serdang Bedagai, Siak dan Pekanbaru; Riau Kepulauan (n=25) adalah pulau-pulau satelit Sumatera yang termasuk dalam wilayah Riau, yaitu: Pulau Bengkalis, Pulau Mendol, Pulau Kundur, Pulau Combol, dan Pulau Batam; Pulau Bangka (n=5); dan Pulau Jawa (n=5).
Korelasi Pearson digunakan untuk menguji keberadaan hubungan antara hitungan sisik dan panjang tubuh (Pmk) biawak. Kelayakan data morfometrik dan
meristik tubuh dan bagian kepala ditentukan dengan uji asumsi. Jika asumsi terpenuhi, maka dilakukan uji Analysis of Variance (ANOVA) atau Analisis Ragam Satu Arah dengan α=0.05 untuk membandingkan rerata hitungan sisik dan morfometrik antara V. s. macromaculatus dari 5 UTO. Hasil Analisis Ragam Satu Arah yang berbeda nyata diuji lebih lanjut dengan Uji Duncan untuk menunjukkan tingkat perbedaan rerata hitungan sisik dan morfometrik.
Principal Components Analysis atau Analisis Komponen Utama (AKU) terhadap karakter meristik, morfometrik tubuh, dan kepala digunakan untuk mereduksi kumpulan data karakter meristik dan morfometrik sehingga menghasilkan beberapa variabel atau komponen baru, yang memiliki nilai variansi tinggi pada ruang dua dimensi. Variabel-variabel komposit baru merupakan kombinasi linear dari variabel asli. Analisis ini menggambarkan skor objek unit taksonomi operasional sekunder dalam sistem koordinat yang diberikan oleh dua komponen utama yang paling penting. Komponen utama yang paling penting dipilih berdasarkan proporsi variansi kumulatif yang lebih dari 75%. Variabel yang memiliki nilai vektor ciri tertinggi (nilai koefisien komponen utama) adalah variabel yang digunakan.
Dendrogram karakter morfometrik dan meristik pada biawak air dibuat berdasarkan jarak menggunakan metode Euclidean dan hierarchial clustering
(hclust). Program R 3.12 digunakan dalam semua analisis statistik (R Core Team 2014).
Hasil
Morfometrik tubuh dan kepala
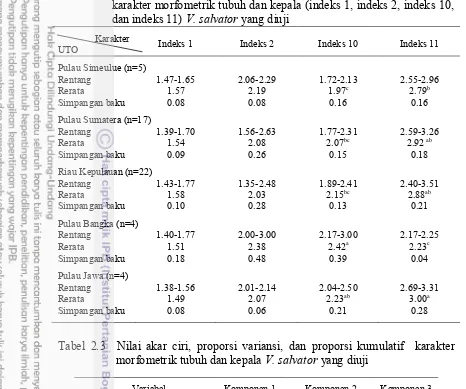
Tabel 2.2 Data rentang, rerata, simpangan baku, dan hasil uji Duncan pada karaktermorfometrik tubuh dan kepala (indeks 1, indeks 2, indeks 10, dan indeks 11) V. salvator yang diuji
Karakter Indeks 1 Indeks 2 Indeks 10 Indeks 11 UTO
Pulau Simeulue (n=5)
Rentang 1.47-1.65 2.06-2.29 1.72-2.13 2.55-2.96 Rerata 1.57 2.19 1.97c 2.79b Simpangan baku 0.08 0.08 0.16 0.16 Pulau Sumatera (n=17)
Rentang 1.39-1.70 1.56-2.63 1.77-2.31 2.59-3.26 Rerata 1.54 2.08 2.07bc 2.92 ab Simpangan baku 0.09 0.26 0.15 0.18 Riau Kepulauan (n=22)
Rentang 1.43-1.77 1.35-2.48 1.89-2.41 2.40-3.51 Rerata 1.58 2.03 2.15bc 2.88ab Simpangan baku 0.10 0.28 0.13 0.21 Pulau Bangka (n=4)
Rentang 1.40-1.77 2.00-3.00 2.17-3.00 2.17-2.25 Rerata 1.51 2.38 2.42a 2.23c Simpangan baku 0.18 0.48 0.39 0.04 Pulau Jawa (n=4)
Rentang 1.38-1.56 2.01-2.14 2.04-2.50 2.69-3.31 Rerata 1.49 2.07 2.23ab 3.00a Simpangan baku 0.08 0.06 0.21 0.28
Tabel 2.3 Nilai akar ciri, proporsi variansi, dan proporsi kumulatif karakter morfometrik tubuh dan kepala V. salvator yang diuji
Variabel Komponen 1 Komponen 2 Komponen 3
Akar ciri 1.473 1.046 0.856 Proporsi variansi 0.368 0.261 0.214 Proporsi kumulatif 0.368 0.630 0.844
Tabel 2.4 Hubungan karakter morfometrik tubuh dan kepala V. salvator yang diuji dengan unit taksonomi operasional berdasarkan analisis AKU
