BIOLOGI PEMBUNGAAN DAN
KEBERHASILAN REPRODUKSI
Zingiber spectabile
DAN
Tapeinochilos ananassae
Oleh:
Megaria
A24061455
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
BIOLOGI PEMBUNGAAN DAN
KEBERHASILAN REPRODUKSI
Zingiber spectabile
DAN
Tapeinochilos ananassae
Skripsi sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian pada Fakultas Pertanian Institut Pertanian Bogor
Oleh Megaria A24061455
DEPARTEMEN AGRONOMI DAN HORTIKULTURA
FAKULTAS PERTANIAN
RINGKASAN
MEGARIA. Biologi Pembungaan dan Keberhasilan Reproduksi Zingiber spectabile dan Tapeinochilos ananassae. Dibimbing oleh ENDAH RETNO PALUPI dan DEBORA HERLINA ADRIYANI.
Zingiberaceae merupakan salah satu tanaman hias tropis yang keindahannya mampu menyaingi tanaman hias sub tropis. Kurangnya informasi mengenai karakteristik tanaman serta rendahnya keragaman varietas yang dibudidayakan di Indonesia menjadi kendala dalam pengembangan tanaman Zingiberaceae, khususnya untuk bunga potong. Informasi biologi dan fenologi pembungaan terkait dengan viabilitas polen dan masa reseptif stigma serta sistem perkawinan tanaman sangat diperlukan untuk melakukan persilangan dalam rangka meningkatkan keragaman dan memperbaiki karakter bunga.
Penelitian ini bertujuan untuk mempelajari biologi dan fenologi pembungaan Zingiber spectabile dan Tapeinochilos ananassae serta mempelajari keberhasilan reproduksi dari penyerbukan alami (terbuka) dan penyerbukan buatan, yaitu penyerbukan sendiri dan penyerbukan silang dari Zingiber spectabile dan Tapeinochilos ananassae. Penelitian dilakukan di Balai Penelitian Tanaman Hias (Balithi) Segunung dengan ketinggian 1 100 m dpl pada Februari hingga Agustus 2010.
Bunga Z. spectabile mulai mekar pukul 11.00 dan layu pukul 16.00 pada hari yang sama. Viabilitas polen pada pukul 08.00 masih rendah sekitar 20 % tetapi meningkat terus dan mencapai sekitar 62 % pada pukul 12.00. Setelah itu viabilitas polen mulai menurun. Berdasarkan pengamatan sekresi pada permukaan stigma dan percobaan penyerbukan silang, pukul 10.00-12.00 merupakan waktu yang paling tepat untuk penyerbukan. Z. spectabile termasuk tanaman partially self incompatible (ISI=0.92). Penyerbukan terbuka tidak menghasilkan biji diduga karena rendahnya penyerbukan yang disebabkan oleh terbatasnya serangga penyerbuk. Jumlah biji yang dihasilkan dari penyerbukan sendiri dan penyerbukan silang masing-masing sebesar 24.0±4.5 dan 11.7±6.3 biji/buah.
Kuncup bunga T. ananassae mulai keluar pukul 07.00 dan mekar pada hari yang sama atau keesokan harinya. Persentase polen yang berkecambah sangat rendah. Sekresi di permukaan stigma mulai muncul pada pukul 12.00-14.00. Jumlah biji hasil penyerbukan paling banyak dihasilkan dari penyerbukan pukul 08.00 dengan rata-rata 12.7 biji/buah. Sedangkan penyerbukan pukul 12.00 dan 14.00 tidak menghasilkan biji, sehingga waktu yang tepat untuk penyerbukan adalah pukul 09.00-11.00.
Persentase pembentukan buah T. ananassae relatif rendah. Penyerbukan silang menghasilkan jumlah buah rata-rata 13.3 % dengan 7.3 biji/buah. Penyerbukan sendiri berhasil membentuk 5.5 biji/buah. Berdasarkan data tersebut,
Judul :
BIOLOGI
PEMBUNGAAN DAN KEBERHASILAN
REPRODUKSI
Zingiber spectabile
DAN
Tapeinochilos ananassae
Nama :
MEGARIA
NIM :
A24061455
Menyetujui,
Pembimbing I Pembimbing II
Dr. Ir. Endah Retno Palupi, MSc. Ir. Debora Herlina Adriyani, MS.
NIP: 19580518 198903 2 002 NIP: 19530408 198102 2 001
Mengetahui,
Ketua Departemen Agronomi dan Hortikultura Fakultas Pertanian IPB
Dr. Ir. Agus Purwito, MSc. Agr.
NIP: 19611101 198703 1 003
RIWAYAT HIDUP
Penulis dilahirkan di Jakarta pada tanggal 4 Januari 1988. Penulis merupakan anak ketiga dari lima bersaudara dari Bapak Tjhen Mie Hian dan Ibu Tjong Njuk Khiun.
Penulis menyelesaikan pendidikan Sekolah Dasar pada tahun 2000 di SD Budi. Tahun 2003 penulis menyelesaikan pendidikan SLTP di SLTP Methodist Jakarta kemudian menyelesaikan pendidikan Sekolah Lanjutan Tingkat Atas di SMAK Methodist Jakarta pada tahun 2006. Pada tahun yang sama, penulis diterima di Institut Pertanian Bogor dan pada tahun 2007 penulis diterima di Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor.
Selama menjalankan pendidikan di Institut Pertanian Bogor, penulis bergabung dalam Unit Kegiatan Mahasiswa ‘Persekutuan Mahasiswa Kristen’ (PMK) Institut Pertanian Bogor. Penulis aktif dalam kegiatan-kegiatan PMK seperti menjadi sekretaris dalam Retreat Komisi Kesenian PMK IPB pada tahun 2008, sekretaris Festival Seni PMK IPB pada tahun 2008, dan sekretaris Komisi Kesenian PMK IPB 2008/2009.
