RESPON GENOTIPE TANAMAN KEDELAI (
Glycine max
L.
Merrill) DARI BERBAGAI NEGARA TERHADAP KONDISI
LINGKUNGAN TUMBUH AGROEKOSISTEM
TROPIKA BASAH
DEWI VALENTINA BUTAR BUTAR
AGRONOMI DAN HORTIKULTURA FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN SUMBER
INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Respon Genotipe Tanaman Kedelai (Glycine max L. Merrill) dari Berbagai Negara Terhadap Kondisi Lingkungan Tumbuh Agroekosistem Tropika Basah adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
ABSTRAK
DEWI VALENTINA BUTAR BUTAR. Respon Genotipe Tanaman Kedelai (Glycine max L. Merrill) dari Berbagai Negara Terhadap Kondisi Lingkungan Tumbuh Agroekosistem Tropika Basah. Dibimbing oleh ISKANDAR LUBIS.
Penelitian ini bertujuan untuk mempelajari respon pertumbuhan dan produksi 130 genotipe kedelai dari berbagai negara terhadap kondisi lingkungan tumbuh agroekosistem tropika basah. Percobaan ini disusun dengan rancangan augmented dalam rancangan acak lengkap (RAL). Benih kedelai yang digunakan sebagai pembanding (kontrol) adalah kedelai varietas Tanggamus, Anjasmoro, Argo Mulyo, Wilis dan Tachinagaha (varietas Jepang) yang terdiri dari 5 ulangan sehingga terdapat 150 satuan percobaan. Hasil penelitian menunjukkan bahwa genotipe GmWMC192 yang berasal dari Nepal berdaya hasil tinggi dan beradaptasi baik pada lahan kebun percobaan IPB Sawah Baru. Hal ini dilihat dari persentase tanaman yang tumbuh di lapangan, tinggi tanaman, jumlah buku, jumlah polong per tanaman, dan persentase biji penuh yang tinggi. Selain itu, genotipe GmWMC019 dan GmWMC042*2 yang berasal dari Korea Utara dan Republik Rakyat Cina berumur sangat genjah (66 hari) serta beradaptasi baik pada lahan kebun percobaan IPB Sawah baru. GmWMC019 dan GmWMC042*2 merupakan genotipe yang peka terhadap suhu tinggi.
Kata kunci: biji penuh, jumlah buku, rancangan augmented, tropika basah
ABSTRACT
DEWI VALENTINA BUTAR BUTAR. Response of Soybean Genotypes (Glycine max L. Merrill) from Various Countries to Environmental of Grow Conditions at Wet Tropical Agroecosystem. Supervised by ISKANDAR LUBIS
This research aims to study the response of growth and production of 130 soybean genotypes from various countries to the condition at wet tropical agroecosystem. The experimental design used was augmented design in randomized complete design (RCB). Soybean seed that was used as check varieties are Tanggamus, Anjasmoro, Argo Mulyo, Wilis and Tachinagaha (Japan soybean variety) consist of 5 replicates so there are 150 units of the experiment. The results indicated that the genotype GmWMC192 from Nepal has high result and adaptable at the field of Bogor Agricultural University. It is seen from the percentage of the survived plants, plant height, the number of node, number of pods per plant, and higher percentage of full seeds. In addition, genotypes GmWMC042*2 and GmWMC019 from North Korea and China have very fast growth (66 days) and also adaptable at the field of Bogor Agricultural University. Both of the genotypes may be useful as a source in soybean breeding.
RESPON GENOTIPE TANAMAN KEDELAI (
Glycine max
L.
Merrill) DARI BERBAGAI NEGARA TERHADAP KONDISI
LINGKUNGAN TUMBUH AGROEKOSISTEM
TROPIKA BASAH
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Agronomi dan Hortikultura
AGRONOMI DAN HORTIKULTURA FAKULTAS PERTANIAN INSTITUT PERTANIAN BOGOR
BOGOR 2014
Judul Skripsi : Respon Genotipe Tanaman Kedelai (Glycine max L. Merrill) dari Berbagai Negara Terhadap Kondisi Lingkungan Tumbuh Agroekosistem Tropika Basah
Nama : Dewi Valentina Butar Butar NIM : A24100039
Disetujui oleh
Dr Ir Iskandar Lubis, MS
Pembimbing
Diketahui oleh
Dr Ir Agus Purwito, MSc. Agr Ketua Departemen
PRAKATA
Puji syukur penulis panjatkan kehadirat Tuhan Yang Maha Esa karena telah memberi kekuatan dan kelancaran sehingga skripsi ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Februari 2014 ini adalah seleksi genotipe kedelai dari berbagai negara, dengan judul Respon Genotipe Tanaman Kedelai (Glycine max L. Merrill) dari Berbagai Negara Terhadap Kondisi Lingkungan Tumbuh Agroekosistem Tropika Basah.
Penulis mengucapkan terimakasih kepada semua pihak yang telah membantu dalam penyusunan skripsi ini, terkhusus penulis sampaikan kepada:
Bapak almarhum Abel Butar Butar, Ibu Almarhumah Rosmaida Sitinjak, serta seluruh keluarga besar yang telah memberikan dukungan baik materi dan moril
Dr Ir Iskandar Lubis, MS selaku dosen pembimbing yang telah memberikan pengarahan sampai skripsi ini berhasil diselesaikan
Dr Ir Hajrial Aswidinnoor, MSc selaku dosen pembimbing akademik yang telah memberikan pengarahan selama mengikuti perkuliahan di IPB
Prof Dr Tatsuhiko Shiraiwa dan Dr Koki Homma selaku dosen di Universitas Kyoto, Jepang yang telah memberikan benih kedelai Jepang untuk penelitian ini
National Institute of Agrobiological Sciences (NIAS) yang telah memberikan benih kedelai dari berbagai negara untuk penelitian ini.
Prof Dr. Ir. Munif Ghulamahdi, MS dan Dr. Ir. Trikoesoemaningtyas, MSc yang telah memberikan galur-galur kedelai untuk penelitian ini
Dr Willy Bayuardi Suwarno, SP, MSi yang telah memberikan pengarahan pengolahan data penelitian
Bapak Rahmat selaku penanggung jawab Laboratorium Ilmu dan Teknologi Benih, Departemen Agronomi dan Hortikultura, Institut Pertanian Bogor
Gatra Saputra selaku alumni angkatan 46 yang telah membantu penulis selama penelitian
Para petani yang bekerja di Sawah Baru, Kecamatan Dramaga, Kabupaten Bogor
Semua teman-teman yang telah membantu penulis selama penelitian. Semoga skripsi ini dapat bermanfaat.
DAFTAR ISI
DAFTAR TABEL viii
DAFTAR GAMBAR viii
DAFTAR LAMPIRAN viii
PENDAHULUAN
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis Penelitian 2
TINJAUAN PUSTAKA
Botani dan Morfologi Kedelai 3
Bunga 3
Perkembangan Polong 3
Biji 3
Fase Tumbuh 4
Respon Kedelai Terhadap Lingkungan Tumbuh 4
Rancangan Augmented 7
METODE
Waktu dan Tempat 7
Bahan dan Alat 7
Metode Penelitian 8
Analisis Data 8
Pelaksanaan Penelitian 8
Pengamatan 9
HASIL DAN PEMBAHASAN
Kondisi Umum Penelitian 9
Keragaan Karakter Agronomi dan Hasil Genotipe-Genotipe Kedelai 10
Persentase Tumbuh 11
Umur Berbunga (Fase R1) 13
Waktu Muncul Polong (Fase R3) 15
Waktu Perubahan Polong Menjadi Kuning, Coklat, Matang (Fase R7) 16
Umur Panen (Fase R8) 18
Tinggi Tanaman pada Fase R1 19
Tinggi Tanaman pada Fase R7 21
Jumlah Buku pada Fase R8 23
Jumlah Cabang per Tanaman 24
Bobot Kering Tajuk 27
Jumlah Polong per Tanaman 27
Ukuran Biji dan Warna Biji 30
Persentase Kondisi Biji 31
Kadar Air Biji 34
Korelasi Antarkarakter 37
SIMPULAN DAN SARAN 40
DAFTAR PUSTAKA 40
LAMPIRAN 43
DAFTAR TABEL
5 Hasil analisis sifat kimia dan fisik tanah kebun percobaan IPBSawah Baru 10 10 Jumlah buku rata-rata pada fase R1 dari 118 genotipe kedelai
yang diuji dan kelima varietas pembanding 23 11 Jumlah buku rata-rata pada fase R8 dari 118 genotipe kedelai
yang diuji dan kelima varietas pembanding 25 12 Jumlah cabang rata-rata dari 118 genotipe kedelai yang diuji
dan kelima varietas pembanding 26
13 Bobot kering tajuk rata-rata dari 118 genotipe kedelai yang
diuji dan kelima varietas pembanding 28
14 Jumlah polong rata-rata per tanaman dari 118 genotipe kedelai
yang diuji dan kelima varietas pembanding 29
15 Pengelompokan biji berdasarkan ukuran 30
16 Pengelompokan warna biji dari masing-masing genotipe 31 17 Persentase biji penuh, keriput, rusak, dan unfertile dari 125
genotipe kedelai yang diuji dan kelima varietas pembanding 32 18 Kadar air benih dari 118 genotipe kedelai yang diuji dan
kelima varietas pembanding 35
19 Korelasi fenotipik antarkarakter varietas/galur kedelai pada
pengujian di kebun percobaan IPB Sawah Baru 39
DAFTAR GAMBAR
1 Tampilan daya tumbuh benih dilapangan 13
2 Tampilan benih yang dorman 13
3 Tampilan serangan lalat bibit 13
4 Umur berbunga (fase R1) 14
6 Tampilan serangan ulat penggulung daun 14
7 Tampilan serangan penyakit rebah semai 14
8 Tampilan gulma yang dominan tumbuh dilapangan 14
9 Waktu muncul polong (fase R3) 15
10 Tampilan tanaman pada fase R3 16
11 Tampilan serangan kepik polong 16
12 Waktu perubahan warna polong (fase R3) 16
13 Tampilan tanaman pada fase R7 17
14 Tampilan tanaman yang terkena penyakit karat daun 17
15 Umur panen (fase R8) 18
16 Tampilan tanaman pada fase R8 19
17 Tampilan kegiatan panen 19
18 Tampilan persentase kondisi benih 34
19 Tampilan alat dan pengunaan Moisture Tester 36
DAFTAR LAMPIRAN
1 Identitas genotipe kedelai yang digunakan di percobaan 44
2 Deskripsi varietas benih kedelai pembanding 50
3 Tata letak percobaan dilapangan 54
4 Pengelompokan benih berdasarkan iklim 55
PENDAHULUAN
Latar Belakang
Kedelai (Glycine max L. Merril) termasuk komoditas tanaman pangan terpenting setelah padi dan jagung. Kedelai merupakan salah satu sumber protein yang paling banyak dikonsumsi masyarakat Indonesia karena harganya yang relatif terjangkau. Kebutuhan kedelai akan terus meningkat seiring dengan meningkatnya permintaan sebagai bahan baku industri pangan. Konsumsi kedelai pada tahun 2013 sebesar 2.5 juta ton (BAPPENAS 2014) sedangkan produksi kedelai nasional tahun 2013 sebesar 807.57 ribu ton biji kering atau turun 4.22 % dibandingkan tahun 2012 (BPS 2013). Peningkatan konsumsi kedelai nasional tersebut sangat dipengaruhi oleh peningkatan jumlah penduduk Indonesia. Jumlah penduduk Indonesia pada tahun 2013 sebesar 242 juta jiwa (BPS 2013). Kekurangan kedelai harus dipenuhi lewat impor sebesar 624 000 ton sampai akhir tahun 2013, sehingga total kedelai impor nasional setiap tahun terus meningkat. Konsumsi kedelai yang terus meningkat tersebut membutuhkan adanya upaya untuk meningkatkan produktivitas nasional.
