EKSTRAKSI SENYAWA POLIFENOL MELALUI DEGRADASI
BIOMASSA LIGNOSELULOSA KULIT KOPI MENGGUNAKAN
KONSORSIUM AKTINOMISET
NURLENI KURNIAWATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul “Ekstraksi Senyawa Polifenol Melalui Degradasi Biomassa Lignoselulosa Kulit Kopi Menggunakan Konsorsium Aktinomiset” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2015
Nurleni Kurniawati
RINGKASAN
NURLENI KURNIAWATI. Ekstraksi Senyawa Polifenol Melalui Degradasi Biomassa Lignoselulosa Kulit Kopi Menggunakan Konsorsium Aktinomiset. Dibimbing oleh ANJA MERYANDINI dan TITI CANDRA SUNARTI.
Kulit kopi merupakan limbah dari pengolahan primer buah kopi yang tinggi akan kandungan senyawa metabolit sekunder yaitu polifenol; yang terdiri atas tanin, flavonol, katekin, asam hidroksimat, dan antosianin. Senyawa polifenol dapat digunakan sebagai antioksidan, pewarna alami makanan, dan mengatasi penyakit diabetes serta kardiovaskular. Polifenol yang terdapat pada kulit kopi dilindungi oleh komponen serat yaitu selulosa, hemiselulosa, dan lignin. Untuk melepaskan senyawa polifenol tersebut maka perlu dilakukan degradasi komponen serat yang dapat dilakukan oleh enzim selulase, xilanase, dan peroksidase yang dihasilkan oleh bakteri melalui kultivasi. Pada penelitian ini enzim selulase, xilanase, dan peroksidase dapat dihasilkan oleh aktinomiset
Streptomyces exfoliatus 42, sedangkan xilanase yang tinggi dihasilkan oleh
Streptomyces costaricanus 45I-3 melalui fermentasi atau kultivasi media padat. Oleh karena itu penelitian ini bertujuan untuk meningkatkan perolehan ekstrak senyawa polifenol dengan mendegradasi komponen lignoselulosa kulit kopi menggunakan konsorsium isolat aktinomiset dan kultivasi media padat.
Dibandingkan dengan fermentasi spontan buah kopi, penambahan kultur konsorsium aktinomiset mampu mempercepat degradasi komponen lignoselulosa kulit kopi, dan secara signifikan berpengaruh pada hasil senyawa bioaktif seperti polifenol, antosianin, tanin, dan katekin. Penurunan tertinggi komponen lignoselulosa terjadi setelah hari ke-6 fermentasi yaitu 29.11% selulosa, 4.19% lignin, dan hari ke-9 sebesar 17.56% untuk hemiselulosa. Perolehan hasil ekstrak tertinggi dari komponen ekstrak bioaktif terjadi pada inkubasi setelah hari ke-6 yaitu total polifenol (1.20 mg mL-1), antosianin (109.95 mg mL-1) dan katekin pada inkubasi hari ke-3 (10.38 mg mL-1). Penggunaan konsorsium Streptomyces
sp. selama fermentasi juga dapat menurunkan perolehan ekstrak tanin, sehingga dapat meningkatkan pemanfaatan kulit kopi yang terfermentasi sebagai pakan ternak. Dengan demikian penggunaan konsorsium aktinomiset dapat meningkatkan perolehan ekstrak polifenol dan meningkatkan nilai guna limbah kulit kopi.
SUMMARY
NURLENI KURNIAWATI. Extraction of Polyphenol Compounds by Lignocellulose Biomass Degradation of Coffee Pulp Using Actinomycetes Concortia. Supervised by ANJA MERYANDINI and TITI CANDRA SUNARTI.
Coffee pulp is a by-product from coffee primary processing unit which contains high concentration of secondary metabolite compounds like polyphenols i.e tannins, flavonols, catechins, hydroxycinamic acid and anthocyanins. The compounds can be used as antioxidants and natural food dyes, which can be used to overcome diabetic, and cardiovascular symphtoms. Polyphenols in coffee pulp are binding by cellulose, hemicellulose, and lignin fiber components. Polyphenol can be easily released by the degradation of fiber components by cellulase, xylanase, and peroxidase which resulting from cultivation of bacteria by using solid state fermentation. On this study, the cellulase, xylanase, and peroxidase can be produced by Streptomyces exfoliatus 42, whereas the high xylanase produced by Streptomyces costaricanus 45I-3. The aim of this study is to enhance the yield of polyphenol extract by degrading the fiber component using concortia of
Streptomyces exfoliatus 42 and Streptomyces costaricanus 45I-3.
Compared to spontaneous fermentation of coffee cherries, the introduction of actinomycetes starter accelerated the fermentation which cause increasing the degradation of lignocellulose components of coffee pulp, and significantly effects on the yield of bioactive compounds mainly for polyphenols, anthocyanins, tannins, and catechins contents. The highest decreasing in the lignin and cellulose levels in coffee pulp was obtained after 6th days as 4.19% of lignin, 29.11% of cellulose, and after 9th days for hemicellulose (16.89%). The highest yield of bioactive components extracts produced from 6th days incubation for total polyphenols (1.20 mg mL-1), anthocyanin (109.95 mg mL-1) and the highest yield of catechin (10.38 mg mL-1) produced form 3th days incubation, and introduction of actinomycetes also reduced the tannin contents after fermentation. This made the fermented coffee pulp were suitable as feed source.
Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
EKSTRAKSI SENYAWA POLIFENOL MELALUI DEGRADASI BIOMASSA LIGNOSELULOSA KULIT KOPI MENGGUNAKAN
KONSORSIUM AKTINOMISET
NURLENI KURNIAWATI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Bioteknologi
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Tesis : Ektraksi Senyawa Polifenol Melalui Degradasi Biomassa Lignoselulosa Kulit Kopi Menggunakan Konsorsium Aktinomiset Nama : Nurleni Kurniawati
NIM : P051120121
Disetujui oleh Komisi Pembimbing
Diketahui oleh
Ketua Program Studi Bioteknologi
Prof Dr Ir Suharsono, DEA
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian : 5 Juni 2015 Tanggal Lulus :
Prof Dr Anja Meryandini, MS Dr Ir Titi Candra Sunarti, MSi
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari 2014 sampai Desember 2014 ialah Ekstraksi Senyawa Polifenol Melalui Degradasi Biomassa Lignoselulosa Kulit Kopi Menggunakan Konsorsium Aktinomiset.
Terima kasih penulis ucapkan kepada Prof Dr Anja Meryandini, MS sebagai ketua komisi pembimbing dan Dr Ir Titi Candra Sunarti, MSi sebagai anggota komisi pembimbing yang telah banyak memberikan ilmu, saran, motivasi, nasehat waktu konsultasi, serta solusi dari setiap permasalahan yang dihadapi penulis selama melaksanakan penelitian dan penyusunan karya ilmiah ini. Selain itu penulis ucapkan terima kasih kepada penguji luar komisi Dr Suryani, SP MSc, dan Prof Dr Ir Suharsono, DEA selaku ketua Program Studi Bioteknologi IPB yang telah memberikan masukan pada saat ujian sidang tesis, serta motivasi selama studi. Kepada DIKTI melalui Beasiswa Unggulan selama menempuh pendidikan pascasarjana di IPB, dan terima kasih atas hibah penelitian tahun 2014 a.n. Dr Ir Titi Candra Sunarti, MSi sehingga penelitian yang penulis lakukan dapat terlaksana dengan baik.