KATA PENGANTAR
Puji syukur penulis panjatkan ke hadirat Tuhan Yang Maha Esa atas berkat, anugerah serta pimpinanNya sehingga penelitian ini dapat diselesaikan dengan baik. Penelitian mengenai biologi pembungaan dan keberhasilan reproduksi Zingiber spectabile dan Tapeinochilos ananassae pada penyerbukan alami dan buatan didasarkan pada kondisi pasar tanaman hias Indonesia saat ini yang semakin didominasi oleh tanaman hias daerah subtropis. Kekayaan tanaman hias tropis Indonesia kurang dikenal dan diminati konsumen karena terbatasnya informasi dan masih sedikitnya teknologi yang dikembangkan untuk perbaikan tanaman, khususnya pemuliaan tanaman. Penelitian yang dilaksanakan di Balai Penelitian Tanaman Hias (Balithi) Segunung ini diharapkan menghasilkan informasi yang berguna bagi para pemulia untuk mengembangkan tanaman tropis Indonesia, terutama famili Zingiberaceae.
Penulis menyampaikan terima kasih kepada:
1. Seluruh keluarga, mama, papa, Viana, Veronika Dewi, Rini Puspita, Risa yang selalu mendukung dan mendoakan penulis dalam menyelesaikan penelitian 2. Dr. Ir. Endah Retno Palupi, MSc. selaku pembimbing I dan Ir. Debora Herlina
Adriyani, MS. sebagai pembimbing II yang telah memberikan bimbingan dan pengarahan kepada penulis sehingga penelitian ini dapat diselesaikan.
3. Dr. Desta Wirnas, SP. MSi selaku dosen penguji atas kritik dan saran yang telah diberikan.
4. Seluruh Dosen Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor atas pengajaran dan bimbingannya.
5. Bapak Dr. Budi Winarto atas bimbingannya selama penelitian di Balai Penelitian Tanaman Hias Segunung
6. Bapak Suyono Sri Suwarno selaku pembimbing lapang yang memberikan pengarahan selama penelitian di lapangan
8. Teman-teman seperjuangan di BALITHI Segunung; Ester Yentina, Paramyta Nila Permanasari, dan Hilaria Primapuspita atas bantuan, dukungan, dan doa selama penelitian
9. Teman-teman AGH 43; Sadewi Maharani, teman satu bimbingan (Andini Safitri dan Satrio Tunggul Pratomo), dan Nehemia J. A. Sinaga atas dukungan dan doa yang telah diberikan
10.Teman-teman Wisma Ananda 1: Vera, Novi, Yoan, Yessica, Diamond; Ian atas support yang diberikan
11.Untuk semua pihak yang tidak dapat disebutkan satu per satu.
Penulis mengharapkan penelitian ini dapat bermanfaat bagi ilmu pengetahuan dan semua pihak yang memerlukan.
Bogor, Januari 2011
DAFTAR ISI
DAFTAR TABEL ... ix
DAFTAR GAMBAR ... x
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan ... 3
TINJAUAN PUSTAKA ... 4
Tanaman Zingiberaceae ... 4
Perbanyakan Tanaman ... 6
Deskripsi Tanaman ... 7
Struktur Bunga ... 9
Fenologi Pembungaan ... 10
Keberhasilan Reproduksi dan Penyerbukan ... 11
Perkembangan Buah dan Biji ... 12
BAHAN DAN METODE ... 14
Tempat dan Waktu ... 14
Bahan dan Alat ... 14
Metode Penelitian ... 15
Pengamatan ... 19
HASIL DAN PEMBAHASAN ... 20
Kondisi Umum ... 20
Zingiber spectabile ... 22
Struktur Braktea dan Bunga ... 22
Viabilitas Polen ... 26
Masa Reseptif Stigma ... 27
Keberhasilan Reproduksi ... 30
Tapeinochilos ananassae ... 33
Struktur Braktea dan Bunga ... 33
Viabilitas Polen ... 36
Masa Reseptif stigma ... 37
Keberhasilan Reproduksi ... 39
KESIMPULAN DAN SARAN ... 42
DAFTAR TABEL
No Halaman
1 Karakteristik braktea Z. spectabile pada lima fase
perkembangan……….. 23
2 Karakteristik bungaZ. spectabile……… 25 3 Perubahan pada stigma dan antera Z. spectabile selama bunga
mekar………... 28
4 Jumlah biji Z. spectabile yang dihasilkan pada masing-masing
perlakuan waktu penyerbukan silang……… 29 5 Jumlah buah dan biji/buah pada masing-masing tipe
penyerbukan pada Z. spectabile……….. 31 6 Karakteristik braktea T. ananassae pada lima fase
perkembangan………. 33
7 Karakteristik bunga T. ananassae………... 35 8 Persentase polen T. ananassae yang berkecambah pada tiap
waktu pengambilan sampel………. 37 9 Perubahan stigma dan antera T. ananassae selama bunga
mekar………. 38
10 Jumlah biji yang dihasilkan pada masing-masing perlakuan
waktu penyerbukan pada T. ananassae……….. 38 11 Jumlah buah dan biji yang terbentuk dari tiap tipe pnyerbukan
DAFTAR GAMBAR
No Halaman
1 Bentuk daun: (a) lancealate (b) ovate……….. 5 2 (A) T. ananassae (B) Z. spectabile (C) Larutan stok
Brewbaker and Kwack (D) Mikroskop binokuler
(E) Mikroskop Stereo (F) Haemacytometer……… 15 3 Pengamatan diameter braktea: (A) Z. spectabile
(B) T. ananassae………. 16
4 Prosedur penghitungan jumlah polen dengan menggunakan
haemacytometer: (A) Struktur bunga (B) Pengamatan panjang pistil dan antera pada T. ananassae (C)
Haemacytometer yang ditetesi larutan polen (D) Bagian-bagian haemacytometer (E) Kotak-kotak untuk menghitung jumlah sel yang tampak di bawah mikroskop (F) Polen dalam
haemacytometer dilihat dari bawah mikroskop……… 17 5 Z. spectabile: (A) Lokasi penelitian (B) Hama ulat yang
menyerang (C) Bunga yang terserang (D) Braktea berwarna
kehitam-hitaman akibat kotoran cacing………... 21 6 T. ananassae: (A) Lokasi penelitian (B) Braktea yang
kehitam-hitaman akibat semut dan kutu daun (C) Kutu daun
famili Aphidae………. 21
7 Perkembangan braktea Z. spectabile: (A) Fase pertama (B)
Fase dua (C) Fase tiga (D) Fase empat (E) Fase lima……….. 23 8 Bunga Z. spectabile: (A) Kuncup (B) Mulai mekar
(C) Mekar………. 24
9 Bunga Z. spectabile: (A) Bagian-bagian bunga (B) Antera
(C) Stigma……… 25
10 Polen Z. spectabile yang dikecambahkan dalam media BK… 26 11 Periode viabilitas polen Z. spectabile berdasarkan
perkecambahan………. 27
12 Perkembangan diameter ovarium Z. spectabile setelah
penyerbukan pada tiga tipe penyerbukan………. 30 13 Perkembangan ovarium Z. spectabile setelah penyerbukan
ketiga lokus……….. 32 14 Perkembangan braktea T. ananassae: (A) Fase pertama (B)
Fase dua (C) Fase tiga (D) Fase empat (E) Fase lima
(F) Braktea abnormal……… 34 15 Bunga T. ananassae: (A) Kuncup (B) Mulai mekar
(C) Mekar penuh……….. 35
16 Bunga T. ananassae: (A) Struktur bunga (B) Struktur alat
reproduksi……… 36
17 Polen T. ananassae dalam media BK……….. 36 18 Perkembangan diameter ovarium T. ananassae setelah
penyerbukan………. 39
19 (A) Ovarium yang mulai pecah (B) Biji sudah masak
PENDAHULUAN
Latar Belakang
Hutan tropis Indonesia menyimpan keanekaragaman hayati, termasuk di dalamnya flora dan fauna. Keanekaragaman tersebut merupakan sumber daya bagi perekonomian, pariwisata, kesehatan, dan budaya bangsa. Secara alami, penyebaran keanekaragaman hayati itu tidak merata di setiap wilayah karena sangat bergantung pada ekosistem wilayahnya.
