INTRODUKSI DAN EKSPRESI GEN FRAGMEN ANTIBODI
UNTAI TUNGGAL (scFv) ANTI-EGFRvIII
PADA
Pichia pastoris
AI HERTATI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa Tesis yang berjudul Introduksi dan Ekspresi Gen Fragmen Antibodi Untai Tunggal (scFv) Anti-EGFRvIII pada
Pichia pastoris adalah karya saya bersama komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir Tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor dan LIPI.
Bogor, April 2013
RINGKASAN
AI HERTATI. Introduksi dan Ekspresi Gen Fragmen Antibodi Untai Tunggal (scFv) Anti-EGFRvIII pada Pichia pastoris. Dibimbing oleh ANJA
MERYANDINI DAN ASRUL MUHAMAD FUAD
EGFRvIII merupakan salah satu varian mutan Epidermal Growth Factor Receptor (EGFR) yang mengalami mutasi pada bagian ekstraselularnya.
EGFRvIII hanya ditemukan pada sel kanker dan tidak pada sel normal, sehingga merupakan target terapi kanker yang sangat menarik. Molekul antibodi utuh telah digunakan sebagai agen terapi kanker. Akan tetapi penggunaannya dalam
konstruksi obat dengan sistem penghantaran terarah memiliki kelemahan karena ukurannya yang relatif besar (150 kDa). Hal ini disebabkan karena ukuran yang besar dapat mengurangi kemampuan untuk penetrasi ke dalam sel tumor. Permasalahan tersebut diatasi dengan membuat suatu fragmen antibodi untai tunggal (scFv) anti-EGFRvIII yang memiliki ukuran 25 kDa. Fragmen antibodi terdiri dari bagian untai berat (VH) dan untai ringan (VL) yang dihubungkan oleh suatu peptida yang disebut linker (G4S)3. Tujuan dari penelitian ini adalah untuk mengkonstruksi scFv, mengintroduksi dan mengekspresikannya ke dalam Pichia pastoris. Fragmen antibodi dibuat dalam 2 versi, yaitu versi 1 (VH-L-VL) dan versi 2 (VL-L-VH), yang dimaksudkan untuk mempelajari sifat afinitasnya terhadap antigen. Fragmen antibodi versi 1 dirancang dan dibuat secara sintetik dan kemudian diklon ke dalam vektor pPICZαA dan ditransformasikan ke dalam
E. coli. Plasmid rekombinan (pPICZαA-scFv-1) dianalisis dengan menggunakan PCR, enzim restriksi dan sekuensing. Plasmid rekombinan dengan urutan DNA yang tepat kemudian difusikan dengan suatu gen penanda yaitu Green
Fluorescent Protein (GFP) lalu ditransformasikan ke dalam P. pastoris
SMD1168H. Nilai efisiensi transformasi yang diperoleh adalah 1,2x 104 cfu/µg DNA plasmid. Hasil analisis dengan teknik SDS-PAGE dan Slot blot terhadap media kultur bebas sel menunjukkan bahwa fusi scFv-GFP telah berhasil
diekspresikan secara ekstraselular dengan ukuran 60 kDa. Fragmen antibodi versi 2 dibuat dengan menggunakan teknik overlap extension PCR lalu diklon ke dalam vektor pTZ57r/t dan ditransformasikan ke dalam E. coli.
SUMMARY
AI HERTATI. Introduction and Expression of Single Chain Fragment Variable (scFv) Anti-EGFRvIII Gene Into Pichia pastoris. Supervised by ANJA
MERYANDINI and ASRUL MUHAMAD FUAD
EGFRvIII is one of EGFR (epidermal growth factor receptor) mutant variant that has deletion in its extracellular domain. EGFRvIII is one of important targets for cancer therapy since its only found in cancer cell. EGFRvIII antibody binds to truncated extracellular domain of EGFRvIII. Intact antibody has been used as cancer therapeutic agent. The size of intact antibody (150 kDa) could be an obstacle in targeted drug delivery system. it reduces ability to penetrate the cancer cell. It is well known that bigger molecule has bigger immunogenicity. These problems might be solved by using smaller molecule such as single chain fragment variable (scFv) that has a smaller size (25 kDa). The scFv consist of variable light (VL) and variable heavy (VH) domains joined by a flexible peptide linker (G4S)3. The aimed of this studies were to construct scFv, transforms and express this gene into P. pastoris. Two version of scFv genes was constructed in this study. First version of scFv (VH-L-VL) was designed and made synthetically. While the second version (VL-L-VH) was made by overlap extension PCR
technique. One of those constructs (scFv-1) was cloned into pPICZαA and
transformed into E. coli. A recombinant plasmid (pPICZαA-scFv-1) was analyzed using PCR, restriction enzyme and DNA sequencing. Some clones of scFv with proper DNA sequence and orientation have been obtained. EGFP gene was then fused with the recombinant plasmid (pPICZα-scFv-1) and transformed into P. pastoris. Transformation of P. pastoris was performed using electroporation method. The transformation efficiency observed was 1,2x104 cfu/µg DNA plasmid. SDS-PAGE and Slot Blot analysis of free-cell culture medium showed the presence of scFv-GFP protein fusion having a size of 60 kDa and expressed extracellularly. scFv-2 that has structure VL-L-VH has been made by overlap extension PCR technique. It was cloned to pTZ57r/t and transformed into E. coli.
© Hak Cipta milik IPB, tahun 2013
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; pengutipan tersebut tidak merugikan kepentingan IPB & LIPI
INTRODUKSI DAN EKSPRESI GEN FRAGMEN ANTIBODI
UNTAI TUNGGAL (scFv) ANTI-EGFRvIII PADA
Pichia pastoris
AI HERTATI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Bioteknologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN
Judul Penelitian : Introduksi dan Ekspresi Gen Fragmen Antibodi Untai Tunggal (scFv) Anti-EGFRvIII pada Pichia pastoris
Nama : Ai Hertati
NIM : P051090141
Disetujui oleh
Komisi Pembimbing
Prof Dr Anja Meryandini, MS Dr Asrul Muhamad Fuad, MSi
Ketua Anggota
Diketahui oleh,
Ketua Program Studi Dekan Sekolah Pascasarjana
Bioteknologi
Prof Dr Ir Suharsono, DEA Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: Tanggal Lulus:
PRAKATA
Puji serta syukur penulis ucapkan kehadirat Alloh SWT karena berkat Rahmat dan Hidayah-Nya penulis dapat menyelesaikan penelitian dengan judul “Introduksi dan Ekspresi Gen Fragmen Antibodi Untai Tunggal (scFv) Anti-EGFRvIII pada Pichia pastoris”. Tesis ini merupakan hasil penelitian yang dilakukan di Laboratorium Protein, Vaksin dan Sistem Penghantaran Terarah, Pusat Penelitian Bioteknologi-LIPI.
Pada kesempatan ini, penulis mengucapkan terima kasih yang sebesar-besarnya kepada :
1. Prof. Dr. Anja Meryandini, MS dan Dr. Asrul Muhamad Fuad, M.Si selaku dosen pembimbing yang telah memberikan bimbingan dan arahan selama penelitian hingga penyusunan tesis ini.
2. Drs. Djadjat Tisnadjaja, M.Tech yang telah memberikan kesempatan dan dukungan sehingga penulis dapat mengikuti program pendidikan pascasarjana. 3. Teh Goli, Prety, Dian, bu Ami, Rithami, Maya dan Riyona atas bantuannya selama penelitian di Laboratorium Protein, Vaksin dan Sistem Penghantaran Terarah. Nurlaili, dr. Ela dan Herman dari Laboratorium Biopharmaceutical
atas pengertiannya selama penulis mengerjakan penelitian. 4. Teman-teman Program Studi BTK 2009
Ungkapan terima kasih juga disampaikan kepada Mamah, Apa, Mas dan Fikri tersayang atas segala doa, dukungan dan kasih sayangnya.
Penulis menyadari akan keterbatasan yang dimiliki, maka masukan berupa saran dan kritik guna penyempurnaan penulisan karya ilmiah ini sangat
diharapkan. Penulis berharap semoga karya ilmiah ini dapat bermanfaat bagi para pembaca dan terutama pengembangan ilmu penulis.