Penghargaan penulis sampaikan kepada staf dan laboran di laboratorium Bioteknologi Hewan dan Biomedis, PPSHB IPB dan laboratorium Kimia Analitik IPB yang telah membantu selama penelitian. Penulis juga mengucapkan terima kasih kepada Rike Tri KD, Nurkholis, Yeni, Tika, Hamtini, Ike Apriani, Anik, Novianty, Rahmi, Lia, Ika, Debi, Ira, Fithe, Wahyu, Wida, Fitri, Nuril serta seluruh teman-teman di Laboratorium Kimia Analitik IPB, atas dukungan, motivasi, dan bantuannya selama penelitian. Ucapan terima kasih tak terhingga juga penulis ucapkan kepada bapak Darmanto SPd, ibu Sumarni, Adek Nurhavid Kurniawan tersayang, dan Tomi Sutrisno, SSi atas doa, dukungan, kasih sayang, motivasi, dan semangat yang diberikan. Terima kasih untuk teman-teman seperjuangan di Sekolah Pascasarjana Bioteknologi IPB angkatan 2012 serta seluruh pihak yang telah memberikan doa dan dukungannya, penulis ucapkan terima kasih.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2015
DAFTAR ISI
DAFTAR TABEL ii
DAFTAR GAMBAR ii
DAFTAR LAMPIRAN ii
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Manfaat Penelitian 2
Ruang Lingkup Penelitian 3
TINJAUAN PUSTAKA
Kulit Kopi 3
Biomassa Lignoselulosa 4
Aktinomiset 6
Senyawa Bioaktif di Kulit Kopi 6
METODE 7
Waktu dan Tempat Penelitian 7
Alat dan Bahan 7
Karakterisasi Bahan Baku 7
Peremajaan Isolat dan Produksi Enzim 8
Degradasi Komponen Serat Pada Biomassa Kulit Kopi
Persiapan Substrat dan Kultur Starter 8
Proses Kultivasi 8
Kinerja Hasil Degradasi 9
Analisis Data Penelitian 9
HASIL PEMBAHASAN
Karakterisasi Bahan Baku 10
Produksi Enzim 11
Degradasi Biomassa Lignoselulosa Kulit Kopi 13
SIMPULAN 21
DAFTAR PUSTAKA 22
DAFTAR TABEL
1 Komponen kimia pada kulit kopi 9
2 Komposisi komponen substrat kulit kopi setelah kultivasi pada suhu
27 °C 15
3 Kandungan gula hasil ekstraksi setelah kultivasi pada suhu 27 °C 18 4 Komposisi perolehan hasil ekstraksi senyawa bioaktif kulit kopi setelah
kultivasi pada suhu 27 °C 19
DAFTAR GAMBAR
1 Struktur buah kopi 3
2 Struktur dinding sel selulosa, hemiselulosa, dan lignin pada
lignoselulosa 4
3 Aktivitas dan pertumbuhan Streptomyces exfoliatus 42 pada media
sintetik xilan dan CMC 11
4 Aktivitas dan pertumbuhan Streptomyces costaricanus 45I-3 pada
media sintetik xilan 11
5 Aktivitas enzim oleh konsorsium aktinomiset pada media kolit kopi 1% 13 6 Persentase susut bobot substrat setelah kultivasi metode padat inkubasi
pada suhu 27 °C 15
7 Persentase penurunan komponen serat setelah kultivasi pada 27 °C 16
DAFTAR LAMPIRAN
1 Analisis proksimat 29
2 Komponen serat 30
3 Kandungan gula 32
4 Kandungan senyawa bioaktif 32
5 Aktivitas enzim selulase Streptomyces exfoliatus 42 34 6 Aktivitas enzim xilanase Streptomyces exfoliatus 42 dan Streptomyces
costaricanus 45I-3 34
7 Kurva standar xilosa dan glukosa 35
8 Bobot komponen serat 36
1
PENDAHULUAN
Latar Belakang
Kopi merupakan komoditi perkebunan yang sangat penting di dunia. Ada sekitar 60 negara penghasil kopi, dan Indonesia menempati posisi ketiga setelah Brasil dan Vietnam (FAO 2012). Berdasarkan Ditjenbun (2012) produksi kopi Indonesia pada luas lahan 1 240 919 ha mencapai 748 109 ton pertahun. Buah kopi tidak bisa dikonsumsi langsung, namun perlu diproses menjadi biji kopi kering dengan metode pengolahan basah, kering dan semi kering (Schewan et al.
2012). Metode basah merupakan metode yang paling umum digunakan untuk pengolahan kopi. Hal ini karena memerlukan waktu yang relatif lebih singkat dibanding dengan metode kering dan semi kering. Metode pengolahan basah dilakukan melalui pengupasan kulit terluar secara mekanik, selanjutnya biji kopi difermentasi secara spontan. Fermentasi tersebut bertujuan untuk menghidrolisis lapisan lendir yang menempel pada bagian biji kopi selama proses fermentasi. Proses fermentasi spontan dilakukan oleh mikrob yang kompleks yang melibatkan beberapa mikroorganisme seperti khamir, bakteri, dan cendawan (Silva et al.
2008). Namun fermentasi spontan ini kurang efektif karena menghasilkan produk biji kopi yang kurang seragam.
Penambahan kultur bakteri dan khamir umumnya dilakukan untuk mengatasi permasalahan pada proses fermentasi spontan. Silva et al. (2013) melaporkan bahwa kultur starter bakteri yang ditambahkan dapat mengoptimalkan proses fermentasi sehingga proses fermentasi berjalan lebih cepat dan meningkatkan kualitas hasil biji kopi fermentasi. Mikroorganisme spesifik yang dipilih untuk kultur starter selama proses fermentasi kopi penting untuk meningkatkan kualitas proses fermentasi dan untuk meningkatkan kualitas sensorik dari minuman kopi (Massawe dan Lifa 2010). Fermentasi spontan juga masih menyisakan permasalahan yaitu 40% limbah yang belum termanfaatkan dengan baik. Limbah tersebut terdiri atas limbah cair sisa fermentasi dan limbah padat berupa biomassa kulit kopi. Biomassa kulit kopi merupakan biomassa lignoselulosa dengan kandungan serat tinggi. Komponen serat pada kulit kopi yaitu 49% selulosa, 24.5% hemiselulosa, dan lignin 7.63% (Diniyah et al. 2013).
Komponen lignoselulosa dapat didegradasi oleh agen biologi seperti aktinomiset. Beberapa aktinomiset mampu menggunakan polisakarida seperti pati, selulosa, hemiselulosa sebagai nutrisi untuk metabolismenya karena dapat menghasilkan enzim hidrolitik ekstraseluler (Kokulya et al. 2002). Berdasarkan penelitian Tuncer et al. (2004) diketahui bahwa Streptomyces sp. F2621 menghasilkan enzim peroksidase, xilanase, dan endoglukanase. Keuntungan dari menggunakan konsorsium dibandingkan dengan kultur tunggal dalam degradasi telah dibuktikan. Hal ini dapat dikaitkan dengan efek sinergis antara anggota konsorsium. Isolat Streptomyces exfoliatus 42 dilaporkan memiliki kemampuan menghasilkan selulase, xilanase, dan lignin peroksidase yang mampu mendegradasi lignoselulosa dari substrat tandan kosong kelapa sawit (TKKS), bagas tebu dan tongkol jagung (Apriani 2013; Astuti 2012). Menurut Nur (2008),
2
Kultur Aspergillus tamari yang ditambahkan pada fermentasi kulit kopi dapat meningkatkan perolehan polifenol (Gonjales et al. 2011). Kultur tersebut melepaskan salah satu komponen polifenol yaitu asam hidroksinamat yang terikat pada lignoselulosa kulit kopi yang dapat dimanfaatkan sebagai antioksidan. Polifenol merupakan golongan senyawa metabolit pada kulit kopi. Polifenol pada kulit kopi antara lain tanin, flavonol, plavan-3-ol, katekin, epikatekin, asam hidroksimat, asam ferulat dan senyawa aldehid seperti kafein (Ramirez et al.
2004). Senyawa antosianin pada kulit kopi juga dapat digunakan sebagai pewarna alami untuk makanan (Prata dan Olivera 2006). Senyawa-senyawa polifenol tersebut dapat diperoleh dengan merusak atau mendegradasi komponen lignoselulosa yang mengikatnya seperti lignin, protein, dan karbohidrat (selulosa, hemiselulosa). Gonzales et al. (2011) menyatakan bahwa senyawa polifenol terikat kuat secara kovalen pada dinding sel. Oleh karena itu, perolehan ekstrak polifenol akan lebih banyak jika komponen yang mengikat senyawa tersebut didegradasi terlebih dahulu.
Terdegradasinya komponen lignoselulosa kulit kopi maka diharapkan dapat meningkatkan perolehan ekstrak polifenol yang terikat pada komponen sel kulit kopi. Pendegradasian kulit kopi untuk meningkatkan perolehan ekstrak polifenol akan meningkatkan nilai guna dari kulit kopi dan meningkatkan nilai ekonomis dari limbah kulit kopi yang selama ini kurang termanfaatkan secara maksimal. Selain itu juga diharapkan akan memudahkan proses pengolahan kopi. Dalam penelitian ini digunakan konsorsium Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3 untuk memaksimalkan hasil degradasi kulit kopi. Dengan demikian pendekatan bioteknologi perlu dilakukan untuk meningkatkan perolehan polifenol limbah kulit kopi robusta dengan kultivasi media padat atau Solid State Fermentation (SSF).
Tujuan Penelitian
Penelitian ini bertujuan untuk meningkatkan perolehan ekstrak senyawa polifenol dengan mendegradasi komponen lignoselulosa kulit kopi menggunakan konsorsium isolat aktinomiset dan kultivasi media padat.
Manfaat Penelitian
3
Ruang Lingkup
Ruang lingkup dalam penelitian ini adalah sebagai berikut :
1. Pemanfaatan buah kopi yang digunakan dalam penelitian ini adalah jenis Robusta (Coffea canephora) segar yang baru dipanen dan berasal dari Lampung.
2. Peremajaan isolat dan produksi enzim konsorsium Streptomyces exfoliatus
42 dan Streptomyces costaricanus 45I-3 meliputi pengukuran aktivitas selulase dan xilanase.
3. Proses kultivasi buah kopi menggunakan konsorsium Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3 dilakukan selama 9 hari pada suhu 27 °C.
4. Penentuan kinerja hasil kultivasi menggunakan konsorsium Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3 meliputi kandungan serat (selulosa, hemiselulosa dan lignin), kandungan gula (total gula dan gula reduksi) dan senyawa bioaktif.
5. Senyawa bioaktif yang diamati yaitu tergolong polifenol yang meliputi total polifenol, tanin, katekin, dan antosianin.
TINJAUAN PUSTAKA
Kulit kopi
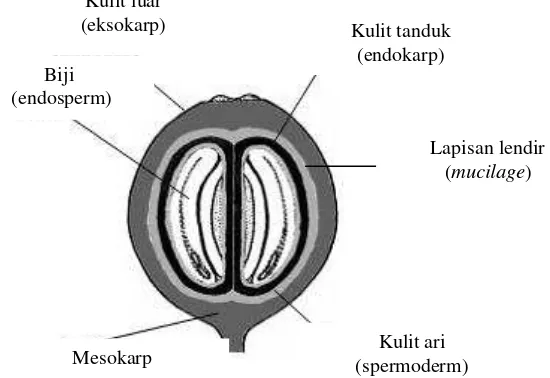
Kopi merupakan salah satu komoditas unggulan dalam sektor perkebunan di Indonesia. Indonesia memiliki dua jenis kopi utama yaitu jenis Arabika dan Robusta. Buah kopi tersebut diolah atau diproses menjadi produk olahan kopi sebelum diekspor ke negara lain. Hasil samping dari pengolahan kopi tersebut berupa limbah kulit buah kopi. Buah kopi memiliki beberapa bagian (Gambar 1).