Peningkatan produktivitas kedelai nasional saat ini dihadapkan pada berbagai tantangan seperti adanya persaingan dengan komoditas lain, alih fungsi lahan dan perubahan iklim yang mengakibatkan intensitas serangan organisme pengganggu tanaman (OPT) meningkat sehingga berdampak nyata pada upaya peningkatan produksi kedelai. Tantangan tersebut mengakibatkan pemenuhan kedelai nasional sebagian besar masih bergantung pada negara lain. Berbagai penelitian terus dilakukan untuk meningkatkan produktivitas kedelai, salah satu upaya yang dilakukan adalah program pemuliaan tanaman. Tantangan yang akan dihadapi dalam program pemuliaan tanaman semakin besar dengan adanya pemanasan global. Pemanasan global mengakibatkan terjadinya peningkatan suhu di sentra-sentra produksi kedelai. Sumarno dan Manshuri (2007) menyatakan suhu yang tinggi mengakibatkan terjadinya aborsi polong pada tanaman kedelai sehingga berpengaruh terhadap penurunan pertumbuhan dan produktivitas tanaman kedelai.
setelah hibridisasi. Koleksi berbagai genotipe (plasma nutfah) dapat berupa nutfah lokal atau yang diintroduksikan dari luar negeri yang digunakan sebagai keragaman sumber gen untuk mendapatkan genotipe yang diinginkan untuk tujuan pemuliaan. Tahapan selanjutnya adalah kegiatan seleksi dengan cara memilih beberapa tanaman terbaik dari suatu populasi tanaman. Kajian ini penting digunakan untuk mendapatkan genotipe-genotipe yang beradaptasi baik dan berpenampilan stabil pada lingkungan tropika basah tersebut. Rasyad dan Idwar (2010) menjelaskan galur atau varietas yang stabil umumnya mempunyai keragaman yang kecil jika ditanam pada kondisi lingkungan yang berbeda atau memiliki keragaan yang tetap pada berbagai lingkungan. Oleh karena itu, varietas yang stabil akan memberikan tanggap hasil yang relatif sama meskipun lingkungannya berbeda. Kestabilan suatu varietas bukan hanya ditunjukkan oleh hasilnya saja tetapi juga diperlihatkan oleh kestabilan sifat-sifat agronomis lain seperti komponen hasilnya.
Varietas kedelai dari wilayah subtropika yang sesuai untuk panjang hari 14-16 jam apabila ditanam di Indonesia yang panjang harinya 12 jam maka akan mempercepat pembungaan pada umur 20-22 hari walaupun batang tanaman masih pendek, dan tanaman sudah berbunga. Varietas asal subtropika di tempat aslinya, berbunga pada umur tanaman sekitar 50 hari saat batang kedelai sudah tumbuh setinggi 60-70 cm (Sumarno dan Manshuri 2007). Oleh karena itu, penelitian respon genotipe tanaman kedelai dari berbagai negara terhadap kondisi lingkungan tumbuh agroekosistem tropika basah perlu dilakukan untuk mempelajari pengaruh perbedaan lingkungan tumbuh genotipe-genotipe kedelai tersebut terutama akibat suhu terhadap pertumbuhan dan produksinya. Genotipe yang memiliki karakter unggul tersebut, selanjutnya dapat diusulkan kepada pemulia tanaman untuk dikembangkan sebagai sumber gen dalam melakukan persilangan tanaman untuk merakit varietas unggul nasional.
Tujuan Penelitian
Penelitian ini bertujuan mempelajari respon pertumbuhan dan produksi 130 genotipe kedelai dari berbagai negara terhadap lingkungan tumbuh agroekosistem tropika basah.
Hipotesis
1. Terdapat perbedaan respon pertumbuhan dan produksi diantara genotipe-genotipe yang diuji.
2. Terdapat satu atau lebih genotipe yang memiliki pertumbuhan dan produksi lebih tinggi daripada pembanding.
TINJAUAN PUSTAKA
Botani dan Morfologi Kedelai
Tanaman kedelai (Glycine max L. Merril) merupakan anggota dari famili Leguminosae, subfamili Papilionideae dan tergolong kedalam genus Glycine L. Kedelai merupakan tanaman semusim, tanaman tegak dengan tinggi 40-90 cm, bercabang, memiliki daun tunggal dan daun bertiga, bulu pada daun dan polong tidak terlalu padat, umur tanaman antara 72-90 hari dan memiliki akar tunggang. Kedelai introduksi umumnya tidak memiliki atau memiliki sangat sedikit percabangan (Adie dan Krisnawati 2007).
Bunga
Kedelai merupakan tanaman menyerbuk sendiri yang bersifat kleistogami. Periode perkembangan vegetatif bervariasi tergantung pada varietas dan keadaan lingkungan, panjang hari dan suhu. Tanaman memasuki fase reproduktif saat tunas aksiler berkembang menjadi kelompok bunga dengan 2 hingga 35 kuntum bunga setiap kelompok. Ada dua tipe pertumbuhan batang dan permulaan pembungaan pada kedelai. Tipe pertama adalah indeterminit, yaitu tunas terminal melanjutkan fase vegetatif selama pertumbuhan. Tipe kedua adalah determinit dimana pertumbuhan vegetatif tunas terminal terhenti ketika terjadi pembungaan. Periode berbunga dipengaruhi oleh waktu tanam, berlangsung 3-5 minggu. (Adie dan Krisnawati 2007).
Perkembangan Polong
Jumlah polong bervariasi mulai 2-20 dalam satu pembungaan dan lebih 400 dalam satu tanaman. Satu polong berisi 1-15 biji, namun pada umumnya berisi 2-3 biji per polong. Polong masak berwarna kuning muda sampai kuning kelabu, coklat, atau hitam. Panjang polong maksimum dicapai 20-25 hari setelah berbunga. Lebar dan tebal polong maksimum dicapai sekitar 30 hari setelah berbunga. Periode pengisian biji (seed filling period) pada kedelai merupakan fase paling kritis dalam pencapaian hasil optimal. Pada fase tersebut terjadinya kekurangan atau kelebihan air, serangan hama atau penyakit, dan sebagainya akan berpengaruh buruk pada proses pengisian biji (Adie dan Krisnawati 2007).
Biji
Fase Tumbuh
Adie dan Krisnawati (2007) menjelaskan penentuan waktu perlakuan agronomis berdasarkan umur tanaman dapat memberikan hasil yang berbeda dibandingkan yang berdasarkan fase tumbuh karena setiap varietas kedelai memiliki fase tumbuh yang berbeda. Selain ditentukan oleh varietas.
Fehr dan Caviness (1977) menjelaskan karakteristik pertumbuhan tanaman kedelai fase vegetatif dan fase generatif pada Tabel 1 dan 2, serta rentang waktu tumbuh tanaman setiap fase yang disajikan pada Tabel 3.
Tabel 1 Karakteristik fase tumbuh vegetatif pada tanaman kedelai Sandi
fase Fase Pertumbuhan Keterangan
Ve Kecambah Tanaman baru muncul di atas tanah
Vc Kotiledon
Daun keping (kotiledon) terbuka dan dua daun tunggal di atasnya juga mulai terbuka
V1 Buku kesatu
Daun tunggal pada buku pertama telah berkembang penuh, dan daun berhurufi tiga pada buku di atasnya telah terbuka berkembang penuh, dan daun pada buku kelima telah terbuka
V4 Buku keempat
Daun berangkai tiga pada buku keempat telah berkembang penuh, dan daun pada buku kelima telah terbuka
Vn Buku ke-n Daun berangkai tiga pada buku ke-n telah berkembang penuh
Respon Kedelai Terhadap Lingkungan Tumbuh
Suhu berinteraksi dengan panjang penyinaran (photo period) dalam menentukan waktu berbunga dan pembentukan polong. Suhu yang tinggi berakibat pada aborsi polong, sebaliknya suhu di bawah 15 °C menghambat pembentukan polong. Suhu di atas 30 °C berpengaruh negatif terhadap kualitas biji dan daya tumbuh benih. Suhu di atas 27 °C kurang optimum untuk kualitas biji sebagai benih, berkaitan dengan laju pengisian dan pemasakan biji yang kurang optimal (Sumarno dan Manshuri 2007).