Gambar 2.3 AKU karakter morfometrik tubuh dan kepala V. salvator yang diuji. Simbol: ■ : Pulau Simeulue, ▲: Pulau Sumatera, ● : Riau Kepulauan, ♦ : Pulau Bangka, : Pulau Jawa. Komponen 1 memiliki proporsi variansi sebesar 36,80% dan komponen 2 memiliki proporsi variansi 26.10%
Analisis AKU menunjukkan keragaman karakter morfometrik tubuh dan kepala dapat digambarkan oleh 3 komponen utama, yaitu KU 1, KU 2, dan KU 3 yang mencapai keragaman kumulatif sebesar 84.4 (Tabel 2.3). Ketiga komponen tersebut diwakili secara berturut–turut oleh indeks 10, indeks 11, dan indeks 1 (Tabel 2.4). Hasil paduan antara komponen 1 dan 2 mengindikasikan biawak air yang berasal dari Pulau Bangka terpisah dari kelompok lainnya, tetapi kelompok yang lain saling timpang tindih pada komponen dua (Gambar 2.3).
Dendrogram karakter morfometrik tubuh dan kepala menunjukkan bahwa biawak air yang berasal dari pulau Bangka mengelompok tersendiri, tetapi kelompok yang lain masih saling tumpang tindih (Gambar 2.4). Hasil tersebut
Rentang 33.00-79.00 52-63 103-124 150-168 157-186 142-178 87-121
Rerata 57.80 54.20 108.00a 157. 60a 168.60 155.00 98.60 Keterangan: Singkatan karakter merujuk pada Tabel 2.1. Hasil uji Duncan karakter meristik
menunjukkan berbeda nyata jika angka dalam satu kolom diikuti dengan huruf tika atas yang berbeda. Karakter hitungan sisik dianggap konsisten jika dapat membedakan satu dengan setiap lainnya yang diuji.
dapat digunakan sebagai karakter pengidentifikasi spesies karena karakter tersebut tidak terpengaruh oleh ukuran tubuh (Pmk).
Hasil analisis statistik pada karakter meristik biawak air secara terbatas menunjukkan adanya keterpisahan di antara unit taksonomi biawak air yang diuji. Karakter Q dan S merupakan karakter yang penting (Tabel 2.5 dan Lampiran 6). Karakter Q dapat membedakan kelompok biawak air yang berasal dari Pulau Simeulue dan Pulau Bangka dengan biawak air yang berasal dari Riau Kepulauan dan Pulau Sumatera. Karakter S tidak dapat membedakan antara kelompok biawak air yang berasal dari Pulau Simeulue dan Sumatera, antara kelompok yang berasal Pulau Bangka dan Pulau Jawa.
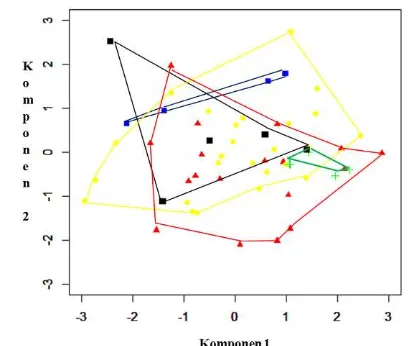
Keragaman karakter hitungan sisik dapat digambarkan oleh 3 komponen utama, yaitu KU1, KU2, dan KU3 yang mencapai keragaman kumulatif sebesar 80.2 (Tabel 2.6). Ketiga komponen tersebut diwakili secara berturut–turut oleh karakter XY, TN, dan Q (Tabel 2.7). Hasil paduan antara komponen 1 dan 2 mengindikasikan biawak air yang berasal Pulau Bangka sedikit terpisah dari Pulau Simeulue, tetapi tumpang tindih dengan kelompok lainnya (Gambar 2.5). Tabel 2.6 Nilai akar ciri, proporsi variansi, dan proporsi kumulatif karakter
hitungan sisik V. salvator yang diuji
Variabel Komponen 1 Komponen 2 Komponen 3 Akar ciri 3.228 1.038 0.546 Proporsi variansi 0.538 0.173 0.091 Proporsi kumulatif 0.538 0.711 0.802
Tabel 2.7 Hubungan karakter meristik V. salvator yang diuji dengan unit taksonomi operasional (UTO) berdasarkan analisis AKU
Variabel Komponen 1 Komponen 2 Komponen 3 P 0.419 0.042 -0.192 Q 0.402 0.372 -0.624
S 0.370 0.576 0.416 TN 0.358 -0.625 0.183
XY 0.462 0.005 0.524
m 0.430 -0.370 -0.304 Keterangan: Nilai koefisien komponen utama tertinggi pada komponen 1, 2 dan 3 dipilih
(ditebalkan) untuk menjelaskan karakter pembeda diantara UTO.