Gambar 1 Struktur buah kopi (Avallone et al. 2002)
Kulit luar (eksokarp)
Biji (endosperm)
Kulit tanduk (endokarp)
Mesokarp (spermoderm) Kulit ari
Lapisan lendir
4
Kulit kopi atau kulit buah kopi merupakan bagian terluar dari buah kopi disebut juga eksokarp, sedangkan bagian kulit dengan daging kulit yang berasa manis dan mempunyai kandungan air yang cukup tinggi disebut dengan mesokarp. Endokarp atau kulit tanduk merupakan kulit kopi paling keras tersusun oleh selulosa dan hemiselulosa. Kulit ari merupakan kulit yang paling tipis dan menempel pada kulit kopi dan endosperm atau keping biji merupakan bagian buah kopi yang dimanfaatkan untuk diolah menjadi kopi bubuk. Pada bagian kulit luar, mesokarp, dan kulit tanduk pada kulit kopi mengandung karbohidrat (35%), protein (5.2%), serat (30.8%), dan mineral (10.7%) sedangkan di bagian mucilage
mengandung air (84.2%), protein (8.9%), gula (4.1%), dan abu (0.7%). Selain itu kulit kopi mengandung beberapa senyawa metabolit sekunder seperti kafein dan golongan polifenol. Menurut Ramirez et al. (2004) senyawa polifenol yang terkandung dalam limbah kulit kopi seperti tanin, flavonol, plavan-3-ol, asam hidroksimat dan aldehid seperti kafein.
Kulit kopi merupakan limbah lignoselulosa yang dapat digunakan sebagai salah satu bahan baku berbagai olahan seperti pembuatan etanol, asam karboksilat, sorbitol, xilitol, asam amino atau produk-produk lain yang lebih kompleks seperti protein sel tunggal. Lignoselulosa terdiri atas tiga penyusun utama, yaitu selulosa, hemiselulosa, dan lignin yang saling terikat satu sama lainnya. Kulit buah kopi atau pulp kopi diketahui memiliki kandungan 49% selulosa, 24.5% hemiselulosa, dan 7.63% lignin (Diniyah et al. 2013).
Biomassa Lignoselulosa
Bahan lignoselulosa merupakan komponen organik berlimpah di alam, yang terdiri dari tiga polimer yaitu selulosa, hemiselulosa dan lignin (Gambar 2). Lapisan S1 dan S3 memiliki selulosa amorf dan hemiselulosa. Lapisan S2 memiliki daerah lapisan selulosa kristal (Lee 2005). Dengan tingginya kandungan bahan organik tersebut maka lignoselulosa dapat dijadikan biomaterial untuk menghasilkan produk yang memiliki nilai yang lebih tinggi. Kulit kopi mempunyai komposisi 49% selulosa, 24.5% hemiselulosa, dan kandungan lignin 7.63%.
Gambar 2 Struktur dinding sel selulosa, hemiselulosa, dan lignin dari bahan lignoselulosa (Lee 2005)
Selulosa merupakan homopolisakarida yang mempunyai molekul berbentuk linear, tidak bercabang dan tersusun atas 10 000 sampai 15 000 unit glukosa yang dihubungkan dengan ikatan ß-1.4 glikosidik (Nelson dan Michael 2000). Bangun
Lignin
Hemiselulosa
Selulosa
S1
S2
5 dasar selulosa berupa suatu selubiosa yaitu dimer dari glukosa. Rantai panjang selulosa terhubung secara bersama melalui ikatan hidrogen dan Van der Waal (Perez et al. 2002). Ikatan β-1.4 glikosidik yang kuat dari selulosa dapat membentuk kristal mikrofibril yang kemudian secara bersama-sama membentuk serat selulosa yang tidak larut.
Enzim selulase pada umumnya diklasifikasikan menjadi tiga kelompok tergantung spesifisitas dalam menghidrolisis selulosa, yaitu endoglukanase (EC 3.2.1.4), eksoglukanase (EC 3.2.1.91), dan ß-glukosidase (EC 3.2.1.21). Ketiga kelompok enzim ini bekerjasama menghidrolisa selulosa yang tidak dapat larut menjadi glukosa, sehingga aktivitas gabungan ketiga enzim ini dapat diukur dengan memantau jumlah glukosa yang dihasilkan. Hidrolisis selulosa dapat dilakukan oleh beberapa agen biologis seperti cendawan ataupun bakteri dari genus Streptomyces. Semedo et al. (2004) melaporkan bahwa Streptomyces drozdowiczii M7aT hasil isolasi dari sampel tanah di Rio de Janeiro, Brazil, yang tergolong bakteri termofilik dan memiliki aktivitas selulolitik pada suhu 70-100 o
C.
Hemiselulosa adalah salah satu penyusun dinding sel tumbuhan yang terdiri dari kumpulan beberapa unit gula atau heteropolisakarida dan dikelompokkan berdasarkan residu gula utama sebagai penyusunnya seperti xilan, mannan, galaktan dan glukan (Fengel dan Wegener 1995). Komponen utama hemiselulosa adalah xilan yang memiliki ikatan rantai β-1.4 xilosida. Xilan merupakan komponen penyusun hemiselulosa terbesar yang merupakan polimer dari pentosa atau xilosa dengan ikatan ß-1.4 dan tersusun atas 150-200 monomer xilosa dimana unit struktur sering disubstitusi pada posisi C2 atau C3 dengan arabinofuranosil, asam 4-O-metil glukuronik, asetil atau fenolik (Moure et al. 2006). Xilan lebih cepat diurai oleh mikroba dibandingkan selulosa karena ikatannya yang cenderung lemah. Ikatan tersebut dapat diputus dengan bantuan enzim xilanase. Xilanase dapat dihasilkan dari bakteri, fungi maupun khamir antara lain Aspergillus niger, Penicillium chrysogenum, Streptomyces, Bacillus, Aureobasidium, Fusarium, Chaetomium, Phanerochaete, Rhizomucor, Humicola, Talaromyces, Cryptococcus dan Trichoderma viridae (Goswami dan Pathak 2013).
Lignin merupakan polimer aromatik yang berasosiasi dengan polisakarida pada dinding sel sekunder tanaman. Pada umumnya lignin mengandung tiga jenis alkohol aromatik yaitu koniferil, sinapil, dan p-koumaril (Howard et al. 2003). Di dalam jaringan tanaman, lignin sulit didegradasi karena mempunyai struktur kompleks dan heterogen yang berikatan dengan selulosa dan hemiselulosa. Bakteri mampu melakukan delignifikasi pada berbagai bahan lignoselulosa dengan mensekresikan enzim peroksidase. Menurut Bugg et al. (2011) bahwa enzim mengenai bakteri pendegradasi lignin belum banyak diketahui dibandingkan dengan enzim pendegradasi lignin yang dihasilkan oleh cendawan. Mikroorganisme dari golongan cendawan yang mampu mendegradasi lignin yaitu
6
Streptomyces viridosporus T7A dengan Solid State fermentation melakukan depolimerisasi lignin dalam mendegradasi selulosa dan memproduksi APPL (acid-precipitable polymeric lignin) yang larut dalam air sebagai produk utama degradasi lignin (Ramachandra et al. 1988).
Aktinomiset
Berdasarkan analisis gen 16S rRNA, aktinomiset diklasifikasikan dalam domain Bacteria, filum Actinobacteria, kelas Schizomycetes, dan ordo Actinomycetales (Hayakawa 2003). Aktinomiset adalah mikrob uniseluler yang dikelompokkan ke dalam bakteri Gram positif dengan DNA yang kaya kandungan G dan C, yaitu sekitar 57-75% (Lo et al. 2002). Aktinomiset bereproduksi dengan spora aerial (konidia) atau melalui fragmentasi miselia. Aktinomiset memiliki dua macam miselia, yaitu miselia aerial dan miselia substrat, kedua miselia ini mampu menghasilkan pigmen yang menyebabkan perbedaan warna pada masing-masing koloni.
Streptomyces sp. dapat tumbuh pada berbagai sumber karbon dan memiliki kemampuan untuk mendegradasi selulosa, hemiselulosa, dan lignin yang banyak terdapat pada tanaman. El-Sersy et al. (2010) melaporkan bahwa Streptomyces ruber mampu menggunakan selulosa jerami padi untuk meproduksi enzim selulase. Menurut Yassien et al. (2013) bahwa Streptomyces sp.hasil isolasi dari tanah di Arab Saudi teridentifikasi sebagai Streptomyces longispororuber. Streptomyces sp. SKK1-8 mampu menghasilkan xilanase yang dapat memecah substrat p-NP-ß-D-xilanopiranosida, p-NP-a-L-arabinofuranosida, p NP-a-D-glukopiranosida dan p-NP-a-D-galaktopiranosida, akan tetapi tidak memecah substrat p-NP-asetat (Meryandini et al. 2008). Streptomyces sp. 7b dilaporkan oleh Bajaj et al. (2010) mampu memproduksi xilanase pada dedak gandum melalui kultivasi media padat (Solid State Fermentation).