Tabel 2 Karakteristik fase tumbuh reproduktif pada tanaman kedelai Sandi
fase Fase Pertumbuhan Keterangan
R1 Mulai berbunga Terdapat satu bunga mekar pada batang sepanjang 5 mm pada batang utama R4 Polong berkembang menunjukkan warna matang (berwarna abu-abu atau kehitaman)
R8 Polong matang penuh 95% telah matang (kuning kecoklatan atau kehitaman)
Tabel 3 Rentang waktu dari fase tumbuh ke fase tumbuh selanjutnya
Fase tumbuh Rata-rata waktu
(hari)
yang berdampak pada fotosintesis tanaman. Ukuran biji, rata-rata pertumbuhan biji dan lama pengisian biji merupakan salah satu komponen hasil yang dipengaruhi faktor lingkungan dan dibatasi oleh faktor karakteristik genetik kultivar (Soedradjad dan Avivi 2005).
Pertumbuhan kedelai akan terhambat jika ketinggian tempat lebih dari 750 m dpl namun masih berproduksi baik pada ketinggian 110 m dpl. Kedelai dapat tumbuh baik pada berbagai jenis tanah dengan syarat drainase dan aerasi tanah cukup baik. Kecuali untuk tanah (podzolik merah kuning) dan tanah-tanah yang banyak mengandung pasir kwarsa pertumbuhannya kurang baik. Oleh karena itu, perlu diberikan pupuk organik dan kapur pertanian dalam jumlah cukup. Sebaiknya kedelai ditanam pada bulan-bulan yang agak kering tetapi air tanah masih cukup tersedia agar pertumbuhan optimal. Air diperlukan sejak pertumbuhan awal sampai pada periode pengisian polong (Kementan 2013).
Genotipe tanaman yang adaptif terhadap lingkungan tumbuh umumnya mengembangkan strategi adaptasi yang unik untuk mendapatkan unsur hara tertentu dari dalam tanah sedangkan genotipe yang tidak adaptif umumnya mengandalkan pupuk sebagai sumber hara yang siap tersedia (Bertham dan Abimanyu 2011).
Penanaman kedelai pada tanah basah akan menghambat perkecambahan dan pertumbuhan awal karena kekurangan oksigen untuk pertumbuhan biji maupun akar tanaman. Biasanya populasi tanaman yang tumbuh akan berkurang pada tanah-tanah yang kelebihan air. Perbaikan drainase pada tanah-tanah seperti ini akan meningkatkan populasi tanaman, perakaran menjadi lebih baik sehingga tanaman menjadi lebih tegak dan berproduksi meningkat (Irwan 2005).
Kondisi ternaungi membuat tanaman kedelai tumbuh lebih memanjang dibandingkan dengan pada keadaan terbuka. Pemberian naungan pada suatu penambahan kontinu di lapang tidak dapat memperbaiki keadaan pertumbuhan tanaman membuat tanaman menjadi lebih tinggi. Tanaman yang tumbuh lebih dari 1 meter tampak rebah dan tumbuh merayap sehingga tidak menguntungkan pada tanaman kedelai (Agusta dan Santosa 2005).
Intensitas cahaya rendah menyebabkan kepadatan trikoma berkurang. Kondisi ini sangat menguntungkan tanaman kedelai karena jumlah cahaya yang akan direfleksikan menjadi sedikit sehingga daun semakin efisien dalam menyerap cahaya (Muhuria et al. 2006).
Tabel 4 Perbedaan agroklimat dan teknik budidaya di wilayah tropika (Indonesia) dan di wilayah subtropika (Amerika Serikat)
Komponen Wilayah tropika Wilayah subtropika Kesuburan tanah Sedang-subur Sangat subur Lapisan olah tanah Dangkal-sedang Sangat dalam Penyiapan lahan Tanpa olah/minimal Intensif – optimal
Panjang hari 12 jam 14-16 jam
Curah hujan Sering berlebih, kering Hujan rintik-rintik Kesesuaian lahan Sangat beragam Sangat sesuai
Hama Sangat banyak Minimal
Penyakit Sangat banyak Minimal
Rancangan Augmented
Tahap awal pemuliaan tanaman pada umumnya terdapat genotipe dalam jumlah besar tetapi jumlah benih yang tersedia tiap genotipe terbatas sehingga untuk mengatasi masalah ini dan mengoptimalkan sumberdaya yang ada biasanya pemulia menggunakan rancangan perbesaran (rancangan augmented). Rancangan perbesaran digunakan untuk menyaring genotipe-genotipe dalam percobaan pemuliaan (Syukur et al. 2010). Rancangan ini pada genotipe baru yang diuji tidak diulang tetapi genotipe pembanding yang diulang (Federer et al.2001). Scott dan Milliken (1993) menjelaskan pembanding dalam rancangan perbesaran ini digunakan untuk menduga pengaruh blok dan pengaruh lingkungan. Menurut Peterson (1994) tujuan dari rancangan perbesaran yaitu merupakan suatu cara membandingkan antara genotipe yang diuji dengan pembanding dan untuk menyesuaikan hasil genotipe yang berbeda dari blok ke blok.
METODE PENELITIAN
Waktu dan Tempat
Penelitian ini dilaksanakan di kebun percobaan IPB Sawah Baru, Dramaga, Bogor. Penelitian berlangsung pada tanggal 17 Februari sampai 30 Juni 2014.
Bahan dan Alat
Metode Penelitian
Penelitian ini disusun dengan rancangan augmented dalam rancangan acak lengkap (RAL) dengan genotipe kedelai sebagai perlakuan (P) tunggal. Benih kedelai yang digunakan sebagai pembanding (kontrol) adalah kedelai varietas Tanggamus, Anjasmoro, Argo Mulyo, Wilis dan Tachinagaha (varietas Jepang) terdiri dari 5 ulangan sehingga terdapat 150 satuan percobaan. Setiap petak percobaan berukuran 5 m × 2 m dengan jumlah 10 tanaman per baris. Tata letak percobaan terdapat pada Lampiran 3. Model matematika percobaan ini mengikuti model Gomez dan Gomez (2007) :
Yij = µ + τi + εij, dimana i = 1, 2, 3..., 130 ; j = 1, 2, 3, 4, 5 Keterangan :
Yij = respon pengamatan genotipe kedelai ke-i (i = 1, 2,3..,130), ulangan ke-j (j = 1, 2, 3, 4, 5)
µ = rataan umum
τi = pengaruh perlakuan genotipe kedelai ke-i (i =1, 2,3..,130)
εij = pengaruh galat percobaan perlakuan genotipe kedelai ke-i (i = 1, 2,3..,130),
ulangan ke-j (j = 1, 2, 3, 4, 5)
Analisis Data
Data yang diperoleh diuji melalui uji F menggunakan aplikasi SAS dan jika menunjukkan adanya pengaruh nyata maka akan dilanjutkan dengan uji BNT pada taraf � = 5% (pada selang kepercayaan 95%).
Pelaksanaan Penelitian
Persiapan dan pengolahan tanah dilakukan dua minggu penanaman untuk membersihkan gulma. Masing-masing petak percobaan dibuat dengan ukuran 5 m × 2 m dan saluran drainase atau selokan dengan lebar 0.5 meter untuk menghindari genangan air.
Penanaman dilakukan dengan ditugal sebanyak 2 butir/lubang dengan jarak tanam 50 cm × 20 cm. Benih direndam dengan legin selama 4-5 menit dan saat penanaman diberikan karbofuran 5-7 butir pada masing-masing lubang tanam.
Pemupukan dilakukan pada saat penanaman dengan cara dialur di samping barisan tanaman. Dosis pupuk yang digunakan adalah 50 kg/ha urea, 150 kg/ha SP-36 dan 100 kg/ha KCl.
Pengendalian gulma dilakukan secara manual setiap minggu dan pengendalian hama dan penyakit dilakukan secara kimiawi apabila diperlukan.
Pengamatan dimulai pada satu minggu setelah tanam (MST) dengan menghitung daya tumbuh benih dilapang. Penyulaman dilakukan dengan mencabut tanaman kedelai yang tumbuh pada lubang tanam secara hati-hati agar akar utama tidak terputus dan dilakukan pada sore hari.
Pengamatan
Pengamatan dilakukan terhadap 3 tanaman contoh untuk setiap genotipe kedelai. Peubah tanaman yang diamati dilapangan terdiri atas :
1. Persentase tanaman tumbuh : diamati setelah 2 MST,
2. Tinggi tanaman (cm) : diukur dari pangkal batang hingga bagian ujung (titik tumbuh) batang utama pada saat fase R1 dan R7,
3. Umur berbunga (hari) : diamati saat 50% jumlah populasi berbunga, 4. Waktu muncul polong (hari) : diamati pada saat terdapat satu atau lebih
polong sepanjang 5 mm pada batang utama (fase R3),
5. Jumlah buku per tanaman : diamati pada saat fase R1 dan R3, 6. Warna bunga : diamati pada saat fase R1,
7. Waktu perubahan polong menjadi matang (hari): diamati pada fase R7, 8. Waktu panen (hari) : diamati apabila 95% polong telah matang (kuning
kecoklatan atau kehitaman),
9. Bobot kering tajuk (g) : Bagian tajuk tanaman dipisahkan dari akar dengan cara memotong pada bagian pangkal batang lalu tajuk tersebut dioven pada suhu 80 °C selama 3 hari (kadar air 0%),
10.Jumlah polong per tanaman: diamati pada saat setelah panen, 11.Jumlah cabang per tanaman : diamati pada saat fase R8, 12.Warna biji : diamati pada saat setelah panen,
13.Kadar air benih (%) : diamati dengan menggunakan alat moisture tester agar benih tidak rusak,
14.Bobot 100 biji (g) : adalah bobot biji yang dihasilkan setelah dipanen dan dijemur di bawah sinar matahari selama 3-5 hari (kadar air 14%),
15.Persentase kondisi biji yaitu menghitung benih penuh/sempurna, keriput, rusak dan unfertile,
16.Ukuran biji : diamati setelah menimbang bobot 100 biji.