Indonesia merupakan salah satu daerah tropis yang memiliki ekosistem berupa flora dan fauna yang tergolong cukup beragam dibandingkan negara-negara lain di dunia. Famili Zingiberaceae, Heliconiaceae, Musaceae, dan Costaceae adalah sebagian kecil dari beragamnya jenis tanaman tropis yang ada di Indonesia. Pemanfaatan tanaman-tanaman tersebut sangat beragam mulai dari pangan, pengobatan, industri, sampai tanaman hias. Keseluruhan manfaat dari masing-masing tanaman sangat mendukung perekonomian Indonesia dan yang mulai ditingkatkan saat ini adalah sebagai tanaman hias.
Tanaman hias merupakan komoditas hortikultura yang berpotensi untuk dikembangkan dalam dunia bisnis saat ini. Jenis tanaman yang banyak diminati masyarakat saat ini adalah tanaman yang berasal dari negara-negara subtropis seperti krisan, mawar, gerbera, anyelir, dan tulip. Rendahnya minat konsumen akan jenis tanaman hias tropis diakibatkan kurangnya pengenalan akan tanaman hias tersebut, khususnya di Indonesia. Tanaman hias tropis yang ada di Indonesia memiliki nilai estetika yang mampu menyaingi keindahan tanaman-tanaman subtropis. Salah satu yang memiliki potensi cukup tinggi untuk dikembangkan dan keindahan yang unik adalah dari famili Zingiberaceae.
Famili Zingiberaceae yang berbunga merupakan famili terbesar dalam ordo Zingiberales, terdiri atas sekitar 50 genus dan lebih dari 1300 spesies. Tanaman aromatik ini dapat tumbuh di daerah lembab baik wilayah tropis maupun subtropis dengan populasi utama dan penyebaran spesies terpusat di Asia Selatan dan Tenggara (Branney, 2005). Walaupun dikatakan famili terbesar, namun jenis-jenis yang dikenal hanya beberapa saja, diantaranya genus Zingiber
Pemanfaatan tanaman dari famili Zingiberaceae ini belum optimal, sebagian besar digunakan sebagai obat-obatan, kosmetik, dan bumbu masakan sedangkan bagian bunganya sangat unik dan menarik. Salah satu kendala dalam budidaya bunga tanaman ini adalah kurangnya lahan untuk pertanaman dan dibutuhkan waktu yang cukup lama serta kurangnya informasi mengenai karakteristik dan teknik budidaya tanaman tersebut. Dilihat dari keunikan bunganya, famili Zingiberaceae memiliki prospek yang cerah dalam bisnis tanaman hias. Oleh karena itu perlu diupayakan pengembangan jenis/variasi baru melalui pemuliaan tanaman baik konvensional maupun mutasi.
Upaya yang dapat dilakukan untuk mengembangkan varian baru dan perbaikan varietas adalah dengan pemuliaan tanaman, baik induksi mutasi ataupun persilangan konvensional (hibridisasi). Namun teknik mutasi kurang efektif karena khimera (hasil mutasi) tidak dapat diwariskan, sehingga teknik yang dilakukan adalah persilangan konvensional. Menurut Suharsono (1996), persilangan antara dua tanaman yang berbeda dapat menghasilkan suatu tanaman yang mempunyai sifat-sifat baru yang merupakan kombinasi dari kedua tetuanya. Persilangan ini sangat penting dalam perakitan tanaman hias baik dalam spesies yang sama maupun antar spesies. Namun kegiatan ini memerlukan waktu yang relatif lama.
Tujuan
Penelitian ini bertujuan untuk:
1. Mempelajari biologi dan fenologi pembungaan Zingiber spectabile dan
Tapeinochilos ananassae.
2. Mempelajari keberhasilan reproduksi dari penyerbukan alami (terbuka) dan penyerbukan buatan, yaitu penyerbukan sendiri dan penyerbukan silang dari
TINJAUAN PUSTAKA
Tanaman Zingiberaceae
Famili Zingiberaceae terdiri atas 1300 spesies yang terbagi ke dalam 50 genus. Tanaman ini ditemukan di sepanjang wilayah tropis dan subtropis, dengan populasi terbesar dan keragaman spesies terpusat di Asia Tenggara dan Selatan (Branney, 2005). Beberapa tanaman Zingiberaceae seperti buah pelaga (Elettaria cardamomum), kunyit (Curcuma longa atau C. domestica), jahe (Zingiber officinale), dan lengkuas (Alpinia galanga) digunakan sebagai bumbu dan tanaman obat-obatan.