Bogor, April 2013
DAFTAR ISI
Sistem Ekspresi Pichia pastoris 4
Green Fluorescent Protein (GFP) 6
METODOLOGI 7
Waktu dan Tempat Penelitian 7
Bahan dan Metode 7
Prosedur Kerja 8
HASIL DAN PEMBAHASAN 13
Subkloning scFv-1 (VH-L-VL) ke dalam Vektor Ekspresi 13
Fusi Gen scFv-1 dengan gen GFP 18
Introduksi ke dalam Pichia pastoris 24
Seleksi Sel Pichia pastoris Transforman 27 Uji Ekspresi dan Analisis Ekspresi Protein Rekombinan Skala Kecil 31
Konstruksi scFv Versi 2 (VL-L-VH) 33
DAFTAR TABEL
Halaman
1 Primer yang digunakan dalam penelitian 8
2 Hasil transformasi plasmid rekombinan pPICZαA-scFv(1)-GFP pada E. coli TOP10F’
21
3 Hasil penentuan konsentrasi dan kemurnian plasmid rekombinan pPICZαA-scFv(1)-EGFP dengan menggunakan nanofotometer
26
4 Kondisi pada saat elektroporasi plasmid rekombinan pPICZα A-scFv(1)-EGFP dengan menggunakan alat elektroporator
26
5 Jumlah koloni hasil transformasi P. pastoris dengan plasmid pPICZαA-scFv(1)-EGFP
27
6 Persentase hasil seleksi P. pastoris transforman 28
DAFTAR GAMBAR
Halaman 1 Skema struktur protein pada EGFR wild-type dan EGFRvIII 3
2 Peta vektor pPICZαA 5
3 Skema struktur plasmid pAF_scFv-01 7
4 Target konstruksi scFv anti-EGFRvIII 7
5 Skema PCR untuk konstruksi scFv versi 2 12 6 Produk amplifikasi PCR plasmid pAF-scFv-1 dengan pasangan
primer VH101-F-NdeXho dan VL101-R-NdeXho
13
7 Hasil potong vektor pPICZαA dan gen scFv hasil PCR dengan enzim XhoI
14
8 Hasil transformasi pPICZαA-scFv-1 ke dalam E. coli TOP10F’
dengan metoda kejut panas
14
9 Posisi penempelan primer VH101-F dan VL101-R pada plasmid rekombinan pPICZαA_scFv-1
15
10 Hasil PCR koloni transforman pPICZαA-scFv-1 dengan menggunakan primer VH101-F dan VL101-R
16
11 Posisi penempelan primer AOX-F dan VH101-R pada plasmid rekombinan pPICZαA-scFv-1
16
12 Hasil PCR penentuan orientasi gen scFv dalam konstruk pPICZαA-scFv-1 dengan pasangan primer AOX-F dan VH101-R
17
13 Hasil pemotongan dengan menggunakan enzim restriksi XhoI terhadap plasmid rekombinan pPICZαA_scFv
18
14 Hasil amplifikasi (tahap pertama) dengan menggunakan pasangan primer EGFP-F dan EGFP-R
19
15 Hasil amplifikasi gen egfp dengan PCR tahap kedua
menggunakan pasangan primer G4S-ClaI dengan
R-ClaI
16 Hasil transformasi pPICZαA_scFv(1)_EGFP pada E. coli
TOP10F’ 20
17 Posisi penempelan primer pada proses PCR koloni dengan menggunakan primer AOX-F dan AOX-R
21
18 Hasil PCR koloni transforman dengan menggunakan pasangan primer AOX-F dan AOX-R
22
19 Posisi penempelan primer pada proses orientasi konstruk dengan menggunakan pasangan primer EGFP-F dan AOX-R
22
20 Hasil PCR penentuan orientasi gen egfp di dalam konstruk dengan menggunakan pasangan primer EGFP-F dan AOX-R
23
21 Hasil pemotongan plasmid rekombinan
pPICZαA_scFv(1)_EGFP dengan menggunakan enzim restriksi
ClaI
23
22 Hasil isolasi DNA plasmid pPICZαA_scFv(1)_EGFP 25 23 Hasil potong plasmid pPICZαA_scFv(1)_EGFP dengan
menggunakan enzim restriksi SacI
25
24 Hasil transformasi pPICZαA_scFv(1)_EGFP pada P. pastoris 27 25 Hasil seleksi transforman P. pastoris pada media YPD agar yang
mengandung zeocin 100 µg/ml
29
26 Hasil seleksi transforman P. pastoris pada media YPD agar yang mengandung zeocin 200 µg/ml
29
27 Hasil seleksi transforman P. pastoris pada media YPD agar yang mengandung zeocin 300 µg/ml
30
28 Hasil seleksi transforman P. pastoris pada media YPD agar yang mengandung zeocin 1000 µg/ml
30
29 Hasil analisis ekspresi protein rekombinan dengan teknik SDS-PAGE
31
30 Hasil analisis immunoblotting menggunakan teknik Slot Blot terhadap sampel protein beberapa galur P. pastoris transforman
32
31 Hasil pengamatan sel transforman P.pastoris yang mengandung gen penanda GFP yang berfluoresen hijau
32
32 Hasil amplifikasi PCR fragmen VH, VL dan Linker 34 33 Hasil amplifikasi PCR dan purifikasi fragmen scFv-2
menggunakan pasangan primer VL-F dan VH-R
35
34 Hasil transformasi plasmid rekombinan pTZ_scFv ke dalam E. coli DH5α
35
35 Hasil PCR koloni E. coli transforman pTZ_scFv-2 36 36 Hasil isolasi DNA plasmid pTZ_scFv-2 dari beberapa klon E.
coli transforman
36
37 Hasil PCR plasmid rekombinan pTZ_scFv dengan menggunakan pasangan primer VL101-F dan VH101-R
37
38 Hasil pemotongan plasmid rekombinan pTZ_scFv dengan menggunakan enzim XbaI
DAFTAR LAMPIRAN
Halaman 1 Komposisi dan cara pembuatan media, larutan dan buffer 43 2 Bahan yang digunakan dalam pembuatan gel poliakrilamid 45 3 Hasil sekuen DNA plasmid rekombinan pPICZαA_scFv-1 46 4 Hasil sekuen DNA plasmid rekombinan
pPICZαA_scFv(1)_EGFP
48
PENDAHULUAN
Latar Belakang
Terapi kanker terarah berbasis antibodi merupakan salah satu jenis terapi kanker generasi baru yang diharapkan dapat bersifat selektif sehingga dapat mengurangi efek samping terhadap sel normal. Faktor yang sangat penting dalam pembuatan terapi kanker terarah adalah pemilihan target terapi yang tepat. Salah satu target terapi yang telah diketahui mengalami overekspresi pada beberapa jenis sel kanker adalah epidermal growth factor receptor (EGFR) (Gupta et al. 2010 , Kuan et al. 2001).
Epidermal growth factor receptor (EGFR) merupakan suatu reseptor glikoprotein yang memiliki berat molekul sebesar 170 kDa. EGFR adalah anggota famili reseptor tirosin kinase yang memiliki peran penting dalam pertumbuhan dan proliferasi sel (Pedersen et al. 2001). Fakta bahwa EGFR ditemukan pada sel normal mengharuskan para peneliti dalam bidang onkologi mencari alternatif antigen lain yang lebih spesifik pada sel kanker. EGFRvIII merupakan salah satu varian mutan EGFR yang diketahui mengalami overekspresi pada beberapa jenis sel kanker seperti kanker paru-paru (16%), kanker payudara (78%) dan kanker ovarium (73%) (Pedersen et al. 2001). EGFRvIII merupakan target terapi yang sangat ideal karena hasil mutasi delesi ekson 2-7 menghasilkan suatu residu asam amino glisin dan membentuk sekuen peptida unik yang menjadikannya sebagai antigen spesifik. Selain itu ekspresi EGFR vIII tidak ditemukan pada sel normal (Pedersen et al. 2001, Gupta et al. 2010).
Antibodi yang digunakan dalam terapi kanker terarah dapat berupa molekul antibodi utuh atau berupa fragmen antibodi. Kedua molekul ini berperan sebagai ligan untuk menghantarkan agen sitotoksik pada sel target yang dituju. Terdapat beberapa hambatan apabila digunakan molekul antibodi utuh sebagai ligan. Hambatan ini terutama disebabkan karena ukuran molekul antibodi yang relatif besar (150 kDa) dapat mengurangi kemampuannya untuk penetrasi melewati membran sel tumor. Selain itu semakin besar ligan yang digunakan semakin besar pula imunogenisitas dari ligan tersebut. Oleh karena itu dicari ligan lain yang berukuran lebih kecil seperti fragmen antibodi untai tunggal (scFv). ScFv terdiri atas domain variable untai berat (VH) dan domain variable untai ringan (VL) yang dihubungkan oleh suatu polipeptida penghubung (linker). ScFv memiliki ukuran yang lebih kecil (25 kDa) sehingga memiliki kemampuan penetrasi membran sel tumor yang lebih baik dan imunogenisitas yang lebih rendah (Ahmad et al. 2011, Jafari et al. 2011)
ScFv telah diekspresikan pada beberapa sel inang mulai dari sel prokariot (E. coli) hingga sel mamalia (Chinese Hamster Ovary). Ekspresi scFv pada E. coli
memiliki beberapa kelemahan seperti scFv yang dihasilkan tidak mengalami proses modifikasi pasca translasi sehingga terkadang menghasilkan scFv yang tidak fungsional. Selain itu ekspresi scFv pada E. coli juga dapat menghasilkan
insoluble inclusion bodies yang memerlukan beberapa langkah tambahan agar menghasilkan scFv yang aktif (Naumann et al.2011, Peterson et al. 2006)
Sistem ekspresi khamir dapat menggabungkan keuntungan dari kedua sistem yang dijelaskan di awal. Sistem ekspresi khamir memiliki kemudahan proses seperti E. coli dan memungkinkan terjadinya proses modifikasi pasca translasi sehingga dihasilkan molekul scFv yang aktif. Ekspresi pada khamir dapat terjadi secara intraselular maupun ekstraselular. Pichia pastoris merupakan salah satu khamir yang telah banyak digunakan sebagai sel inang untuk ekspresi beberapa jenis protein rekombinan termasuk scFv (Daly & Hearn 2005). Beberapa keunggulan P. pastoris sebagai sel inang diantaranya adalah mampu tumbuh dengan densitas sel yang tinggi pada media yang sederhana dan memiliki promotor AOX1 yang sangat efisien sehingga mampu memproduksi protein rekombinan dalam jumlah yang cukup besar (Cupit et al. 1999)
Green fluorescent protein (GFP) merupakan suatu protein yang dapat berperan sebagai penanda bagi beberapa proses biologis seperti ekspresi suatu protein atau untuk menentukan lokasi dari protein target di dalam sel atau organisme tertentu. Fusi scFv dengan GFP telah diketahui memiliki beberapa manfaat seperti dalam proses analisis tumor, penemuan obat baru serta mempelajari proses internalisasi dan farmakodinamika terapi gen (Naumann et al.
2011).
Tujuan Penelitian
Penelitian ini bertujuan untuk membuat fragmen antibodi untai tunggal (scFv) yang digunakan sebagai ligan untuk membawa agen sitotoksik.
Manfaat Penelitian
Fragmen antibodi (scFv) anti-EGFRvIII diharapkan dapat menjadi suatu ligan spesifik yang berperan membawa agen antikanker yang dapat bermanfaat untuk pengembangan terapi kanker terarah.
TINJAUAN PUSTAKA
EGFR dan EGFRvIII
Epidermal growth factor (EGF) yang ditemukan pada tahun 1950-an oleh Levi-Montalcini dan Stanley Cohen adalah faktor pertumbuhan yang pertama kali ditemukan. Pada tahun 1970-an protein membran plasma yang berikatan dengan EGF ditemukan dan diberi nama EGFR. EGFR merupakan suatu glikoprotein dengan berat molekul 170 kDa dan termasuk ke dalam famili reseptor tirosin kinase (Carpenter & Cohen 1979).