HASIL DAN PEMBAHASAN
Kondisi Umum Penelitian
yaitu terdapat miselium cendawan pada polong. Berikut disajikan hasil analisis sifat kimia dan fisik tanah kebun percobaan IPB Sawah Baru pada Tabel 5. Tabel 5 Hasil analisis sifat fisik dan kimia tanah kebun IPB Sawah Baru
pH 1:1 Sumber : Laboratorium Departemen ITSL IPB 2014
Selain itu, kriteria kesesuaian agroklimat untuk pertanaman kedelai adalah pH 6.0-6.5, unsur K sedang-tinggi, Ca dan Mg sedang, serta kejenuhan liat sekitar 36-43 (Sumarno dan Manshuri 2007). Berdasarkan kriteria penilaian sifat kimia dan fisik tanah menurut Hardjowigeno (1995), maka data hasil analisis sifat kimia dan fisik tanah kebun percobaan IPB Sawah Baru pada Tabel 5 termasuk dalam kriteria rendah-sedang dengan tingkat keasamaan tanah (pH) agak masam. Oleh karena itu, lahan percobaan tersebut layak digunakan untuk penelitian budidaya kedelai walaupun curah hujan tinggi karena penelitian ini bertujuan untuk mempelajari respon genotipe kedelai dari wilayah iklim yang berbeda-beda pada kawasan Indonesia yang tropika basah terhadap pertumbuhan dan produksinya.
Keragaan Karakter Agronomi dan Hasil Genotipe-Genotipe Kedelai
Tabel 6 Rekapitulasi hasil sidik ragam berbagai karakter genotipe tanaman kedelai
Persentase biji penuh/sempurna tn tn tn a)26.49
Persentase biji keriput ** tn ** a)21.96
Persentase biji rusak tn tn tn a)28.02
Persentase biji unfertile tn tn tn b)23.95
** = Berbeda sangat nyata pada taraf uji 1%, * = Berbeda nyata pada taraf uji 5%, tn = Tidak berpengaruh nyata, G = Genotipe, K = Kontrol (varietas pembanding), G*V = Interaksi, a) = Data ditransformasikan dengan log(x+2), b) = Data ditransformasikan dengan log(x+3).
Persentase Tumbuh
Pengamatan terhadap persentase daya tumbuh benih di lapangan dilakukan pada 2 MST. Persentase daya tumbuh benih di lapangan disajikan pada Tabel 7 dengan memberikan nomor kode lapangan (NKL) yang dijelaskan pada Lampiran 1 dan 2. Menurut Koesrini dan William (2004) hasil observasi rata-rata daya tumbuh kedelai di lahan sulfat masam pada 2 MST cukup tinggi yaitu antara 89.8-97.9%. Oleh karena itu, dapat dilihat pada Tabel 7 dibawah ini bahwa ada 99 genotipe yang memiliki persentase tumbuh yang tinggi yaitu diatas 80% dan 19 genotipe yang memiliki persentase tumbuh rendah yaitu dibawah 50%.
Tabel 7 Persentase daya tumbuh dari 125 genotipe kedelai yang diuji dan kelima Angka-angka di dalam tabel merupakan nomor kode lapangan (NKL)
Tabel 7 menunjukkan bahwa genotipe dengan nomor 27, 31, 54, 61, 70, 116, dan 117 merupakan genotipe yang paling tidak adaptif pada lingkungan tropika basah karena tidak ada satu pun tanaman yang hidup di lapangan. Beberapa faktor penyebab hal tersebut adalah rendahnya viabilitas benih, benih yang doman, dan adanya perbedaan lingkungan tumbuh asalnya terutama akibat suhu, kelembaban udara, dan intensitas cahaya matahari. Kartahadimaja et al. (2013) menyatakan bahwa gejala turunnya viabilitas benih akibat penyimpanan pada periode 4 tahun setelah diuji di laboratorium dan di lapangan terjadi karena benih tetap berkecambah, tetapi tidak normal, atau karena benih tidak tumbuh sama sekali atau karena benih sudah mati. Beberapa genotipe kedelai dari wilayah subtropika yang memiliki persentase tumbuh yang rendah (<50%) seperti genotipe dengan nomor 27, 29, 30, 50, 55, dan 61 yang berasal dari Korea, genotipe dengan nomor 28, 31, dan 54 yang berasal dari Republik Rakyat Cina (RRC), genotipe dengan nomor 70 dan 80 yang berasal dari Pakistan dan Nepal. Genotipe dari wilayah subtropika tersebut memiliki persentase tumbuh yang rendah diduga akibat berbedanya iklim di lingkungan tumbuh percobaan dengan iklim negara asal. Genotipe dengan nomor 76 yang berasal dari Thailand juga memiliki persentase tumbuh yang rendah meskipun berasal dari wilayah tropika, diduga diakibatkan rendahnya viabilitas benih sehingga daya berkecambah benih di lapangan rendah. Selain itu, terdapat satu genotipe yang mengalami dormansi benih dengan NKL 84 yang berasal dari Nepal. Genotipe tersebut mengalami dormansi benih akibat kulit benih yang tebal walaupun sebelumnya telah diberi perlakuan dengan cara melakukan pengamplasan.
ditimbulkan akibat lalat bibit kedelai adalah terdapat bintik cokelat pada keping biji dan daun sehingga tanaman menjadi layu, mengering, dan akhirnya mati. Serangan yang ditimbulkan akibat lalat bibit kedelai sekitar 10%. Berikut disajikan tampilan daya tumbuh benih di lapangan, tampilan benih yang dorman dan serangan lalat bibit pada Gambar 1, 2, dan 3.
Umur Berbunga (Fase R1)
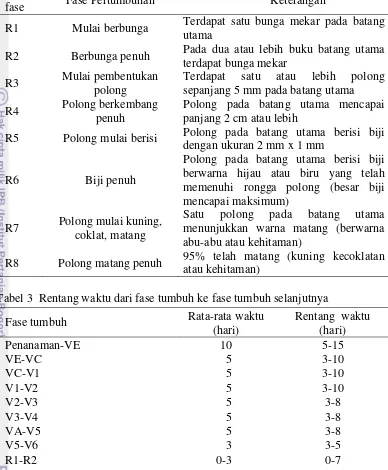
Pengamatan umur berbunga dilakukan pada saat 28-52 HST. Dapat dilihat pada Gambar 4 bahwa Tachinagaha merupakan varietas pembanding yang berbunga paling cepat yaitu pada saat 31 HST sedangkan Wilis merupakan varietas pembanding yang berbunga paling lama yaitu pada saat 45 HST sehingga kedua varietas ini menjadi acuan untuk membandingkan genotipe-genotipe yang diuji berbunga cepat atau lambat. Menurut Susanto dan Sundari (2011) rentang umur berbunga tanaman kedelai pada lingkungan tidak tenaungi berkisar antara 36-48 hari HST dengan rata-rata 41 HST. Dapat dilihat pada Gambar 4 bahwa ada 21 genotipe yang berbunga paling cepat terjadi pada saat 28 HST yaitu Enrei, KOUKOU 6514-2, Athow, LD003309, TIEFENG 18, GmWMC001, GmWMC006, GmWMC015, GmWMC019, GmWMC024, GmWMC035, GmWMC036, GmWMC038, GmWMC042*2, GmWMC046, GmWMC070, GmWMC071, GmWMC073, GmWMC084, GmWMC086, GmWMC107, GmWMC113, GmWMC119, GmWMC123, dan GmWMC159 yang tergolong kedalam tipe determinit. Sebaliknya terdapat 1 genotipe yang berbunga paling lama terjadi pada saat 52 HST yaitu GmWMC191 yang tergolong kedalam tipe indeterminit. Oleh karena itu, data hasil penelitian ini menunjukkan bahwa beberapa genotipe tersebut memiliki umur berbunga yang lebih cepat dan lebih lama dari literatur. Genotipe yang berbunga paling cepat tersebut sebagian besar merupakan kedelai yang berasal dari wilayah subtropika yaitu Jepang, Korea, dan Republik Rakyat Cina. Hal ini sesuai dengan Sumarno dan Manshuri (2007) bahwa varietas kedelai dari wilayah subtropika yang sesuai untuk panjang hari 14-16 jam apabila ditanam di Indonesia yang panjang harinya 12 jam maka akan mempercepat pembungaan pada umur 20-22 hari walaupun batang tanaman masih pendek, tanaman sudah berbunga. Berikut disajikan umur berbunga (fase R1) dari genotipe yang diuji dan varietas pembanding (kontrol) pada Gambar 4.
Hama yang menyerang tanaman pada saat fase R1 ini adalah ulat penggulung daun (Lamprosema indicata). Gejala yang timbul akibat ulat penggulung daun adalah adanya daun-daun yang tergulung menjadi satu sehingga bila gulungan dibuka dapat dilihat ulat atau kotorannya yang berwarna coklat hitam. Serangan yang ditimbulkan akibat ulat penggulung daun sekitar 15%.
Penyakit yang menyerang tanaman pada saat fase R1 adalah penyakit rebah semai. Marwoto dan Hardaningsih (2007) menjelaskan gejala yang timbul akibat penyakit ini adalah tanaman menjadi mendadak layu. Daun-daun yang terinfeksi membentuk bercoklat bulat berwarna cokelat tua kemudian mengering.