Menurut Simpson (2006), Zingiberaceae merupakan famili besar yang biasanya diklasifikasikan ke dalam empat suku, yaitu Hedychieae (daun sejajar dengan rimpang, mahkota bunga menyamping, tidak menyatu dengan labellum), Zingibereae (stilus mencuat melewati antera dan terbungkus oleh pembungkus antera), Alpinieae (daun tegak lurus dengan rhizoma, mahkota bunga kecil dan menyatu dengan labellum), dan Globbeae (filamen memanjang dan melengkung, ovariumsatu lokus).
Semua jahe-jahean tumbuh dari rhizoma tebal yang merupakan batang bagian bawah yang termodifikasi sebagai cadangan makanan. Bentuk dan ukuran rhizoma bervariasi antara genus yang satu dan yang lain. Rhizoma memiliki sejumlah mata tunas yang dorman selama di atas tanah. Akar jahe-jahean muncul dari rhizoma dan pada beberapa anggota famili Zingiberaceae dihasilkan umbi di ujung akar tersebut. Umbi ini terletak di bagian tanah yang sangat dalam, terutama pada genus Siphonochilus dan berperan sebagai sistem penyimpanan makanan tambahan yang hilang akibat cekaman iklim (Branney, 2005).
Batang yang terlihat pada sebagian besar tanaman Zingiberaceae bukan batang secara keseluruhan. Sebaliknya, batang tersebut adalah perpanjangan pelepah dari daun-daun yang tampak. Menurut Branney (2005), bentuk pelepah yang menghubungkan struktur ini dinamakan pseudostem (batang semu). Batang yang sebenarnya umumnya sangat pendek.
seperti pada Kaempferia. Sebagian besar daun Zingiberaceae memiliki tulang daun yang sangat mencolok dan terkadang berbeda warna dengan helaian daun. Pada spesies Curcuma, semua urat daun timbul dan memberikan efek menarik di sepanjang permukaan daun. Daun variegata relatif jarang pada famili ini, meskipun beberapa kultivar memiliki garis daun berwarna putih atau krem. Sebagian besar tanaman berwarna merah, perunggu atau keperakan, baik pada batang maupun pada susunan tulang dan urat daun.
(a) (b)
Gambar 1. Bentuk daun: (a) lanceolate (b) ovate
Sumber: http://www.smccd.net/accounts/leddy/leafshapes.htm
Belum lama ini, Costaceae diklasifikasikan sebagai Costoideae, subfamili dari Zingiberaceae. Di Asia, Costaceae hanya Costus dan Tapeinochilos (Larsen
et al., 1999). Secara botani, Costaceae dipisahkan dari Zingiberaceae karena tiga alasan. Pertama, Costaceae memiliki daun yang tersusun spiral di atas batang, sedangkan pada Zingiberaceae daun tersusun pada batang secara vertical dalam dua sisi yang berhadapan. Kedua, pada Costaceae semua stamen menyatu dengan
labellum, sedangkan pada Zingiberaceae hanya stamen bagian dalam yang menyatu dengan labellum. Terakhir, Costaceae tidak memiliki aroma seperti pada semua tanaman Zingiberaceae (Branney, 2005).
Syarat Tumbuh
Air sangat berperan dalam perkembangan rimpang, apabila kekurangan air, perkembangan rimpang akan sangat terhambat. Menurut Januwati et al. (1992) tanaman jahe tumbuh baik pada curah hujan antara 2500-4000 mm/tahun dan pada curah hujan rata-rata 3000 mm/tahun tanaman jahe akan tumbuh subur. Tanaman jahe dapat tumbuh pada ketinggian 0-1500 m dpl, namun ketinggian yang optimum bagi pertumbuhannya adalah 300-900 m dpl.
Cahaya mempengaruhi pertumbuhan dan produksi melalui proses fotosintesis dan reaksi fotoperiodisitas. Namun pengaruh intensitas cahaya terhadap petumbuhan tanaman lebih besar dibanding pengaruh dari perubahan dalam mutu penyinaran (Januwati et al., 1992)
Kisaran suhu yang memungkinkan pertumbuhan jahe optimum adalah 25-30 ºC. Suhu di atas 35 ºC akan menghanguskan daun, kemudian daun mongering, sedangkan makin rendah suhu maka umur tanaman akan semakin panjang. Struktur tanah yang cocok bagi tanaman jahe adalah tanah lempung berpasir, liat berpasir dan tanah laterit (Suprapti, 2003). Tanaman jahe dapat tumbuh pada keasaman tanah (pH) 4.3-7.4, tetapi pH optimum untuk jahe adalah 6.8-7.0 (Januwati et al., 1992)
Perbanyakan Tanaman
Famili Zingiberaceae dapat diperbanyak secara generatif maupun vegetatif. Perbanyakan secara generatif yaitu melalui biji jarang dilakukan karena relatif sulit. Menurut Branney (2005) hanya Roscoea, Cautleya dan spesies
Hedychium pada topografi yang lebih tinggi yang secara teratur memproduksi biji ketika ditanam pada kondisi outdoor di Inggris. Biji pada kebanyakan spesies Zingiberaceae memiliki viabilitas rendah dan keberhasilan perkecambahan yang menurun seiring dengan lamanya penyimpanan.
banyak titik tumbuh yang berkembang menjadi dua atau lebih tanaman baru di akhir musim pertumbuhan. Tanaman seperti ini dapat dipisahkan kapan saja selama masa dormansi, selama tidak ada kerusakan pada jaringan dan tidak ada pemotongan pada permukaan rhizoma.
Perbanyakan melalui stek juga mungkin dilakukan pada famili Zingiberaceae. Beberapa genus yang pernah dilaporkan dapat diperbanyak dengan stek adalah pada genus Globba, Hedychium, dan genus Zingiber yang paling banyak disebut. Bagian yang digunakan adalah batang semu sekitar 15-45 cm atau keseluruhan panjangnya, tergantung varietas dan yang memiliki satu atau dua daun (Branney, 2005). Batang semu yang sudah menghasilkan bunga tidak sesuai untuk digunakan sebagai bahan perbanyakan melalui stek, tetapi apabila batang semu telah mengeras, ada kemungkinan dapat digunakan.