Karena peranannya sebagai mediator dalam proses pertumbuhan dan proliferasi sel, EGFR diasosiasikan dengan sebagian besar kasus malignancy pada manusia termasuk diantaranya kanker payudara, kanker paru-paru, kanker otak, kanker prostat dan kanker mulut (Harari 2004).
Mekanisme yang menyebabkan EGFR menjadi onkogenik bervariasi yaitu : proses autocrine growth factor loops, amplifikasi dan overekspresi EGFR, serta mutasi yang menyebabkan perubahan aktivasi reseptor yang semula bersifat terinduksi ligan menjadi bersifat konstitutif (Harari 2004).
Mutan EGFR ini biasa ditemukan pada beberapa kasus tumor manusia dan mutasi yang paling sering terjadi adalah delesi pada daerah ekstraselular. Delesi ini mengakibatkan EGFR bersifat konstitutif. Salah satu varian mutan EGFR yang mengalami mutasi adalah EGFRvIII (Kuan et al. 2001, Mendelsohn & Baselga 2003).
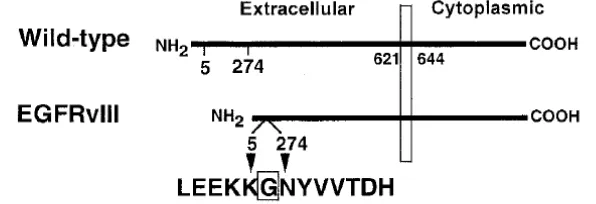
EGFRvIII merupakan suatu EGFR yang telah mengalami mutasi pada bagian ekstraselularnya. Bentuk mutasi yang terjadi adalah delesi pada 801 pasang basa yang terdapat pada ekson 2-7 pada mRNA, menghasilkan kehilangan asam amino nomor 6-273 pada bagian ekstraselular. EGFRvIII memiliki berat molekul sebesar 145 kDa. EGFRvIII tetap memiliki kemapuan untuk masuk ke dalam membran sel karena tidak ada perubahan pada signal peptida. Perbedaan struktur EGFR dengan EGFRvIII dapat lihat pada Gambar 1 (Kuan et al. 2001).
Gambar 1 Skema Struktur Protein pada EGFR Wild-type dan EGFRvIII (Kuan et al. 2001)
Sistem Ekspresi Pichia pastoris
Sistem ekspresi prokariot biasanya digunakan untuk memproduksi protein heterolog rekombinan dari cDNA eukariot. Akan tetapi, dari hasil beberapa penelitian diketahui bahwa protein yang berasal dari sel eukariot membutuhkan proses modifikasi pasca translasi agar menjadi suatu bentuk protein yang stabil dan aktif secara biologis. Sistem ekspresi prokariot tidak mempunyai perangkat untuk melakukan proses modifikasi pasca translasi ini. Kelemahan sistem ekspresi prokariot yang lain adalah senyawa toksin dari bakteri dan yang bersifat pirogen dapat mengkontaminasi produk protein rekombinan yang dihasilkan. Untuk mengatasi masalah tersebut maka dikembangkan sistem ekspresi protein eukariot, yaitu ragi, serangga, atau sel mamalia sehingga protein yang dihasilkan bebas dari kontaminasi, stabil dan memiliki aktivitas biologis (Daly & Hearn 2005).
Ekspresi protein dalam S. cerevisiae telah berhasil digunakan untuk ekspresi berbagai protein dari sumber yang berbeda. Pada beberapa hasil penelitian hasil ekspresinya rendah. Pada penelitian lain disebutkan bahwa protein rekombinan yang diproduksi pada sistem ekspresi S. cerevisiae ini mengalami proses hiperglikosilasi dengan jumlah residu manosa 50-150 residu pada rantai samping N-linked oligosakarida. Inti oligosakarida S. cerevisiae memiliki ikatan 1,3 glikan pada ujungnya yang diduga menyebabkan efek hiper antigenik sehingga protein yang dihasilkan tidak dapat digunakan sebagai agen terapetik. Selain itu, pada S. cerevisiae apabila densitas sel tinggi maka akan memproduksi etanol yang bersifat toksik bagi sel. Oleh karena itu, jumlah protein yang disekresikan harus lebih rendah (Cregg et al. 1989, Daly & Hearn 2005).
Peneliti lalu mencari sistem ekspresi eukariot lain yang dapat berperan sebagai sel inang yang efektif untuk memproduksi protein rekombinan tanpa kendala yang telah diuraikan dibagian sebelumnya. P. pastoris sebagai sistem ekspresi eukariot memiliki kemampuan untuk memproduksi protein, pelipatan protein dan modifikasi pasca translasi. Namun disisi lain P. pastoris mudah untuk dimanipulasi seperti E. coli dan S. cerevisiae. P. pastoris merupakan khamir metilotropik, yaitu khamir yang mampu menggunakan metanol sebagai sumber karbonnya (Cereghino & Cregg 2000).
Terdapat tiga faktor yang menyebabkan P. pastoris sangat baik untuk produksi protein rekombinan, yaitu :
1. Biomassa yang dihasilkan tinggi, lebih tinggi 10 kali lipat dibandingkan dengan kultur S. cerevisiae
2. Memiliki promotor yang sangat kuat sehingga tingkat ekspresinya tinggi. Promotor gen AOX1 adalah promotor dari enzim alkohol oksidase yang dapat diinduksi oleh metanol. Jumlah alkohol oksidase di dalam sel bisa mencapai 30% dari total protein sel saat ditumbuhkan dengan substrat metanol.
3. Mensekresikan sangat sedikit protein native sehingga mempermudah proses purifikasi protein rekombinan yang disekresikan.
4. Tidak terjadi proses hiperglikosilasi pada protein rekombinan yang dihasilkan, dimana residu manosa yang ditambahkan hanya 8-14 residu untuk setiap rantai samping.
Vektor ekspresi P. pastoris biasanya dibuat untuk dapat berintegrasi ke dalam kromosom sehingga tidak akan terjadi masalah sedikitnya jumlah produk yang dihasilkan karena ketidakstabilan plasmid. Gen target dan marka seleksi ragi disisipkan ke kromosom melalui proses rekombinasi homolog antara DNA pada vektor dengan daerah yang homolog pada genom P. pastoris (Cregg et al. 2008).
Vektor pPICZ dan pPICZα merupakan vektor ekspresi untuk P. pastoris
yang memiliki tingkat ekspresi yang tinggi. Kedua vektor ini mempunyai marker seleksi antibiotik Zeocin sehingga proses seleksi dapat dilakukan secara langsung. Zeocin juga dapat digunakan untuk E. coli sehingga dapat mengurangi penggunaan antibiotik lain (Gambar 3) (Invitrogen 2008).
Vektor pPICZ dan pPICZα memiliki : (1) promoter AOX1 untuk mengekspresikan protein dengan level tinggi yang diinduksi oleh metanol. (2) c-terminal c-myc epitope dan urutan 6xhis untuk memudahkan proses deteksi dan purifikasi protein. (3) gen 5’AOX1 yang berfungsi untuk mengarahkan integrasi ke genom P. pastoris. Vektor pPICZα juga memiliki signal sekresi α-faktor yang berfungsi untuk menargetkan sekresi protein rekombinan ke medium (Invitrogen 2008).
Gambar 2 Peta vektor pPICZαA (Invitrogen 2008)
Vektor untuk P. pastorisyang lain, seperti pGAPZ dan pGAPZα memiliki
sifat yang berbeda dengan kedua vektor yang disebut terlebih dulu. Vektor ini dapat mengekspresikan protein tanpa memerlukan metanol sebagai penginduksi. Vektor ini memiliki promoter GAP untuk mengekspresikan protein dengan level tinggi, marker seleksi Zeocin untuk seleksi langsung, C-terminal c-myc epitope
serta 6xHis untuk memudahkan proses deteksi dan purifikasi protein. pGAPZα juga mengandung signal sekresi α-faktor yang berfungsi untuk menargetkan sekresi protein rekombinan ke medium.
modifikasi pasca-translasi seperti pada sel mamalia, sel serangga atau sel tanaman. P. pastoris dilaporkan mampu mensintesa beberapa produk protein mamalia seperti human-Insulin, h-IFN-α1, porcine-Lactoferin, h-α-Glucosidase, h-EPO mutan (Skoko et al. 2003) dan beberapa protein antibodi (scFv) (Shi et al. 2003). Sementara itu produksi protein heterolog pada P. pastoris dilaporkan cukup beragam, mulai dari beberapa mg/L sampai beberapa g/L kultur. Perbedaan tingkat ekspresi tersebut dapat disebabkan oleh beberapa hal seperti sifat dan asal protein target, perbedaan kodon, konstruksi vektor ekspresi, serta efisiensi sekresi protein ekstrasel. Peningkatan ekspresi protein rekombinan dapat dilakukan dengan cara optimasi kodon dari gen target dengan kodon preferensi dari sel inang (Cereghino & Cregg 2000).
Green Fluorescent Protein (GFP)
Green fluorescent protein (GFP) adalah protein yang ditemukan dari ubur-ubur (Aequorea victoria) oleh Osamu Shimomura pada tahun 1962 pada saat meneliti aequorin. GFP berhasil diklon pertama kali pada tahun 1992 oleh Prasher dan kemudian diketahui terdiri atas 238 asam amino. Ketiga residu asam amino yaitu serin-tirosin-glisin pada posisi 65-67 berperan sebagai gugus kromofor yang menjadikan GFP dapat berfluoresensi. GFP telah diketahui bersifat non toksik dan dapat diekspresikan dalam jumlah yang cukup tinggi pada berbagai organisme yang berbeda dan sama sekali tidak berpengaruh pada efek fisiologinya. Ketika GFP difusikan dengan suatu protein target yang sedang diteliti maka protein tersebut dapat diekspresikan dan tetap aktif. Di sisi lain GFP sendiri tetap berfluoresensi (Tsien 1998).