Senyawa Bioaktif pada Kulit Kopi
Senyawa bioaktif adalah senyawa yang dihasilkan tanaman dan memiliki efek farmakologis atau toksikologi pada manusia dan hewan (Bernhoft 2010). Senyawa bioaktif dihasilkan sebagai metabolit sekunder. Kulit buah kopi memiliki beberapa senyawa bioaktif seperti senyawa polifenol dan alkaloid. Polifenol merupakan kelompok zat kimia yang ditemukan pada tumbuhan. Zat ini memiliki tanda khas yaitu memiliki banyak gugus fenol dalam molekulnya. Komponen fenolik merupakan komponen yang terdiri atas cincin aromatik dengan satu atau lebih gugus hidroksil yang terdapat pada tanaman dan memiliki kemampuan sebagai antioksidan, antara lain seperti: asam fenolik, tannin, dan flavonoid (Balasundram et al. 2006).
7 tergolong tanin terkondensasi, banyak terdapat pada buah-buahan, biji-bijian dan tanaman pangan, sementara yang tergolong tanin terhidrolisis terdapat pada bahan non pangan (Makkar 1993).
Antosianin adalah komponen flavonoid yang termasuk kelompok polifenol banyak terdapat pada tumbuhan seperti buah-buahan, sayuran, dan bunga. Antosianin memiliki lima subkelas, yaitu peralgonidin, cyanidin, peonidin, malvidin, dan delphinidin (Ajila et al. 2011). Di antara semua polifenol, antosianin dikenal sangat penting karena berpotensi sebagai antioksidan dan paling banyak dikonsumsi (Prior 2003) dan Mayer-meibach et al. (2012). Menurut Jurgonski et al. (2013) antosianin berpotensi mengatasi penyakit diabetes dan kardiovaskuler.
METODE PENELITIAN
Waktu dan Tempat
Penelitian ini dilaksanakan pada Februari 2014 sampai dengan Desember 2014 di Laboratorium Bioteknologi Hewan dan Biomedis PPSHB-LPPM dan Kimia Analitik Institut Pertanian Bogor.
Alat dan Bahan
Bahan-bahan yang digunakan dalam penelitian ini meliputi buah kopi, isolat
Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3. Bahan kimia yang digunakan meliputi media Yeast Starch Agar (YSA), media Carboxymetyl cellulose (CMC), media xylan beechwood, Dinitrosalicylic acid (DNS), etanol, butanol-HCL, H2SO4, Fenol, Neutral Detergent Solvent (NDS), Acid Detergent
Solvent (ADS).
Alat yang digunakan antara lain: Laminar Air Flow, spektrofotometer UV-Vis, High Performance Liquid Chromatografi (HPLC), neraca analitik, Hammer mill, cockborer, autoklaf, oven, waterbath shaker, mikropipet, tanur, desikator, tabung kjehdahl, cawan porselin, pembakar gas, gegep besi, sentrifugasi rotor
fixed angle, dan alat-alat gelas.
Karakterisasi Bahan Baku
8
Peremajaan Isolat dan Produksi Enzim
Isolat Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3 merupakan koleksi Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi Institut Pertanian Bogor (PPSHB IPB). Isolat tersebut diremajakan pada media YSA, dan diinkubasi selama 7 hari pada suhu ruang. Hasil peremajaan isolat digunakan untuk propagasi kultur. Untuk 100 mL media propagasi terdiri atas 80 mL media YSA yang disuplementasi 1 g serbuk kulit buah kopi kering, selanjutnya kultur diinkubasi selama 7 hari pada suhu 27 °C.
Produksi enzim dilakukan dengan menentuan aktivitas enzim selulase serta xilanase untuk Streptomyces exfoliatus 42 dan aktivitas enzim xilanase untuk
Streptomyces costaricanus 45I-3. Pengukuran aktivitas enzim selulase dilakukan berdasarkan pembentukan gula pereduksi hasil hidrolisis CMC menggunakan metode Dinitrosalicylic acid atau DNS (Miller 1959) (Lampiran 5), dengan pengamatan setiap hari selama 8 hari.
Pengukuran aktivitas enzim xilanase dilakukan berdasarkan pembentukan gula pereduksi menggunakan metode DNS (Miller 1959) (Lampiran 6), menggunakan media beechwood xylan dan dilakukan pengamatan setiap hari selama 8 hari. Untuk pertumbuhan sel masing-masing bakteri dihitung berdasarkan biomassa berat kering yang menggunakan kertas saring kemudian dikeringkan ke dalam oven selama 24 jam pada suhu 80 °C (Yee et al. 1996) (Lampiran 7).
Degradasi Komponen Serat Pada Biomassa Kulit Kopi Persiapan Substrat dan Kultur Starter
Sebanyak 2 cockborer isolat Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3, masing-masing diinokulasikan ke dalam 100 mL media cair yang disuplementasi 1 g kulit buah kopi kering. Media yang telah berisi kultur selanjutnya diinkubasi selama 10 hari pada suhu 27 °C. Pengamatan dilakukan setiap 24 jam untuk mengetahui pertumbuhan bakteri. Selanjutnya untuk kultivasi, digunakan 500 g buah kopi Robusta (Coffea canephora) segaryang baru dipanen. Buah kopi dicuci dengan air mengalir kemudian dimasukkan ke dalam wadah fermentasi dan selanjutnya disterilkan menggunakan sinar UV selama 60 menit.
Proses Kultivasi
9
Kinerja Hasil Kultivasi
Keberhasilan konsorsium aktinomiset dalam mendegradasi kulit kopi ditandai dengan adanya penurunan susut bobot, perubahan komponen serat, perolehan ekstrak gula dan senyawa metabolit sekunder. Buah kopi setelah dikultivasi selanjutnya dikupas untuk memisahkan kulit kopi dengan biji kopi. Kulit kopi selanjutnya dikeringkan di oven pada suhu 50 °C selama 48 jam. Selanjutnya kulit kopi digunakan untuk analisis yang meliputi kandungan selulosa, hemiselulosa, dan lignin serta zat ekstrakif menggunakan metode Van Soest et al.
(1963) (Lampiran 2).
Hasil degradasi komponen lignoselulosa kulit kopi juga berupa monomer gula dan senyawa bioaktifnya. Untuk analisis komponen tersebut, kulit kopi yang sudah kering digiling hingga diperoleh ukuran 40 mesh. Proses ekstraksi dilakukan secara maserasi menggunakan etanol : air (80 : 20) selama 24 jam. Analisis gula meliputi gula total dan gula reduksi. Total gula diukur dengan metode fenol-H2SO4 (Dubois et al. 1956) (Lampiran 3), gula pereduksi dengan metode DNS (Miller 1959) (Lampiran 3). Analisis senyawa bioaktif dari kulit kopi meliputi total polifenol, tanin, antosianin, dan katekin. Total polifenol dianalisis dengan metode Folin-Ciocalteu oleh Singleton dan Rossi (1965) menggunakan spektrofotometer UV-Vis pada panjang gelombang 700 nm, tanin dengan metode Butanol-HCl oleh IAEA (1999), katekin dianalisis dengan menggunakan HPLC menurut Guyot et al. (2001), dan antosianin diukur berdasarkan metode Iglesias et al. (2008) (Lampiran 4).
Analisis Data
Data yang diperoleh dianalisis menggunakan analisis statistik. Parameter yang diamati yaitu komponen serat (selulosa, hemiselulosa, dan lignin), kandungan gula (total gula dan gula reduksi), dan senyawa bioaktif (total polifenol, tanin, antosianin, dan katekin). Data yang diperoleh ditampilkan dalam bentuk rataan ± standar deviasi dari 2 ulangan, selanjutnya dianalisis menggunakan Rancangan Acak Lengkap (RAL) menggunakan software
10
HASIL DAN PEMBAHASAN
Karakteristik Kulit Buah Kopi
Penggunaan bakteri untuk ekstraksi polifenol dari kulit kopi memerlukan persyaratan substrat yang memungkinkan bakteri untuk hidup. Bakteri memerlukan protein sebagai salah satu nitrogen organik dalam pertumbuhannya (Rachman 1999). Hasil analisis kulit kopi (Tabel 1) memberikan informasi bahwa kulit kopi merupakan suatu bahan yang menyediakan sumber nitrogen dan sumber karbon untuk pertumbuhan mikrob. Hal ini karena kulit kopi merupakan komponen kompleks yang terdiri atas serat kasar, protein, lemak, karbohidrat, gula total, dan gula reduksi (Tabel 1). Hasil yang sama dikemukakan oleh Brand
et al. (2001) yang menyatakan bahwa kulit buah kopi yang terdiri dari kulit luar, pulp, dan perkamen, mengandung (% DM): karbohidrat (21-35), protein (5.2), serat kasar (30.8), lemak (2.0-7.0), dan mineral (10.7). Dengan demikian maka kulit kopi dapat digunakan sebagai substrat tunggal untuk pertumbuhan konsorsium aktinomiset tanpa adanya penambahan komponen lain sebagai nutrisi pada proses kultivasi. Substrat dengan kandungan karbohidrat dan protein yang tinggi berpotensi untuk dimanfaatkan dalam bioproses (Ashock et al. 2000). Tabel 1 Komponen kimia kulit buah kopi
Kandungan bahan Komposisi (% bk) Kandungan bahan Komposisi (% bk)
Bahan kering 37.34 Komponen serat
Kadar abu 3.43 Lignin 6.10
Serat kasar 13.39 Selulosa 53.42
Protein 3.90 Hemiselulosa 24.10
Lemak 1.51 Senyawa bioaktif
Karbohidrat (by
difference) 28.50 Polifenol 0.73 mg mL
-1
Kandungan gula Tanin 2.0 %
Gula total 11.8 mg mL-1
Gula reduksi 2.0 mg mL-1
11
Produksi Enzim
Aktivitas enzim yang dihasilkan serta pertumbuhan sel Streptomyces sp. diamati selama 8 hari. Aktivitas enzim xilanase Streptomyces exfoliatus 42 pada media xilan 0.5% tertinggi pada hari ke-4 sebesar 205.5 mU mL-1 dan biomassa sel sebesar 1.15 g L-1, sedangkan Streptomyces costaricanus 45I-3 pada hari ke-4 sebesar 248.31 mU mL-1 dan biomassa sel sebesar 1.25 g L-1. Aktivitas enzim selulase Streptomyces exfoliatus 42 pada media CMC 1% tertinggi pada hari ke-4 sebesar 25.5 mU mL-1 dan biomassa sel sebesar 1.15 g L-1.