Dendrogram karakter hitung sisik menggunakan metode Euclidean dan
Gambar 2.5 AKU karakter meristik V. salvator yang diuji. Simbol + : Pulau Simeulue, ▲: Pulau Sumatera, ● : Riau Kepulauan, ■ : Pulau Bangka, ■ : Pulau Jawa. Komponen 1 memiliki proporsi variansi sebesar 53.80% dan komponen 2 memiliki proporsi variansi 17.30%
Pola warna
Biawak air yang berasal dari wilayah Sumatera terbagi dalam dua kelompok berdasarkan pola warna ventral dan dorsalnya (Tabel 2.8, Gambar 2.7, dan Lampiran 7). Pola warna ventral maupun dorsal kelompok pertama terdiri atas individu-individu yang berasal dari Riau Kepulauan (Pulau Bengkalis, Pulau Mendol, Pulau Combol, Pulau Kundur, dan Pulau Batam) dan Pulau Bangka menyerupai pola warna kelompok yang berasal dari Pulau Sumatera (Meulaboh, Serdang Berdagai, Pekanbaru, dan Siak), dalam hal ini diwakili individu yang berasal dari Siak. Pola warna tubuh kelompok pertama tidak memiliki garis melintang gelap pada bagian ventralnya (Gambar 2.5A), tetapi memiliki ocelli
(bintik terang) yang cenderung terpisah pada baris pertama garismelintang dorsal tubuhnya (Tabel 2.5 dan Gambar 2.5B). Biawak air yang berasal dari Pulau Simeulue menunjukkan pola warna tubuh yang berbeda dari kelompok pertama. Pola warna bagian ventral tubuhnya memiliki beberapa garis melintang gelap (Gambar 2.5C), sedangkan pada bagian dorsalnya terdapat bintik terang yang unik (kecil, tidak berbentuk lingkaran) dengan jarak antargaris melintang yang lebih jauh (Gambar 2.5 D). Secara umum, pola warna bagian dorsal biawak air yang berasal dari wilayah Sumatera cenderung berbeda dari pola warna biawak air yang berasal dari Pulau Jawa. Pola warna pada bagian dorsal V. salvator yang berasal dari Pulau Jawa terdapat bintik terang yang cenderung menyatu pada baris pertama garis melintang (Tabel 2.8, Gambar 2.5E) dan pada bagian ventral tanpa garis melintang gelap. Sebagai catatan, ada dua individu yang berasal dari Serdang Bedagai, Sumatera Utara yang teramati hampir tanpa pola pada bagian dorsal dan teridentifikasi ditemukan di pinggiran aliran sungai dengan vegetasi yang rimbun.
Tabel 2.8 Persentase V. salvator yang diuji berdasarkan pola warna Pola warna
C: bintik terang (ocelli) pada garis pertama tidak menyatu D: bintik terang pada garis pertama menyatu
I : V. salvator yang berasal dari Meulaboh, Serdang Bedagai, Pekanbaru, Siak, Pulau Bengkalis, Pulau Mendol, Pulau Combol, Pulau Kundur: Pulau Batam, dan Pulau Bangka
II : V. salvator yang berasal dari Pulau Simeulue III: V. salvator yang berasal dari Bogor (Pulau Jawa).