Berdasarkan sifat GFP di atas, maka GFP memiliki peranan yang sangat penting dalam penelitian di bidang biologi. GFP dapat digunakan sebagai pewarta dalam mempelajari proses-proses yang terjadi di dalam sel seperti proses transfer protein, mempelajari organel-organel yang terikat pada membran sel dan juga berperan dalam proses pencitraan sel serta jaringan (Huang & Shusta 2006). Roger Tsien (1998) menyebutkan bahwa GFP selain dapat diekspresikan pada
E.coli dan C. elegans juga berhasil diekspresikan pada khamir Sacharomyces cerevisiae dan sel mamalia. Pada saat yang bersamaan juga diketahui bahwa GFP berhasil diekspresikan pada Drosophila melanogaster. Penemuan-penemuan tersebut menunjukkan bahwa GFP dapat diekspresikan pada keempat jenis organisme model sehingga GFP dapat digunakan sebagai penanda genetik universal dalam penelitian di bidang biologi. Roger Tsien (1998) juga menyebutkan bahwa pembentukan gugus kromofor pada GFP terjadi pada proses pasca translasi dan hanya memerlukan molekul oksigen untuk mengaktifkannya. Oleh karena itu, GFP dapat diekspresikan pada semua organisme.
Pada salah satu penelitian fusi GFP dengan fragmen antibodi untai tunggal (scFv) yang dilakukan oleh Huang dan Shusta (2006) diketahui bahwa fusi GFP dengan protein target akan lebih resisten terhadap proses degradasi apabila ditempatkan pada ujung karboksi.
METODOLOGI
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dari bulan Januari 2011 sampai Oktober 2012, bertempat di Laboratorium Protein, Vaksin dan Sistem Penghantaran Terarah, Pusat Penelitian Bioteknologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI), Cibinong, Bogor.
Bahan dan Metode
Gen, Plasmid dan Mikroorganisme
Gen scFv anti EGFRvIII dibuat secara sintetik dan diklon dalam plasmid pJ201 di DNA 2.0 Inc USA berdasarkan sekuen protein anti-EGFRvIII (Weber et al. 2009). Sekuen gen anti-EGFRvIII sintetik telah dioptimasi dengan kodon preferensi P. pastoris. Plasmid yang dihasilkan disebut pAF-scFv-01 (Gambar 3). Plasmid pPICZαA (Invitrogen, USA) digunakan sebagai vektor ekspresi gen
scFv pada sistem ekspresi P. pastoris. E. coli yang digunakan untuk kloning plasmid rekombinan adalah E. coli DH5α dan E. coli TOP10F’. P. pastoris
SMD1168H (Invitrogen, USA) digunakan sebagai sel inang untuk ekspresi protein rekombinan. Plasmid pTZ_EGFP digunakan sebagai sumber gen gfp. Sekuen gen gfp pada pTZ_EGFP berasal dari pEGFP1 (Clontech, USA)
Gambar 3 Skema struktur plasmid pAF_scFv-01
Keterangan : VH= domain untai berat, VL= domain untai ringan, L= Linker, 6xH= polihistidin tag, X= kodon stop; XhoI, ClaI dan XbaI= situs restriksi, KanR= penanda resistensi antibiotik kanamisin.
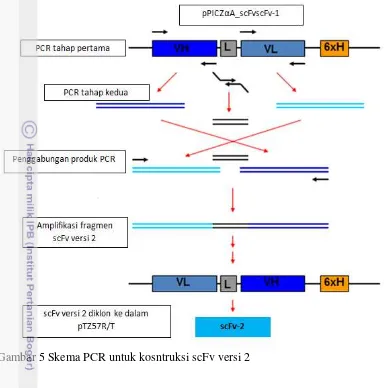
Fragmen antibodi scFv dibuat dalam 2 versi yang berbeda (Gambar 4). Fragmen scFv versi 1 (scFv-1) kemudian difusikan dengan gen gfp sebagai penanda. Fragmen scFv versi 2 (scFv-2) yang memiliki struktur VL-L-VH dibuat dengan teknik overlap extension PCR.
Gambar 4 Target konstruksi scFv anti-EGFRvIII
Media
Media-media yang digunakan dalam penelitian ini adalah Luria Bertani (LB), Low Salt Luria Bertani (LSLB), YPD, YPDS, BMGY, BMMY dan 2xYT. Komposisi media dapat dilihat pada lampiran. Primer DNA yang digunakan dalam penelitian dapat dilihat pada Tabel 1.
Tabel 1 Primer yang digunakan dalam penelitian.
NO. NAMA PRIMER URUTAN NUKLEOTIDA
1. VH101-F 5’-CAA GTT CAA TTG GTT GAG TCA GGA G-3’
2. VH101-R 5’-AGA CGA CAC TGT TAC CAG CGT-3’
3. VL101-F 5’-GAT ATT GTT ATG ACC CAA ACA CCA TTG TC-3’
4. VL101-R 5’-TTT GAT TTC GAC TTT AGT TCC TTG ACC A-3’
5. Linker101-F 5’- GGT CAA GGA ACT AAA GTC GAA ATC AAA GGA GGT GGC GGT TCC GGT GGTGGTGGT AGT GGT-3’
6. Linker101-R 5’- CTC CTG ACT CAA CCA ATT GAA CTT GTG AAC CTC CTC CAC CAC TAC CAC CACCAC CGG AA-3’
7. VH101-F-NdeXho 5’- CAT ATG CTC GAG AAG AGG GAG-3’
Subkloning gen scFv-1 (VH-L-VL) ke dalam Vektor Ekspresi
Gen scFv pada plasmid pAF-scFv-01 memiliki struktur VH-L-VL (disebut scFv-1). VH adalah fragmen variabel untai berat, VL fragmen variabel untai ringan dan L adalah peptida penghubung (linker) yang menghubungkan VH dan VL. Linker (L) yang digunakan memiliki sekuen (G4S)3.
rekombinan kemudian dianalisis dengan cara pemotongan menggunakan enzim restriksi XhoI. Klon yang menunjukkan hasil positif kemudian dipersiapkan untuk analisis urutan nukleotida (sekuensing DNA). Plasmid yang diperoleh disebut pPICZαA_scFv_1. Klon dengan urutan nukleotida yang tepat digunakan dalam langkah berikutnya yaitu fusi gen scFv dengan gen gfp.
Fusi Gen scFv dengan gen gfp
Plasmid rekombinan pPICZαA_scFv_1 digunakan untuk fusi gen scFv
dengan gen gfp. Gen gfp difusikan pada ujung-C dari scFv. Gen gfp diamplifikasi dari plasmid pTZ-EGFP menggunakan pasangan primer EGFP-F-G4S-ClaI (forward) dan EGFP-R-ClaI (reverse). Gen gfp hasil PCR dan plasmid pPICZαA_scFv dipotong dengan enzim restriksi ClaI. Selanjutnya gen gfp
diligasikan dengan plasmid pPICZαA_scFv. Hasil ligasi ditransformasikan ke dalam E.coli TOP10F’. Sel tansforman diseleksi pada medium LSLB yang mengandung antibiotik zeocin (25 µg/ml). Untuk memperoleh klon plasmid rekombinan pPICZαA_scFv(1)_EGFP yang tepat, dilakukan analisis lebih lanjut dengan teknik PCR. Pertama, PCR koloni dilakukan terhadap beberapa klon yang tumbuh dalam media seleksi menggunakan primer AOX1-F (forward) dan AOX1-R (reverse). Klon yang memberikan hasil positif kemudian diekstraksi dari
E. coli. Kedua, PCR plasmid dilakukan terhadap klon yang positif mengandung gen sisipan (egfp) menggunakan primer EGFP-F (forward) dan AOX-R (reverse) untuk menentukan orientasi gen pada plasmid. Plasmid rekombinan yang diperoleh (pPICZαA_scFv(1)_EGFP) kemudian dipotong dengan enzim restriksi
ClaI. Beberapa klon plasmid rekombinan dengan fusi scFv-EGFP yang tepat kemudian disiapkan untuk analisis urutan nukleotida. Klon dengan urutan DNA yang tepat digunakan untuk tahap selanjutnya dan plasmid yang diperoleh disebut sebagai pPICZαA_scFv(1)_EGFP.
Introduksi Plasmid Rekombinan ke dalam P. pastoris
volume beragam pada setiap cawan (50, 25 dan 20 µl). Cawan diinkubasi selama 2-4 hari pada suhu 30°C sampai terlihat koloni yang tumbuh. Selanjutnya koloni tersebut ditumbuhkan kembali pada media YPD baru yang mengandung zeocin dengan konsentrasi zeocin meningkat untuk proses seleksi.
Sel P. pastoris transforman diseleksi pada medium YPD dengan beberapa konsentrasi zeocin meningkat, yaitu 200, 300 dan 1000 µg/ml untuk mendapatkan transforman yang memiliki stabilitas genetik tinggi dan jumlah salinan gen lebih dari satu. Pengujian tersebut dilakukan dengan memilih koloni yang tumbuh baik pada media YPD dengan konsentrasi zeocin 100 µg/ml. Koloni tersebut kemudian digoreskan pada medium YPD dengan konsentrasi zeocin 200 µg/ml. Kultur kemudian diinkubasi selama 1-2 hari pada suhu 30°C. Koloni yang tumbuh baik pada medium tersebut kemudian digoreskan pada media dengan konsentrasi zeocin 300 µg/ml dan diinkubasi pada suhu 30°C selama 1-2 hari. Hal yang sama dilakukan pada media dengan zeocin 1000 µg/ml. Koloni yang tumbuh dengan baik pada media dengan konsentrasi zeocin tertinggi digunakan untuk uji ekspresi protein.
Uji Ekspresi Protein Rekombinan Skala Kecil
Sebanyak 2 ml media BMGY diinokulasi dengan satu koloni transforman kemudian diinkubasi dalam incubator shaker pada suhu 28-30°C dengan agitasi 200 rpm selama 16-18 jam sehingga kultur mencapai nilai OD600 = 2-6. Sel kemudian dipanen dengan cara sentrifugasi pada 3000 g selama 5 menit pada suhu ruang. Supernatan didekantasi kemudian sel pelet diresuspensi dengan 5 ml medium BMMY hingga OD600 = 1. Kultur kemudian diinkubasi dalam incubator shaker pada suhu 28-30 °C, 200 rpm, selama 72 jam. Metanol ditambahkan ke dalam media induksi hingga konsentrasi akhir 0,5% (v/v) setiap 12 jam untuk induksi ekspresi protein rekombinan. Pengambilan contoh sebanyak 300 μl untuk analisis ekspresi protein rekombinan dilakukan pada jam ke 0, 24, 48 dan 72.
