STUD1
ARSITEKTUR POHON DALAM HUBUNGANNYA
DENGAN PERTUMBUHAN DAN PERKEMBANGAN DURIAN
Oleh
ARlS MUNANDAR
AGR 93512
PROGRAM PASCASARJANA INSTITUT PERTANIAN BOGOR
ABSTRAK
ARlS MUNANDAR, 2001. Studi Arsitektur Pohon dalarn Hubungannya dengan Perturnbuhan dan Perkernbangan Durian. Di bawah birnbingan SRI SETYATI HARJADI sebagal Ketua Kornisi dan A. SURKATI ABiDIN, BAMBANG S. PURWOKO, SUDARSONO dan SUHIRMAN sebagai Anggota.
Dalarn rangka mernpelajari hubungan perilaku percabangan
,
bentuk pohon, strategi perturnbuhan dan perkernbangan serta produktivitas durian telah dilakukan empat set penelitian tentang (1) arsitektur sernai (bibit generatif) dan bibit klonal (bibit vegetatif), (2) perilaku percabangan. (3) struktur tajuk, analisis perturnbuhan dan fenologi trubus dan (4) fisiologi source-sink. Penelitian dila- kukan di Kebun Percobaan IPB dan daerah sekitar Bogor pada tahun 1995-1 999. Hasil penelitian rnenunjukkan bahwa terdapat hubungan paralel antara perilaku percabangan, bentuk tajuk, strategi pertumbuhan dan perkembangan tanarnan dan potensi produksi durian. lntervensi fisik (dengan pernangkasan dan training) dapat mengarahkan tanarnan menuju strategi "model" (keseim- bangan pertumbuhan dan perkernbangan tanaman) yang rnendukung potensi produksi, kontinuitas produksi dan kelestarian pohon. lntervensi secara biologi (dengan penernpelan) rnenggunakan bibit generatif yang diketahui asal-usulnya sebagai batang bawah belurn dapat mernberi kepastian bahwa batang bawah tersebut menginte~ensi bentuk tajuk dan sudut cabang karena semai mengalami segregasi.Berdasarkan hubungan paralel bentuk tajuk bibit dengan bentuk tajuk pohon dewasa (tanaman belurn rnenghasilkan rnaupun tanarnan rnenghasilkan, TBM atau TM) dan hasil simulasi pernangkasan dan training disimpulkan bahwa durian menarnpakkan perilaku kendali apikal yang spesifik klon. Perilaku ini tidak dipengaruhi faktor ontogeni maupun pernangkasan dan training. Berdasarkan respons perturnbuhan dan perilaku dirnorfisme percabangan dua klon durian yang berbeda level kendali apikalnya dapat disirnpulkan bahwa pemangkasan dan training untuk suatu jenis durian bersifat unik. Durian dengan kendali apikal kuat (Klon Monthong) sebaiknya dipangkas dan dibentuk dengan pola terbuka tengah (open center), sedangkan yang lernah (Klon Matahari) dipangkas dan dibentuk dengan pola terbuka tengah, palmette leader atau central leader yang diikuti dengan perundukan.
Dalarn keterbatasan waktu, kendala iklim dan faktor ontogeni, pengaruh pernangkasan dan training terhadap perkembangan tanarnan sebagairnana didekati dengan studi fenologi trubus belurn dapat diungkap.
Sukrosa dan oligosakarida (diduga stachiosa), gula non-reduksi yang mobil yang rnempunyai peranana penting dalam fisiologi source-sink, dapat digunakan untuk memprediksi produktivitas durian yang mengalami pernangka- san dan training. Studi lanjut rnengeksplorasi mobilisasi oligosakarida, aktivitas enzim yang terlibat dalarn interkonversi cadangan gula oligosakarida dan mekanisrne pengendalian pelepasannya dari komparternen penyimpanan diper- lukan untuk meninakatkan ~roduktivitas durian dan rnencari rnanfaat praktis seperti aptikasi pupuc mikro delalui daun.
ABSTRACT
ARlS MUNANDAR, 200f. Study on Tree Architecture i n Relation t o Growth and Development of Durian. Under a team o f supervisors w i t h SRI SETYATI HARJADI as chairman, and A. SURKATI ABIDIN, BAMBANG S. PURWOKO, SUDARSONO and SUHIRMAN as members.
In an attempt to study the relationship of branching habit, tree form. strategy of growth and development and productivity of durian four parallel researches concerning (1) generative and vegetative seedling architecture. (2) branching habit, (3) canopy structure, growth analysis and flushing phenology and (4) source-sink physiology were conducted. Those four sets researches were conducted at IPB experimental farm and in area of Bogor District during 1995-1 999.
The results showed that there were parallel relationship between branch- ing habit, tree form, strategy of growth and development and the "potential" productivity of durian. Physical intervention (by pruning and training) could direct the plant toward "model" strategy (balance of vegetative and reproductive growth and development) that support high "potential" productivity, continuous production and tree longevity. Biological intervention (through budding) using generative seedling as identified rootstock was unable to affect characters of the vegetative seedling due to segregation.
Based on parallel relationship between vegetative seedling form. juvenile- to-mature tree form and result of pruning and training simulation, it can be concluded that apical control on durian is clone-specific and it does not depends on ontogeny and pruning or training. Based on the growth response and branching dimorphism of two durjan clones with different level of apical control, it can be concluded that pruning and training for certain clone is unique. Strong apical control clone (Monthong) should be pruned and trained in open center, but the weaker one (Matahari) should be in open center, palmette leader or in central leader followed by bending. This clone-specific practical application improve the previous (the first generation) pruning and training practices that only be gene- ralized at species level.
In limited time, restricted humid climate and ontogenic factor, the training and pruning did not affect tree phenology.
Sucrose and oligosaccharide (suggested as stachyose), which are non- reductive mobile sugars that play important role in source-sink physiology of durian, can be used to predict the productivity of trained and pruned durian. Further study is needed to explore the oligosaccarides mobilization, the activity of enzyme that interconvert the storage of that sugar. and the control mechanism of the release from storage compartments, which can be beneficial for other practical application. such as micro-element foliar fertilizer application.
SURAT PERNYATAAN
Melalui surat ini saya rnenyatakan bahwa disertasi yang berjudul:
STUD1 ARSlTEKTUR POHON DALAM HUBUNGANNYA DENGAN PERTUMBUHAN DAN PERKEMBANGAN DURIAN
adalah benar merupakan hasil karya sendiri dan belum pernah dipublikasi. Semua sumber data dan inforrnasi yang digunakan telah dinyatakan secara jelas
dan dapat diperiksa kebenarannya.
Bogor, November 2001
STUD1 ARSITEKTUR POHON DALAM HUBUNGANNYA
DENGAN PERTUMSUHAN DAN PERKEMBANGAN DURIAN
Oleh
ARlS MUNANDAR
AGR
93512
Disertasi sebagai salah satu syarat untuk memperoleh gelar
D o k t o r pada
Program Pascasarjana lnstitut Pertanian Bogor
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Disertasi :Studi Arsitektur Pohon dalam Hubungannya dengan Pertumbuhan dan Perkembangan Durian
Narna Mahasiswa : Aris Munandar
Nomor Pokok :93532
Program Studi : Agronomi
Menyetujui Komisi Pembimbin.g
Prof. Dr Ir Sri Setyati Hariadi ~ e t u a -
4
Prof. Dr Ir A-Surkati
Anggota Anggota
Dr lr Sudarsono Anggota
?
i,
?4%2,L<&??--.
Dr Ir Suhirrnan Anggota
2. Ketua Program Studi Agronomi
&
k
-Dr Ir Sudirman Yahya
gram
l
a
IPB/ /
,.
Dr Ir Sudirman Yahya
" L ' b :
-?=q,. Piw'Syafrida Manuwoto-- . .
-
6
N D V 2381Penulis Lahir di Bangkalan, Jawa Tirnur, pada tanggal 28 Desember 1956
dari ayah bernama Abdul Rachern ( a h ) dan Ibu Hj Siti Maryarn. Penulis sebagai anak pertarna dari delapan bersaudara.
Penulis rnenempuh sekolah dasar di SD Negeri Plaosan II, Magetan dan SD
Negeri
Kranan Bangkalan hingga tahun 1969, sekolah rnenegah di SMP Negeri I Bangkalan tahun 1969-1972 dan SPMA (Sekolah Pertanian Menengah Atas) Negeri Malang tahun 1972-1975. Penulis rnernperoleh getar Sarjana Pertanian Bidang Agronomi dari Fakultas Pertanian IPB pada tahun 1982 dan Magister Sains pada Bidang llrnu Pengelolaan Surnberdaya Alarn dan Lingkungan. Pada tahun 1993 penulis mengikuti Program Doktor di bidangAgronorni pada Program Pascasarjana IPB.
Pada tanggal 25 Januari 1990 penutis rnenikah dengan Eni Yuiiantini.
Penulis dikaruniai tiga orang putri yaitu lpo Darliyanti. Fadila Rarnadini dan Nadira Elkalarn dan mengangkat seorang putra Sigit Mulyansyah.
Penulis bekerja sebagai staf teknis Balai Penelitian Tanaman Pangan pada tahun 1976-1982. Sejak 1983 penulis bekerja sebagai dosen pada
KATA PENGANTAR
Puji dan syukur penulis panjatkan ke hadirat Allah SWT yang telah melirnpahkan rahrnat-Nya sehingga penulis dapat menyelesaikan disertasi ini. Tulisan ini rnerupakan hasil penelitian disertasi yang ciiinspirasi oleh keinginan menghubungkan aspek arsitektur pohon dengan perturnbuhan dan perkembang- an tanaman durian. Tulisan ini rnenguraikan empat set penelitian yaitu (1)
arsitektur semai (bibit generatif) dan bibit klonal (bibit vegetatif), (2) perilaku percabangan. (3) struktur tajuk, analisis perturnbuhan dan fenologi trubus dan (4)
fisiologi source-sink. Masing-masing hasil penelitian disajikan sebagai bab-bab tersendiri (manuskrip) yang dihantar dengan suatu pendahuluan dan ditutup dengan pembahasan urnum.