Gulma dominan yang tumbuh pada saat tanaman mengalami fase R1 adalah gulma dari golongan rumput seperti Eleusine indica, Cynodon dactylon, Digitaria ciliaris dan Cyperus sp. dari golongan gulma teki. Gulma tersebut cepat mengalami pertumbuhan karena curah hujan yang cukup tinggi. Berikut disajikan tampilan tanaman pada fase R1, serangan ulat, serangan ulat dan gulma pada
Waktu Muncul Polong (Fase R3)
Pengamatan waktu muncul polong dimulai pada saat 35 HST dan berakhir pada saat 55 HST. Berikut disajikan waktu muncul polong (fase R3) dari genotipe yang diuji dan varietas pembanding (kontrol) pada Gambar 5.
Gambar 9 Waktu muncul polong (fase R3)
Gambar 9 menunjukkan bahwa Tachinagaha merupakan varietas pembanding yang paling cepat membentuk polong yaitu pada saat 35 HST sedangkan Anjasmoro merupakan varietas pembanding yang paling lama membentuk polong yaitu pada saat 50 HST karena benih Anjasmoro yang digunakan memiliki viabilitas yang rendah akibat umur simpan yang lama. Oleh karena itu, kedua varietas ini menjadi acuan untuk membandingkan genotipe-genotipe yang diuji berpolong cepat atau lama. Berdasarkan hasil pengamatan, rentang waktu dari fase R1 menuju fase R3 adalah 6-10 hari sehingga waktu muncul polong pada penelitian ini menunjukan waktu yang lebih cepat dibandingkan Fehr dan Caviness (1977) yang menyatakan bahwa rentang waktu fase R1 sampai R3 sekitar 15 hari. Dapat dilihat pada Gambar 9 bahwa ada 26 genotipe yang membentuk polong paling cepat yang terjadi pada 35 HST yaitu Enrei, KOUKOU 6514-2, Athow, LD003309, GmWMC001, GmWMC006, GmWMC015, GmWMC019, GmWMC024, GmWMC035, GmWMC036, GmWMC038, GmWMC042*2, GmWMC046, GmWMC070, GmWMC071, GmWMC073, GmWMC084, GmWMC086, GmWMC113, GmWMC107, GmWMC119, GmWMC123, dan GmWMC159. Genotipe-genotipe tersebut merupakan genotipe yang paling cepat berbunga sedangkan 2 genotipe yang membentuk polong paling lama yang terjadi pada 55 HST yaitu GmWMC173 dan GmWMC191. GmWMC173 dan GmWMC191 merupakan genotipe yang paling lama waktu munculnya bunga 50% dari populasi sehingga hal ini berkorelasi dengan lamanya pembentukan polong. Genotipe yang membentuk polong paling cepat tersebut sebagian besar berasal dari wilayah subtropika yaitu Jepang, Korea, Republik Rakyat Cina dan Nepal sedangkan genotipe yang paling lama membentuk polong adalah yang berasal dari Taiwan dan Filipina.
Hama yang menyerang pada masa pembentukan polong adalah ulat penggulung daun (Lamprosema indicata) dan kepik polong (Riptortus linearis). Serangan yang ditimbulkan akibat kepik polong adalah pertumbuhan polong dan biji kempes lalu mengering kemudian polong gugur. Berikut disajikan tampilan tanaman pada fase R3 dan serangan kepik polong pada Gambar 10 dan 11.
Waktu Perubahan Polong Menjadi Kuning, Coklat, Matang (Fase R7)
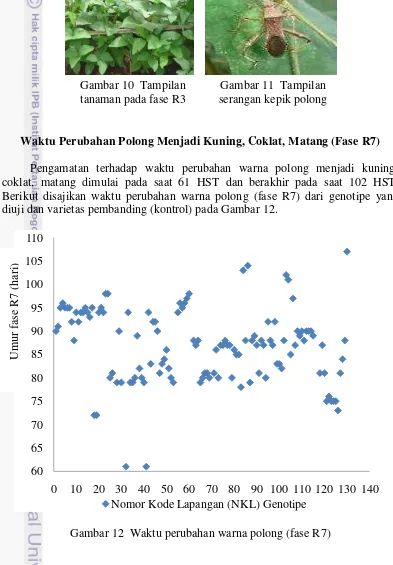
Pengamatan terhadap waktu perubahan warna polong menjadi kuning, coklat, matang dimulai pada saat 61 HST dan berakhir pada saat 102 HST. Berikut disajikan waktu perubahan warna polong (fase R7) dari genotipe yang diuji dan varietas pembanding (kontrol) pada Gambar 12.
Gambar 12 Waktu perubahan warna polong (fase R7) 60
65 70 75 80 85 90 95 100 105 110
0 10 20 30 40 50 60 70 80 90 100 110 120 130 140 Nomor Kode Lapangan (NKL) Genotipe
Gambar 10 Tampilan tanaman pada fase R3
Gambar 11 Tampilan serangan kepik polong
Umur
f
ase
R
7 (h
Gambar 12 menunjukkan bahwa Tanggamus merupakan varietas pembanding yang paling cepat mengalami fase R7 yaitu pada saat 85 HST sedangkan Anjasmoro merupakan vaietas pembanding yang paling lama mengalami fase R7 terjadi pada saat 97 HST. Oleh karena itu, kedua varietas ini menjadi acuan untuk membandingkan genotipe-genotipe yang diuji mengalami fase R7 tercepat dan terlama. Berdasarkan hasil pengamatan, rentang waktu dari fase R3 menuju fase R7 adalah 26-47 hari sehingga waktu perubahan polong menjadi matang pada hasil penelitian ini lebih cepat dibandingkan Fehr dan Caviness (1977) yang menyatakan bahwa rentang waktu fase R3 sampai R7 berkisar antara 36 sampai 91 hari. Dapat dilihat pada Gambar 11 bahwa ada 5 genotipe yang mengalami fase R7 paling cepat terjadi pada saat 61, 72 dan 73 HST yaitu GmWMC019, GmWMC042*2, 317 RINGGIT, SUMBING dan M200-13-47-7 sedangkan 3 genotipe yang mengalami fase R7 paling lama terjadi pada 103, 104 dan 107 HST yaitu GmWMC115, GmWMC192, GmWMC191. GmWMC019 dan GmWMC042*2 merupakan genotipe yang mengalami fase tumbuh paling cepat dilihat dari waktu pada saat fase R1, R3 sampai R7 sedangkan GmWMC191 merupakan genotipe yang mengalami fase tumbuh paling lama dilihat dari waktu pada saat fase R1, R3 sampai R7.
Hama yang menyerang pada saat tanaman mengalami fase R7 tidak berbeda pada saat fase R3 yaitu ulat penggulung daun (Lamprosema indicata) dan kepik polong (Riptortus linearis). Serangan hama yang diakibatkan ulat penggulung daun dan kepik polong tersebut sekitar 50% sehingga dilakukan penyemprotan insektisida 2 kali seminggu secara rutin.
Umur Panen (Fase R8)
Pengamatan umur panen dimulai pada saat 66 HST dan berakhir pada saat 112 HST. Berikut disajikan umur panen (fase R8) dari genotipe yang diuji dan varietas pembanding (kontrol) pada Gambar 15.
Gambar 15 Umur panen (fase R8)
Berdasarkan data dari fase R1 sampai R8 dapat dilihat bahwa Tachinagaha merupakan varietas pembanding yang mengalami masa reproduktif paling lama, Argo Mulyo merupakan varietas pembanding yang mengalami dari pembentukan polong sampai polong matang penuh (fase R3-R7) paling cepat. Tanggamus tergolong varietas pembanding yang mengalami waktu perubahan polong menjadi kuning, coklat, matang sampai polong matang penuh (fase R7-R8) yang cepat. Wilis merupakan varietas pembanding yang mengalami masa vegetatif paling lama. Anjasmoro merupakan varietas pembanding yang mengalami fase pertumbuhan paling lama saat umur berbunga sampai polong matang penuh (fase R1-R8). Hal ini disebabkan karena benih yang digunakan memiliki viabilitas rendah sehingga mempengaruhi pertumbuhan tanaman di lapangan.
Gambar 15 menunjukkan bahwa Tanggamus dan Argo Mulyo merupakan varietas pembanding yang paling cepat mengalami fase R8 terjadi pada 94 HST dan Anjasmoro merupakan varietas pembanding yang paling lama mengalami fase R8 terjadi pada 105 HST. Oleh karena itu, kedua varietas ini menjadi acuan untuk membandingkan genotipe-genotipe yang diuji mengalami fase R8 tercepat dan terlama. Berdasarkan data hasil pengamatan dapat dilihat bahwa semua varietas pembanding yang digunakan pada penelitian ini yaitu Anjasmoro, Tanggamus, Argo Mulyo, dan Wilis memiliki umur panen yang lebih lama dari deskripsi varietas. Menurut Adisarwanto et al. (2007) umur panen 1-2 hari lebih lama dari deskripsi varietas menunjukkan tingkat kadar airnya rendah dan mempunyai vigor diatas 95%.
Tambaguro 100, dan Pangrango. Dapat dilihat bahwa ada 2 genotipe yang selalu mengalami fase pertumbuhan paling cepat mulai dari fase R1 sampai fase R8 yaitu GmWMC019 dan GmWMC042*2 yang berasal dari Korea Utara dan Republik Rakyat Cina sedangkan 1 genotipe yang selalu mengalami fase pertumbuhan paling lama mulai dari fase R1 sampai fase R8 yaitu GmWMC191 yang berasal dari Filipina. Hal ini menunjukkan bahwa pada beberapa genotipe semakin cepat umur berbunga maka akan semakin cepat pula umur panen. Sebaliknya jika semakin lama umur berbunga maka akan semakin lama pula umur panennya. Hal ini sesuai dengan Hakim (2012) bahwa koefisien korelasi antara umur berbunga dengan umur polong masak adalah nyata. Adie dan Krisnawati (2007) menjelaskan bahwa umur tanaman kedelai dikelompokkan menjadi genjah (< 80 hari), sedang (80-85 hari), dan dalam (>85 hari). Oleh karena itu, GmWMC019 dan GmWMC042*2 tergolong kedelai berumur genjah sedangkan GmWMC191 tergolong kedelai berumur dalam. Berikut disajikan tampilan tanaman pada fase R8 dan kegiatan panen pada Gambar 16 dan 17.