Gambar 3 Aktivitas enzim selulase pada media CMC 1% ( ), xilanase pada media beechwood xylan 0.5% ( ) dan pertumbuhan
Streptomyces exfoliatus 42 ( ) yang diinkubasi pada suhu 27 °C
Gambar 4 Aktivitas xilanase pada media beechwood xylan 0.5% ( ) dan pertumbuhan Streptomyses costaricanus 45I-3 ( ) diinkubasi pada suhu 27 °C
12
Streptomyces exfoliatus 42 pada inkubasi hari ke-1 diketahui memasuki fase lag, dimana sel mikrob yang dipindahkan ke media cair xilan mengalami adaptasi terlebih dahulu. Fase tersebut ditandai dengan pertambahan biomassa sel
Streptomyces exfoliatus 42 yang tidak terlalu besar. Hari ke-3 sampai dengan hari ke-4 merupakan fase pertumbuhan atau fase eksponensial yang terlihat dari kenaikan biomassa sel dan aktivitas xilanase yang mencapai optimum. Hari ke-5 hingga hari ke-7 merupakan pertumbuhan sel memasuki fase stasioner yang ditandai dengan bobot biomassa sel dan aktivitas enzim yang cenderung stabil. Xilanase mulai diproduksi pada saat sel mengalami fase eksponensial, substrat mulai dimanfaatkan oleh mikrob untuk memproduksi xilanase. Peningkatan aktivitas enzim xilanase ini berhubungan dengan pola pertumbuhan sel, laju pertumbuhan sel akan maksimum ketika waktu fase eksponensial telah tercapai dan aktivitas enzim akan berkurang atau stabil selama fase stasioner (Amraini 2008). Streptomyces exfoliatus 42 pada media cair xilan (Gambar 3) juga memperlihatkan fase-fase pertumbuhan yang dialami isolat tersebut. Pola peningkatan populasi selama rentang waktu tertentu akan membentuk fase-fase pertumbuhan. Fase-fase tersebut meliputi fase lag, fase eksponensial, fase stasioner dan fase kematian (Sunatmo 2009).
Enzim selulase juga dihasilkan oleh Streptomyces exfoliatus 42. Aktivitas selulase yang dihasilkan oleh Streptomyces exfoliatus 42 meningkat seiring dengan pertumbuhan biomassa selnya. Aktivitas selulase mulai mengalami peningkatan pada inkubasi hari ke-1. Hal ini menunjukkan bahwa selulase dihasilkan untuk memanfaatkan CMC sebagai sumber karbon. Tamburini et al.
(2004) dalam penelitiannya mengemukakan bahwa aktivitas selulase Streptomyces rochei A2 dalam mendegradasi CMC berlangsung selama fase awal pertumbuhan yang cepat, dan aktivitasnya sebanding dengan kenaikan bahan sel. Menurut Datshban et al. (2010) hidrolisis selulosa membutuhkan gabungan enzim endoglukanase, eksoglukanase dan ß-glukosidase yang bekerja secara sinergis. Mikrob yang mampu tumbuh pada substrat CMC memiliki endoglukanase yang dapat memutus ikatan ß-1.4 glikosida (Wang et al. 2008; Maki et al. 2009). Enzim tersebut akan mendegradasi substrat CMC dengan memutuskan ikatan selulosa pada bagian internal area amorf. Selanjutnya ketika sel mencapai fase stasioner, aktivitas enzim selulase juga memperlihatkan pola yang statis. Fase stasioner ditandai dengan kecepatan pembelahan sel sama dengan kecepatan kematian sel dan lisis sel (Martina et al. 2002). Hal ini menyebabkan aktivitas enzim juga statis bahkan sedikit cenderung turun.
13 ke-5 hingga hari ke-8 merupakan pertumbuhan sel memasuki fase stasioner yang ditandai dengan bobot biomassa dan aktivitas xilanase yang cenderung stabil.
Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3 merupakan bakteri Gram positif yang memiliki dua macam miselia, yaitu miselia aerial dan miselia substrat. Aktinomiset tersebut mampu menghasilkan senyawa enzim hidrolitik berupa selulase dan xilanase yang dapat bermanfaat pada bioproses. Pada ekstraksi dengan dihasilkannya enzim tersebut akan membantu melepaskan senyawa metabolit dari tanaman. Menurut Vattem dan Setti (2003) perolehan polifenol dapat ditingkatkan dengan bantuan enzim β-glukosidase yang berasal dari Aspergillus lentinusedodes melalui SSF. Salah satu karakteristik SSF yaitu rendahnya agitasi dan bakteri sebagai pelaku proses fermentasi. Agitasi diketahui memiliki efek buruk yang dapat menyebabkan gangguan keterikatan bakteri dengan substrat, dan kerusakan miselia (Lonsane et al. 1992). Dengan karakteristik Streptomyces sp. yang memiliki miselia atau berfilamen tersebut maka mendukung untuk dilakukan proses fermentasi secara SSF.
Degradasi Komponen Serat pada Biomassa Kulit Kopi
Aktivitas Enzim Oleh Konsorsium Isolat Aktinomiset pada Media Kulit Kopi
Aktivitas enzim selulase dan xilanase oleh konsorsium aktinomiset pada media kulit kopi diukur untuk menentukan waktu yang diperlukan untuk persiapan mikrob konsorsium aktinomiset sebagai starter yang digunakan dalam proses kultivasi. Aktivitas enzim selulase mengalami peningkatan tertinggi pada hari ke-4 sebesar 21.61 mU mL-1, dan aktivitas xilanase pada hari ke-6 sebesar 98.11 mU mL-1 (Gambar 5). Hasil yang berbeda diperlihatkan oleh penelitian Vats
14
Aktivitas selulase dan xilanase oleh konsorsium aktinomiset pada kulit kopi cenderung lambat dibanding pada aktivitas enzim xilanase dan selulase kultur tunggal (Gambar 4). Aktivitas xilanase dan selulase pada awal inkubasi terlihat belum mengalami peningkatan yang cukup besar. Hal ini dikarenakan hari ke-1 dan ke-2 inkubasi merupakan fase adaptasi atau fase lag dari mikrob konsorsium Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3. Mikrob yang telah dikultur dan selanjutnya dipindahkan ke lingkungan yang baru maka mikrob tersebut akan menyesuaikan diri terlebih dahulu (Stanbury dan Whitaker 1993). Peningkatan aktivitas enzim yang cenderung lambat juga dikarenakan konsorsium aktinomiset menggunakan komponen yang paling sederhana yaitu gula pereduksi berupa glukosa yang terdapat pada kulit kopi tanpa harus menghidrolisis selulosa maupun xilan untuk digunakan sebagai sumber karbon. Fontes et al. (2000) melaporkan bahwa pertumbuhan mikrob penghasil xilanase yang diuji menggunakan glukosa dan xilan sebagai sumber karbon menghasilkan pertumbuhan sel lebih cepat pada medium glukosa dibandingkan dengan pertumbuhan sel pada media xilan. Pada substrat kulit kopi hari ke-3 sampai dengan hari ke-4 merupakan fase eksponensial yang terlihat dari puncak aktivitas enzim xilanase dan selulase yang dihasilkan dan kenaikan aktivitas enzimnya yang sangat besar. Puncak aktivitas enzim tertinggi pada hari ke-6. Hal tersebut mengindikasikan bahwa pertumbuhan konsorsium aktinomiset mencapai titik maksimum atau fase eksponensial. Hasil penelitian juga menunjukkan bahwa konsorsium aktinomiset menghasilkan aktivitas xilanolitik yang lebih tinggi dibandingkan dengan selulolitik meskipun selulosa merupakan komponen utama pulp kopi. Hasil penelitian tersebut sama dengan penelitian yang dikemukakan Tuncer et al. (2004) bahwa xilanase dan selulase oleh Streptomyces sp. FP2621 dihasilkan pada fase pertumbuhan (eksponensial).
Aktivitas enzim oleh konsorsium aktinomiset juga relatif rendah dibandingkan dengan aktivitas enzim tunggal pada media CMC (Gambar 3) dan xilan sintesis (Gambar 3 dan 4). Hal ini terjadi karena adanya persaingan nutrisi dalam penggunaan substrat yang dibutuhkan mikrob dalam pertumbuhan. Interaksi antar spesies tidak hanya bersifat sinergisme atau komensalisme, tetapi dapat juga bersifat kompetisi dan penghambatan (Kato et al. 2005). Perbedaan yang lain juga disebabkan oleh adanya polifenol yang mampu menjadi inhibitor bagi enzim. Polifenol mampu berikatan pada sisi aktif enzim sehingga menghambat aktivitas enzim selulase (McDaugall et al. 2005 dan Jurgonski et al. 2013). Polifenol tersebut merupakan polifenol larut air yang muncul karena ukuran substrat yang dikecilkan menjadi 40 mesh. Selain itu juga perbedaan aktivitas enzim terjadi karena adanya perbedaan dan konsentrasi substrat. Enzim memiliki spesifitas yang tinggi terhadap substrat. Penggunaan media yang berbeda menyebabkan perbedaan aktivitas enzim yang dihasilkan (White 1995).