Pada kesempatan ini penulis menyampaikan terima kasih kepada:
1. Ibu Prof. Dr Ir Sri Setyati Harjadi sebagai Ketua, Bapak Prof. Dr Ir A. Surkati.
Bapak Dr Ir Bambang S. Puwoko, Bapak Dr Ir Sudarsono dan Bapak Dr Ir Suhirman sebagi Anggota Komisi Pembirnbing yang telah mernberikan birnbingan dan pengarahan selama melaksanakan penelitian dan penyusun-
an disertasi ini.
2. Bapak Prof Dr Ir G. A. Wattimena dan Bapak Dr Winarno yang telah memberikan saran-saran perbaikan disertasi ini.
3. Dekan Fakultas Pertanian IPB periode 1990-1997 dan Dekan periode 1997- 2001 yang telah memberikan kesempatan dan ijin belajar sarnbil bekerja
serta rnenyediakan Kebun Percobaan Cikabayan, Sukarnantri dan Bara- nangsiang untuk tempat penelitian.
4. Bapak H. Aden, Ir Wijaya, MS dan staf Kebun Cipaku (Departemen
Pertanian) atas bantuan penyediaan bibit durian sebagai materi penelitian; Saudara Endi Rohendi, SP; Ir Kosasih (Lab Kirnia Terpadu); Ir Ketty Suketi, MS dan Saudara Fitri; para staf kebun Percobaan Cikabayan. Sukarnantri
5. Kedua orangtua penulis almarhum rarnanda Abdul Rachem dan bunda Hj.
Sjti Maryarn yang telah mengasuh, mernbesarkan dan rnendidik penulis. 6. lstri penulis Eny Yuliantini dan putra-putri penulis Ipo Darliyanti, Fadila
Ramadini, Nadira Elkalam dan Sigit Mulyansyah atas kesetiaan, kesabaran dan ketabahan serta dorongan sernangat yang diberikan selama penulis rnengikuti pendidikan hingga menyelesaikan disertasi ini.
7 . Keluarga dan semua pihak yang telah membantu baik selama pendidikan, pelaksanaan percobaan rnaupun penyusunan disertasi.
Akhirnya penulis menyadari bahwa tulisan ini masih jauh dari sernpurna. Walapun demikian penulis berharap semoga hasil-hasil yang dituangkan dalam
disertasi ini bermanfaat bagi pengembangan tanarnan tropika. Amin.
Bogor, Mei 2001
DAFTAR
IS1
DAFTAR TABEL . .
. .
. . .. . .
. . . .. . .
. . ..
.. . .
. . ..
,.
. . .. . .
. . .. .
, DAFTAR GAMBAR ..... . ...
.. . . .. . ..
.....
.
...
... ....
.. . . . ....
..
. . .....
. .
. .. . DAFTAR LAMPIRAN . . . .. . .
. . . .. . . .
. . ..
.. . .
. . .. . .
. . ..
. . .. . .
. . . . PENDAHULUANLatar Belakang
...
...
...
... ...
......
...
.. .
...
......
...
... . .....
... . Tujuan Penelitian...
... . .....
......
...
...
...
...
...
...
... ... ... ... ,...
Kegunaan dan Relevansi Penelitian...
...
... ... ... .. .
... ... ... .
Daftar Pustaka...
......
... ......
... ... ... ..., ... ... ..,. SET PENELITIAN-1: STUD1 ARSITEKTUR SEMAl DAN BlBlT KLONALAbstrak .
. . .
. . .. . . .
. . .. . .
. . .. . .
..
. . . .. . .
. . ..
.
..
. . .. . . .
. . .. .
,.
Pendahuluan
Latar Belakang ...
...
... .....
.
... . .. ... . .. ... ... ... .. . ... ...
. .. Tujuan Penelitian.
.. ... ......
... .. ....
. .. ......
.. . ... .. . . .. .. . .. . Bahan dan MetodeSub-set Penelitian-I a: Karakterisasi Morfologi Semai . Sub-set Penelitian-I b: Pengaruh Batang Bawah
terhadap Karakter Morfologi Bibit Ternpelan . . . .. Pengamatan
. . . .
. . . .. . .
. . .. .
. . .. . .
. . ..
. . .. . . .
. . , Analisis Statistika...
... ...... ... ... .
..
...... ..
. .....
. .
..
.. . ....
Hasil dan PembahasanSub-set Penelitian-la: Karakterisasi Morfologi Sernai
..
Sub-set Penelitian-lb: Pengaruh Batang Bawahterhadap Karakter Morfologi Bibit Ternpelan ... .. Kesimpulan dan Saran
Kesimpulan .. . .
. . . .
. . .. ..
..
. . .. .. . .
. . . ... . . .
. . ...
.
,.
. . . ...
Saran. . .
. . .. .
. . .. . . .
. . .. . .
.
. . . .. . . .
. . . ,. . .
. . .. . .
.. Daftar Pustaka . . ..
.. . . .
. . .. . . .
. . . . ...
.. . . .
. . . .. . .
. . .. . .
..
..
. . . .. . . .
:.. SET PENELITIAN-2: PERILAKU PERCABANGAN. BENTUK TAJUK DAN ASPEK MORFOMETRI CABANGAbstrak
.
..
. .. .. .
. . . . .. ... .
. . . ... . . .
. . .. . .... .
. .. . ...
. . .
. .. . ... . .
. . .. .. . .
. . PendahuluanLatar Belakang
. .
...
...
......
... ......
...
...
... .,....
... ... . Tujuan Penel~tran ...... ...
......
......
...
......
...
...
......
... . Bahan dan MetodeSub-set Penelitian-2a: Model Tajuk Bibit Klonal . .. . .. Sub-set Penelitian-2b: Hubungan Pernangkasan dan
xiii
Training dengan Kendali Apikal dan Bentuk Tanaman Belum Menghasilkan ...
Sub-set Penelitian-2c: Dimorfisme dan Aspek-aspek Morfometri dan Percabangan ...
Pengamatan ...
.
.
...Analisis Statistika
...
Hasil dan PembahasanSub-set Penelitian-Pa: Model Tajuk Bibit Klonal ... Sub-set Penelitian-2b: Hubungan Pemangkasan dan
Training dengan Kendali Apikal dan Bentuk Tanaman Belum Menghasilkan ...
Sub-set Penelitian-PC: Dimorfisme dan Aspek-aspek Morfometri dan Percabangan ...
Kesimpulan dan Saran
Kesimpulan ...
Saran ...
Daftar Pustaka ...
SET PENELITIAN-3: STRUKTUR TAJUK
.
ANALISIS PERTUMBUHAN DAN FENOLOGI TRUBUSAbstrak ...
Pendahuluan
Latar Belakang ...
Tujuan Penelitian ...
Bahan dan Metode
...
Rancangan Penelitian
Pengamatan dan Analisis ...
Hasil dan Pembahasan
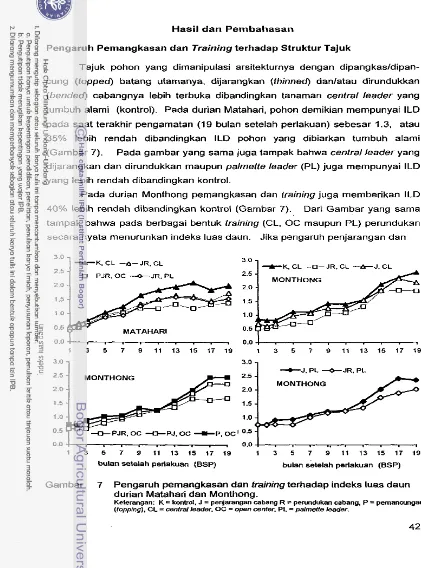
Pengaruh Pemangkasan dan Training terhadap Struktur Tajuk
...
Pengaruh Pemangkasan dan Training terhadapPertumbuhan Tanaman
...
Pengaruh Pemangkasan dan Training terhadapFenologi Trubus ...
Kesimpulan dan Saran
Kesimpulan
...
Saran...
Daftar Pustaka...
SET PENELITIAN-4: FlSlOLOGl SOURCE-SINKAbstrak ...
Pendahuluan
Bahan dan Metode
Sub-set Penelitian4a: Dinamika gula Tanaman Durian yang Mengalami Defoliasi, Pemangkasan Bentuk dan Training ...
Sub-set Penelitian-4b: Fraksi dan Kandungan Gula dalam Floern Cabang dan Daun Cabang "on" dan Cabang "off'
...
Sub-set Penelitian4c: Hubungan Sudut Cabang dengan Fraksi dan Kandungan Gula Floem Cabang dan Daun
...
Pengamatan dan Analisis...
Hasil dan Pembahasan
Sub-set Penelitian4a: Dinamika gula Tanaman Durian yang Mengalami Defoliasi, Pernangkasan Bentuk dan Training
...
Sub-set Penelitian-4b: Fraksi dan Kandungan Gula dalam Floern Cabang dan Daun Cabang "on" dan Cabang "off' ...
Sub-set Penelitian4c: Hubungan Sudut Cabang dengan Fraksi dan Kandungan Gula Floem Cabang dan Daun ...
Kesirnpulan dan Saran
...
Kesirnpulan
Saran ...
Daftar Pustaka ...
PEMBAHASAN DAN KESIMPULAN UMUM
Pernbahasan Urnum
...
Kesimpula Umum...
Daftar Pustaka ...GLOSARIUM
...
LAMPIRAN ...DAFTAR TABEL
Proporsi (%) komponen karakter rnorfologi semai durian
Lokal Bogor Pagar. Mentega, Silodong dan Koclak
...
Frekuensi (%) sernai berdasarkan bentuk tajuk dan sudut cabang
...
Pengaruh penempelan terhadap sudut cabang dan tinggi tanarn- an batang atas durian lokal Pagar. Mentega dan Silodong ...
Pengaruh batang bawah terhadap frekuensi bentuk tajuk, sudut cabang dan keberhasilan penempelan bibit klonal Monthong dan Hepe umur 6 bulan ...