Analisis Ekspresi Protein Rekombinan
Sel Pichia transforman dan non transforman diamati dengan mikroskop fluoresens Leica DM 1000. Pengamatan dilakukan terhadap ekspresi EGFP yang akan berpendar hijau dibawah pengamatan mikrokop fluoresens. Sampel adalah sel yang diambil setiap 24 jam setelah induksi dengan methanol, yaitu sampel setelah 24, 48 dan 72 jam induksi. Sebanyak 5-10 μl sel diteteskan pada kaca preparat dan dilihat dengan menggunakan mikroskop fluoresens.
Protein rekombinan yang diekspresikan dianalisis menggunakan SDS-PAGE (konsentrasi poliakrilamid 12%) dan Slot Blot (immunoblotting). Prosedur SDS-PAGE dilakukan sesuai metode dari Ausubel et al (2002) dengan pewarnaan
coomassie blue. SDS-PAGE dimulai dengan pembuatan gel poliakrilamid 12% yang terdiri dari stacking dan separating gel (lampiran). Sampel yang dianalisis merupakan supernatan yang telah dipresitasi sebelumnya dengan menggunakan larutan TCA 100% (trichloroacetic acid) menurut Sanchez (2001).
membran niroselulosa. Membran nitroselulosa kemudian direndam dan digoyang dengan larutan blocking (lampiran) selama 1 jam. Membran lalu dicuci dengan larutan washing (lampiran) sebanyak 3 kali masing-masing selama 15, 5 dan 5 menit. Membran kemudian direndam dan digoyang dengan antibodi primer yang dicampur dengan larutan blocking dengan perbandingan 1:2500 selama 1 jam lalu dicuci seperti pada langkah sebelumnya. Membran lalu direndam dan digoyang lagi dengan antibodi sekunder yang telah dicampur dengan larutan blocking
dengan perbandingan 1:2500 selama 1 jam. Membran kemudian dicuci lagi sesuai dengan langkah sebelumnya dan ditambahkan developer yaitu Western Blue Stabilized Substrate for Alkaline Phosphatase (Promega, USA).
Oleh karena fusi protein scFv-EGFP mengandung His-tag, maka antibody primer yang digunakan adalah mouse anti-His (Promega, USA). Antibodi sekunder yang digunakan adalah goat anti-mouse IgG-AP conjugate (Santa Cruz, USA).
Konstruksi scFv-2 (VL-L-VH)
Fragmen scFv-2 dibuat dengan menggunakan sekuen scFv-1 sebagai cetakan. Dua pasang primer VH-F/VH-R dan VL-F/VL-R digunakan masing-masing untuk amplifikasi fragmen VH dan VL. Pasangan primer linker –F dan linker-R digunakan untuk mengkonstruksi fragmen linker yang masing-masing memiliki daerah overlap dengan fragmen VH dan VL. Fragmen VH, VL dan linker masing-masing diamplifikasi secara terpisah. Produk PCR selanjutnya dipurifikasi dari gel agarose dengan menggunakan kit dari GeneAid. Fragmen scFv-2 (versi-2) ini dikonstruksi dengan metode PCR menggunakan teknik
HASIL DAN PEMBAHASAN
Subkloning scFv-1 (VH-L-VL) ke dalam Vektor Ekspresi
Fragmen gen scFv-1 yang terdapat di dalam plasmid pAF_scFv-1 diklon ke dalam vektor ekspresi pPICZαA. Fragmen scFv diperoleh melalui amplifikasi gen scFv-1 pada pAF_scFv-1 dengan teknik PCR menggunakan pasangan primer VH101-F-NdeXho dan VL101-R-NdeXho. Situs restriksi XhoI yang diperlukan untuk proses subkloning gen ke dalam pPICZαA ditambahkan pada kedua ujungnya. Hasil amplifikasi PCR dari gen scFv-1 dapat dilihat pada Gambar 6.
Gambar 6 Produk amplifikasi PCR plasmid pAF-scFv-1 dengan pasangan primer VH101-F-NdeXho dan VL101F-R-NdeXho.
Keterangan : M= Marka DNA berukuran 1 kb, Lajur 1 dan 2= produk PCR gen scFv
Gambar 7 Hasil potong vektor pPICZαA dan gen scFv hasil PCR dengan enzim
XhoI.
Keterangan : M= Marka DNA berukuran 1 kb, Lajur 1= Vektor pPICZαA (tidak dipotong), Lajur 2= Vektor pPICZαA dipotong dengan enzim XhoI, Lajur 3= gen scFv, Lajur 4= gen scFv dipotong dengan enzim restriksi XhoI.
Proses kloning berikutnya adalah ligasi dan transformasi ke dalam E. coli
TOP10F’ dengan metoda kejut panas (heat shock). Dari hasil transformasi diperoleh sebanyak 42 koloni yang diduga membawa fragmen scFv yang diinsersikan (Gambar 8). Nilai efisiensi transformasi yang dihasilkan adalah sebesar 2,47 x 103 cfu/µg DNA plasmid. Rendahnya nilai efisiensi transformasi dapat disebabkan oleh beberapa faktor, diantaranya adalah konsentrasi DNA plasmid yang digunakan pada saat proses ligasi terlalu rendah, perbandingan antara komponen DNA plasmid dan DNA gen sisipan yang belum optimal, suhu inkubasi pada saat proses ligasi yang belum optimal dan aktivitas enzim ligase yang digunakan. (Brown 2006; Glick & Pasternak 1994).
Gambar 8 Hasil Transformasi pPICZαA_scFv-1 ke dalam E. coli TOP10F’ dengan metoda kejut panas.
Koloni E. coli yang tumbuh diduga mengandung sisipan gen scFv. Koloni E. coli
tersebut diverifikasi dengan beberapa cara, diantaranya adalah teknik PCR koloni, pengecekan orientasi gen dalam konstruksi, pemotongan menggunakan enzim restriksi XhoI dan analisis sekuen DNA.
Gambar 9 Posisi penempelan primer VH101-F dan VL101-R pada plasmid rekombinan pPICZαA_scFv-1
PCR koloni dilakukan untuk memverifikasi keberadaan sisipan gen scFv
di dalam plasmid rekombinan pPICZαA_scFv. PCR koloni dilakukan dengan menggunakan pasangan primer VH101-F dan VL101-R (Tabel 1). Posisi penempelan primer pada plasmid rekombinan pPICZαA_scFv-1 dapat dilihat pada Gambar 9. Hasil PCR koloni menunjukkan bahwa dari 42 sampel koloni yang diperiksa diperoleh 16 sampel yang diduga positif mengandung sisipan gen
scFv. Dengan kata lain hanya 38% sampel yang diduga positif mengandung gen
Gambar 10 Hasil PCR koloni transforman pPICZαA_scFv dengan menggunakan primer VH101-F dan VL101-R.
Keterangan : M= Marka DNA 1 kb, (K+)= Hasil PCR koloni dengan cetakan plasmid pAF-scFv-01, Lajur 1-10= transforman pPICZαA_scFv
Gambar 11 Posisi penempelan primer AOX-F dan VH101-R pada plasmid rekombinan pPICZαA_scFv-1
Penentuan orientasi struktur scFv dilakukan dengan teknik PCR menggunakan pasangan primer AOX-F dengan VH101-R. Penentuan orientasi konstruk dilakukan untuk mengetahui posisi dari sisipan gen scFv telah tepat atau belum, karena terdapat kemungkinan terjadinya orientasi yang salah dari gen scF.
Gambar 12 Hasil PCR penentuan orientasi gen scFv dalam konstruk pPICZαA_scFv dengan menggunakan pasangan primer AOX-F dan VH101-R.
Keterangan : M = marka DNA 1 kb, lajur 1-12 = sampel transforman
Penentuan orientasi konstruk yang dilakukan terhadap 16 sampel klon yang diduga mengandung DNA sisipan diperoleh 8 klon yang mengandung gen scFv
dengan orientasi yang tepat yaitu sebesar 50%. Hasil ini lebih rendah dari nilai yang diharapkan yaitu sekitar 80%. Orientasi gen sisipan di dalam konstruk menentukan aktivitas dari protein yang diekspresikan. Orientasi yang salah akan menyebabkan protein target tidak terekspresi. Gambar 12 menunjukkan sebagian hasil pengecekan orientasi konstruk yang positif, yaitu klon pada lajur ke-2, 3, 4, 5, 7 dan 8.
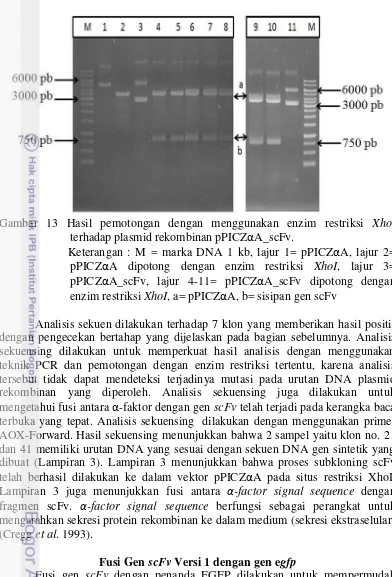
Pemotongan dengan menggunakan enzim restriksi XhoI dilakukan untuk memastikan keberadaan sisipan gen scFv yang terfusi pada situs restriksi tertentu (XhoI). Gambar 13 memperlihatkan hasil pemotongan 8 sampel yang dipotong dengan menggunakan enzim restriksi XhoI. Klon pada kolom ke-4 sampai 10 memberikan 2 pita yaitu pita a yang berukuran 3600 pb dan pita b yang berukuran 750 pb. Pita a diduga merupakan vektor pPICZαA sedangkan pita b merupakan sisipan gen scFv. Hasil pemotongan menggunakan enzim restriksi XhoI dari 8 sampel yang dipotong diperoleh 7 klon sampel yang diduga mengandung sisipan gen scFv. Gambar 13 lajur 11 menunjukkan bahwa satu klon yang dipotong tidak memberikan pita seperti yang diharapkan, yaitu pita a (3600 pb) dan pita b (750 pb). Terdapat beberapa kemungkinan yang menyebabkan tidak terpotongnya plasmid rekombinan pada kedua situs restriksi XhoI. Diantaranya adalah kemungkinan terjadinya mutasi pada salah satu situs restriksi XhoI yang terdapat pada klon tersebut.