Kinerja Hasil Degradasi
15 substrat. Interaksi antara substrat dengan bakteri menyebabkan dihasilkannya aktivitas enzim endo glukanase, β glucanase, dan xilanase oleh konsorsium
Clostridium dan Thermoanaerobacterium yang mengakibatkan penurunan bobot substrat jerami padi, bagas, dan tongkol jagung setelah difermentasi selama 7 hari (Wangwilaiwalin et al. 2010). Persentase susut bobot (%) pada biomassa kulit kopi (Gambar 6) selama 9 hari lama inkubasi masing-masing sebesar 19.02% pada hari ke-3, 34.24% pada hari ke-6, dan 39.29% pada hari ke-9.
Gambar 6 Persentase susut bobot substrat kopi setelah kultivasi metode padat pada suhu 27 °C
Kemampuan konsorsium aktinomiset dalam berinteraksi dengan substrat diperlihatkan dengan pertumbuhan koloni Streptomyces sp. pada permukaan substrat kulit buah kopi dengan warna koloni abu-abu dan putih, sedangkan pada perlakuan kontrol terdapat sedikit koloni yang berwarna putih. Pertumbuhan aktinomiset juga terlihat dari banyaknya uap air yang terdapat pada wadah kultivasi. Menurut Dalzel et al. (1997) dalam perombakan bahan organik oleh mikroba dibutuhkan air dan oksigen dan nutrien dari bahan organik sebagai sumber energi dan kemudian melepaskan CO2, air dan energi panas sehingga menyebabkan bobot bahan semakin berkurang. Terjadinya penurunan bobot juga diakibatkan oleh pelepasan CO2 dan senyawa-senyawa lainnya. Menurut Tuomela
et al. (2000) bahan organik dari lignoselulosa dimanfaatkan mikrob sebagai sumber karbon untuk proses metabolisme yang menghasilkan energi dan melepaskan CO2 dan produk akhir yang lebih sederhana sesuai dengan enzim ekstraseluler yang dihasilkan. Penyusutan bobot juga menunjukkan bahwa konsorsium aktinomiset yang digunakan mampu merombak komponen serat yang meliputi selulosa, hemiselulosa, dan lignin pada kulit kopi.
Perubahan kandungan komponen substrat dikaitkan dengan adanya mikroorganisme dan kemampuannya dalam mensekresikan enzim ekstraseluler. Pada perlakuan kontrol (fermentasi spontan atau tanpa penambahan kultur konsorsium Streptomyces sp.) terjadi penurunan komposisi komponen serat tidak signifikan dibandingkan dengan perlakuan sampel (Tabel 2). Hal ini terjadi karena hanya terdapat bakteri indigenous pada fermentasi buah kopi tersebut. Menurut Silva et al. (2008), pada fermentasi spontan proses pengolahan kopi secara alami
16
terdapat mikroorganisme seperti Bacillus sp. Bakteri tersebut mampu menghidrolisis selulosa karena adanya kemampuan menghasilkan selulase (Coughlan dan Mayer 1991). Pada perlakuan konsorsium aktinomiset, penurunan komposisi serat berkaitan dengan kemampuan mikrob dalam mensekresikan beberapa enzim ekstraseluler untuk pertumbuhan, yaitu selulase dan xilanase (Gambar 3) serta lignin peroksidase (Apriyani 2012). Kemampuan mikrob dalam mengasimilasi bahan organik tergantung dengan kemampuannya dalam memproduksi enzim yang dibutuhkan untuk mendegradasi komponen substrat, semakin kompleks substrat yang digunakan maka semakin komplek enzim yang dibutuhkan (Tuomela et al. 2000). Menurunnya kandungan serat selulosa menjadi 29% dan hemiselulosa menjadi 5.4% pada Borage officinalis disebabkan karena penggunaan enzim komersial yaitu selulase dan xilanase secara konsorsium (Soto
et al. 2008). Keberadaan enzim-enzim yang dihasilkan oleh konsorsium aktinomiset efektif digunakan dalam mendegradasi komponen organik yang kompleks menjadi molekul sederhana. Molekul-molekul sederhana yaitu berupa monosakarida yang dapat digunakan sebagai sumber karbon bagi sel mikrob. Faktor lain yang ikut mempengaruhi degradasi komponen serat yaitu aktivitas enzim yang dimiliki. Semakin besar aktivitas enzim maka akan semakin tinggi pula kemampuannya dalam mendegradasi komponen lignoselulosa. Kammerer et al. (2005) menyatakan bahwa penggunaan enzim pada ekstraksi polifenol dari limbah pengolahan anggur harus memiliki proses yang relevan seperti jenis enzim, rasio enzim-substrat dan suhu. B 23.31±0.27a 22.92±0.7b 15.68±0.7c 10.28±0.4d
Lignin A 6.10±0.15
Ket: A: Sampel (dengan kultur konsorsium Streptomyces sp.) B: Kontrol (tanpa kultur konsorsium Streptomyces sp.)
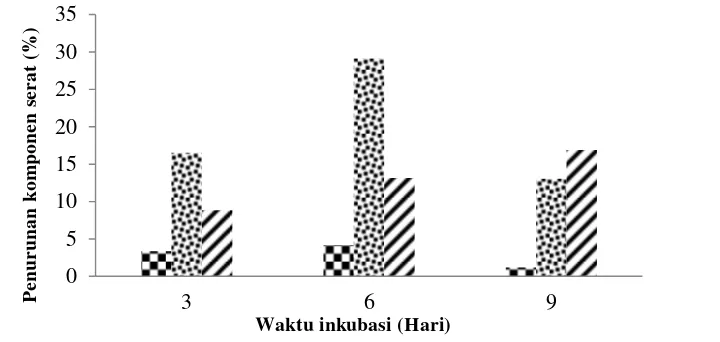
17 mampu mendegradasi lignin pada tongkol jagung sehingga menyebabkan penurunan lignin mencapai 4.8%. Enzim peroksidase seringkali dihubungkan dengan degradasi lignin karena kerja enzim relatif tidak spesifik. Substrat yang mampu didegradasi adalah senyawa seperti fenol, aromatik amines dan beberapa komponen seperti alkil peroksida (Jing Li et al. 2009). Penurunan kandungan lignin di kulit kopi terjadi pada inkubasi hari ke-3, dan penurunan terbesarnya pada hari ke-6. Penurunan tersebut seiring dengan meningkatnya aktivitas enzim peroksidase. Tuncer et al. (2004) menyatakan juga bahwa Streptomyces sp. FP2621 memiliki aktivitas lignoselulolitik berupa peroksidase. Aktivitas peroksidase meningkat seiring pertumbuhan biomassa sel dan mencapai aktivitas optimum setelah 4 hari inkubasi.
Gambar 7 Penurunan lignin ( ), selulosa ( ), dan hemiselulosa ( ) setelah diinkubasi pada suhu 27 °C
.
Kandungan selulosa pada kulit kopi (Gambar 7) mengalami penurunan tertinggi pada hari ke-6 yang berkaitkan dengan enzim selulase oleh isolat konsorsium aktinomiset Streptomyces exfoliatus 42. Hasil yang berbeda dilaporkan bahwa penurunan kandungan selulosa tongkol jagung mencapai 51% oleh konsorsium bakteri hasil isolasi dari tanah setelah mengalami inkubasi selama 8 hari (Feng et al. 2008). Menurut Jager et al. (2010) bahwa kristalinitas selulosa merupakan faktor utama yang menentukan penguraian selulosa oleh enzim selulolitik sehingga mikrob mampu menghidrolisis kristal selulosa yang terdapat di alam. Menurut Yoon et al. (2005) penurunan kandungan serat pangan tak larut berupa selulosa dilakukan oleh enzim selulase yang mendegradasi dinding sel sebagai komponen tidak larut menjadi komponen sederhana yang lebih larut. Wang et al. (2008) mengemukakan bahwa selulase merupakan enzim yang dihasilkan untuk memanfaatkan selulosa yang melibatkan gabungan enzim hidrolisis, terdiri atas endoglukanase, eksoglukanase (cellobiohydrolase), dan ß-glukosidase yang bekerja secara sinergis.
Penurunan kandungan hemiselulosa tertinggi pada hari ke-9 inkubasi. Penurunan tersebut lebih besar dibandingkan dengan penurunan selulosa. Hal tersebut berkaitan besarnya aktivitas xilanase yang dihasilkan oleh kedua isolat konsorsium. Xilanase yang dihasilkan menghidrolisis xilan sebagai komponen
18
terbesar penyusun hemiselulosa. Selama proses hidrolisis, enzim xilanase menghidrolisis xilan menjadi fraksi-fraksi penyusunnya. Hidrolisis xilan diperlukan beberapa enzim berbeda yaitu endo1.4-ß-xilanase yang menghidrolisis struktur dasar xilan secara acak menjadi xilooligosakarida, 1.4 ß-D-xilosidase yang memutus xilooligosakarida menjadi xilosa. Gugus penyusun samping xilan akan dibebaskan oleh α-L-arabinofuranosidase, α-D-glukorodase, dan asetil xilan esterase menjadi arabinosa, glukuronat, dan asetat (Subraminayan dan Prema 2002). Mekanisme hidrolisis xilan diawali oleh α-rabinofurabinosidase yang mampu menghidrolisis xilan menjadi L-arabinosa dan xilobiosa. Pemutusan rantai
cabang akan memudahkan hidrolisis xilan oleh eksoxilan dan β-xilosidase. Produk utama dari hidrolisis ketiga enzim tersebut adalah xilosa, arabinosa, dan xilotetrosa yang sedikit bercampur dengan xilotriosa (Puspaningsih 2004).