Beberapa anggota Zingiberaceae secara alami menghasilkan keturunan, dengan anakan kecil yang terbentuk dari perbungaan (spika), terutama genus
Globba (Branney, 2005). Sebagian besar spesies Globba menghasilkan umbi dalam jumlah besar pada bagian dasar perbungaan. Alpinia purpurata tropis dan
Hedychium greenii juga memproduksi anakan yang berkembang dari perbungaan yang memudar (fading inflorescence).
Kultur jaringan adalah suatu teknik untuk menumbuhkan sel dan jaringan dalam medium buatan dalam lingkungan aseptik untuk tumbuh dan berkembang menjadi tanaman sempurna atau anakan (Armini, 2008). Beberapa tanaman tidak siap untuk diperbanyak dengan metode ini, tetapi jahe-jahean dapat diperbanyak melalui metode kultur jaringan. Peningkatan jumlah jahe-jahean merupakan hasil dari perbanyakan melalui kultur jaringan di beberapa Negara di Eropa, Asia dan Amerika. Spesies Curcuma, Globba, Hedychium, dan Kaempferia telah diperbanyak dengan metode kultur jaringan dalam jumlah yang sangat besar (Branney, 2005).
Deskripsi Tanaman
dari 150 spesies, yang berasal dari Asia Selatan dan Tenggara, terutama Thailand dan Cina Selatan.
Zingiber merupakan tanaman jahe-jahean berukuran sedang, yaitu dengan tinggi tanaman mulai dari 30–180 cm. Tanaman ini tumbuh tegak dan biasanya memiliki batang semu tebal yang menopang daun berbentuk lanceolate. Sebagian besar spesies memiliki periode pembungaan yang singkat dan muncul langsung dari rhizoma, tidak jauh dari batang semu. Bunganya berbentuk kerucut, terkadang sangat panjang, terkadang pendek dan tidak cerah. Sebagian besar spesies memiliki kumpulan braktea yang tersusun erat (kompak), braktea yang mekar penuh biasanya berwarna kuning, merah atau cokelat kastanye. Braktea ini umumnya terlihat sangat mencolok dan memiliki masa pajang yang sangat panjang. Tanaman ini merupakan salah satu spesies yang dibudidayakan terutama untuk perdagangan bunga potong (Branney, 2005).
Zingiber spectabile diberi nama oleh William Griffith sebagai Zingiber
yang mengesankan (spectabile), tanaman ini juga terkenal sebagai jahe sarang lebah untuk menggambarkan perbungaannya yang mengagumkan (Branney, 2005). Zingiber spectabile merupakan tanaman asli dari Thailand Selatan, semenanjung Malaysia, dan Sumatera, yang tumbuh di hutan tropis yang padat dan ternaungi.
Tanaman ini bisa mencapai tinggi hingga 3 m. Braktea Z. spectabile
berwarna kuning dan coklat keunguan dengan bintik-bintik kuning pada mahkota bunga sejatinya (www.plant-group.com). Z. spectabile dengan perubahan warna braktea selama pendewasaan dan masa pajang sekitar dua minggu menjadi alasan penggunaan bunga ini sebagai bunga potong (Larsen et al., 1999).
Zingiber spectabile tumbuh baik di dataran rendah sampai dataran tinggi. Selama pertumbuhannya, tanaman ini memerlukan naungan. Penggunaan naungan paranet yang meneruskan intensitas cahaya 45% sangat baik untuk pertumbuhannya.
tetap bersih sampai waktu panen. Untuk persiapan tanam, lahan perlu digemburkan menggunakan bahan organik, yaitu sekam, kompos, atau pupuk kandang. Lahan dibuat bedengan dengan lebar 2 m. Jarak antar tanaman sebaiknya minimal 1 m x 1 m (Adriyani, 2007).
Genus Tapeinochilos terdiri dari 8-10 spesies berasal dari New Guinea,
Australia, dan Indonesia. Beberapa ahli mengklasifikasikan genus ini ke dalam famili Costaceae. Spika bunga Tapeinochilos umumnya muncul langsung dari rhizoma. Spesies ornamental lain memiliki braktea yang bervariasi mulai dari warna merah dan kuning sampai hampir hitam. Jenis ini termasuk tanaman tropis karena itu relatif jarang dibudidayakan di wilayah Amerika (Llamas, 2003).
Tapeinochilos ananassae atau dikenal dengan nama bunga kasturi merupakan spesies yang berasal dari Sulawesi. Spesies tanaman dari famili Costaceae ini memiliki dua kultivar, yaitu kultivar yang mempunyai braktea berwarna merah darah (Sekar Sauli) dan kultivar yang mempunyai braktea berwarna merah jingga (Sekar Manise). Tanaman ini termasuk herba perennial yang mempunyai rimpang non aromatik. Daun tersusun spiral dengan lamina tunggal, berbentuk lonjong dengan permukaan licin. Daun terkonsentrasi pada batang bagian atas. Rangkaian bunga berbentuk seperti bunga pinus, terletak terminal pada tunas atau terpisah dari batang berasal dari rhizoma (Adriyani, 2007). Braktea muncul dari ujung batang dan tunas samping. Warna braktea merah dengan helaian yang kaku dan keras serta bagian ujung runcing. Bunga sejati berwarna kuning (Adriyani dan Suwarno, 2002).
Struktur Bunga
Perbungaan pada famili Zingiberaceae dapat dikelompokkan menjadi dua, yaitu secara terminal (muncul dari ujung batang) dan secara radikal (muncul dari rhizome). Secara terminal, terdapat tangkai yang panjang dengan braktea di bagian atas. Hal yang paling jarang adalah ketika perbungaan secara terminal dan radikal muncul pada tanaman yang sama. Fenomena ini dapat dilihat pada
Secara umum perbungaan Zingiberaceae dideskripsikan seperti tombak atau rangkaian. Ketika braktea tumbuh tumpang tindih (overlap), rangkaian bunga (spika) tampak seperti bunga pinus. Pembentukan braktea biasanya tersusun spiral ketika berjumlah banyak; kadang-kadang jumlahnya sedikit, individual, dan berbentuk seperti mangkok (seperti pada Camptandra). Pada beberapa genus (Curcuma, Etlingera, Zingiber) braktea utamanya besar dan memiliki warna pink, kuning, jingga, ungu atau putih yang indah. Keistimewaan ini sering dimanfaatkan untuk tujuan komersial.