Pengaruh pernbentukan pohon terhadap rata-rata dan distribusi kuartil diameter cabang primer, sudut cabang, nisbah kerunduk- ... an, plagiotropi dan reiterasi durian T5M Matahari
Karakter tajuk durian yang ditanam alami (tanpa pernbentuk- an pohon, Kebun Tarnan Buah Mekarsari, Cileungsi, Bogor ..
Pengaruh aktivitas sink terhadap konsentrasi gula fioem cabang durian dengan sudut cabang relatif datar (cabang plagiotrop) ...
Pengaruh aktivitas sink terhadap konsentrasi gula daun tua .... durian dengan sudut cabang relatif datar (cabang plagiotrop)
Kerangka studi arsitektur pohon dalam hubungannya dengan perturnbuhan dan perkernbangan tanaman durian
...
Penjelasan grafis perlakuan kontrol (K), pemancungan batang utarna (P), penjarangan cabang (J) dan pemndukan cabang (R) untuk menghasilkan tanaman dengan pola central leader (CL), open center (OC) dan palmette leader (PL)
...
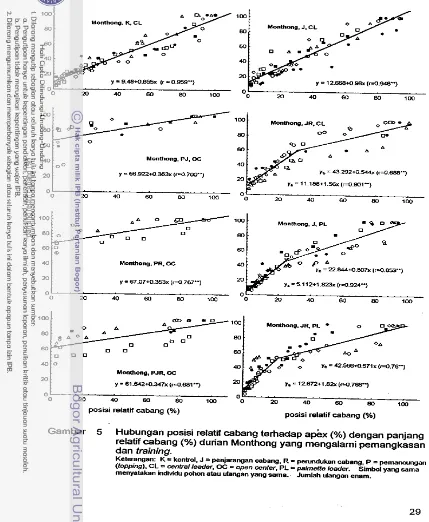
Hubungan posisi relatif cabang terhadap apex (%) dengan panjang relatif cabang (%) bibit klonal durian Matahari dan
...
MonthongHubungan posisi relatif cabang terhadap apex (%) dengan panjang relatif cabang (%) durian Matahari yang mengalami pemangkasan dan training
...
Hubungan posisi relatif cabang terhadap apex (%) dengan panjang relatif cabang (%) durian Monthong yang mengalarni pemangkasan dan training
...
Pengaruh pemangkasan dan training terhadap nisbah volume1 area batang dan cabang durian Matahari dan Monthong ...
Pengaruh pemangkasan dan training terhadap indeks luas daun durian Matahari dan Monthong
...
Pengaruh pemangkasan dan training terhadap bobot kering total durian Matahari dan Monthong
...
Pengaruh pemangkasan dan training terhadap laju tumbuh absolut (LTA) durian Matahari dan Monthong
...
Pengaruh pernangkasan dan training terhadap laju tumbuh
...
relatif (LTR) durian Matahari dan MonthongPengaruh pemangkasan dan training terhadap laju asirnilasi bersih (LAB) durian Matahari dan Monthong
...
Pengaruh pemangkasan dan training terhadap partisi bahan kering durian Matahari
...
Pengaruh pemangkasan dan training terhadap nisbah bahan
kering cabangfbatang durian Matahari dan Monthong
...
49 Pengaruh pemangkasan dan training terhadap ritme trubus15 Gambar skernatik lokasi dan bentuk asimilat yang terlibat dalam penyimpanan sernentara (transient storage) asirnilat (Dietz dan
Keller, 1997) ... 59
16 Pengaruh bentuk training dan aktivitas sink trubus terhadap
kandungan gula floem cabang ... 66
97 Pengaruh bentuk training dan aktivitas sink trubus terhadap
kandungan gula daun tua
...
6618 Pengaruh bentuk training dan aktivitas sink trubus terhadap
kandungan gula daun rnuda ... 69
19 Hubungan sudut cabang dan aktivitas sink dengan kandungan
[image:25.526.57.451.18.570.2]DAFTAR LAMPIRAN
1 Percabangan durian dengan perlakuan pemangkasan, penja- rangan dan perundukan atau PJR dibandingkan kontrol, dibiar-
kan alami atau K pada tanaman urnur 3.5 tahun
... ...
... ...... ... ...
89 2 Koefisien korelasi sudut cabang dengan kadar beberapa jenisgula pada floem batang utama, floem cabang dan daun dengan
PENDAHULUAN
Latar Belakang
Durian merupakan tanaman indigenus Asia Tenggara yang mempunyai prospek baik. Produksinya di Indonesia selama 1989-1998 meningkat dari 139.2 ribu rnenjadi 405.1 ribu ton atau sekitar 12% per tahun (Ditjen Tanaman Pangan dan Hortikultura, 1999). Walaupun lndonesia terrnasuk net-importer, namun
nilai ekspor durian tercatat pernah mencapai US $ 698.2 ribu pada tahun 1997, sebelum krisis ekonorni. Tanaman ini dibudidayakan secara alamiah di peka- rangan atau kebun berskala kecil dan produktivitasnya rendah yaitu 6.2-13.5 ton1 ha. Berdasarkan analogi terhadap potensi produksi buah-buahan yang secara alamiah tajuknya berstruktur sederhana seperti nenas, pepaya dan pisang (Borchert. 1976). kelapa sawit atau pohon dengan tajuk "bentukan" seperti kopi (Halle, Oldeman dan Tornlinson, 1978), apel, pir dan buah-buahan famili Rosa-
ceae (Ryugo, 1988). potensi produksi durian 25 tonlha (Verheij dan Coronel, 1992) diperkirakan dapat dicapai dengan pembentukan pohon. Pembentukan pohon in1 dllakukan dengan penernpelan, penyambungan (Hartmann, Kester dan Davies, 1990) pemangkasan dan training (Ryugo, 1988).
Secara botani durian menarnpilkan model arsitektur Roux (Halle et al., 1978 dan Subhadrabandhu, Scheemann dan Verheij, 1992). Tanaman dengan model ini berbatang rnonopodial dengan ritme pertumbuhannya kontinyu dan cabangnya rnengalami dimorfisme, seperti halnya kopi. Observasi pertanarnan durian di Bogor, Cianjur. Cileungsi dan Leuwilang menunjukkan bahwa durian yang ditanam atau turnbuh atami rnempunyai karakter sebagai berikut (1) tajuk
kompleks, (2) cabang bersudut besar relatif lama terbentuk (lebih dari 5 tahun), hanya terdapat pada basal tajuk dan diarneternya relatif kecil (3) pohon sangat
tinggi (mencapai 40 meter) karena cabang basal mengalami self-pruning secara gradual, (4) berbuah alternate bearing dan (5) trubus (flushing) terjadi bersamaan dengan perkernbangan reproduktif, (6) pada kondisi tajuk yang gelap organ reproduktif berdediferensiasi. menjadi organ vegetatif. Pernbentukan pohon
...
Perilaku percabang- Dimorfisme cabangan spesies durian, model arsitektur Roux (Halle et a/., 1978)
1 I
I
Intervensi: penem- Bentuk tajuk alarni
[
Bentuk tajuk bentukanI
pelan (SP-I). pe- I I
mangkasan-din training (SP-21314)
plagiotrop <c. plagiotrop >>.
> I buah (CL
sudut cc
Kendali apikal (KA)
+
bentuk tajuk (cane.1997 dan Wifson. 2000). bergantung jenis (SP-112)
Strategi
pertumbuhan dan perkernbangan
Pertumbuhan dan
perkembangan 0 cabang <c
-
ca cabang >> (SP-3)-
trubus berlebihan-
* twbus proporsionalreaterasi
Fenomena defisit kar-
bohidrat (SP4) dan pruning da n anernale
manajemen kebun (PHTl bearing (cadangan
IPM, poltinasi buatan, perneli- asirnilat cukup)
haraan fruitsel dan panen) Manajemen mudah
Produktivitas Prod. tinggi (prod.,
[image:28.526.43.466.28.572.2]kontinuitas & kelestarian)
...
Gambar 3 . Kerangka studi arsitektur pohon dalam hubungannya dengan pertumbuhan dan perkembangan tanaman durian.
Keterangan: SP-1f21314 berturut-turul rnenunjukkan set penelitianfstudi. SP. (1)
arsitektur semai dan bibit klcnal. (2) psrilaku percabangan. (3) struktur tajuk, analisis perturnbuhan dan fenologi lrubus dan (4) fisiologi source-sink durian. CL = central leader. OC = open center dan PL = palmette leader. Bentuk
a
= kolumnar atau kubah, bentuk A = kerucut. LTA = Iaju turnbuh absolut. LTR = laju turnbuh relatif dan LAB = laju asimilasi becsih. Tanda >> = banyaklbesar dan c< = sedikitlkeul. Seni membentuk pohon untuk rnemperoleh tanaman superior telah dimu-lai sejak lama (sekitar 100 tahun yang lalu) pada tanadan bernilai ekonomi tinggi kakao, kopi (Halle et a/., 1978). tanaman sub tropika apel, pir, peach
(Verheij dan Coronel. 1992) dan famili Rosaceae lain (Ryugo. 1988). Namun demikian kecuali terhadap mangga, seni ini belum banyak dipraktekkan untuk
Pemangkasan dan training durian baru dimulai tahun 1985 di Thailand (Yaacob dan Subhadrabandhu. 1995) dan awal 1990 di Malaysia (Muhammed,
1994). Pemilihan batang antara (interstock) yang unggul di Thailand diorienta- sikan untuk merekayasa ketahanan terhadap penyakit Phytophtora (Poenvanto.
1996), belum diorientasikan untuk memanipulasi bentuk pohon. Pembentukan pohon pada era tersebut di atas terhadap tanaman model Roux dan Rauh sebagaimana tanaman bernilai ekonomi tersebut terdahulu, bertujuan menginter- vensi perilaku dimofisme dan polimorfisme percabangan dengan atau tanpa meniadakan dominansi apikal (Halle et a/., 1978 dan Verheij dan Coronel, 3992).
Dengan intewensi demikian diharapkan terbentuk cabang plagiotrop atau spurs yang lebih banyak dengan pertumbuhan yang lebih baik. Mengingat cabang plagiotrop pada tanaman model Roux merupakan tempat menempelnya buah maka cabang ini merupakan aset yang bernilai ekonomi. Dimorfisrne merupa- kan suatu perilaku yang spesifik bagi spesies-spesies dalarn kelompok pohon dengan model arsitektur Roux (Halle et a/., 1978). Pembentukan pohon dalam
era tersebut dapat dikatakan merupakan "generasi pertama".