Terjadinya perubahan kandungan serat lignoselulosa pada kulit buah kopi yaitu selulosa dan hemiselulosa, berpengaruh juga terhadap perolehan komponen gula dan senyawa bioaktif. Penentuan komponen tersebut dilakukan dengan cara mengekstraksi menggunakan pelarut polar yaitu etanol. Oleh karena itu, dalam penelitian ini komponen yang terukur dari hasil ekstraksi merupakan komponen bioaktif yang bersifat polar. Polifenol merupakan kelompok senyawa polar yang banyak mengandung gugus OH (Shi et al. 2003). Pengukuran komponen gula yaitu total gula dan gula reduksi, sedangkan senyawa bioaktif meliputi total polifenol, antosianin, tanin dan katekin. Perolehan hasil ekstraksi setelah proses kultivasi menggunakan konsorsium aktinomiset dapat dilihat pada Tabel 3.
Tabel 3 Kandungan gula hasil ekstraksi setelah kultivasi pada suhu 27 °C
Komponen Perlakuan Waktu inkubasi (hari)
0 3 6 9
Ket: A: Sampel (dengan kultur konsorsium Streptomyces sp.) B: Kontrol (tanpa kultur konsorsium Streptomyces sp.)
19 Pada perlakuan kontrol, gula reduksi hanya sedikit saja mengalami peningkatan atau cenderung konstan, yang berbeda dengan perlakuan sampel. Dalam hal ini memungkinkan karena keterbatasan hadirnya mikroba yang hanya sedikit saja peningkatan gula reduksi oleh aktivitas mikrob indigeneus. Gula reduksi berupa monomer gula merupakan produk hasil degradasi serat selulosa dan xilan oleh aktivitas enzim xilanase dan selulase oleh konsorsium aktinomiset. Menurut Saha (2004) hidrolisis selulosa akan menghasilkan monomer glukosa dan oligomer selobiosa, sedangkan xilan berupa xilosa, arabinosa, dan xilo-oligosakarida. Menurut Yoon et al. (2005) hidrolisis serat menggunakan enzim selulase diperoleh beberapa komponen gula yaitu monosakarida (glukosa, fruktosa, galaktosa, dan arabinosa), selooligosakarida (selopentosa, selotetrosa, selotriosa, dan selobiosa), dan galaktooligosakarida (galaktotetrosa dan galaktotriosa).
Nilai gula reduksi meningkat seiring dengan peningkatan waktu inkubasi substrat oleh konsorsium aktinomiset. Perolehan gula reduksi meningkat setelah 3 hari inkubasi dan perolehan tertinggi terjadi pada inkubasi hari ke-6. Perihal tersebut terkait dengan hasil degradasi tertinggi komponen serat kompleks (Gambar 7). Pada hari ke-3 substrat belum terhidrolisis secara sempurna oleh xilanase dan selulase sehingga masih ada peningkatan gula reduksi pada hari ke-6. Molekul xilosa memiliki struktur dengan gugus karbonil yang berada pada ujung rantai karbon, yang menandakan bahwa xilosa mempunyai gugus aldehid bebas yang reaktif sehingga dimasukkan dalam kategori gula pereduksi (Yang et al.
2005).
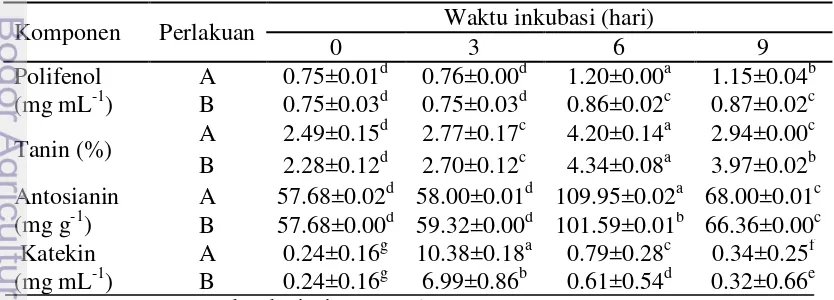
Terdegradasinya komponen selulosa, hemiselulosa dan lignin sebagai pengikat senyawa metabolit sekunder, maka berpengaruh juga terhadap perolehan hasil ekstraksi senyawa bioaktif pada kulit kopi (Tabel 4). Proses ekstraksi yang telah dilakukan diketahui perolehan total polifenol mengalami peningkatan yang lebih besar pada perlakuan sampel dibandingkan dengan kontrol. Hasil analisis
menggunakan α 0.05 juga menunjukkan bahwa penggunaan konsorsium
aktinomiset berpengaruh nyata terhadap perolehan hasil ekstrak senyawa bioaktif yaitu total polifenol dan uji lanjut Duncan juga menunjukkan adanya perbedaan yang nyata antara perlakuan kontrol dengan sampel.
Tabel 4 Komposisi perolehan hasil ekstraksi senyawa bioaktif kulit kopi setelah kultivasi pada suhu 27 °C
Komponen Perlakuan Waktu inkubasi (hari)
0 3 6 9
20
Pada perlakuan kontrol, keterbatasan mikroorganisme yang ada selama fermentasi menyebabkan perolehan ekstrak polifenol meningkat sedikit saja. Mikroorganisme hidrolitik yang hadir dalam fermentasi spontan hanya Bacillus sp. (Coughlan dan Mayer 1998). Pada fermentasi adanya enzim hidrolitik menyebabkan degradasi dinding sel tetapi juga dapat mempengaruhi stabilitas senyawa fenolik. Hidrolisis menggunakan enzim mampu menurunkan viskositas substrat, mengurangi gaya tarik menarik antara molekul-molekul dan menurunkan stabilitas interaksi antara asam uronik, protein, dan tanin. Selain itu pemecahan polimer dinding sel meningkatkan permeabilitas dan porositas sel, serta kelarutan yang lebih besar pada komponen internal sel, sehingga akan berakibat peningkatan konsentrasi senyawa fenolik dan aktivitas antioksidan dari ekstrak (Cerda et al. 2013). Oleh karena itu, penambahan kultur konsorsium aktinomiset mampu meningkatkan perolehan senyawa bioaktif terutama ekstrak senyawa polifenol. Analisis perolehan senyawa metabolit sekunder meliputi senyawa bioaktif kulit kopi dilakukan terhadap 4 macam, yaitu total polifenol, tanin, antosianin, dan katekin. Menurut Ramirez-Coronel et al. (2004) menemukan senyawa bioaktif pada kulit kopi terdiri atas empat kelas utama polifenol dalam kulit buah kopi yaitu Flavan-3-ols, asam hidroksinamat, flavonols, dan antosianidin.
Total polifenol pada perlakuan sampel (Tabel 4) diketahui tertinggi pada hari ke-6. Meningkatnya perolehan polifenol tersebut karena lisisnya komponen dinding sel oleh enzim selulase, xilanase dari konsorsium aktinomiset. Menurut Huang et al. (2007) bahwa campuran Aspergillus oryzae and Trichoderma reesei
yang mampu menghasilkan enzim selulase dan xilanase mampu meningkatkan 23% perolehan ekstrak asam elagat yang tergolong polifenol pada buah Valonia acorn. Aguilar et al. (2007) melaporkan bahwa fermentasi menggunakan
Aspergillus niger GH1 selama 4 hari dengan SSF mampu meningkatkan
perolehan polifenol dari kulit buah Delima. Laroze et al. (2010); Collao et al.
(2007) juga melaporkan bahwa penggunaan campuran beberapa enzim komersial seperti selulase dan hemiselulase dapat meningkatkan perolehan ekstrak polifenol yang dapat digunakan sebagai antioksidan dari buah resberry dan Oenothera biennis. Maier et al. (2008) menyatakan bahwa penggunaan campuran dua jenis enzim komersial dapat meningkatkan perolehan ekstrak polifenol pada buah anggur. Ekstraksi fenolik antioksidan dari sayuran menggunakan enzim dapat terjadi melalui degradasi hidrolitik polisakarida pada dinding sel. Fenolat tersebut terikat pada lignin dan polisakarida oleh ikatan hidrogen atau hidrofobik. Selain itu mekanisme lain yang mungkin juga dilakukan oleh enzim yaitu langsung merusak ikatan eter atau ester antara fenol dan polimer dinding sel tanaman (Pinelo et al. 2008). Peningkatan senyawa total polifenol juga terjadi karena terlepasnya ikatan antara komponen selulosa, hemiselulosa dan lignin dengan senyawa polifenol. Senyawa polifenol terikat secara kovalen pada dinding sel. Menurut Gonzales et al. (2011), fermentasi padat menggunakan Aspergillus tamarii mampu meningkatkan perolehan ekstrak senyawa fenol berupa asam hidroksinamat. Ekstrak asam hidroksinamat dari kulit kopi tersebut dapat digunakan sebagai senyawa antioksidan.