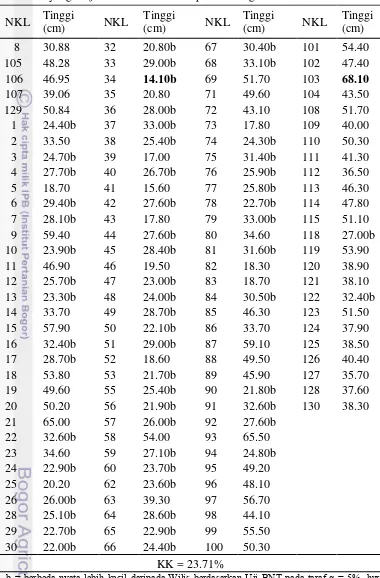
Tinggi Tanaman pada Fase R1
Hakim (2012) menjelaskan varietas kedelai yang batangnya tinggi cenderung mempunyai jumlah cabang, jumlah buku subur, dan jumlah polong per tanaman yang lebih banyak dan bobot brangkasan lebih tinggi daripada varietas yang berbatang pendek. Selain itu, genotipe kedelai yang batangnya tinggi berpotensi memberikan bobot biji dan hasil yang tinggi.
Data tinggi tanaman rata-rata pada fase R1 disajikan pada Tabel 8. Berdasarkan uji lanjut BNT, varietas pembanding yang terbaik adalah Wilis dengan NKL 129. Dapat dilihat pada Tabel 8 bahwa GmWMC022 dengan NKL 34 merupakan genotipe yang paling rendah dan nyata lebih rendah dibandingkan Wilis sedangkan GmWMC191 dengan NKL 103 merupakan genotipe paling tinggi namun tidak berbeda nyata lebih tinggi dibandingkan Wilis. Berdasarkan data hasil pengamatan, GmWMC022 merupakan tanaman yang berumur pendek (genjah) sedangkan GmWMC191 merupakan tanaman yang berumur dalam. Selain itu, sebagian besar genotipe yang berasal dari Jepang memiliki nilai tengah tinggi tanaman yang rendah dan berumur pendek. Hal ini menunjukkan bahwa pada beberapa genotipe semakin rendah tanaman maka fase pertumbuhan akan semakin cepat. Sebaliknya jika semakin tinggi tanaman maka akan semakin lama pula fase pertumbuhannya dan hasil ini sesuai dengan Hakim (2012) yang menjelaskan bahwa koefisien korelasi antara umur polong masak dengan tinggi tanaman adalah nyata.
Gambar 16 Tampilan tanaman pada fase R8
Tabel 8 Tinggi tanaman rata-rata pada fase R1 dari 118 genotipe kedelai yang diuji dan kelima varietas pembanding
NKL Tinggi
b = berbeda nyata lebih kecil daripada Wilis berdasarkan Uji BNT pada taraf α = 5%, huruf
Tinggi Tanaman pada Fase R7
Pengukuran tinggi tanaman pada fase R7 bertujuan membandingkan tinggi tanaman pada fase R1 dengan R7 sehingga dapat dilihat tipe pertumbuhan tanaman dari masing-masing genotipe. Data tinggi tanaman pada fase R7 disajikan pada Tabel 9. Berdasarkan uji lanjut BNT, varietas pembanding (kontrol) yang terbaik adalah Wilis dengan NKL 129. Dapat dilihat pada Tabel 9 bahwa GmWMC022 yang berasal dari Korea Utara dengan NKL 34 merupakan genotipe yang paling rendah dan nyata lebih rendah dibandingkan Wilis sedangkan GmWMC183 yang berasal dari Taiwan dengan NKL 98 merupakan genotipe yang paling tinggi dan nyata lebih tinggi dibandingkan Wilis.
Menurut Adie dan Krisnawati (2007) perbedaan pertumbuhan vegetatif pada tipe determinit dan indeterminit adalah pada tipe determinit, pertumbuhan vegetatif akan berhenti setelah berbunga (fase R1) sedangkan tipe indeterminit, pertumbuhan vegetatif akan berlanjut setelah berbunga. Selain itu, varietas kedelai yang ada di Indonesia pada umumnya bertipe tumbuh determinit. Berdasarkan hasil pengamatan tinggi tanaman rata-rata pada fase R1 dan R7 menunjukkan bahwa GmWMC022 tergolong tipe determinit sedangkan GmWMC183 tergolong tipe indeterminit. Sebagian besar genotipe yang diuji pada penelitian ini memiliki tipe pertumbuhan determinit, dan hanya ada 3 genotipe yang memiliki tipe pertumbuhan indeterminit. Oleh karena itu, dapat dilihat bahwa sebagian besar genotipe yang diuji tidak lagi mengalami pertumbuhan vegetatif setelah berbunga. Ketiga genotipe yang memiliki tipe pertumbuhan indeterminit tersebut merupakan genotipe yang berasal dari Taiwan, dan Nepal dengan NKL 87, 98, dan 104. Genotipe tersebut mengalami pertambahan tinggi yang sangat pesat pada saat fase pertumbuhan R7.
Jumlah Buku pada Fase R1
Berdasarkan Hakim (2012) pada penelitian sebelumnya mengenai korelasi jumlah buku terhadap tinggi tanaman, jumlah cabang, jumlah polong per tanaman dan bobot brangkasan sehingga perhitungan jumlah buku tanaman kedelai perlu dilakukan pada penelitian ini.
Tabel 10 Jumlah buku rata-rata pada fase R1 dari 118 genotipe kedelai yang diuji
b = berbeda nyata lebih kecil daripada Tanggamus berdasarkan Uji BNT pada taraf α = 5%, huruf yang dicetak tebal dalam tabel menandakan genotipe yang memiliki nilai tengah tertinggi dan terendah, data ditransformasikan dengan log(x+2), NKL = Nomor kode lapangan
Jumlah Buku pada Fase R8
perhitungan jumlah buku pada fase R8 perlu dilakukan untuk menentukan jumlah buku setelah berbunga dan membandingkannya dengan jumlah buku pada fase R1 sehingga dapat ditentukan tipe pertumbuhan tanaman dari masing-masing genotipe.
Data jumlah buku pada fase R8 disajikan pada Tabel 11. Berdasarkan uji lanjut BNT, varietas pembanding (kontrol) yang terbaik adalah Tanggamus dengan NKL 105. Dapat dilihat pada Tabel 11 bahwa GmWMC115 dengan NKL 60 yang berasal dari Amerika Serikat merupakan genotipe yang memiliki jumlah buku tanaman pada fase R8 yang paling sedikit dan berbeda nyata lebih rendah dibandingkan Tanggamus sedangkan GmWMC192 dengan NKL 104 yang berasal dari Nepal merupakan genotipe yang memiliki jumlah buku pada fase R8 yang paling banyak dan berbeda nyata lebih tinggi dibandingkan Tanggamus. Berdasarkan data hasil pengamatan bahwa GmWMC115 tidak mengalami pertambahan jumlah buku tanaman yang besar dari fase R1 sampai fase R8 sedangkan GmWMC192 mengalami pertambahan jumlah buku tanaman yang pesat dari fase R1 sampai fase R8. Menurut Adie dan Krisnawati (2007) perbedaan jumlah buku setelah berbunga pada tipe determinit dan tipe indeterminit adalah tipe determinit, jumlah buku tanaman setelah berbunga tidak mengalami pertambahan sedangkan tipe indeterminit, jumlah buku tanaman setelah berbunga mengalami pertambahan. Oleh karena itu, GmWMC115 tergolong kedalam tipe determinit sedangkan GmWMC192 tergolong kedalam tipe indeterminit.
Jumlah Cabang per Tanaman
Data jumlah cabang per tanaman disajikan pada Tabel 12. Berdasarkan uji lanjut BNT, varietas pembanding (kontrol) yang terbaik adalah Wilis dengan NKL 129. Dapat dilihat pada Tabel 12 bahwa terdapat 2 genotipe yaitu GmWMC123*2 dan GmWMC169 dengan NKL 65 dan 90 yang berasal dari Nepal dan Republik Rakyat Cina merupakan genotipe yang memiliki cabang paling sedikit dan berbeda nyata lebih rendah dibandingkan Wilis sedangkan SUMBING dengan NKL 19 yang berasal dari Indonesia merupakan genotipe yang memiliki cabang yang paling banyak dan berbeda nyata lebih tinggi dibandingkan Wilis.
memiliki daya hasil lebih tinggi dibandingkan kelima varietas pembanding tersebut jika dilihat dari jumlah cabang yang sangat banyak
Tabel 12 Jumlah cabang rata-rata dari 118 genotipe kedelai yang diuji dan
a= Berbeda nyata lebih besar daripada Wilis dan b = berbeda nyata lebih kecil daripada Wilis
Bobot Kering Tajuk
Hakim (2012) menjelaskan pada penelitian sebelumnya bahwa bobot kering tajuk berkorelasi terhadap tinggi tanaman, jumlah buku, jumlah cabang, dan jumlah polong per tanaman sehingga pengamatan bobot kering tajuk tanaman kedelai pada penelitian ini perlu dilakukan.