Gambar 2.7 Pola warna ventral dan dorsal tubuh V. salvatoryang diuji. A. Tanpa garis melintang gelap. B. Bintik terang terpisah pada garis melintang pertama. C. Garis melintang gelap. D. Jarak antargaris melintang jauh. E. Bintik terang menyatu pada garis melintang pertama
Pembahasan
V. salvator jantan dan betina yang diamati di alam adalah monomorfik, tetapi ada beberapa metode yang membantu penentuan seks. Berdasarkan ukuran tubuh, ukuran tubuh individu jantan lebih besar daripada betina pada umur yang sama (Shine et al. 1996), tetapi untuk biawak yang diperoleh dari alam sulit menentukan umurnya secara langsung. Penggunaan cloaca probe untuk menentukan adanya hemipenis pada jantan dapat memberikan hasil yang membingungkan karena betina memiliki hemiclitoral sacculae (Böhme 1995). Beberapa biawak air betina dijumpai lebih kusam dibandingkan yang jantan.
Karakter morfometrik berupa indeks 10 dan indeks 11 merupakan karakter diagnostik dari kelompok biawak air yang berasal dari Pulau Bangka dengan kelompok lainnya. Hasil ini mengindikasikan bahwa kepala biawak air yang berasal dari Pulau Bangka memiliki rerata panjang kepala relatif lebih panjang terhadap lebar kepala, dan rerata panjang kepala relatif lebih pendek terhadap tinggi kepala sehingga kepalanya tampak lebih panjang dan pipih dibandingkan kelompok lainnya. Kelompok biawak air yang berasal dari Pulau Sumatera, Riau Kepulauan, Pulau Simeulue, dan Pulau Jawa menunjukkan ketumpangtindihan karakter meristik.
Hitungan sisik Q dan S tidak berkorelasi dengan ukuran tubuh (Pmk)
berdasarkan uji korelasi Pearson) sehingga karakter Q dan S dapat digunakan sebagai karakter pengidentifikasi spesies V. salvator. Tidak berkorelasinya karakter Q dan S dengan ukuran tubuh dapat dikatakan sebagai age–independent, yaitu tidak bergantungnya karakter ini pada umur individu, seperti dicontohkan pada dua spesies biawak Australia, V. caudolineatus dan V. gilleni (Aplin et al.
Karakter Q dan S merupakan karakter meristik penting berdasarkan Uji Duncan. Jika dibandingkan dengan spesies biawak lainnya, hitungan sisik Q pada kelompok V. salvator yang berasal dari Pulau Simeulue (rerata 92 ± 2) tumpang tindih dengan hitungan sisik Q pada V. marmoratus (rerata 91 ± 7) (Welton et al.
2014a). Ketumpangtindihan juga ditunjukkan hitungan sisik Q antara V. salvator
yang berasal dari Pulau Bangka (rerata 108 ± 9) dan V. ramusseni yang berasal dari kepulauan Sulu, Filipina (114.50 ± 3.54) (Koch et al. 2010b). Karakter S biawak air yang berasal dari Pulau Bangka dan Pulau Jawa (157.40 ± 8.23) tumpang tindih dengan karakter S pada V. ramusseni (154.50 ± 3.54) (Koch et al.
2007, Koch dan Böhme 2010, Koch et al. 2010b, Welton et al. 2014a). Ketumpangtindihan hitungan sisik Q dan S dengan kata lain menempatkan individu-individu pada ruang morfologi yang agak berbeda.
Pola warna bagian dorsal dan ventral tubuh pada kelompok V. salvator yang berasal dari Pulau Simeulue merupakan karakter diagnostik terhadap V. s. macromaculatus yang berasal dari Pulau Sumatera, Riau Kepulauan, dan Pulau Bangka. Karakter pola warna bagian ventral pada V. salvator yang berasal dari dari Pulau Simeulue diduga terkait dengan terisolasinya Pulau Simeulue dari Pulau Sumatera (Voris 2000) dan adanya adaptasi morfologi spesies tersebut pada habitatnya. Biawak air di Pulau Simeulue teramati di daerah rawa nipah yang bervegetasi rapat. Sebaliknya individu-individu biawak air yang berasal dari Pulau Sumatera, Riau Kepulauan, dan Pulau Bangka cenderung terdapat di daerah rawa dekat aliran sungai dan daerah pinggir pantai tanpa naungan sehingga cenderung memiliki bintik terang berbentuk cincin yang besar. Pola warna yang sama ditemukan pada V. s. macromaculatus yang berasal dari Thailand, yang terdapat di daerah cekungan sungai Chao Phraya (Cota et al. 2009). Pola warna pada kelompok V. salvator yang berasal dari Pulau Bangka yang belum menunjukkan perbedaan mengindikasikan bahwa keterpisahan tersebut mungkin belum terlalu lama dibandingkan dengan biawak air yang berasal dari Pulau Simeulue.