Gambar 13 Hasil pemotongan dengan menggunakan enzim restriksi XhoI
terhadap plasmid rekombinan pPICZαA_scFv.
Keterangan : M = marka DNA 1 kb, lajur 1= pPICZαA, lajur 2= pPICZαA dipotong dengan enzim restriksi XhoI, lajur 3= pPICZαA_scFv, lajur 4-11= pPICZαA_scFv dipotong dengan enzim restriksi XhoI, a= pPICZαA, b= sisipan gen scFv
Analisis sekuen dilakukan terhadap 7 klon yang memberikan hasil positif dengan pengecekan bertahap yang dijelaskan pada bagian sebelumnya. Analisis sekuensing dilakukan untuk memperkuat hasil analisis dengan menggunakan teknik PCR dan pemotongan dengan enzim restriksi tertentu, karena analisis tersebut tidak dapat mendeteksi terjadinya mutasi pada urutan DNA plasmid rekombinan yang diperoleh. Analisis sekuensing juga dilakukan untuk mengetahui fusi antara α-faktor dengan gen scFv telah terjadi pada kerangka baca terbuka yang tepat. Analisis sekuensing dilakukan dengan menggunakan primer AOX-Forward. Hasil sekuensing menunjukkan bahwa 2 sampel yaitu klon no. 21 dan 41 memiliki urutan DNA yang sesuai dengan sekuen DNA gen sintetik yang dibuat (Lampiran 3). Lampiran 3 menunjukkan bahwa proses subkloning scFv telah berhasil dilakukan ke dalam vektor pPICZαA pada situs restriksi XhoI. Lampiran 3 juga menunjukkan fusi antara α-factor signal sequence dengan fragmen scFv. α-factor signal sequence berfungsi sebagai perangkat untuk mengarahkan sekresi protein rekombinan ke dalam medium (sekresi ekstraselular) (Cregg et al. 1993).
Fusi Gen scFv Versi 1 dengan gen egfp
Fusi gen scFv dengan penanda EGFP dilakukan untuk mempermudah proses pendeteksian ekspresi protein rekombinan pada P. pastoris (Morino et al.
2001).
menambahkan situs restriksi ClaI yang diperlukan untuk proses fusi gen scFv
dengan gen egfp.
Gambar 14 Hasil amplifikasi (tahap pertama) dengan menggunakan pasangan primer EGFP-F dan EGFP-R
Keterangan : M= Marka DNA 1 kb, lajur 1&2= produk PCR gen
egfp tahap pertama.
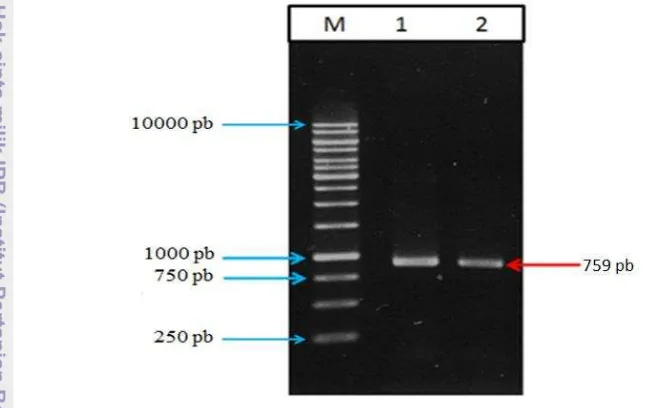
Pada amplifikasi gen egfp tahap pertama dengan pasangan primer EGFP-F dan EGFP-R diperoleh produk PCR dengan ukuran 717 pb (Gambar 14). Produk PCR yang diperoleh kemudian dijadikan sebagai cetakan untuk proses PCR tahap kedua. Proses PCR tahap kedua yang menggunakan pasangan primer EGFP-G4S-ClaI dan EGFP-R-EGFP-G4S-ClaI diperoleh produk PCR dengan ukuran 759 pb (Gambar 15). Amplifikasi gen egfp dengan teknik PCR 2 tahap ini diperlukan untuk mengintroduksi situs restriksi ClaI yang dibutuhkan untuk proses subkloning ke dalam vektor. Pasangan primer yang digunakan pada PCR tahap kedua memiliki situs restriksi ClaI sehingga pada waktu proses subkloning hanya diperlukan 1 situs restriksi. Produk ini kemudian dipurifikasi menggunakan Gel/PCR DNA Fragment Extraction Kit dari GeneAid.
Gambar 15 Hasil amplifikasi gen egfp dengan PCR tahap kedua menggunakan pasangan primer EGFP-G4S-ClaI dengan EGFP-R-ClaI.
Keterangan : M= marka DNA 1 kb, lajur 1&2= produk PCR gen egfp
tahap kedua.
pPICZαA_scFv-1 melalui proses ligasi dan transformasi plasmid rekombinan ke dalam E. coli TOP10F’ dengan metoda kejut panas (heat shock). Gen egfp
difusikan pada ujung-3’ dari gen scFv. Ligasi antara plasmid rekombinan pPICZαA_scFv dengan gen egfp menghasilkan plasmid rekombinan pPICZαA_scFv(1)_EGFP. Gambar 16 menunjukkan hasil transformasi plasmid rekombinan pPICZαA_scFv(1)_EGFP pada E. coli TOP10F’. Hasil transformasi E. coli dengan plasmid rekombinan pPICZαA_scFv(1)_EGFP yang tumbuh pada media LSLB agar yang mengandung zeocin 25 µg/ml mencapai 865 koloni (Tabel 2). Sementara itu, dari proses religasi plasmid tanpa DNA sisipan yang digunakan sebagai kontrol tumbuh 442 koloni. Melihat hasil transformasi tersebut maka diduga hanya sekitar 51% koloni yang tumbuh pada media seleksi tersebut merupakan koloni yang membawa plasmid rekombinan pPICZαA_scFv(1)_EGFP. Salah satu faktor penyebab rendahnya presentase koloni yang diduga membawa plasmid rekombinan pPICZαA_scFv(1)_EGFP adalah kemungkinan pada saat proses defosforilasi enzim alkalin fosfatase tidak bekerja secara optimal karena suhu dan waktu inkubasi yang tidak tepat. Akibatnya masih banyak plasmid yang mengalami religasi (Glick & Pasternak 1994). Proses yang dapat dilakukan untuk mengidentifikasi koloni yang membawa plasmid rekombinan pPICZαA_scFv(1)_EGFP antara lain dengan teknik verifikasi menggunakan PCR koloni terhadap keberadaan gen egfp, penentuan orientasi konstruk fusi scFv dengan gen egfp, pemotongan plasmid rekombinan dengan enzim restriksi ClaI dan analisis sekuensing plasmid rekombinan yang diperoleh.
Tabel 2 Hasil Transformasi Plasmid Rekombinan pPICZαA_scFv(1)_GFP pada E. coliTOP10F’
Plate A B Religasi
1 1 87 120
2 20 97 198
3 33 209 124
4 33 229 -
5 35 121 -
Jumlah 122 743 442
Keterangan : A= Ligasi pPICZαA_scFv dengan EGFP dengan perbandingan 1:4, B= Ligasi pPICZαA_scFv dengan EGFP dengan perbandingan 1:8, Religasi = pPICZαA_scFv.
Gambar 17 Posisi penempelan primer pada proses PCR koloni dengan menggunakan primer AOX-F dan AOX-R
PCR koloni dilakukan terhadap 115 koloni transforman yang tumbuh yang dipilih secara acak dari semua plate. PCR koloni dilakukan dengan menggunakan pasangan primer AOX-F dan AOX-R. Posisi penempelan primer AOX-F dan AOX-R pada plasmid rekombinan dapat dilihat pada Gambar 17. Hasil analisis pada gel agarosa (Gambar 18) menunjukkan terdapat 2 pita produk PCR yang berukuran 2000 pb dan 1200 pb (lajur no. 1 dan 5). Sampel transforman yang memberikan produk PCR dengan ukuran sekitar 2000 pb diduga membawa DNA sisipan gen egfp. Sedangkan transforman yang memberikan produk PCR berukuran 1200 pb diduga tidak mengandung gen sisipan egfp. Hal ini diperoleh dari ukuran komponen plasmid rekombinan, yaitu α- faktor (500 pb), gen scFv
Gambar 18 Hasil PCR koloni transforman dengan menggunakan pasangan primer AOX-F dan AOX-R
Keterangan : M = marka DNA 1 kb, lajur 1-12 = sampel transforman
Gambar 19 Posisi penempelan primer pada proses orientasi konstruk dengan menggunakan pasangan primer EGFP-F dan AOX-R
Gambar 20 Hasil PCR penentuan orientasi gen egfp di dalam konstruk dengan menggunakan pasangan primer EGFP-F dan AOX-R.