Bunga sejati sangat khas dan biseksual. Mulai dari yang kecil sekali seperti pada genus Globba sampai tipe besar seperti Alpinia. Bunga jahe-jahean sangat rapuh dan hanya mampu bertahan sebentar saja. Bunga ini memiliki ovarium yang kurang baik, yang memungkinkan satu lokus dengan ovul pada tiga plasenta di sepanjang dinding, atau yang lebih sering, 3 lokus dengan plasenta aksilar.
Kelopak adalah pipa dengan tiga atau kadang dua helaian. Mahkota terdiri atas sebuah daun mahkota tipis berbentuk pipa dan tiga daun mahkota yang hampir serupa, di mana daun mahkota bagian atas biasanya sedikit berbeda bentuknya. Secara sederhana, bunga Zingiberaceae mirip bunga anggrek yang memiliki bibir atau “labellum”.
Dua benang sari samping akan berubah menjadi struktur seperti mahkota pada genus Globba dan Hedychium atau menjadi dua lidah kecil pada Alpinia dan
Zingiber. Benang sari fertil memiliki tangkai sari yang bervariasi panjangnya dan berakhir pada kotak sari, yang pada kebanyakan spesies membuka dengan celah membujur (Larsen et al., 1999).
Fenologi Pembungaan
Polen yang berhasil berkecambah di atas stigma akan mulai memanjang masuk ke dalam saluran stilus menuju bakal buah (Darjanto dan Satifah, 1990).
Polen merupakan jaringan hidup yang memiliki umur terbatas kemudian mati. Polen yang baik adalah polen dari kuncup bunga yang telah dewasa yang hampir merekah karena pada saat itu ruang sari pada polen tersebut belum pecah dan terisi penuh dengan polen yang memiliki daya tumbuh yang tinggi. Suhu yang tinggi dan cekaman lain selama pendewasaan polen dan penyerbukan diduga memberi pengaruh buruk terhadap biji yang dihasilkan (Mascarenhas and Altschuler, 1982). Menurut Darjanto dan Satifah (1990), untuk perkecambahan serbuk sari (polen) umumnya diperlukan suhu berkisar antara 15 ºC sampai 35 ºC. Suhu yang lebih tinggi akan menyebabkan penguapan air sehingga banyak polen yang akan mengering.
Viabilitas polen yang digunakan akan mempengaruhi viabilitas benih yang dihasilkan. Polen dengan viabilitas tinggi akan lebih dahulu membuahi sel telur sehingga akan menghasilkan buah dengan mutu yang baik dan benih dengan viabilitas yang tinggi (Widiastuti dan Palupi, 2008)).
Keberhasilan Reproduksi dan Penyerbukan
Keberhasilan reproduksi pada tanaman diartikan sebagai jumlah ovul yang berkembang sempurna menjadi biji yang viabel dan mampu bertahan hidup dalam batas waktu tertentu. Menurut Wiens et al. (1987) faktor pembatas dari keberhasilan reproduksi antara lain faktor lingkungan (cuaca, unsur hara, serangga, serangan hama dan penyakit) dan faktor genetik (gen dan kromosom).
Penyerbukan adalah peristiwa menempelnya serbuk sari di kepala putik dengan perantara angin, air, serangga, atau hewan lain. Penyerbukan yang berhasil akan menyebabkan fertilisasi yang disertai pembentukan buah dan biji (Mangoendidjojo, 2003). Sistem penyerbukan pada Zingiberaceae belum banyak diteliti. Proses penyerbukan yang diamati pada beberapa spesies dibantu oleh kupu-kupu dan ngengat.
terbatasnya vektor penyerbukan. Menurut Lelivelt (1993) keberhasilan penyerbukan bergantung pada dua genotip pistil dan induk polen serta jarak genetik antara genotipenya. Faktor waktu penyerbukan dan cuaca juga mempengaruhi keberhasilan.
Waktu penyerbukan harus ditentukan agar tepat dan tidak terlambat. Baik putik maupun benang sari harus dalam keadaan segar, sehat dan telah masak. Pertumbuhan serbuk sari dipengaruhi oleh suhu udara. Menurut Darjanto dan satifah (1990), cuaca cerah dan udara yang agak lembab merupakan kondisi yang baik untuk penyerbukan dan pada iklim dingin bunganya tidak cepat layu sehingga dapat lebih lama diserbuki.
Perkembangan Buah dan Biji
Pembuahan terjadi apabila serbuk sari yang menempel di kepala putik berkecambah dan membentuk tabung polen. Tabung polen akan terus memanjang masuk ke dalam stilus menuju ke ovarium dan kantung embrio. Oleh karena itu tabung polen harus lebih panjang daripada tangkai putik. Dua inti sperma yang terdapat dalam tabung polen, satu akan melebur dengan inti sel telur membentuk zigot dan yang lain melebur dengan inti polar dalam kantung embrio membentuk endosperma. Peleburan dua inti sperma (generatif) dengan satu inti sel telur dan dua inti polar disebut pembuahan ganda. Zigot yang terbentuk akan berkembang menjadi embrio, sedangkan endosperma akan menjadi jaringan yang menyediakan zat makanan untuk pertumbuhan embrio. Sebelum tumbuh menjadi embrio, zigot akan mengalami fase istirahat selama beberapa waktu sehingga dalam 1-2 minggu pertama setelah penyerbukan belum dapat diketahui apakah penyerbukan tersebut gagal atau akan berlangsung dengan pembuahan (Darjanto dan Satifah, 1990).
Bentuk buah pada Zingiberaceae umumnya bulat dengan bakal buah tenggelam, beruang tiga dengan bakal biji dalam tiap ruangnya. Endosperma dikelilingi perisperm dan embrio. Biji banyak dan tidak mempunyai endosperma besar sehingga banyak biji yang tidak dapat berkecambah pada saat ditanam (Purseglove dan Tjitrosoepomo dalam Yunira, 2009).