Variasi pola warna tubuh pada V. s. macromaculatus yang berasal dari wilayah Sumatera pada lingkungan berbeda disokong oleh dorongan selektif. Ada dua hipotesis utama, yaitu kripsis dan termoregulasi. Pola warna tersamar berfungsi sebagai penurunan predasi, suatu perilaku melarikan diri secara pasif (antipredator) dan peningkatan efisiensi pencarian makan. Gabungan efek tersebut dapat meningkatkan kesintasan setelah deteksi awalbiawak air terhadap predator. Pola warna tubuh bagian dorsal dengan bintik terang yang lebih besar dan berwarna terang pada biawak diduga berfungsi agar lebih efisien memantulkan kelebihan panas yang diterima pada lingkungan yang kering (sensu Willmer et al.
Simpulan
3
KARAKTERISTIK GENETIKA MOLEKULER BIAWAK
AIR (
Varanus salvator
) ASAL WILAYAH SUMATERA
BERDASARKAN GEN COI
Pendahuluan
Biawak air (Varanus salvator) tergolong spesies tersamar (Koch et al. 2007, Welton et al. 2014a). Divergensi morfologi pada spesies tersamar tidak sejalan dengan divergensi genetik (Lorenz et al. 2005). Hal tersebut mengindikasikan bahwa kelompok taksa yang terdapat dalam spesies tersamar sangat mirip secara morfologi, secara molekuler berbeda.
Salah satu spesies biawak air Filipina, V. marmoratus, teridentifikasi sebagai spesies tersamar (Welton et al. 2013b). Telaah ulang V. marmoratus yang berasal dari Mindoro dan Semenanjung Bicol, Filipina menghasilkan tambahan dua spesies baru, yaitu V. bangonorum dan V. dalubasha secara berturut-turut di samping spesies lamanya, yaitu V. marmoratus yang berasal dari Pulau Luzon. Dua spesies baru tersebut secara morfologi hampir tidak dapat dibedakan, tetapi dapat teridentifikasi berdasarkan posisi mereka pada estimasi filogeni multilokus yang menggunakan lokus mitokondria (ND1, ND2) dan lokus inti (DGL-α, PRLR) (Welton et al. 2014a). V. bangonorum dan V dalubhasa merupakan hasil spesiasi alopatrik akibat adanya isolasi geografi. Penggunaan penanda genetik yang lain, yaitu gen COI telah berhasil digunakan untuk mengidentifikasi spesies-spesies biawak yang termasuk di dalam kelompok V. salvator ssp. (Arida 2014).
V. s. macromaculatus yang berasal dari wilayah Sumatera menarik untuk ditelaah ulang karena subspesies ini memiliki penyebaran terluas (Gambar 1.1) dan tergolong subspesies tersamar (Welton et al. 2013b). Pada studi sebelumnya, biawak air yang berasal dari Pulau Simeulue dan Pulau Bangka secara morfologi teridentifikasi sebagai morfo-spesies yang berbeda dari biawak air yang berasal dari Pulau Sumatera dan Riau Kepulauan (Bab 2). Oleh karena itu, karakterisasi molekuler berdasarkan gen COI perlu dilakukan untuk mengonfirmasi status taksa
V. s. macromaculatus yang berasal dari wilayah Sumatera yang telah berdifferensiasi secara morfologi apakah secara molekuler masih dalam kelompok yang sama atau telah berbeda pula secara molekuler.
Bahan dan Metode
Waktu dan Tempat Penelitian
Bahan
Setiap individu V. salvator yang tertangkap per lokasi pengambilan sampel (Lampiran 2) diambil 0.5-1.0 ml contoh darah di sekitar sepertiga pangkal ekor dengan menggunakan spuit. Spuit ditusukkan pada bagian ventral ekor pada posisi dua atau tiga baris ke kanan atau kiri dengan arah menuju median. Contoh darah tersebut dipreservasi dengan etanol pure grade dengan perbandingan 1 : 5 dan segera dikocok untuk menjamin semua darah dipreservasi dengan baik. Setelah itu, individu biawak air dilepaskan ke habitatnya.