Keterangan : M = marka DNA 1 kb, lajur 1-12 = sampel transforman
Verifikasi dengan cara pemotongan menggunakan enzim restriksi ClaI dilakukan terhadap 14 sampel koloni transforman (Gambar 21). Hasil pemotongan dengan enzim restriksi ClaI seharusnya memberikan hasil 2 buah pita yaitu pita plasmid rekombinan pPICZαA_scFv(1) (4350 pb) dan pita gen egfp (800 pb), akan tetapi pada sampel plasmid hasil pemotongan dengan enzim restriksi ClaI tidak diperoleh kedua pita tersebut. Gambar 21 menunjukkan bahwa hasil pemotongan plasmid rekombinan dengan menggunakan enzim restriksi ClaI hanya diperoleh 1 pita DNA yang berukuran sekitar 5000 pb. Pita tersebut diduga diperoleh dari hasil pemotongan hanya pada satu situs restriksi ClaI sehingga plasmid rekombinan menjadi berbentuk linear akan tetapi tidak dapat mengeluarkan sisipan gen egfp. Hal ini dapat diduga disebabkan oleh beberapa hal seperti aktivitas enzim restriksi ClaI yang berkurang atau karena telah terjadi mutasi pada situs restriksi ClaI. Untuk mengetahui penyebab tidak terbentuknya dua pita pada pemotongan dengan enzim restriksi ClaI maka dilakukan analisis sekuensing terhadap 4 sampel yang dipilih secara acak dari 14 koloni yang diperoleh untuk mengidentifikasi urutan nukleotida klon tersebut.
Gambar 21 Hasil pemotongan plasmid rekombinan pPICZαA_scFv(1)_EGFP dengan menggunakan enzim restriksi ClaI.
Hasil analisis sekuensing terhadap 4 koloni transforman menunjukkan bahwa hanya satu klon, yaitu A3 yang memiliki plasmid rekombinan dengan urutan nukleotida yang tepat (Lampiran 4). Hasil sekuensing menunjukkan bahwa fusi antara gen scFv dengan gen egfp telah terjadi pada situs restriksi ClaI. Dua situs restriksi ClaI terdapat pada kedua ujung gen egfp (ujung-3’ dan ujung-5’). Hasil sekuensing ini belum dapat menjelaskan penyebab tidak terpotongnya plasmid rekombinan pPICZαA_scFv(1)_EGFP oleh enzim restriksi ClaI meskipun tidak terjadi mutasi pada situs restriksi tersebut. Penyebab tidak terpotongnya plasmid rekombinan ini dapat disebabkan oleh beberapa hal, diantaranya adalah aktivitas enzim ClaI yang digunakan kemungkinan belum optimal, suhu dan waktu inkubasi pada saat proses digesti dengan enzim restriksi yang kurang optimal atau kemungkinan karena telah terjadi proses metilasi pada situs restriksi ClaI. Hasil analisis sekuen DNA dari beberapa klon yang lain menunjukkan terjadinya mutasi baik pada bagian gen scFv maupun gen egfp.
Introduksi Plasmid Rekombinan ke dalam P. pastoris
P. pastoris yang digunakan sebagai inang dalam proses transformasi ini adalah P. pastoris SMD1168H yang memiliki genotip His4 dan pep4 serta fenotip Mut+, His- dan pep4-. Proses transformasi plasmid rekombinan pPICZα_scFv(1)_EGFP ke dalam P. pastoris diawali dengan preparasi sel kompeten P. pastoris dan plasmid rekombinan pPICZα_scFv(1)_EGFP. Sel kompeten P. pastoris dibuat sesuai dengan prosedur pembuatan sel kompeten Invitrogen Easy SelectTM Pichia Expression Kit.
Plasmid rekombinan pPICZα_scFv(1)_EGFP diisolasi dari E. coli
TOP10F’ (Gambar 22) dan kemudian dipurifikasi menggunakan Gel/PCR DNA Fragment Extraction Kit dari GeneAid. Kemurnian dari plasmid yang akan ditransformasikan ke dalam P. pastoris akan sangat berpengaruh terhadap hasil transformasi. Pita DNA hasil isolasi (Gambar 22) menunjukkan adanya 3 pita DNA yang terbentuk. Diduga pita yang berada paling atas adalah pita DNA genom oleh karena itu harus dilakukan purifikasi terhadap preparasi plasmid tersebut. Plasmid rekombinan tersebut kemudian dipotong dengan menggunakan enzim restriksi SacI agar diperoleh DNA plasmid yang linear. Menurut Cregg et al. 1989 pemotongan DNA plasmid rekombinan ini bertujuan untuk meningkatkan hasil transformasi, karena DNA yang telah dipotong akan mengarahkan proses rekombinasi secara langsung pada sekuen DNA target, yaitu sekuen promotor AOX1. Di dalam Easy SelectTM Pichia Expression Kit disebutkan bahwa apabila digunakan plasmid DNA yang sudah dipotong (linearized) maka DNA yang diperlukan untuk proses transformasi sebesar 5-10 μg, sedangkan apabila digunakan DNA sirkular, maka dibutuhkan jumlah DNA yang jauh lebih besar yaitu 50-100 μg DNA. Plasmid yang telah dipotong dengan enzim restriksi
Gambar 22 Hasil isolasi DNA plasmid pPICZαA_scFv(1)_EGFP
Keterangan : K = kontrol plasmid pPICZαA_scFv, lajur 1&2= plasmid pPICZαA_scFv_EGFP
Gambar 23 Hasil potong plasmid pPICZαA_scFv(1)_EGFP dengan menggunakan enzim restriksi SacI.
Keterangan : M = marka DNA 1 kb, Lajur 1 = plasmid pPICZαA (tidak dipotong), Lajur 2 = plasmid pPICZαA dipotong dengan enzim restriksi SacI, Lajur 3 = plasmid pPICZαA_scFv(1)_EGFP, Lajur 4-5 = plasmid pPICZαA_scFv(1)_EGFP dipotong dengan enzim restriksi
SacI.
Tabel 3 Hasil penentuan konsentrasi dan kemurnian plasmid rekombinan pPICZαA_scFv(1)_EGFP dengan menggunakan nanofotometer
Sampel A260/280 Konsentrasi (ng/µl)
pPICZαA_scFv_EGFP (ekstraksi ke-1) 2,5 12,1 pPICZαA_scFv_EGFP (ekstraksi ke-2) 3,0 7,3
Proses transformasi pada P. pastoris dilakukan sesuai dengan prosedur yang terdapat pada Easy SelectTM Pichia Expression Kit (Invitrogen) dengan metoda elektroporasi menggunakan alat elektroporator (BioRad). Proses elektroporasi yang dilakukan menggunakan protokol dan kondisi elektroporasi yang optimal untuk transformasi P. pastoris. Kondisi pada saat proses elektroporasi dapat dilihat pada Tabel 4. DNA plasmid rekombinan pPICZαA_scFv(1)_EGFP sebanyak 20 µl (0,24 µg) ditransformasikan ke dalam 40 µl sel kompeten P. pastoris. Ke dalam campuran antara plasmid rekombinan dan sel kompeten kemudian ditambahkan 1 ml sorbitol 1 M segera setelah proses elektroporasi. Penambahan sorbitol bertujuan untuk menjaga tekanan osmotik sel agar tidak pecah setelah proses elektroporasi dan membantu sel untuk memperbaiki diri. Selain itu sorbitol juga berfungsi untuk menjaga agar DNA yang sudah masuk ke dalam sel tidak kembali lagi keluar sel (Invitrogen 2001).
Tabel 4 Kondisi pada saat elektroporasi plasmid rekombinan pPICZαA_scFv(1)_EGFP dengan menggunakan alat Elektroporator
Keterangan : kontrol (sel kompeten P. pastoris SMD1168H)
Gambar 24 Hasil transformasi pPICZαA_scFv(1)_EGFP pada P. pastoris
Keterangan : A, B & C menunjukkan sel hasil elektroporasi yang disebar masing-masing 50 μl (A), 25μl (B) & 20 μl (C).
Ada beberapa metoda transformasi yang dapat digunakan untuk P. pastoris, diantaranya adalah elektroporasi dan spheroplast. Metoda elektroporasi memiliki beberapa keunggulan dibanding metoda spheroplast. Keungguluan yang pertama adalah sangat efisien karena memungkinkan proses seleksi langsung pada media yang mengandung zeocin. Keunggulan yang kedua adalah metoda ini memberikan hasil transformasi yang lebih tinggi dibandingkan dengan metoda
spheroplast (Cregg et al. 1985). Pada metoda spheroplast, dinding sel yang mengalami kerusakan akan memerlukan waktu untuk pemulihan sehingga meningkatkan sensitivitas sel terhadap zeocin. Oleh karena itu sel yang mungkin saja membawa plasmid rekombinan akan tidak dapat bertahan hidup sebelum sempat mengekspresikan gen resistensi zeocin yang telah dimiliki. Untuk memperoleh nilai efisiensi transformasi yang tinggi selain memperhatikan faktor-faktor yang telai diuraikan di bagian sebelumnya juga harus dilakukan optimasi proses elektroporasi agar diperoleh kondisi elektroporasi yang optimal.
Seleksi P. pastoris Transforman
Seleksi P. pastoris transforman dilakukan untuk mencari transforman yang memiliki stabilitas genetik dan kemungkinan jumlah salinan gen lebih dari 1. Dari 504 transforman yang diperoleh sebanyak 130 transforman diseleksi kembali pada media dengan konsentrasi zeocin yang meningkat, yaitu 200, 300 dan 1000 µg/ml. Hasil seleksi dan uji tumbuh galur transforman pada medium seleksi tersebut dapat dilihat pada Tabel 6 dan Gambar 25-28.