Data bobot kering tajuk rata-rata disajikan pada Tabel 13. Berdasarkan uji lanjut BNT, varietas pembanding (kontrol) yang terbaik adalah Tanggamus dengan NKL 105. Dapat dilihat pada Tabel 13 bahwa GmWMC001 dengan NKL 25 yang berasal dari Swedia merupakan genotipe yang memiliki bobot kering tajuk paling rendah dan berbeda nyata lebih rendah dibandingkan Tanggamus sedangkan GmWMC191 dengan NKL 103 yang berasal dari Filipina merupakan tanaman yang memiliki bobot kering tajuk yang paling tinggi dan berbeda nyata lebih tinggi dibandingkan Tanggamus. Berdasarkan data hasil pengamatan dapat dilihat bahwa GmWMC191 memiliki tinggi tanaman, jumlah buku, dan jumlah cabang yang paling besar sehingga karakter tersebut yang mempengaruhi bobot kering tajuk rata-rata GmWMC191 paling tinggi dibandingkan genotipe lainnya. Hal ini sesuai dengan penelitian Hakim (2012) bahwa varietas kedelai yang batangnya tinggi cenderung mempunyai jumlah cabang, jumlah buku subur, dan jumlah polong per tanaman yang lebih banyak dan bobot brangkasan lebih tinggi daripada varietas yang berbatang pendek. Berikut data bobot kering tajuk rata-rata yang disajikan pada Tabel 13.
Jumlah Polong per Tanaman
Berdasarkan uji lanjut BNT, varietas pembanding (kontrol) yang terbaik adalah Wilis dengan NKL 129. Dapat dilihat pada Tabel 14 bahwa Tambaguro 100 yang berasal dari Jepang dengan NKL 24 merupakan genotipe yang memiliki jumlah polong per tanaman paling sedikit dan berbeda nyata lebih rendah dibandingkan Wilis sedangkan GmWMC192 dengan NKL 104 yang berasal dari Nepal memiliki jumlah polong per tanaman yang paling banyak namun tidak berbeda nyata lebih tinggi dibandingkan Wilis. Berdasarkan hasil pengamatan bahwa Wilis merupakan varietas pembanding yang paling banyak memiliki jumlah polong per tanaman namun hasil ini berbeda dengan penelitian Ghulamahdi (2011) yang menjelaskan bahwa varietas Tanggamus menghasilkan jumlah polong lebih terbanyak dibandingkan Wilis, Anjasmoro, dan Slamet pada teknologi budidaya kedelai jenuh air maupun tanpa pengairan. Hal ini juga berbeda diduga akibat adanya perbedaan lingkungan tumbuh.
Tabel 13 Bobot kering tajuk rata-rata dari 118 genotipe kedelai yang diuji dan kelima varietas
a= Berbeda nyata lebih besar daripada Tanggamus dan b = berbeda nyata lebih kecil daripada
Tanggamus berdasarkan Uji BNT pada taraf α = 5%, huruf yang dicetak tebal dalam tabel menandakan
genotipe yang memiliki nilai tengah tertinggi dan terendah, NKL = Nomor kode lapangan
Tabel 14 Jumlah polong rata-rata per tanaman dari 125 genotipe kedelai yang
Berdasarkan data hasil pengamatan dapat dilihat bahwa GmWMC192 merupakan genotipe yang memiliki jumlah buku paling banyak pada saat fase R1 dan R8. Hal ini sesuai dengan penelitian Hakim (2012) bahwa varietas kedelai yang mempunyai jumlah buku subur yang banyak cenderung mempunyai jumlah polong yang tinggi, dan jumlah polong per tanaman berkorelasi positif sangat nyata dengan bobot biji per tanaman.
Ukuran dan Warna Biji
Menurut Hakim (2012) bobot 100 biji menunjukkan korelasi positif nyata dengan bobot biji per tanaman dan berkorelasi sangat nyata dengan indeks panen. Bobot 100 biji merupakan salah satu komponen yang mempengaruhi produksi biji. Karakter bobot 100 biji merupakan perbandingan ukuran secara kuantitatif antara biji dari masing-masing genotipe. Ukuran biji dapat ditentukan setelah menghitung bobot 100 biji. Berikut disajikan pada Tabel 15 pengelompokan biji berdasarkan ukuran dengan nomor kode lapangan (NKL) dari masing genotipe. Tabel 15 Pengelompokan biji berdasarkan ukuran
Ukuran biji
bahwa biji besar memiliki umur panen yang lebih lama dibandingkan biji kedelai yang berukuran kecil.
Data pengelompokan warna biji dari masing-masing genotipe disajikan pada Tabel 16. Karakter warna biji diperlukan untuk menentukan pemanfaatan kedelai. Menurut Wirnas et al. (2012) selama ini kedelai hitam umumnya digunakan sebagai bahan kecap, sedangkan kedelai kuning selain untuk kecap juga dimanfaatkan untuk bahan baku industri tempe, tahu, dan susu. Adie dan Krisnawati (2007) menjelaskan bahwa warna kulit biji kedelai bervariasi dari kuning, hijau, coklat, hitam hingga kombinasi berbagai warna atau campuran. Pigmen kulit biji sebagian besar terletak di lapisan palisade, terdiri dari pigmen antosianin dalam vakuola, klorofil dalam plastisida, dan berbagai kombinasi hasil uraian produk-produk pigmen tersebut. Kombinasi berbagai pigmen yang ada di kulit biji dan kotiledon akan membentuk warna biji yang bermacam-macam pada kedelai. Dapat dilihat pada Tabel 16 bahwa pada penelitian ini lebih banyak menggunakan kedelai kuning dan sebenarnya warna biji tidak menjadi karakter utama di dalam penelitian. Berdasarkan data hasil pengamatan dapat dilihat bahwa warna biji tidak berpengaruh terhadap bobot 100 biji dan ukuran biji.
Tabel 16 Pengelompokan warna biji dari masing-masing genotipe Warna biji
Tabel 17 Persentase biji penuh, keriput, rusak, dan unfertile dari 118 genotipe kedelai yang diuji dan kelima varietas pembanding
Tabel 17 Persentase biji penuh, keriput, rusak, dan unfertile dari 118 genotipe kedelai yang diuji dan kelima varietas pembanding (lanjutan)
NKL Penuh Data persentase biji penuh, biji keriput, dan biji rusak ditransformasikan dengan log(x+2), data persentase biji unfertile ditransformasikan dengan log(x+3), huruf yang dicetak tebal dalam tabel menandakan genotipe yang memiliki nilai tengah tertinggi dan terendah, NKL = Nomor kode lapangan
dengan kondisi biji terbaik karena nilai tengah persentase biji penuh tinggi serta persentase biji keriput, rusak, dan unfertile rendah. Berikut disajikan tampilan persentase kondisi biji dari contoh genotipe kedelai hitam dan kuning pada Gambar 18.
Perhitungan persentase kondisi biji perlu dilakukan untuk mengetahui ketersediaan benih dalam kegiatan penelitian selanjutnya. Selain itu, untuk mengetahui respon tumbuh genotipe yang diuji terhadap lingkungan tumbuh. Adanya hama kepik polong (Riptortus linearis) yang menyerang pada saat terjadinya pembentukan polong sampai polong berubah warna (fase R7-R8) mempengaruhi persentase kondisi biji. Marwoto dan Hardaningsih (2007) menjelaskan bahwa serangan yang diakibatkan oleh kepik polong pada fase perkembangan biji dan pertumbuhan polong menyebabkan polong dan biji kempes, kemudian mengering dan polong gugur. Hal ini yang menyebabkan geotipe yang tidak toleran terhadap serangan kepik polong tersebut mengalami penurunan hasil dan kualitas biji seperti genotipe Tambaguro 100.
Kadar Air Biji
Pengukuran kadar air biji dilakukan untuk mengetahui daya umur simpan biji, jenis kemasan dan ruang simpan benih. Pengukuran kadar air biji pada penelitian ini menggunakan metode tidak langsung dengan memakai alat yang disebut Moisture Tester. Tujuan penggunaan alat ini adalah untuk mendapatkan hasil pengukuran kadar air secara cepat dan tidak merusak benih pada saat pengukuran kadar air karena biji-biji yang dalam jumlah terbatas tersebut akan digunakan untuk kegiatan penelitian selanjutnya dalam waktu dekat. Berikut disajikan data kadar air biji rata-rata pada Tabel 18.
Menurut Indartono (2011) kedelai tergolong benih ortodoks. Kadar air optimum dalam penyimpanan bagi sebagian besar benih ortodoks adalah antara 6-11%. Hal ini sesuai dengan penelitian Tatipata (2007) bahwa kadar air awal 12% kurang tepat digunakan untuk menyimpan benih kedelai. Oleh karena itu, semua biji kedelai tersebut dijemur kembali untuk menurunkan kadar air karena kadar airnya masih diatas 11%.
Tabel 18 Kadar air biji rata-rata dari 118 genotipe kedelai yang diuji dan kelima
huruf yang dicetak tebal dalam tabel menandakan genotipe yang memiliki nilai tengah tertinggi dan terendah, NKL = Nomor kode lapangan
menyebabkan deteriorasi benih kedelai selama penyimpanan namun deteriorasi masih relatif kecil yang ditunjukkan dari daya berkecambah dan vigor benih yang masih di atas 80%. Namun kondisi pada ruang penyimpanan benih dari hasil penelitian ini memiliki suhu 20 °C dan RH 36%. Kondisi tersebut disebabkan karena keterbatasan fasilitas ruang penyimpanan benih.