Cara kerja
Sampel darah V. salvator yang telah dikoleksi diambil sebanyak ± 25 mg, dimasukkan ke dalam tabung Eppendorf (Eppendorf tube) 1.5 ml, dicuci dengan menggunakan Tris-asam etilena diamina tetra asetat (EDTA) konsentrasi rendah a t a u d ik e na l low TE. Tabung tersebut divorteks selama10 menit dan disentrifugasi 4500 rpm (revolutions per minute) selama tiga menit. Langkah ini diulang sebanyak tiga kali dengan tujuan untuk menghilangkan kandungan alkohol yang terdapat di dalam sampel jaringan V. salvator. DNA total diekstrak dari sampel jaringan dengan menggunakan Dneasy ® Blood & Tissue Kit cat no 69504 berdasarkan Spin-Column Protocol.
Kualitas DNA dilihat dengan dimigrasikan pada gel agarosa 1.2% dengan menggunakan buffer 1 x TBE (89 mM Tris, 89 mM asam borat dan 2 mM EDTA pH 8.0). Gel agarosa diwarnai dengan etidium bromida (0.5 g/ml). Pengamatan kualitas DNA dilakukan dengan bantuan alat foto dengan sinar Ultra Violet (UV) dengan ukuran 300 nm. DNA total yang dihasilkan digunakan sebagai cetakan DNA untuk proses amplifikasi.
Fragmen DNA pada daerah COI diamplifikasi dengan menggunakan sepasang primer dari Nagy et al. (2012) dengan sedikit modifikasi, yaitu VS-COI_F (5‟-TCTTCTCCACAAACCACAAAGA-3‟), dan VS-COI_R (5‟ACCTC TGGGTGTCCGAAGAATCA-3‟) pada kondisi Polymerase Chain Reaction
(PCR) yang digunakan sebagai berikut: predenaturasi dengan suhu 94 ºC selama 3 menit, 35 siklus dari (94 ºC selama 45 detik untuk denaturasi, 56 ºC selama 30 detik untuk penempelan primer dengan cetakan DNA (annealing), 72 ºC selama 1 menit untuk elongasi, 72 ºC selama 7 menit untuk elongasi akhir), dan inkubasi dengan suhu 15 ºC selama 40 menit. Larutan reaksi PCR dibuat dalam 25 µl dengan komponen dan konsentrasi akhir terdiri atas: 5 µl buffer, 5 µl enhance, 1 µl dNTP, 1 µl primer VS-COI_F dan VS-COI_R 10 pmol, 3-5 µl DNA, dan 0.2 µl TaqH5.
Produk PCR dielektroforesis melalui gel agarosa 1.2% dan divisualisasikan dengan pewarnaan etidium bromida. Pita tunggal pada gel agarose sebagai produk PCR dijadikan sebagai cetakan dalam reaksi pengurutan basa nukleotida dengan menggunakan primer VS-COI_F dan VS-COI_R. Purifikasi dan perunutan hasil amplifikasi dilakukan oleh perusahaan jasa 1st BASE) di Malaysia melalui PT Genetica Science.
dihasilkan berukuran 664 pb (mengacu pada urutan basa nukleotida COI utuh V. salvator yang berasal dari Thailand dengan nomor akses AB980995 dan AB980996 dengan panjang nukleotida total 1596 pb). Semua urutan basa nukleotida tersebut dijajarkan berganda, beberapa situs ambigu ditemukan dan disunting secara manual. Runutan DNA yang diperoleh dari hasil penjajaran berganda tersebut adalah 516 nukleotida (Gambar 3.1), yaitu pada posisi ke 184 sampai dengan posisi ke 699 (mengacu pada COI utuh V. salvator dari GenBank). Bagian urutan basa nukleotida fragmen yang tidak terbaca, yaitu 148 nukleotida dari ujung 5‟ primer VS-COI_F. Hasil suntingan ditranslasi untuk memverifikasi bahwa sekuen hasil suntingan bebas dari stop codon atau kodon penerminalan. Setiap urutan basa nukleotida diverifikasi dengan melakukan Basic Local Alignment Search Tool (BLAST) pada GenBank (wwww.ncbi.nlm.nih.gov) untuk menentukan ada tidaknya gap dan mengidentifikasi kemiripan genetik dengan urutan nukleotida utuh gen COI V. salvator yang ada di GenBank.