Berdasarkan tinjauan (review) Cline (1997) dan Wilson (2000) serta hasil percobaan Cook, Rabe dan Jacob (1999) pada buah-buahan famili Rosaceae. intervensi tidak cukup pada dirnorfisme cabang dan dominansi apikal, tetapi juga perlu berorientasi pada kendali apikal. Fenomena ini merupakan suatu perilaku percabangan yang spesifik varietas atau klon untuk mengendalikan perirnbangan alokasi asimilat antar cabang dan antara cabang dan batang. lntervensi yang spesifik varietas atau klon ini menandai "generasi ke-dua" pembentukan pohon.
Buah-buahan famili Rosaceae menampilkan model arsitektur Rauh model
yang mirip dengan model Roux pada durian (Halie et a/., 1978). Tanarnan kedua model ini berbatang monopodial, tetapi ritme tumbuh model Rauh episodik
sedangkan Roux kontinyu. Percabangan tanaman model Rauh mengalami polimorfisme yang mernbentuk spurs, sedangkan cabang tanaman model Roux mengalami dimorfisme. Respon tanaman berarsitektur Roux, khususnya durian.
terhadap pembentukan pohon belum banyak diketahui.
lntervensi dimorfisme percabangan, dominansi apikal dan kendali apikal baik pada generasi pertama maupun ke-dua pembentukan pohon tersebut diha-
diharapkan dapat rnengurangi ( I ) reiterasi, (2) pertumbuhan vegetatif yang terla-
lu jagur (too vigorous) dan (3) pauperasi (Halle eta!., 1978) yang rnenyebabkan struktur tajuk kornpleks, dengan nisbah volumelarea batang dan cabang kecil. dan kehilangan energi (entropi). Secara akumulatif model keseimbangan ini diharapkan menjamin produktivitas tinggi (Gambar 1). Produktivitas rnenunjuk- kan produksi per satuan has, kontinuitas produksi tahunan dan kelestarian pohon karena partisi bahan kering dan asimilat yang rnenguntungkan. Menurut
kerangka penelitian ink, pariisi bahan kering didekati dengan analisis pertumbuh- an sedangkan partisi asirnilat dengan fisiologi source-sink tanarnan. Bentuk
(frame) pohon yang baik selain menguntungkan dari segi ekofisiologi juga memberikan kernudahan dalarn rnanajemen kebun seperti pengendalian hama terpadu, pelaksanaan polinasi buatan, pemeliharaan dan pemanenan.
Pada banyak tanaman yang telah lama dikenal rnernpunyai nilai ekonorni
baik. seperti kopi dan kakao (Gopat et
a/.,
1975; Janardan et a/.. 1977; Halle et a/. , 1978; Ramaiah dan Venkatararnanan. 1987; dan Balasimha. 1987). zaitun (Stutte dan Martin, 1986), peach (Bassi, Dirna dan Scorza. 1994) rnaupun buah- buahan farnili Rosaceae (Ryugo. 1988), inforrnasi hubungan bentuk pohon dengan perilaku percabanga, pertumbuhan dan perkernbangan tanarnan seba- gaimana dihipotesiskan penelitian ini (Gambar I ) , telah banyak diketahui.Bahkan sejalan dengan kernajuan teknik-teknik analisis khususnya teknik krorna- tografi pada tahun 1980-an dan teknik pemurnian enzirn, informasi fisiologi tanaman telah tersedia hingga tingkat sub selular (Loescher. 1987, Yamaki dan Ishikawa, 1986 dan Negrn dan Loescher, 1981). Dinamika dan fungsi gula selain sukrosa, yaitu gula alkohol seperti sorbitol dalam fisiologi source-sinkdan bahkan
dalarn fisiologi stress telah diketahui dan ditinjau (reviewed) dengan baik (Dietz dan Kelle. 1997 dan lglesias dan Podesta. 1997). Namun demikian pada banyak
tanaman lain yang berbeda sifat dan karakter botani dan fisiologinya, yang tumbuh di alam tropika dan tergolong energy-rich fruit seperti halnya durian, inforrnasi pada umumnya rnasih terbatas pada aspek-aspek morfometri (Verheij dan Coronel. 1992; Muharnrned. 1994; dan Ali. Jaffar dan Lin, 1995). Studi lebih
mendatam rnernpelajari fisiologi tanaman durian b adilakukan oleh Rarnlan et ~
al. (1 996) yaitu rnempelajari kapasitas fotosintesis durian yang ditanarn alarni. Dari uraian tersebut di atas dapat disimpulkan bahwa pada tanaman tropika umumnya rnasih terdapat kesenjangan informasi hubungan bentuk, peri-
itu karena kerniripan kedua model arsitektur, yaitu Rauh dan Roux, ini rnengan- dung perbedaan-perbedaan, rnaka mernpefajari hubungan-hubungan bentuk. perilaku percabangan dan intervensi perilaku percabangan secara fisik dan biologi terhadap pertumbuhan dan perkembangan durian relevan dipelajari.
Tujuan Penelitian
Mengingat keterbatasan bahan dan waktu untuk rnelakukan penelitian serial, rnaka dilakukan penelttian paralel. Penelitian terdiri atas ernpat set
penelitian (SP). Secara urnum keernpat set penelitian paralel bertujuan rneng- eksplorasi dan rnernbuktikan hubungan bentuk tajuk, struktur dan perilaku perca- bangan, strategi pertumbuhan dan perkembangan serta produktivitas (Garnbar
1). Secara khusus masing-masing set penelitian bertujuan sebagai berikut:
1. Mengeksplorasi petuang rnengintervensi bentuk tajuk dan percabangan
rnelalui penempelan dengan rnenggunakan semai (bibit asal biji) sebagai batang bawah.
2. Mernpelajari hubungan perilaku percabangan (kendali apikal dan dimorfisrne)
bentuk tajuk dan aspek rnorfornetri durian fase bibit dan dewasa.
3. Mernpelajari pengaruh pemangkasan dan pola training terhadap struktur
tajuk dan pertumbuhan dan perkernbangan tanaman dua klon durian yang rnernpunyai kendali apikal berbeda.
4. Mernpelajari fisiologi source-sink atau dinarnika gula pada beberapa tahap perkembangan tanaman, pola training dan struktur percabangan.
Kegunaan d a n Relevansi PeneIitian
Studi arsitektur pohon rnernpunyai kegunaan dan relevansi dari segi ( 2 ) produktivitas pohon sebagai individu dalarn sistern produksi pertanian dan ( 2 ) orientasi pohon dalarn ruang (spatial) dalam pengertian ekoIogi dan arsitektur
pertarnanan. Produktivitas tinggi dicapai dengan produksi per pohon yang tinggi dengan jarak tanarn besar atau produksi per pohon sedang tetapi jarak tanam kecil. Pernbentukan arsitektur pohon (pernbentukan pohon) berarti i n t e ~ e n s i secara fisik rnaupun biologi untuk merubah bentuk atqu "frame" tajuk dalam pengertian botani rnenjadi struktur, bentuk atau frame bentukan yang
rnemfasilitasl hubungan efektif dan efisien antara struktur pohon dengan fungsi pertumbuhan dan perkembangan pohon, terjadi hubungan efektif dan efisien
pohon meningkat dari rendah rnenjadi sedang atau tinggi. Struktur tanarnan dengan tajuk bentukan diharapkan dapat (1) menghindari diskontinuitas produksi
karena alternate bearing dan (2) menjaga pohon dan cabang-cabangnya dari self pruning karena ketidakseirnbangan source dan sink dan atau kekurangan cahaya.
Dari segi ekologi dan pertamanan, rnanipuiasi arsitektur pohon mernpu- nyai relevansi dan bermanfaat untuk penataan tanaman secara keruangan
(spatial) dan ekologi (Kuiper,1994). Dalarn konteks ini arsitektur pohon dapat berguna untuk dasar merancang suatu sistern silvikultur yang rnenjarnin tingkat
diversitas yang tinggi dengan input pengelolaan yang rendah dan lestari. Arsitektur pohon mempunyai relevansi dengan penataan tanarnan secara spatial pada taman lingkungan, taman kota dan pekarangan rnenurut struktur etage
bouw. Sebagaimana diketahui, struktur etage bouw yang telah lama dipraktekkan
di pekarangan di Pulau Jawa menampilkan diversitas yang tinggi (Setyati Harjadi, 1989). Berwisata di kebun buah-buahan dengan tajuk dalarn 'skala jangkauan' akan memberikan pengalaman rnenarik.
Dari segi arsitektur pertamanan, arsitektur pohon mempunyai nilai penting karena dapat rnenghasilkan variasi bentuk tajuk secara terstuktur. Di taman lingkungan maupun tarnan kota dapat dibuat pohon dengan tajuk berbentuk
kubah, kerucut, kolumnar atau payung dengan struktur yang rapi, dengan ketinggian bebas cabang tertentu dan sebagainya. lntervensi dapat lebih rneng- ekpose atau rnenonjolkan bunga agar tarnpak lebih ekstensif sebagai gerornbol pada suatu tiers, sepanjang cabang plagiotrop, permukaan tajuk dan sebagainya. Tampilan demikian menimbulkan keindahan visual baik secara temporal rnaupun
spatial. Kehadiran tanaman berbunga dan berbuah intensif pada pohon dengan tajuk bentukan dapat memberikan penekanan (emphasis) dan kontras yang
mernikat (eye catching).
Dalam penelitian ini tanaman durian dipilih sebagai model untuk mern- pelajari arsitektur pohon karena (1) durian secara alami mempunyai bentuk
memperkaya referensi teknik budidaya tanaman dengan model arsitektur Roux yang buahnya padat energi (energy-rich fruit) dan merangsang penelitian lanjut
fisiologi khususnya fisiologi source-sink tanaman tropika pada tingkat jaringan maupun sub-selular.