21 tanin dari buah anggur menggunakan enzim β-galaktosidase meningkat hingga diperoleh 320 mg L-1. Menurut Ramirez-Coronel et al. (2004) kandungan proanthocynidin atau tanin terkondensasi pada kulit kopi mengalami penurunan setelah 3 hari inkubasi. Penggunaan Streptomyces sp. dengan Solid State Fermentation dapat menurunkan kandungan polifenol pada kulit kopi (Orozco et al. 2008). Menurunnya kandungan polifenol terutama tanin di kulit kopi terfermentasi justru menguntungkan karena tanin memiliki dampak negatif yaitu menghambat penyerapan protein. Ozkose et al. (2011) melaporkan bahwa tanin dapat menghambat pertumbuhan bakteri pendegradasi serat pada saluran pencernaan ruminansia. Dengan demikian kandungan tanin yang rendah setelah proses fermentasi menggunakan aktinomiset pada kulit kopi dapat dimanfaatkan sebagai pakan ternak. Peningkatan ekstrak tanin baik kontrol maupun sampel ternyata diikuti dengan peningkatan monomernya yaitu katekin. Perolehan katekin meningkat drastis pada inkubasi hari ke-3.
Peningkatan perolehan hasil ekstraksi senyawa polifenol (Tabel 4) ternyata karena adanya peningkatan salah satu komponennya yaitu antosianin. Perolehan ekstrak antosianin yang lebih tinggi diperoleh pada perlakuan konsorsium aktinomiset. Ekstrak polifenol dan antosianin yang diperoleh dari buah
Blackcurent dapat ditingkatkan dengan menggunakan enzim selulase dan hemiselulase yang dihasilkan oleh Trichoderma spp. (Kapasakalidis et al. 2009). Moreno-peres et al. (2010) melaporkan bahwa ekstrak antosianin yang diperoleh dari buah anggur sbesar 909.48 mg L-1 meningkat karena diekstrak menggunakan
enzim β-galaktosidase. Menurut Jurgonski et al. (2013), penggunaan enzim β -glukosidase dapat meningkatkan peroleh ekstrak antosianin dari Lonicera caerulea hingga 320 mg g-1. Senyawa polifenol seperti halnya antosianin memiliki keterikatan kovalen pada dinding sel. Peningkatan perolehan ekstrak antosianin dari kulit kopi terjadi setelah 3 hari inkubasi (Ramirez-coronel 2004). Prata dan Oliveira (2007) menjelaskan penggunaan kulit buah kopi segar sebagai sumber pewarna alami yang potensial karena kandungan antosianin cyanidin-3-rutinosida. Murthy et al. (2012) melaporkan juga bahwa warna merah pada kulit kopi mengandung cyanindin-3-rutinosida dan cyanidin-3-glukosida yang berpotensi sebagai antioksidan dan pewarwa alami makanan. Selain itu juga Pinelo et al. (2008) menyatakan bahwa dengan adanya pengaruh penggunaan
enzim β-galaktosidase dari Aspergillus niger dan enzim selulase dari Trichoderma reesei mampu meningkatkan perolehan komponen polifenol. Kandungan polifenol terukur meningkat karena adanya peningkatan kandungan asam klorogenik.
SIMPULAN
22
Saran
Perlu dilakukan analisis lebih lanjut mengenai komponen bioaktif non polar seperti komponen alkaloid pada kulit buah kopi hasil fermentasi menggunakan kultur konsorsium Streptomyces exfoliatus 42 dan Streptomyces costaricanus 45I-3
DAFTAR PUSTAKA
Ajila CM, Brar SK, Verma M, Tyagi RD, Godbout S, Valero JR. 2011. Extraction and analysis polyphenol. Biotechnol. 31: 227-249.
Amraini SZ. 2008. Produksi gula dari jagung dengan proses enzimatik secara fermentasi kultur padat. J Teknol Proses. 7: 141-150.
[AOAC] Association of Analytical Communities. 1995. Official methods of analysis of the association of analytical chemists. Washington DC (US): AOAC Int.
Apriani I. 2013. Seleksi isolat aktinomiset untuk degradasi biomassa lignoselulosa [Tesis]. Bogor (ID): Institut Pertanian Bogor.
Aquilar CN, Aquielera-cargo A, Robledo A, Ventura J, Belmares R, Martinez D, Rodriques-herrera R, Contreras J. 2007. Production of antioxidant nutraceuticals by solid-state cultures of pomegranate (Punica granatum) peel and creosote bush (Larrea tridentata) leaves. Food Technol Biotechnol. 46: 218-222.
Aro N, Pakula T, Penttila M. 2004. Transcriptional regulation of plant cell wall
degradation by filamentous fungi. Microbiol. 29: 719-739.
Ashok P, Soccol CR, Nigam P, Brand D, Mohan R, Roussos S. 2000. Biotechnological potential of coffee pulp and coffee husk for bioprocesses.
Biochem Eng. 6:153-162.
Asosiasi Eksportir dan Industri Kopi Indonesia. 2012. Produksi kopi Lampung. [Internet]. Jakarta (ID): AEKI [diunduh 2013 Okt 25]. Tersedia pada:http//http://www.aeki-aice.org/page/bpd-aeki-lampung/id.
Astuti E. 2012. Aktivitas selulolitik dan karakterisasi aktinomiset asal tanah gambut [Tesis]. Bogor (ID): Institut Pertanian Bogor.
Avallone S, Brillouet JM, Guyot B, Olguin E, Guiraud JP. 2002. Involvement of pectolytic micro-organisms in coffee fermentation. Food Sci Technol Int. 37: 191-198.
Balasundram NK, Sundram, Samman S. 2006. Phenolic compounds in plants and agri-industrial by products: antioxidant activity, occurance, and potential uses.
Food Chem. 99: 191-203.
Beg QK, Kapoor M, Mahajan L, Hoondal GS. 2001. Microbial xylanases and their industrial application. Appl Microbiol Biotechnol. 56:326-338.
Brand D, Pandey A, Rodríguez-Leon JA, Roussos S, Brand I, Soccol CR, 2001. Packed bed column fermenter and kinetic modeling for upgrading the nutritional quality of coffee husk in solid-state fermentation. Biotechnol Prog.
23 Bugg TDH, Ahmad M, Hardiman EM, Singh R. 2011. The emerging role for bacteria in lignin degradation and bio-product formation. Curr Opin Biotechnol. 22: 394-400.
Cerda A, Martínez ME, Soto C, Poirrier P, Perez-Correa JR, Vergara-Salinas JR, Zuniga ME. 2013. The enhancement of antioxidant compounds extracted from
Thymus vulgaris using enzymes and the effect of extracting solvent. Food Chem. 139: 138-143
Chandel AK, Chan ES, Rudravaram R, Narasu ML, Rao LV, Ravindra P. 2007. Economics and environmental impact of bioetanol production technologies: Appl Biotechnol Mol Biol Rev. 2: 14-32.
Collao CA, Curotto E, Zuniga ME. 2007. Enzymatic treatment on oil extraction and antioxidant recuperation from Oenothera biennis by cold pressing. Gras Aceit. 58: 10-14.
Dashtban M, Maki M, Leung KT2, Mao C, Qin W. 2010. Cellulase activities in biomass conversion: measurement methods and comparison. Crit Rev Biotechnol. 1-8.
Dalzel HW, Biddlestone AJ, Gray KR, Thurairajan K. 1997. Soil management: compost production and use in tropical and sub tropical environments. Soil Bull. 56: 175-177.
Direktorat Jendral Perkebunan. 2012. Luas areal dan produksi kopi Indonesia. Direktorat Jendral Perkebunan Departemen Pertanian RI (ID): Dirjen Perkebunan.
Diniyah N, Maryanto N, Ahmad S, Demi S, Achmad. 2013. Ekstraksi dan karakterisasi polisakarida larut air dari kulit kopi varietas Arabika (Coffea arabica) dan Robusta (Coffea canephora). J Teknologi Pertanian 14: 73-78. Dubois M, Gilles KA, Hamilton JK, Rebers PA, Smith F. 1959. Colorimetri
method for determination of sugar and related subtance. Anal Chem. 28: 360-356.
El-Sersy NA, Abd-Elnaby H, Abou-Elela GM, Ibrahim HAH, El-Toukhy NMK. 2010. Optimization, economization and characterization of cellulase produced by marine Streptomyces ruber.Afr J Biotechnol. 9: 6355-6364.
Feng Y, Yu Y, Wang X, Qu Y, Li D, Weihua He, Byung Hong Kim. 2011. Degradation of raw corn stover powder (RCSP) by an enriched microbial consortium and its community structure. Bioresour Technol. 102: 742-747. Fengel D dan Wegener G. 1995. Kayu : Kimia, ultrastruktur, reaksi-reaksi.
diterjemahkan oleh Hardjono Sastrohamidjoyo. Yogyakarta (ID): Gajah Mada University Press.
Fontes CM. 2000. A novel cell Vibrio mixtus family 10 xylanase that is both intracellular and expressed under non-inducing conditions. J Microbiol 145: 1959-1967.
[FAO] Food and Agriculture Organization of the United Nations. 2005.
Mechanism of the enzyme cellulase. [diakses 29 Juni 2013]. Tersedia pada: http://www.fao.org/docrep/w7241e/w7241e08.htm.
Gomes DJ, Gomes J, Steiner W. 1994. Factors influencing the induction of endo-xylanase by Thermoascus aurantiacus. J Biotechnol. 33 :87-94.
Gonzalez M, Coronel AR, Mancera M. 2011. Antioxidant activity of fermented and nonfermented coffee (Coffea arabica) pulp extracts. Food Technol