Gambar 3.1 Skema posisi penempelan primer VS-COI_F dan VS-COI_R pada produk amplifikasi
Data urutan basa nukleotida yang teridentifikasi memiliki kemiripan dengan gen COI utuh V. s. macromaculatus yang berasal dari Thailand (nomor akses AB980995 dan AB980996) lebih dari 97% digunakan dalam analisis selanjutnya. Urutan basa nukleotida gen COI parsial Varanus bengalensis yang berasal dari India (kode akses KF766939.1) digunakan sebagai outgroup. Beberapa karakter urutan basa nukleotida gen COI parsial pada V. salvator yang dianalisis, yaitu komposisi basa, keragaman nukleotida tiap kelompok, dan jumlah haplotipe. Karakter tersebut dihitung menggunakan MEGA 6 dan DNA SP versi 5 (Librado dan Rozas 2009). Jaring haplotipe dibuat menggunakan program Network 5.0.0.0.
penelitian ini didasarkan pada rerata jarak genetik bersih antarkelompok. Rerata jarak genetik bersih antarkelompok XY adalah rerata jarak genetik antarkelompok XY dikurangi (rerata jarak genetik dalam kelompok X + rerata jarak genetik dalam kelompok Y)/2). Jarak genetik antar spesies secara umum ditentukan lebih dari 2% yang setara dengan perubahan 14-bp pada panjang sekuen DNA 650-bp (Hebert et al. 2003, Ward 2009).
Metode Neighbor-Joining (NJ) berdasarkan p distance dipilih dalam simulasi konstruksi pohon karena metode ini biasa digunakan pada metode pengelompokan pada beberapa studi DNA (Ratnasingham dan Hebert 2007) dan direkomendasikan sebagai metode standar yang minimum (St. John et al. 2003). Konstruksi pohon NJ dengan model p distance dan butstrap 1000 kali dilakukan dengan menggunakan MEGA 6 (Tamura et al. 2013). Urutan basa nukleotida yang diperoleh akan didepositkan di National Center for Biotechnology Information atau NCBI.
Hasil
Identitas urutan basa nukleotida COI V. salvatorasal wilayah Sumatera
Urutan basa nukleotida sebesar 80% dari total sampel berhasil diamplifikasi (Lampiran 1). Insersi dan delesi tidak terjadi dalam penjajaran urutan basa nukleotida gen COI. Hasil verifikasi setiap urutan basa nukleotida menunjukkan rentang kesamaan urutan basa nukleotida 97-100% dengan urutan basa nukleotida
V. salvatordari GenBank.
Hasil analisis fragmen gen COI sepanjang 516 pb pada semua individu V. salvator yang diteliti terdapat 413 situs conserved (80.04%), 103 situs variable
(19.96%), 28 situs parsimoni informatif (5.43%), 75 situs singleton (14.53%). Proporsi basa nukleotida dari penjajaran urutan basa nukleotida COI pada semua individu yang diteliti menunjukkan persentase basa timin (T) = 26.78%, sitosin (C) = 32.40%, adenin (A) = 25.06% dan guanin (G) = 15.76% (Lampiran 8). Dengan demikian timin + adenin (TA) sebesar 51.84% dan sitosin + guanin (GC) = 48.16% sehingga kandungan GC < AT dan relatif seimbang.
Substitusi basa nukleotida pada studi ini yang paling sering terjadi adalah pada kodon ketiga (78.64%), tetapi hal sebaliknya terjadi pada kodon kedua (8.74%). Hasil ini konsisten dengan pernyataan bahwa gen penyandi protein memiliki kodon ketiga paling variabel, sedangkan kodon kedua paling lestari (Xia et al. 1996, Xia 1998). Substitusi basa nukleotida yang bersifat transisi (78.64%) terjadi lebih banyak dibandingkan yang bersifat transversi (21.36%).
Polimorfisme V. salvator asal wilayah Sumatera berdasarkan gen COI
Tabel 3.1 Haplotipe 52 individu V. salvator yang diuji Kode
Haplotipe Identitas spesimen Kelompok individu Jumlah Hap_1 MB1, MB5, PKU1, PKU3, PKU4, PKU5, BTM3,
Keterangan: Singkatan identitas spesimen merupakan lokasi pengambilan sampel (Lampiran1). Angka pada kode sampel menunjukkan nomor individu yang diuji.
Tabel 3.2 Situs substitusi nukleotida khas sebagai penciri kelompok-kelompok pada V. salvator yang diuji (huruf yang ditebalkan dan diarsir)
ID Klpk