Daftar Pustaka
Ali, S. H., M. Y. Jaffar and T. S. Lin. 1995. Branch size as a factor affecting fruit quality. p:53-59. ;n W.M. Wan Othman et
at.
(Eds). Commercial Production of Fruits, Vegetables and Flowers. Universiti Pertanian Malaysia, Selangor.Bassi, D., A. Dima and R. Scorza. 1994. Tree structure and pruning response of six peach growth form. J. Amer. Soc. Hort. Sci. 119(3):378-382.
Batasimha. D. 1987. Cocoa. p:263-285. In M.R. Sethuraj and A.S. Raghavendra (Eds.) Tree Crop Physiology. Elsevier. Tokyo.
Borchert. R. 1976. Feedback control and age-related changes of shoot growth in seasonal and non-seasonal climates, p:497-515. In P. 6. Tomlinson and M. H. Zimmermann (Eds). Tropical Trees As Living Systems. Cambridge Univ Press, Cambridge.
Cline. M. G. 1997. Concepts and terminology of apical dominance. Arner. J. Bot. 84:1064-1069.
Cook. N. C.. E. Rabe and G. Jacobs. 1999. Early expression of apical control regulates length and crotch angle of sylleptic shoots in peach and nectarine. HortScience 34(4):604-606.
Dietz. K. J. and F. Keller 1997. Transient storage of photosynthates in leaves, p:717-737. In M. Pessarakli (ed). Handbook of Photosynthesis. Marcell Dekker, Inc. New York.
Ditjen Tanaman Pangan dan Hortikultura. 1999. Vademekum Pemasaran 1990- 1999. Departemen Pertanian. Jakarta. 253 p.
Gifford, R. M.. J. H. Thorne, W. D. Hitz and R. T. Giaquinta. 1984. Crop productivity and photoassimilate partitioning. Science. 225801 -808.
Gopal, N. H., K. I. Raju, D. Venkataramanan and K. V. Janardhan. 1975. Physiological studies on flowering in coffee under South Indian condit- ions. Ill. Frowering in relation to foliage and wood starch. Turrialba 25(3):239
Halle, F., R. A.A. Oldeman and P. 6 . Tomlinson. 1978. Tropical Trees and Forest: An Architeciural Analysis, Springer-Verlag, Berlin. 441 p.
Hartmann, H. T.. D. E. Kester and F. T. ~ g v i e s . Jr. 1990. Plant Propagation: Principles and Practices. Fifth Ed. Prentice Hall Career and Technology. New Jersey. 647 p.
Kuiper, L. C. 1994. Architectural Analysis of .Douglas-fir Forests. PhD thesis. Wageningen Agricultural University. The Netherland. 186 p.
Loescher, W. H. 1987. Physiology and metabolism of sugar alcohol in higher plant. Physiol. Plant. 70:553-557.
Muhammed, A. G. 1994. Pemangkasan durian: membentuk pokok dan rneningkat potensi pembuahan. p:47-58. In M. Osman et a/. (Eds). Recent Development in Durian Cultivation. MARDI and Malaysian Soc. Hort. Sci., Serdang.
Negm. F. 8. and W. H. Loescher. 1981. Characterization and partial purification of aldose-6-phospate reductase (aldito-6-phospate:NADP-1-oxidoreduct- ase) from apple leaves. Plant. Physiol. 67:139-142.
Poerwanto, R. 1996. taporan kunjungan ke kebun buah-buahan di Thailand. Laporan Perjalanan Dinas tanggal 4-12 Mei 1996. Fakultas Pertanian IPB, Bogor (tidak dipublikasi).
Ramaiah. P. K. and D. Venkataramanan. 1987. Coffee. p:247-262. In M.R. Sethuraj and A.S. Raghavendra (eds). Tree Crop Physiology. Elsevier. Amsterdam.
Ramlan, M. F., A. S. Mamat. H. Siti Aishah and R. Yusoh. 1996. Photosynthetic capacity of seven durian clones grown under natural condition, p:125- 136. Proc. Int. Conf. Trop. Fruits. Malaysian Agric. Res. Dev. Inst. (MARDI), Kuala Lumpur 23-26 Juli 1996.
Ryugo, K. 2988. Fruit Culture: Its Science and Art. John Wiley and Sons, New York. 344 p.
Setyati Harjadi, S. 1989. Pengantar Hortikultura. PAU llmu Hayati. IPB. Bogor.
Stutte. G.W. and G.C. Martin. 1986. Effects of fight intensity and carbohydrate reserves on flowering in olive. J. Amer. Soc. Hort. Sci. ? 1 I (1):27-31.
Subhadrabandhu, S., J. M. P. Scheernann and E. W. M. Verheij. 1992. Durio zibethinus Murray, p:147-161. In E. W. M. Verheij and R. E. Coronel (Eds). Plant Resource of South East Asia No 2: Edible Fruits and Nuts. PROSEA, Bogor.
Verheij. E. W. M. and R. E. Coronel. 1992. Plant Resource of South-East Asia No.2: Edible Fruits and Nuts. PROSEA Bogor, p:1-56.
Wilson, 6. F. 2000. Apical control of branch growth and angle in woody plants. Amer. J. Bot. 87:601-607.
Yaacob, 0. and S. Subhadrabandhu. q995. The Production of Economic Fruits in Soutl-.-East Asia. Oxford University Press. Kua,la Lurnpur, 419 p.
SET PENELITIAN-I
:
STUD1 ARSITEKTUR
SEMAI DAN BIBIT KLONAL
Abstrak
Dua sub-set penelitian dilakukan untuk mempelajari karakter bibit gene- ratif (semai) dan mengeksplorasi kemungkinan penggunaan sernai tersebut sebagai batang bawah. Dari penelitian ini, diperoleh bahwa diperlukan tujuh kornponen utama dengan 13 karakter untuk menjelaskan sekurang-kurangnya 60% keragaman semai antar jenis durian. Fakta ini menunjukkan bahwa populasi batang bawah bersegregasi dan mernperkuat tinjauan (review) bahwa durian merupakan tanarnan menyerbuk silang dan self sterile. Namun demikian di antara 13 karakter morfologi, bentuk tajuk dan sudut cabang merupakan karakter yang berkontribusi cukup besar. Perbandingan antar dua populasi kategori bentuk (kerucut dan kolumnar) mencapai 17.5 : 82.5 (Mentega) dan 18.1 : 81.9 (Koclak). Perbandingan dua populasi kategori sudut cabang (< 60°
dan
r
60°) mencapai 14.9 : 85.1 (Pagar). Namun perbandingan 50:50 dijumpai pada populasi lainnya.Percobaan penempelan dengan batang atas dan batang bawah semai yang sejenis menunjukkan tidak adanya perubahan kategori karakter sudut cabang. Dengan metode yang lebih tepat diperkirakan batang bawah yang bersudut besar tetap dapat memberikan sudut besar terhadap bibit klonal.
Berdasarkan percobaan ini hubungan bentuk dengan perilaku plagio- tropi, yang direfleksikan oleh sudut cabang, tidak dapat diketahui karena batang bawah bersegregasi dan obsewasi karakter bersifat kategorisasi terhadap suatu populasi. Oleh karena itu penggunaan semai sebagai identified rootstock untuk memperoleh bibit klonal dengan bentuk tajuk dan sudut cabang tertentu belum dapat direkornendasikan, karena masih memerlukan penelitian lebih lanjut dengan pendekatan perbanyakan vegetatif dan pendekatan yang lebih kuantitatif (seperti model topofisik tajuk).
Pendahuluan
Latar Belakang
Di Indonesia durian diperbanyak secara vegetatif dengan penernpelan
atau penyambungan dengan batang bawah yang tidak diketahui (unidentified rootstock) dan dibudidayakan secara alami tanpa pernbentukan pohon sebagai- mana banyak dilakukan pada tanarnan buah-buahan. Pada budidaya banyak pohon buah-buahan manipulasi arsitektur pohon diarahkan untuk rnemperoleh
Manipulasi arsitektur pohon dilakukan mulai fase bibit dan juvenil agar struktur cabang dan pernbentukan jaringan pada cabang tidak rnenyebabkan
gangguan pada sistem transtokasi (Ryugo. 1988 dan Wilson, 1990) dan tidak menuntut pemangkasan dan training yang berlebihan ketika tanaman dewasa (Cline, 1997 dan Cook et
a/.,
1999). Pada fase bibit keberhasilan pernbentukanpohon akan dipengaruhi oleh karakter batang bawah rnaupun batang atas (Hartmann
et
a/.,
1990).Penempelan menggunakan semai sapuan (unidentified rootstock) meru- pakan cara yang larim dilakukan untuk perbanyakan durian. Mengingat peng- gunaan bibit stek (cutting) sebagai batang bawah yang belum memungkinkan. penggunaan semai yang teridentifikasi karakter cfan asal pohon induknya (identified rootstock) diperkirakan mernpunyai prospek yang baik. Oleh karena itu peluang ini dieksplorasi dengan mernpelajari karakter semai dan kornbinasi
sernai sebagai batang bawah dengan batang atas durian unggul.
Sejauh ini sumbanoan karakter bentuk tajuk dan sudut cabang di antara karakter morfologi yang biasa digunakan untuk mendeskripsikan (Tjitrosoeporno. 2000 dan IBPGR, 1980) durian dalam menjelaskan keragaman antar jenis semai belurn diketahui dan karakter tersebut belum digunakan untuk seleksi batang bawah. Di Thailand durian telah ditanam dari bibit klonal dengan batang antara
(interstock) yang diidentifikasi atas dasar ketahanannya terhadap penyakit Phytophtora (Poerwanto, 1996). dan bukan atas dasar bentuk tajuk dan sudut cabang. Oleh karena itu studi tentang struktur semai dan bibit klonal dilakukan sebagai Set Penelitian-1 (SP-1)
Tujuan Penelitian
Set Penelitian-I terdiri atas dua sub set penelitian yang bertujuan:
a. Melakukan karakterisasi rnorfologi semai untuk mempelajari sumbangan ka- rakter sudut cabang dan bentuk tajuk terhadap keragarnan antar jenis?
b. Mernpelajari pengaruh batang bawah yang diidentifikasi terhadap karakter rnorfologi bibit tempelan.