Tabel 6 Persentase hasil seleksi P. pastoris transforman
Konsentrasi zeocin (μg/ml) Koloni yang diuji
Klon P. pastoris transforman yang sebelumnya tumbuh pada media dengan zeocin yang lebih rendah konsentrasinya, ternyata setelah ditumbuhkan kembali pada media dengan zeocin yang konsentrasi yang lebih tinggi klon tersebut tidak tumbuh. Hal ini kemungkinan disebabkan karena proses integrasi DNA pada klon tersebut tidak stabil. Hanya transforman yang DNA plasmid rekombinannya telah terintegrasi ke dalam genom yang dapat tumbuh dengan baik. Seleksi menggunakan media dengan konsentrasi zeocin yang tinggi dapat menghasilkan transforman yang lebih stabil dan plasmid rekombinan yang mengandung marka resistensi zeocin telah terintegrasi stabil di dalam sel (Cereghino et al. 2008). Transforman yang dapat tumbuh pada medium dengan konsentrasi zeocin 100
Gambar 25 Hasil seleksi transforman P. pastorid pada media YPD agar yang mengandung zeocin 100 µg/ml
Keterangan : A= media YPD agar; B, C, D & E= media YPD agar + zeocin 100 µg/ml
Gambar 26 Hasil seleksi transforman P. pastoris pada media YPD agar yang mengandung zeocin 200 µg/ml
Keterangan : A= media YPD agar; B, C, D, E & F= media YPD agar + zeocin 200 µg/ml
Gambar 27 Hasil seleksi transforman P. pastoris pada media YPD agar yang mengandung zeocin 300 µg/ml
Keterangan : A= media YPD agar; B, C & D= media YPD agar + zeocin 300 µg/ml
Gambar 28 Hasil seleksi transforman pada media YPD agar yang mengandung antibiotik zeocin 1000 µg/ml
Uji Ekspresi dan Analisis Protein Rekombinan Skala Kecil
Uji ekspresi protein rekombinan skala kecil dilakukan terhadap transforman yang tumbuh dengan baik pada konsentrasi zeocin 300 µg/ml (21 koloni transforman). Pengujian ini bertujuan untuk menguji transforman tersebut akan mengekspresikan protein rekombinan yang diinginkan.
Proses ekspresi protein rekombinan terbagi dalam 2 tahap yaitu produksi biomassa sel P. pastoris transforman dan proses induksi protein fusi scFv-EGFP rekombinan. Proses produksi biomassa sel berlangsung dengan menggunakan media BMGY sedangkan proses induksi menggunakan media BMMY.
Analisis protein rekombinan dilakukan terhadap sampel yang merupakan cairan kultur bebas sel. Protein dipresipitasi menggunakan larutan TCA. Hasil SDS PAGE (Gambar 29) dengan pewarna coomassie blue menunjukkan bahwa telah terbentuk beberapa pita protein yang diduga merupakan protein target yaitu fusi scFv-EGFP. Ukuran protein target dalam penelitian ini adalah 60 kDa. Gambar 29 memperlihatkan bahwa pita protein yang dihasilkan masih beragam karena protein rekombinan yang dihasilkan belum melalui proses purifikasi. Untuk mengkonfirmasi bila protein tersebut telah berhasil diekspresikan dari P. pastoris, maka diperlukan pengujian tambahan salah satunya dengan teknik immunoblotting dengan menggunakan antibodi spesifik.
Gambar 29 Hasil analisis ekspresi protein rekombinan dengan teknik SDS-PAGE Keterangan : M : marka protein 250 kDa, K1 dan K2 = kontrol pastoris, T1-T7 = P. pastoris transforman
Gambar 30 Hasil analisis immunoblotting menggunakan teknik Slot Blot terhadap sampel protein beberapa galur P. pastoris transforman.
Keterangan : A= Air, M= Media, K= kontrol non transforman, T1-T9= sampel protein dari beberapa galur P. pastoris transforman.
Gambar 30 menunjukkan bahwa galur P. pastoris transforman memiliki tingkat ekspresi yang beragam. Hal ini terlihat dari perbedaan intensitas warna antara galur yang satu dengan yang lainnya. Galur T2 terlihat memiliki intensitas warna ungu yang lebih tebal jika dibandingkan dengan galur yang lain, dan ini berarti bahwa galur T2 memiliki tingkat ekspresi paling tinggi. Sedangkan pada galur lain terlihat sangat tipis bahkan tidak terlihat sama sekali. Tidak terlihatnya warna ungu kemungkinan disebabkan oleh rendahnya afinitas antara antigen dengan antibodi yang digunakan serta preparasi sampel protein yang kurang baik pada proses blotting (Moore 2009).
Sel transforman dan non transforman P. pastoris diamati dengan menggunakan mikroskop fluoresens Leica DM1000 (Gambar 31). Pengamatan ini bertujuan untuk mengetahui keberhasilan proses ekspresi protein rekombinan fusi scFv-EGFP dengan melihat pendaran EGFP pada panjang gelombang 450-490 nm (Morino et al. 2001). Fluoresensi diduga berasal dari EGFP yang berada pada bagian retikulum endoplasma, badan golgi atau terakumulasi di permukaan sel (Naumann et al. 2011).
Gambar 31 Hasil pengamatan sel transforman P.pastoris yang mengandung gen penanda GFP yang berfluoresen hijau.
Keterangan : A= sel pada fase kontras, B= sel pada fase kontras & fluoresen, dengan perbesaran 100x. Nomor pada gambar menunjukkan nomor kloni transforman.
Pengamatan dengan menggukan mikroskop menunjukkan terlihat perbedaan antara kontrol P. pastoris non transforman (K) dengan P. pastoris transforman (diwakili oleh sampel no. 1, 6, 10 dan 100) yaitu pada fase kontras dan fase fluoresen. Pada fase tersebut P. pastoris transforman yang mengandung fusi scFv-EGFP berfluoresen hijau (ditunjukkan oleh panah berwarna merah pada Gambar 30). Hasil pengamatan ini mempermudah proses pendeteksian keberhasilan transformasi fusi scFv-EGFP ke dalam P. pastoris. Selain galur-galur yang terlihat berfluoresensi yang menunjukkan terdapat fusi antara scFv dengan EGFP ditemukan juga beberapa galur yang tidak terlihat fluoresensinya. Penyebab tidak terlihatnya pendaran ini kemungkinan karena ekspresi protein belum optimal, ekspresi sebenarnya ada hanya jumlahnya terlalu sedikit atau sangat lemah sehingga tidak dapat terlihat dan mungkin karena beberapa galur tertentu memiliki tingkat ekspresi yang lebih rendah dibanding yang lain (Morino et al. 2001).
Konstruksi ScFv Versi 2 (VL-L-VH) dengan Menggunakan Teknik OverlapExtension PCR
Proses konstruksi scFv-2 dilakukan dengan teknik PCR menggunakan gen scFv-1 sebagai cetakan. Tahapan proses konstruksi dapat dilihat pada Gambar 5. scFv anti-EGFRvIII dibuat dalam dua versi berbeda dengan tujuan untuk mengetahui pengaruh posisi domain VH, Linker dan VL terhadap aktivitas dan afinitas fragmen antibodi tersebut. Tidak ada suatu ketentuan umum yang menyatakan bahwa konstruk dengan susunan VH-L-VL lebih aktif daripada konstruk dengan susunan VL-L-VH. Aktivitas dan afinitas fragmen antibodi tersebut tergantung kepada gennya itu sendiri (Chang et al. 2008).
Gambar 32 Hasil amplifikasi PCR fragmen VH, VL dan Linker.
Keterangan : M= marka DNA 100 bp, lajur 1= fragmen VH, lajur 2= fragmen VL, lajur 3 fragmen Linker
Proses PCR bagian kedua terdiri dari 2 tahap, yaitu untuk menggabungkan fragmen VL dengan Linker (VL-L) serta Linker dengan VH (L-VH).
Proses ini kemudian diikuti dengan tahapan yang ketiga, yaitu menggabungkan bagian 1 (VL-L) dengan bagian 2 (L-VH). Proses ini dibedakan dalam 2 reaksi PCR. Reaksi yang pertama adalah penggabungan kedua bagian tersebut, sedangkan reaksi yang kedua adalah amplifikasi fragmen VL-L-VH yang telah terbentuk dari reaksi PCR sebelumnya. Rangkaian proses PCR ini menghasilkan fragmen scFv versi 2 dengan struktur VL-L-VH.
Gambar 33 Hasil amplifikasi PCR dan purifikasi fragmen scFv-2 menggunakan pasangan primer VL-F dan VH-R.
Keterangan : M= Marka DNA 1 kb, Lajur 1 dan 2 = fragmen scFv versi 2
Proses berikutnya adalah subkloning fragmen scFv-2 ke dalam vektor pTZ57R/T. Hasil ligasi ditransformasikan pada E. coli DH5α. Hasil transformasi disebar pada media LB agar yang mengandung antibiotik ampicillin (50 μg/ml) yang telah ditambahkan X-Gal dan IPTG. Koloni yang tumbuh setelah diinkubasi selama 16-18 jam sebanyak 114 koloni (Gambar 34). Koloni tersebut semuanya berwarna putih. Koloni putih diduga membawa plasmid rekombinan yang diinginkan yaitu pTZ_scFv. Verifikasi terhadap koloni putih tersebut dilakukan dengan teknik PCR, pemotongan menggunakan enzim restriksi dan analisis sekuen DNA.
Gambar 34 Hasil transformasi plasmid rekombinan pTZ_scFv ke dalam E. coli
DH5α
Verifikasi pertama terhadap E. coli transforman yang diperoleh dilakukan dengan teknik PCR koloni untuk menentukan keberadaan gen sisipan (scFv) menggunakan pasangan primer VL101F-Xho dan VH101R-Xba. Reaksi PCR ini akan menghasilkan pita DNA yang berukuran sekitar 750 bp. PCR koloni dilakukan terhadap 71 klon E. coli yang dipilih secara acak. Koloni E. coli
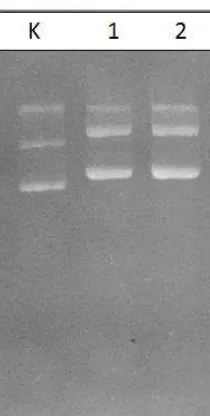
transforman yang diduga membawa plasmid rekombinan pTZ_scFv diperoleh sebanyak 19 klon (Gambar 35). Koloni E. coli transforman yang diduga membawa plasmid rekombinan pTZ_scFv ditunjukkan pada Gambar 35. Sampel tersebut kemudian diisolasi DNA plasmidnya untuk digunakan dalam proses verifikasi selanjutnya (Gambar 36).
Gambar 35 Hasil PCR koloni E. coli transforman pTZ_scFv-2
Keterangan : M= marka DNA 1 kb, (K+)= kontrol positif pPICZαA_scFv, Lajur 1-11= sampel E. coli transforman, a= menunjukkan koloni transforman yang positif.
Gambar 36 Hasil isolasi DNA plasmid pTZ_scFv-2 dari beberapa klon E. coli
ransforman.