(Zingiber spectabile) atau dari ungu kemerahan menjadi merah pada Zingiber ottensi dan ketika setiap braktea sudah membuka sehingga bijinya terlihat. Pemanenan Alpinia dilakukan secara bertahap setelah buah masak yang ditandai dengan perubahan warna. Pada Alpinia purpurata ‘Jungle King’ buah akan berubah warna menjadi merah bila sudah masak, sedangkan pada Alpinia purpurata ‘Jungle Queen’ menjadi merah muda dan kelopak bunga sudah mengering serta kulit buah sudah mulai keriput (Yunira, 2009). Masing-masing benih terbungkus oleh arilus berwarna putih. Pemasakan buah terjadi pada umur
≥ 3 bulan setelah penyerbukan. Genus Alpinia dapat berbunga sepanjang musim namun jarang menghasilkan buah, sedangkan genus Zingiber dapat menghasilkan buah/biji pada tiap malainya hanya saja spesies ini berbunga pada musim-musim tertentu, biasanya pada musim hujan, setelah selesai musim hujan tanaman ini jarang berbunga.
Biji pada anggota famili Zingiberaceae sebaiknya disemai segera sesudah dipanen. Biji dapat disebar di pot atau tray, tergantung jumlah yang tersedia. Sebagian besar biji Zingiberaceae memerlukan penyimpanan pada suhu antara 21–24 ºC pada daerah yang lebih dingin untuk keberhasilan perkecambahan (Branney, 2005).
BAHAN DAN METODE
Tempat dan Waktu
Penelitian ini dilaksanakan di Balai Penelitian Tanaman Hias (Balithi) Segunung dengan ketinggian 1 100 m dpl (di atas permukaan laut). Penelitian dilakukan pada Februari - Agustus 2010.
Bahan dan Alat
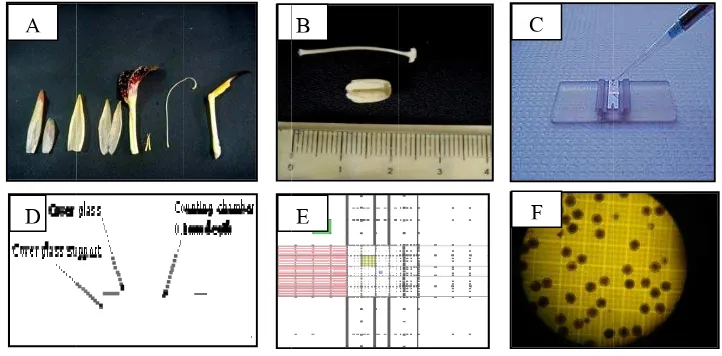
Bahan yang digunakan dalam penelitian ini adalah tanaman Zingiberaceae yang sudah berbunga. Tanaman ini merupakan koleksi Balithi yang ditanam pada tahun 2007. Spesies yang digunakan antara lain Tapeinochilos ananassae
(Gambar 2A) dan Zingiber spectabile (Gambar 2B). Semua tanaman Zingiberaceae yang digunakan tumbuh baik pada kondisi iklim di ketinggian 1 100 m dpl. Suhu rata-rata siang hari berkisar antara 24–26 ºC dan pada malam hari berkisar antara 18 – 20 ºC, dengan kelembaban rata-rata 70 - 90 %.
Alat-alat yang digunakan antara lain jangka sorong, penggaris, label, lup, mikroskop binokuler untuk mengamati jumlah polen, mikroskop stereo untuk mengamati struktur bunga, pinset, scalpel atau pisau, sungkup plastik untuk penyerbukan, gelas objek, dan cawan petri untuk pengamatan viabilitas polen.
Gambar 2. (A) T. ananassae (B) Z. spectabile (C) Larutan stok Brewbaker and
Kwack (D) Mikroskop binokuler (E) Mikroskop stereo (F) Haemacytometer
Metode Penelitian
1. Struktur braktea dan bunga
Struktur braktea dan bunga yang diamati adalah dari genus Tapeinochilos
dan Zingiber. Pengamatan struktur braktea dimulai pada saat braktea masih kuncup sampai mekar untuk mengetahui periode pembukaan braktea, dengan 10 sampel untuk masing-masing kultivar. Struktur bunga diamati pada saat bunga mekar.
Pengamatan struktur braktea meliputi panjang dan diameter braktea (Gambar 3A, B), panjang dan diameter tangkai braktea, warna braktea, dan persentase kemekaran braktea. Pengamatan pada braktea dilakukan setiap minggu sejak braktea kuncup (persentase membuka 10 %) sampai braktea mekar penuh (80 - 100 %) pada saat tidak ada lagi bunga biologi yang muncul. Setelah braktea mekar penuh, helaian braktea dihitung dengan cara dilepas satu per satu untuk memudahkan perhitungan dan menghindari kesalahan karena helaian braktea yang rapat satu sama lain.
A B C
Gambar 3. Pengamatan diameter braktea: (A) Z. spectabile (B) T. ananassae
Pengamatan struktur bunga meliputi panjang bunga, panjang dan lebar helaian mahkota bunga, panjang dan lebar labellum, warna bunga, panjang pistil dan kotak polen, jumlah ovul dan jumlah polen. Pengamatan ini bertujuan untuk mengetahui karakteristik tiap kultivar. Perhitungan jumlah ovul dilakukan di bawah mikroskop stereo karena ukuran ovul yang sangat kecil, sedangkan perhitungan jumlah polen dilakukan dengan menggunakan haemacytometer.
Cara penggunaan haemacytometer sebagai berikut: Sampel bunga diambil, kemudian pistil dan stamen dipisahkan (Gambar 4A). Polen dari kepala sari yang terdiri dari dua antera (Gambar 4B) dilarutkan dalam aquades. Larutan polen kemudian diteteskan ke atas coverglass (Gambar 4C) pada haemacytometer, kemudian ditutup dengan gelas objek (Gambar 4D) dan dihitung di bawah mikroskop. Jumlah cairan yang terdapat antara coverglass dan alat ini mempunyai volume tertentu sehingga satuan isi yang terdapat dalam satu bujur sangkar juga tertentu. Ruang hitung terdiri dari 9 kotak besar dengan luas 1 mm² (Gambar 4E). Pengamatan dilakukan sebanyak 10 ulangan, dimana setiap ulangan dipilih 5 kotak dari 9 kotak besar secara acak. Jumlah polen yang diperoleh dari kelima kotak tersebut dijumlahkan dan dikali 2000, kemudian dirata-rata dari 10 ulangan. Jumlah polen pada satu sampel bunga adalah hasil rata-rata dari 10 ulangan tersebut (Gambar 4F).