Bahan dan Metode
Sub-set Penelitian-la: Karakterisasi Morfologi Semai
Sejalan dengan konteks penelitian untuk mempelajari struktur pohon, penelitian ini bertujuan untuk mempelajari sumbangan karakter rnorfologi sudvt
Sernai durian diarnbil dari biji yang berasal dari pohon yang sama durian unggul lokal Bogor Pagar, Mentega, Silodong dan Koclak. Untuk mengurangi keragam- an, biji untuk bahan benih suatu jenis diarnbil dari pohon yang sama. Pohon induk asal sernai ini yaitu klon Mentega (M), Pagar (P), Silodong (S) dan Koclak (K) masing-masing berturut-turut bertajuk kolumnar dan bersudut besar; kolurn-
nar dan bersudut besar; kerucut dan bersudut besar; kerucut dan bersudut kecil. Penelitian dilakukan di Kebun Jurusan Budidaya Pertanian Fakultas Per- tanian IPB ~aranangsiang (selanjutnya disebut Kebun Baranangsiang) pada tahun 1997. Semai berasal dari biji yang diambil dari Kebun Haji Aden, Desa Sukaiuyu Bogor. Pohon induk durian Pagar. Mentega, Silodong, Koclak sebagai
asal sernai dan batang atas (scion) bibit klonal berumur sekitar 20 tahun. Karakterisasi rnorfologi semai dilakukan pada bulan September-Nopernber 1996 atau urnur sernai 6-8 bulan.
Sub-set Penelitian-9 b: Pengaruh Batang Bawah terhadap Karakter Morfologi Bibit Tempelan
Penelitian ini bertujuan rnernpelajari pengaruh batang bawah yang dise- leksi dan terhadap karakter rnorfologi bibit klonal tempelan. Batang bawah yang digunakan adalah sernai durian lokal Bogor Pagar (P), Mentega (M), Silodong (S)
dan Koclak (K). Batang atas yang digunakan adalah durian unggul nasional Monthong dan Hepe dan durian yang sejenis dengan batang bawahnya. Dengan dernikian penelitian rnernbandingkan karakter dan pertumbuhan:
1. Sernai P dengan bibit klonal P-P. M dengan M-M, S dengan S-S untuk
rnernpelajari pengaruh penempelan dibandingkan sernainya.
2. Bibit klonal Hepe dan Monthong masing-masing dengan batang bawah P, M,
S atau K untuk rnernpelajari pengaruh batang bawah terhadap sudut cabang dan pertumbuhan atau tinggi tanaman.
Mengingat bibit kional mengalami penempelan dan tunas yang diamati tunas yang muncul dari rnata ternpel rnaka semai P, M. S dan K sebagai perlakuan kontrol dipangkas bersamaan dengan masa penernpelan. Tunas semai yang diarnati adalah tunas yang tumbuh setelah pernangkasan.
Pohon induk durian Pagar, Mentega. Silodong, Koclak sebagai asal sernai dan batang atas (scion) bibit klonal berumur sekitar 20 tahun. Batang atas (scion) Hepe dan Monthong diarnbil dari pohon yang telah b e ~ r n u r masing- rnasing 42 tahun dan 8 tahun di Kebun Percobaan atau lnstalasi Penelitian
klonal Monthong dan Hepe dilakukan di Kebun Jurusan Budidaya Pertanian.
Fakuttas Pertanian IPB Baranangsiang Bogor pada bulan Maret 1997. Pengarnatan bibit klonal Hepe, Monthong serta semai (sebagai perlakuan kontrol) durian Pagar, Mentega, Silodong dan Koclak dilakukan pada bulan Nopember 1997 atau pada urnur 8 bulan.
Pengamatan
Karakter morfologi. Karakterisasi rnorfologi didasarkan pada Tropical
Fruit Descriptor yang diterbitkan IBPGR (1980), Suketi (1994), Deskripsi Durian Unggul Nasional (Departernen Pertanian, 1995) dan hasil observasi di Kebun Durian Warso Farm Bogor dan Taman Buah Mekarsari, Cileungsi
Bogor. Pengarnatan rneliputi rnorfologi sebagai berikut:
1. Bentuk tajuk bibit secara keseluruhan yang dibedakan dalarn 2 kategori
yaitu: kerucut dan kolumnar. Bentuk tajuk kerucut rnewakili bentuk-bentuk kerucut tumpul atau kerucut dengan bagan basal lebar, sedangkan
kolumnar selain rnewakili bentuk kolom juga mewakili bentuk kubah.
2. Sudut cabang (khususnya 3 cabang basal) yang dibedakan dalarn 2 kategori yaitu < 60° dan 2 60".
3. Distribusi cabang secara radial yang dibedakan dalarn 3 kategori yaitu: radial, satu sisilbidang dan carnpuran.
4. Warna batang utarna yang dibedakan dalam 4 kategori yaitu hijau, coklat, coklat keabuan dan abu-abu.
5. Bentuk batang utarna yang dibedakan dalarn 3 kategori yaitu segi ernpat, silinder dan lainnya (rnisalnya lonjong atau bulat telur).
6. Bentuk daun yang dibedakan dalarn 3 kategori yaitu (1) elipfical-oblong- lanceolate, (2) oval oblong-linear oblong-obovolanceolate dan (3) obov-
ate-obovate oblong.
7. Bentuk lekukan ujung daun yang dibedakan dalam 5 kategori yaitu acurninate, cuspidate, caudate, cirrhose dan aristate.
8. Bentuk lekukan pangkal daun yang dibedakan dalarn 3 kategori yaitu obtuse, acute dan roundate.
9. Urat tulang daun yang dibedakan dalarn 2 kategori yaitu jelas dan tidak jetas.
11. Warna permukaan bawah daun yang dibedakan dalam 4 kategori yaitu hijau, hijau muda, coklat keemasan dan hijau kecoklatan.
12. Perrnukaan antar tulang daun yang dibedakan dalam 4 kategori yaitu rata, bergelombang, keriput dan berbingkul (bergelombang tidak teratur).
13. Lipatan daun yang dibedakan dalam 3 kategori yaitu rata (mendatar atau tidak berlipat), agak rneiipat dan melipat (seperti V)
Sudut cabang. Sudut cabang menyatakan sudut yang terbentuk antara
cabang primer dengan arah atas batang utarnanya. Sudut cabang diukur dengan busur derajat yang diletakkan mengikuti arah cabang dan diberi bandul (Norman dan Campbell. 1989). Skala yang ditunjukkan oleh benang bandul rnenyatakan skala sudut cabang (dalam derajat). Cabang yang bersudut lebih dari 60' dikategorikan sebagai cabang yang datar.
Tinggi tanaman. Tinggi tanarnan merupakan tinggi tunas dari titik mata ternpel pada bibit klonal atau dari pangkal tunas yang tumbuh setelah semai
dipancung pada semai yang dianggap sebagai perlakuan kontrol.
Analisis Statistika
Data karakter morfologi berupa data ordinal dan nominal (frekuensi)
dianalisis dengan analisis komponen utama yang dapat menjelaskan keragaman sekurang-kurangnya 60% (Suketi, 1994 dan Sahardi, 2000). Analisis ini
dimaksudkan untuk melihat besarnya kontribusi karakter tertentu terhadap keragaman antar jenis. Kategori bentuk dan sudut cabang disajikan sebagai tabel frekuensi. Data tinggi tanaman dan sudut cabang dianalisis dengan uji-t
pada taraf uji 5% dan 1%.
Hasil d a n Pembahasan
Sub-set Penelitian-la: Karakterisasi Morfologi Semai Durian
Hasil analisis kornponen utarna terhadap 13 karakter morfologi rnenunjuk- kan tujuh kornponen utarna pertarna rnenjelaskan 64.1% dari keragaman total (Tabel 1). Masing-masing berturut-turut ketujuh kornponen utama tersebut menyumbangkan keragaman sebesar 12.7, 9.9, 9.4, 8.6,. 8.3, 7.8 dan 7.4%. Dari karakterisasi tersebut diperoleh bahwa untuk menjelaskan tingkat keragam- an sedikitnya 60% diperlukan tujuh kornponen utama. Hasil analisis ini menunjukkan bahwa karakter morfologi antar jenis sangat beragam. Sebagai
[image:39.535.43.479.0.605.2]Tabel 1 : Proporsi (%) komponen karakter morfologi semai durian lokal Bogor Pagar. Mentega. Silodong dan KocIak
diperlukan dua komponen utama (Suketi. 1994). Keragaman yang tinggi ini terjadi karena populasi mengalami segregasi dan fakta ini menegaskan bahwa durian termasuk tanaman yang menyerbuk silang dan self-sterile (Subhadra- bandhu et
a / . ,
1992).Mengingat besarnya keragaman ini analisis lebih jauh dengan diagram pencar maupun dendrograrn tidak dilakukan. Sangatlah sulit menginterpretasi diagram pencar maupun dendrogram yang diderivasi dari lebih 2-3 komponen
utama. Pengelompokan menurut jenis akan sulit diperoleh karena suatu karakter tidak dimiliki secara khas oleh suatu jenis. Namun demikian di antara 13 karakter morfologi tersebut bentuk tajuk dan sudut cabang berkontribusi cukup besar dalam menjelaskan keragaman semai antar jenis durian. Perbandingan antar dua populasi kategori bentuk (kerucut dan kolumnar) mencapai 17.5 : 82.5 (Mentega) dan 18.1 : 81.9 (Koclak). Perbandingan dua populasi kategori sudut
cabang (< 60° dan r 60') nlencapai 14.9 : 85.1 (Pagar). Namun perbandingan
50:50 dijumpai populasi lainnya pada dua kategori tadi (Tabel 2).
Walaupun sumbangan dua karakter ini cukup besar untuk rnenjelaskan
keragaman jenis tertentu, namun mengingat populasi mengalami segregasi maka kedua karakter ini tetap disimpulkan tidak dapat menjelaskan keragaman
semai. Kepentingan dua karakter morfologi ini untuk karakter batang bawah Warna perrnukaan atas daun Warns bawah daun Permukaan antar tulanq daun Sudut permukaan daun
Angka yang dicetak tebal menyatakan lima karakter pertarna yang rnenerangkan keragaman terbesar pada kornponen utarna I-VII.