Karakter kadar air biji berkisar antara 10.90-19.10% sedangkan varietas pembanding memiliki nilai tengah antara 10.62-14.44%. Nilai tengah kadar air biji tidak berbeda nyata dengan varietas pembanding sehingga tidak dilakukan uji lanjut BNT. Tabel 18 menunjukkan bahwa varietas pembanding yang memiliki nilai tengah kadar air paling rendah adalah Anjasmoro sebesar 10.62% sedangkan varietas pembanding yang memiliki nilai tengah kadar air paling tinggi adalah Republik Rakyat Cina dengan nomor 8 yang berasal dari Jepang sebesar 14.44%. GmWMC141 dengan nomor 71 yang berasal dari Indonesia merupakan genotipe yang memiliki nilai tengah kadar air paling rendah sebesar 10.90% sedangkan GmWMC071 dengan nomor 47 yang berasal dari India merupakan genotipe yang memiliki nilai tengah kadar air yang paling tinggi sebesar 19.10%. Berdasarkan data hasil pengamatan dapat dilihat bahwa Tachinagaha memiliki nilai tengah bobot 100 biji sebesar 24.10 gram sedangkan Anjasmoro memiliki nilai tengah bobot 100 biji sebesar 16.88 gram, dan GmWMC071 memiliki nilai tengah bobot 100 biji sebesar 19.33 gram sedangkan GmWMC141 memiliki nilai tengah bobot 100 biji sebesar 11.93 gram. Oleh karena itu, Tachinagaha memiliki bobot biji lebih besar daripada Anjasmoro, dan GmWMC071 memiliki nilai bobot biji lebih besar daripada GmWMC141. Hal ini menunjukkan bahwa bobot biji berkorelasi positif dengan jumlah kadar air biji. Semakin besar bobot biji maka semakin besar kadar air biji. Sebaliknya semakin rendah bobot biji maka semakin rendah kadar air benih. Hal ini sesuai dengan penelitian Hartawan (2013) yang menyatakan bahwa peningkatan cadangan makanan akan meningkatkan bobot biji dan bobot biji berhubungan dengan kadar air. Nilai kadar air biji meningkat sejalan dengan peningkatan kandungan protein, karohidrat, dan lemak. Berikut disajikan tampilan alat dan penggunaan Moisture Tester pada Gambar 19.
Korelasi Antarkarakter
Dapat dilihat pada Tabel 19 bahwa hasil analisis korelasi menunjukkan bahwa umur tanaman pada fase R1 berkorelasi positif sangat nyata terhadap semua karakter pengamatan yaitu umur tanaman pada fase R3, umur tanaman pada fase R7, umur tanaman pada fase R8, tinggi tanaman pada fase R1, tinggi tanaman pada fase R7, jumlah buku pada fase R1, jumlah buku pada fase R8, jumlah cabang, bobot kering tajuk, jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji dengan koefisien korelasi (r) masing-masing 0.961, 0.801, 0.817, 0.791, 0.865, 0.711, 0.646, 0.649, 0.954, 0.648, 0.995, 0.795, dan 0.978. Korelasi antara umur tanaman pada fase R1 dengan umur tanaman pada fase R3 tersebut menunjukkan bahwa genotipe kedelai yang cepat berbunga (R1) cenderung akan lebih cepat berpolong (R3).
Umur tanaman pada fase R3 berkorelasi positif sangat nyata terhadap semua karakter pengamatan yaitu umur tanaman pada fase R7, umur tanaman pada fase R8, tinggi tanaman pada fase R1, tinggi tanaman pada fase R7, jumlah buku pada fase R1, jumlah buku pada fase R8, jumlah cabang, bobot kering tajuk, jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji. Nilai masing-masing koefisien korelasi (r) adalah 0.843, 0.857, 0.774, 0.859, 0.658, 0.857, 0.647, 0.944, 0.626, 0.995, 0.794, dan 0.909. Korelasi antara umur tanaman pada fase R3 dengan umur tanaman pada fase R7 tersebut menunjukkan bahwa genotipe kedelai yang cepat berpolong (R3) cenderung akan lebih cepat mengalami perubahan warna polong menjadi matang (R7).
Umur tanaman pada fase R7 berkorelasi positif sangat nyata terhadap umur tanaman pada fase R8, tinggi tanaman pada fase R1, tinggi tanaman pada fase R7, jumlah cabang, bobot kering tajuk, jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji dengan koefisien korelasi (r) masing-masing 0.995, 0.536, 0.8457, 0.439, 0.863, 0.325, 0.991, 0.667, dan 0.997. Korelasi antara umur tanaman pada fase R7 dengan umur tanaman pada fase R8 tersebut menunjukkan bahwa genotipe kedelai yang cepat mengalami fase tumbuh R7 cenderung akan lebih cepat umur panennya (R8).
Korelasi umur tanaman pada fase R7 berkorelasi positif tetapi tidak nyata dengan jumlah buku pada fase R1 dan R8 (r = 0.316 dan r = 0.316). Hal ini menunjukkan bahwa genotipe kedelai yang memiliki umur fase tumbuh R7 yang cepat memiliki jumlah buku yang sedikit.
Umur tanaman pada fase R8 berkorelasi positif sangat nyata terhadap tinggi tanaman pada fase R1, tinggi tanaman pada fase R7, jumlah cabang, bobot kering tajuk, jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji dengan koefisien korelasi (r) masing-masing 0.542, 0.850, 0.456, 0.872, 0.456, 0.992, 0.643 dan 0.997. Umur tanaman pada fase R7 berkorelasi positif tetapi tidak nyata dengan jumlah buku pada fase R1 dan R8 (r = 0.327, dan r = 0.328). Hal ini menunjukkan bahwa genotipe yang memiliki umur fase R8 yang cepat memiliki jumlah buku yang sedikit.
fase R7 berkorelasi positif sangat nyata terhadap semua karakter yaitu jumlah buku pada fase R1, jumlah buku pada fase R8, jumlah cabang, bobot kering tajuk, jumlah polong, persentase biji penuh, dan kadar air biji. Nilai masing-masing koefisien korelasi dapat dilihat pada Tabel 16. Hal ini menunjukkan bahwa genotipe kedelai yang memiliki batang tinggi cenderung memiliki cabang, jumlah buku, dan jumlah polong per tanaman yang lebih banyak daripada genotipe yang berbatang pendek. Oleh karena itu, hasil data penelitian ini sesuai dengan pernyataan Hakim (2012) pada penelitiannya sebelumnya.
Jumlah cabang berkorelasi positif sangat nyata terhadap bobot kering tajuk, jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji dengan koefisien korelasi masing-masing 0.948, 0.543, 0.924, 0.762, dan 0.899. Hal ini menunjukkan bahwa genotipe kedelai yang bercabang banyak cenderung memiliki bobot kering tajuk yang lebih besar dan jumlah polong yang lebih banyak. Oleh karena itu, hasil data penelitian ini sesuai dengan pernyataan Hakim (2012) pada penelitian sebelumnya.
Bobot kering tajuk berkorelasi positif sangat nyata terhadap jumlah polong, bobot 100 biji, persentase biji penuh, dan kadar air biji dengan koefisien korelasi masing-masing 0.935, 0.974, 0.967, dan 0.946. Hal ini menunjukkan bahwa bobot kering tajuk merupakan salah satu faktor penentu tingginya hasil produksi (jumlah polong, bobot 100 biji, dan persentase biji penuh).
Jumlah polong berkorelasi positif sangat nyata terhadap bobot 100 biji, persentase biji penuh, dan kadar air benih dengan koefisien korelasi masing-masing 0.862, 0.923, dan 0.801. Hal ini menunjukkan bahwa jumlah polong merupakan faktor penentu tingginya persentase biji penuh.
Bobot 100 biji berkorelasi positif sangat nyata terhadap persentase biji penuh dan kadar air benih dengan koefisien korelasi masing-masing 0.929 dan 0.991. Persentase biji penuh berkorelasi positif sangat nyata terhadap kadar air biji dengan koefisien korelasi sebesar 0.913.
SIMPULAN DAN SARAN
Simpulan
Genotipe GmWMC192 yang berasal dari Nepal berdaya hasil tinggi dan beradaptasi baik pada lingkungan tumbuh agroekosistem tropika basah. Hal ini dilihat dari persentase tanaman yang tumbuh di lapangan, tinggi tanaman, jumlah buku, jumlah polong per tanaman, dan persentase biji penuh yang tinggi.
Genotipe GmWMC019 dan GmWMC042*2 yang berasal dari Korea Utara dan Republik Rakyat Cina berumur sangat genjah (66 hari) serta beradaptasi baik pada lingkungan tumbuh agroekosistem tropika basah. GmWMC019 dan GmWMC042*2 merupakan genotipe yang peka terhadap suhu tinggi.
Saran
Perlu dilakukan penelitian lebih lanjut pada waktu musim tanam yang berbeda dan peubah pengamatan dilakukan pada setiap fase.
DAFTAR PUSTAKA
Adie MM, Krisnawati A. 2007. Biologi Tanaman Kedelai. Di dalam : Sumarno, Suyamto, Widjono A, Hermanto, Kasim K, editor. Teknik Produksi dan Pengembangan Kedelai. Bogor (ID): Balai Penelitian dan Pengembangan Pertanian. hlm 45-56.
Adisarwanto T, Subandi, Sudaryono. 2007. Teknologi Produksi Kedelai. Di dalam : Sumarno, Suyamto, Widjono, Hermanto, Kasim K, editor. Teknik Produksi dan Pengembangan Kedelai. Bogor (ID): Balai Penelitian dan Pengembangan Pertanian. hlm 247-248.
Agusta H, Santosa I. 2005. Indeterminasi sekuensi pembungaan dan ketidakmampuan produksi kedelai di lapang akibat penambahan cahaya kontinu pada kondisi terbuka dan ternaungi. Bul.Agron. 33(3):24-32
Anggraeni BW. 2010. Studi morfo-anatomi dan pertumbuhan kedelai (Glycine max (L.) Merr.) pada kondisi cekaman intensitas cahaya rendah [skripsi]. Bogor (ID): Institut Pertanian Bogor.
[BAPPENAS] Badan Perencanaan Pembangunan Nasional 2014. Rencana pembangunan jangka menengah nasional (RPJMN) bidang pangan dan pertanian 2015-2019. [Internet]. [diunduh 2014 Sept 18]. Tersedia pada: http://www.bappenas.go.id.
Bertham, Abimanyu DN. 2011. Mekanisme adaptasi genotipe baru kedelai dalam mendapatkan hara fosfor dari tanah mineral masam. J.Agron.Indonesia 39(1):24-30.