[image:40.537.44.479.33.607.2]Tabel 2 : Frekuensi (%) sema~ berdasarkan bentuk tajuk dan sudut cabang.
yang diidentifikasi memerlukan pengkajian lebih lanjut dengan metode-metode perbanyakan vegetatif.
Karakter morfologi lainnya seperti bentuk batang, lekukan ujung daun dan
morfologi daun lainnya, kecuali warna permukaan bawah daun, walaupun pen- ting untuk mencandra tanaman tampak kurang kontribusinya. Warna permuka- an bawah daun cukup besar kontribusinya dalam menjelaskan keragaman antar jenis.
Jenis durian Pagar Mente~a Silodonq Koclak
Sub-set Penelitian-$6: Pengaruh Batang Bawah terhadap Karakter Morfologi Bibit Tempelan
Pada percobaan penempelan batang bawah dengan batang atas yang
sejenis tampak bahwa rata-rata sudut cabang tidak berbeda, demikian pula frekuensi kategori batang atas bibit tempelan dengan batang bawah yang sejenis
relatif tidak berubah (Tabel 3). Dari Tabel yang sama tampak bahwa di antara ketiga jenis durian tersebut Pagar merupakan jenis durian dengan sudut cabang relatif besar dan populasi Silodong lebih tinggi dibandingkan Pagar maupun Mentega. Narnun demikian perbandingan tinggi populasi bibit klonal terhadap tinggi semai dalam jenis yang sama tidak secara langsung mencerminkan efek kate suatu batang bawah, karena (1) tanaman berasal dari populasi semai yang mengalami segregasi. (2) tanaman masih dalam masa juvenil (3) faktor penyam-
bungan karena regenerasi atau pemulihan pembuluh belum maksimal dan (4) faktor-faktor kompatibilitas batang atas dan batang bawah (Hartmann et a/.,
1990) yang belum diketahui.
Sama halnya dengan percobaan penernpelan batang atas dan batang bawah sejenis, dari hasil percobaan penempelan yang berbeda jenis (Tabel 4) juga belum dapat disimpulkan bahwa batang bawah tertentu mempengaruhi
perilaku batang atas. Namun demikian sesuai dengan tujuan penelitian ini untuk mengeksplorasi maka setidak-tidaknya telah dapat diketahui bahwa batang atas Hepe rnenunjukkan keberhasilan tempelan yang lebih baik dibandingkan Keterangan: Angka dalam kurung rnenunjukkan jumlah individu (n).
Bentuk taiuk Kerucut
41.8 (59) 17.5 (34) 50.0 (52) 18.3 (26)
Sudut cabanq Kolumnar
58.2 (82) 82.5 (160)
50.0 (52) 81.9 (118)
4 60°
14.9 (21) 36.6 (71) 42.3 (44) 38.5 (55)
[image:41.533.41.470.42.600.2]Tabel 3 :Pengaruh penempelan terhadap sudut cabang (di lapang urnur 2 tahun) dan tinggi tanarnan (di pernbibitan urnur 12 bulan) batang atas durian lokal Pagar, Mentega dan Silodong
Catatan: Anaka rata-rata vano diikuti denaan berturut-turut denaan huruf kecil a dan b atau huruf A dsn B dalam sel yang sama mknunjukkan tidak berbkda nyata dengan uji-t pada taraf uji 5% atau 1%. Angka rata-rata pada semai Pagar (P). Mentega (M) dan Silodong (S) yang diikuti dengan huruf kecil p dan q atau P. Q dan R berturut-turut tidak berbeda nyata dengan uji-t pada taraf uji 5% atau 1%. SD = standar deviasi dan n = individu.
Tabel 4 : Pengaruh batang bawah terhadap frekuensi bentuk tajuk, sudut cabang dan keberhasilan penempelan bibit klonal Monthong dan Hepe urnur 6 bulan.
Monthong. Hal ini terjadi diperkirakan karena batang atas Hepe berasal dari pohon induk yang berurnur cukup tua (42 tahun).
Dari penelitian yang bersifat eksploratif ini diperoleh material tanarnan
[image:42.535.43.478.22.617.2]Kesimpulan dan Saran
Kesimpulan
1. Keragaman dalam jenis sangat besar dan populasi batang bawah berse- gregasi, namun demikian dapat djsimpulkan bahwa di antara 13 karakter morfologi yang berperan dalam keragaman tersebut, bentuk tajuk dan sudut cabang merupakan karakter yang berkontribusi cukup besar.
2. Berdasarkan hasil percobaan penempelan, penggunaan semai sebagai identified rootstock untuk memperoleh bibit klonal dengan bentuk tajuk dan
sudut cabang tertentu belum dapat direkomendasikan, karena masih memer- lukan penelitian lebih lanjut dengan pendekatan perbanyakan vegetatif
.
3. Hubungan bentuk tajuk dengan perilaku plagiotropi, yang direfleksikan olehsudut cabang, tidak dapat diketahui karena populasi bersegregasi dan obser-
vasi karakter morfologi yang bersifat kategorisasi terhadap suatu populasi.
Saran
3 . Perlu dilakukan penelitian lebih lanjut untuk rnernpelajari pengaruh batang
bawah terhadap bentuk tajuk dan sudut cabang atau perilaku plagiotropi dengan teknik "layerage" (Hartmann ef a/.. 1990) terhadap tunas-tunas baru
identified rootstock yang telah diketahui secara individual memperbaiki
bentuk tajuk dan sudut cabang bibit klonal.
2. Untuk memperbaiki rnetode pengamatan bentuk tajuk, bentuk tajuk perlu
dinyatakan sebagai model-model topofisik tajuk.
Dafhr Pustaka
Bowman. K. 13. 1997. Inheritance of procumbent habit from 'cipo' sweet orange in crosses with 'Clementine' mandarin. J. Amer. Soc.. Hort. Sci. 122(3):696-701.
Cline, M. G. 1997. Concepts and terminology of apical dominance. Amer. J. Bot. 84~1064-1069.
Cook, N. G., E. Rabe and G. Jacobs. 1999. Early expression of apical control regulates length and crotch angle of sylleptic shoots in peach and nectarine. HortScience 34(4):604-606.
Departemen Pertanian. 1995. Deskripsi Durian Varietas Unggul Nasional. Tidak dipublikasi.
Hartmann, H. T., D. E. Kester and F. T. Davies, Jr. 1990. Plant Propagation: Principles and Practices. Fifth Ed. Prentice Hall Career and Technology, New Jersey, 647 p.
IBPGR. 1980. Tropical Fruit Descriptors. International Board for Plant Genetic Resources. South-East Asia Regional Committee. 11 p.
Norman, J . M. and G. S. Campbell. 1989. Canopy structure, p:301-325.
In
R. W. Pearcy eta/.
(eds). Plant Physiological Ecology. Chapman and Hall, London, 439 p.Poerwanto, R. 1996. Laporan Kunjungan ke Kebun Buah-buahan di Thailand. Laporan Perjalanan Dinas tanggal 4-12 Mei 1996. Fakultas Pertanian IPB. Bogor (tidak dipublikasi).
Ryugo, K. 1988. Fruit Culture: Its Science and Art. John Wiley and Sons. New York, 344 p.
Sahardi. 2000. Studi Karakteristik Anatorni dan Morfologi serta Pewarisan Sifat Toleransi terhadap Naungan pada Padi Gogo
( O v a sativa
L.).Disertasi Program Pascasarjana IPB, Bogor, 96 p.
Subhadrabandhu, S., J. M. P. Scheemann and E. W. M. Verheij. 1992. Durio
zibethinus
Murray, p:147-161.In
E. W. M. Verheij and R. E. Coronel(Eds). Plant Resource of South East Asia No 2: Edible Fruits and Nuts. PROSEA. Bogor.
Suketi, K. 1994. Studi Karakterisasi Bibit Klonal Durian Berdasarkan Morfologi Daun dan Pola Pita Isozirn. Tesis Program Pascasarjana IPB, Bogor, 82 p.
Tjitrosoepomo. G. 2000. Morfologi Tumbuhan. Gajah Mada University Press. Yogyakarta. 266 p.
SET PENELITIAN-2: PERlLAKU PERCABANGAN,
BENTUK TAJUK DAN ASPEK MORFOMETRI CABANG
A b s t r a k
Untuk mempelajari hubungan perilaku percabangan (dirnorfisme dan kendali apikal), bentuk tajuk dan aspek morfometri cabang . durian fase bibit dan dewasa telah dilakukan dua unit percobaan paralel dan dua unit 0 b s e ~ a s i pada dua durian Klon Mothong dan Matahari. Hasil penelitian menunjukkan adanya hubungan paralel (1) antara model (bentuk atau "frame") tajuk fase bibit dan dewasa. (2) antara model alami tajuk dengan kendali apikal dan (3) antara model tajuk dengan dimorfisme cabang pada durian bertajuk kerucut (Klon Matahari) dan kolumnar (Klon Monthong).
Berdasarkan simulasi dengan pemangkasan dan training diperoleh model tajuk dengan perubahan gradien tajuk dua fase (pada Klon Monthong) hingga tiga fase (pada Klon Matahari). Gradien tajuk Matahari lebih besar dibandingkan gradien tajuk Monthong pada fase bibit maupun dewasa dan pada perlakuan kontrol maupun pernangkasan dan training. Fakta-fakta ini rnenunjukkan bahwa (1) cabang basal kedua jenis durian mernpunyai kemarnpuan tumbuh yang berbeda yang dikendalikan oleh suatu rnekanisme kendali apikal. (2) faktor ontogeni maupun perlakuan tidak rnempengaruhi kendali apikal durian dan (3) kendali apikat berhubungan paralel dengan bentuk tajuk.
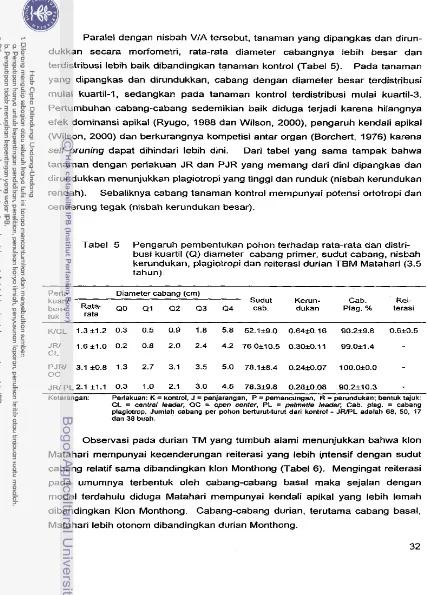
Kendali apikal durian Matahari lebih lemah dibandingkan Monthong, atau dengan kata lain tingkat otonomi cabang basal durian Matahari lebih besar, sehingga tajuknya berbentuk kerucut, dibandingkan Monthong (bertajuk kolum- nar). Dari observasi diperoleh bahwa pada kondisi alarniah, tanarnan dengan kendali apikal lernah (Matahari) cenderung rnernbentuk reiterasi yang lebih banyak atau cabang plagiotrop yang lebih sedikit dibandingkan yang rnempunyai kendali apikal kuat (Monthong). Pernangkasan dan training secara umurn memperbaiki aspek-aspek morfornetri percabangan, diameter dan nisbah VIA.
Pendahuluan
Latar Belakang
Perilaku percabangan telah dirnanfaatkan sebagai dasar rnemanipulasi bentuk, struktur atau "frame" pohon. Dirnorfisme percabangan, suatu perilaku
yang spesifik kelompok tanaman model Roux, dijadikan dasar rnernanipulasi pohon kopi yang mempunyai persamaan model dengan durian (Halle et a/.,
1978) agar cabang plagiotropnya menyangga bunga dan buah sebanyak mungkin, sehingga produktivitas meningkat. Cabang plagiotrop diarahkan sernakin mendatar atau bersudut cabang besar (Ryugo, 7988) dan runduk (Bowman, 1997). Cabang-cabang ortotrop yang potensial mengalami reiterasi (Hafle et a/., 1978) telah dijarangkan (thinned) sehingga pohon berstruktur sederhana, berbatang tunggal. Dengan demikian kompetisi antar organ diha-
struktur demikian produktivitas kopi telah dapat dipacu mencapai produktivitas potensialnya yaitu 50 tonlhattahun. Produktivitas durian sebagai energy rich
fruit
diperkirakan akan dapat ditingkatkan dari 6.2-16.5 tontha menjadi 2 4 kali lipat yaitu 25 ton/ha (Verheij dan Coronel. 1992).
Polimorfisme percabangan, suatu perilaku cabang yang spesifik kelom- pok tanaman dengan arsitektur Rauh, telah dimanfaatkan sebagai dasar mema- nipulasi bentuk pohon peach (Scorza
et
al., 1986 dan Bassiet
a/., 1994), ape1 dan famili Rosaceae lainnya (Ryugo. 1988). Manipulasi selain diarahkan untuk menangkap dan mengintersepsi cahaya matahari secara maksimum di daerahyang jauh dari katulistiwa (Johnson dan Lakso. 1991), juga diarahkan untuk meningkatkan
spur
yang merupakan cabang yang produktif (Scorza et al., 1986 dan Bassi eta / . .
1994).Secara rnorfometri, struktur pohon yang sederhana sebagaimana dimak-
sudkan di atas direfleksikan sebagai perbandingan volume cabang dan batang dengan luas permukaannya atau nisbah volumelarea (Halle
et a / . ,
1978) yang disingkat VIA. Nisbah VIA ini menurut Halleet
a/. (1978) dapat menggambarkan strategi perturnbuhan dan perkembangan tanaman. Pohon yang mengalami pauperasi (strategi-r) atau reiterasi sebagaimana kerangka penelitian ini (GambarI ) akan bercabang banyak, tetapi masing-masing cabang berukuran kecil sehing- ga luas permukaan cabangnya meningkat lebih besar dibandingkan volumenya. Nisbah VIA ini menurut Halle et a / . (1978) juga merefleksikan perbandingan distribusi energi internal untuk pertumbuhan dan perkembangan dengan energi
eksternal untuk adaptasi terhadap lingkungan. Nisbah VIA yang besar mere- fleksikan energi investasi internal yang besar. Sebaliknya nisbah VIA yang kecil merefleksikan energi yang digunakan untuk adaptasi ke lingkungan atau entropi semakin besar.
Jika pada tanarnan herba bentuk dewasa
(mature)
tanarnan dikendalikan oleh dominansi apikal, pada tanaman berbentuk pohon bentuk tersebut dikendalikan selain oleh dominansi apikal juga oleh kendali apikal atau apicalcontrol
(Cline, 1997 dan Wilson, 2000). Perilaku kendali apikal rnempengaruhi(thinned) maka cabang di bawah (basal) tidak lebih panjang, atau hanya,sedikit lebih panjang, dibandingkan cabang atasnya. Akibatnya pohon akan berbentuk kerucut yang tajam, kolumnar atau hirarkhi. Sebaliknya di bawah kendali apikal yang lemah cabang basal akan turnbuh dan berkembang lebih otonom, walaupun cabang di atasnya dijarangkan, sehingga pohon berbentuk kerucut tumpul.
Bahkan cabang basal dapat menggantikan fungsi apexi sehingga tanarnan bersifat poliarkhi (lawan hirarkhi), dapat memiliki lebih dari satu axis.
Penelitian Cook et a/. (1999) pada peach menunjukkan bahwa kendali apikal bewariasi antar kultivar. Kultivar peach Zaigina dan Mayglo diketahui merupakan jenis yang pa1ir.g poliarkhi (lawan hirarkhi), kendali apikalnya paling lemah, dibandingkan kultivar Fiesta Red atau Oom Sarel. Sebagaimana diurai-
kan terdahulu arsitektur pohon peach dan tanaman famili Rosaceae tergolong sebagai model Rauh, sedangkan arsitektur pohon durian adalah Roux, mernpu-
nyai persamaan dan perbedaan-perbedaan tertentu dibandingkan model Rauh. Dari hasil studi tersebut (Cook eta!.. 1999) dan pohon natal (Cline. 1997). diperoleh bahwa perilaku kendali apikal cabang diketahui dapat digunakan untuk meramalkan kesesuaian pola training pohon peach dan seleksi pohon natal pada fase dini. Berdasarkan analogi ini dan paralel dengan tujuan pembentukan pohon, maka studi perilaku percabangan pada durian perlu dilakukan.
Tujuan Penelitian
Untuk dapat mernberikan penjelasan dasar-dasar botanis pengendalian arsitektur pohon sebagaimana diuraikan tersebut dilakukan tiga sub-set pene[iti- an sebagai satu set penelitian yaitu Set Penelitian-2. Ketiga sub-set penelitian tersebut bertujuan sebagai berikut:
1. Mengkuantifikasi bentuk ("frame) tajuk dalam model tajuk.
2. Mempelajari hubungan pemangkasan dan training dengan kendali apikal dan model tajuk.
3. Mempelajari aspek rnorfornetri dan dimorfisrne percabangan
Bahan dan Metode
Sub-set Penelitian-2a: Model Tajuk Bibit Klonal
Sub-set Penelitian-2a ini merupakan suatu observasi terhadap bibit
klonal (tempelan) durian Klon Monthong dan Matahari masing-masing sebanyak 10 tanaman. Kedua jenis durian ini dipilih karena secara visual bentuk tajuk
Matahari berbentuk kerucut. Kedua populasi durian tersebut ditanam alami. tanpa perlakuan pembentukan tajuk.
Di samping itu kedua klon ini dipilih karena sama jenisnya dengan klon TBM yang memperoleh perlakuan pemangkasan dan training pada Sub-set Penelitian-2b maupun observasi durian TM pada Sub-set Penelitian-2c. Dengan
demikian ketiga populasi pada ketiga sub-set penelitian paralel ini dapat dianggap mengatasi faktor ontogeni.
Eibit tanarnan diambil dari Kebun Agropromo Baranangsiang, Jurusan Budidaya Pertanian, Fakultas Pertanian IPB, Bogor. Bibit berumur sekitar 12 bulan sejak penempelan dan ditanam dalam polibag.
Sub-set Penelitian-2b: Hubungan Pemangkasan dan Training dengan Kendali Apikal dan Bentuk Tanaman Belum Menghasilkan (TBM)
Sub-set Penelitian-2b ini terdiri atas dua percobaan paralel masing- rnasing dengan Klon Matahari dan Klon Monthong. Kedua klon durian ini ber- asal dari bibit tempelan dengan batang bawah tidak diketahui (unidentified rootstock). Percobaan dengan Klon Matahari dilakukan di Kebun Percobaan
Fakultas Pertanian IPB Cikabayan (KP Cikabayan), Bogor (* 250 m dpl). sedangkan dengan Klon Monthong di KP Sukamantri Bogor (* 600 rn dpl) yang ditanam tahun 1995. Pada saat akhir percobaan (tahun 1999) tanaman berumur 3,5 tahun dan belum menghasilkan (TBM).
Percobaan dengan Klon Matahari (selanjutnya disebut Percobaan Klon Matahari) merupakan percobaan satu faktor dengan rancangan acak lengkap. Jumlah ulangan lima. Perlakuan merupakan pernangkasan bentuk dan training pohon. Perlakuan mulai diterapkan pada saat tanaman berumur 2 tahun dan
diakhiri 19 bulan kemudian. Perlakuan dan bentuk training adalah sebagai berikut:
I. Kontrol (K = kontrol), central leader (CL)
Tanaman dibiarkan tumbuh alami. Perlakuan ini menghasilkan tanaman dengan panutan (batang) utama (central leader atau CL). Pohon tidak dipangkas {tidak dipancung dan tidak dijarangkan) dan tidak dirundukkan. Bentuk tajuk secara umum kerucut.
2. Penjarangan yang diikuti perundukan cabang (JR, J
=
jarang dan R = runduk), central leader. Cabang primer tanarnan dijarangkan (thinned) dan dirun- dukkan (bended). Perlakuan ini menghasilkan tanaman dengan panutan