3. Penentuan masa reseptif
Masa reseptif stigma ditentukan berdasarkan pengamatan morfologi stigma (adanya sekresi), fleksibilitas stilus dan pengamatan antera serta percobaan penyerbukan pada waktu yang berbeda-beda sejak bunga mekar mulai pukul 08.00 setiap satu jam sampai pukul 14.00. Penyerbukan dilakukan pada waktu yang berbeda mulai dari bunga mekar sampai layu agar diperoleh waktu yang paling tepat. Waktu penyerbukan yang menghasilkan persentase pembentukan buah dan biji yang paling tinggi menunjukkan masa reseptif stigma. Masing-masing waktu diserbuki lima sampel bunga dengan tiga ulangan sehingga totalnya adalah 15 bunga tiap waktu penyerbukan.
4. Penyerbukan
Penyerbukan yang dilakukan adalah penyerbukan buatan, yaitu menyerbuki kepala putik dengan polen dari satu antera dengan menggunakan pinset. Setelah dilakukan penyerbukan, kepala putik ditutup dengan sungkup agar terlindung dari kontaminasi oleh serbuk sari yang lain. Percobaan penyerbukan dilakukan dengan perincian sbb.:
1. Penyerbukan alami (terbuka): bunga dibiarkan terbuka masing-masing pada lima sampel dengan tiga ulangan.
2. Penyerbukan buatan: penyerbukan yang dilakukan dengan bantuan manusia, terdiri dari:
a. Penyerbukan sendiri: bunga diemaskulasi kemudian diserbuk dengan polen dari bunga yang sama,dilakukan masing-masing pada lima sampel dengan tiga ulangan.
b. Penyerbukan silang: bunga diemaskulasi kemudian diserbuk dengan polen dari bunga yang berbeda masing-masing pada lima sampel dengan tiga ulangan.
Zapata dan Arroyo dalam Tangmitcharoen dan Owens (1997) mengungkapkan bahwa intensitas self incompatibility (Index of self incompatibility) dapat ditentukan berdasarkan rumus sbb.:
Index of self incompatibility dapat dikelompokkan menjadi beberapa kategori yaitu:
1. Completely self incompatible (ISI=0) 2. Mostly self incompatible (0<ISI<0.2) 3. Partialy self incompatible (0.2<ISI<1) 4. Completely self compatible (ISI≥1)
Pengamatan
1. Pengamatan struktur braktea dan bunga meliputi:
- Struktur braktea: panjang dan diameter braktea, jumlah helaian braktea, dan lama perkembangan braktea.
- Individu bunga: panjang bunga, panjang dan lebar labellum, panjang dan lebar mahkota bunga, warna mahkota (ditentukan dengan menggunakan
colour chart), panjang pistil dan panjang antera, jumlah ovul (dihitung di bawah mikroskop) serta jumlah polen.
2. Penentuan viabilitas polen melalui pengamatan persentase polen yang berkecambah.
3. Penentuan masa reseptif stigma melalui pengamatan sekresi dan perubahan di permukaan stigma dilakukan dengan mengamati:
- kemekaran - sekresi - posisi stylus
- antera, dan
- persentase biji/buah yang terbentuk
4. Keberhasilan penyerbukan melalui pengamatan persentase buah dan biji/buah yang terbentuk.
Analisis Data
HASIL DAN PEMBAHASAN
Kondisi Umum
Zingiber spectabile ditanam dengan naungan paranet 45 % pada lahan seluas 20 m x 20 m dan terdapat 18 bedengan di dalamnya. Masing-masing bedengan memiliki ukuran 8 m x 1 m (Gambar 5A). Tanaman Z. spectabile
varietas Silvana yang ditanam merupakan hasil perbanyakan dengan metode stek. Periode pembungaan Z. spectabile adalah Februari-Juli.
Kendala yang dihadapi selama pelaksanaan penelitian adalah curah hujan yang tinggi dan jumlah bunga yang tidak mencukupi sehingga menghambat pengamatan. Hama yang menyerang Z. spectabile cukup banyak. Ulat merupakan hama yang paling banyak (Gambar 5B) dan mengganggu karena menyerang braktea dan bunga yang digunakan untuk penyerbukan (Gambar 5C). Penanggulangan yang dilakukan adalah dengan menyemprotkan insektisida dengan bahan aktif Tiodikarb 75 % setiap dua minggu dengan konsentrasi 1 gr/liter. Hama lain yang mengganggu adalah cacing yang tinggal dalam braktea
Z. spectabile dan meninggalkan kotoran sehingga braktea menjadi kehitam-hitaman (Gambar 5D).
Selama masa pembungaan (generatif) tanaman tidak dipupuk. Penyiraman juga tidak dilakukan karena curah hujan yang cukup tinggi. Pemeliharaan yang dilakukan adalah membersihkan gulma di sekitar bedengan dan pemangkasan batang dan daun yang sudah tua setiap dua bulan agar pertumbuhan tanaman tidak terhambat dan untuk mengurangi kelembaban akibat tanaman yang terlalu rapat.
Tapeinochilos ananassae yang digunakan dalam penelitian ini adalah kultivar ‘Sekar Manise’ yang memiliki warna merah jingga. Tanaman ditanam pada bedengan yang bersebelahan dengan Z. spectabile. Bunga T. ananassae
Gambar 5. Z. spectabile: (A) Lokasi penelitian (B) Hama ulat yang menyerang (C) Bunga yang terserang ulat (D) Braktea yang kotor dan kehitam-hitaman akibat kotoran cacing
Hama yang mengganggu pertumbuhan T. ananassae adalah ulat. Selain itu semut yang membuat sarang di celah braktea, membuat helaian braktea menjadi kehitam-hitaman (Gambar 6B). Selain ulat dan semut, T. ananassae banyak diserang kutu daun (Aphideae) yang juga mengakibatkan helaian braktea berwarna kehitaman (Gambar 6C). Pemeliharaan T. ananassae sama dengan Z. spectabile, yaitu penyiangan gulma di sekitar bedengan dan pemangkasan.
Gambar 6. T. ananassae: (A) Lokasi penelitian (B) Braktea yang kehitaman akibat semut dan kutu daun (C) Kutu daun famili Aphideae
A
B
C
D
Zingiber spectabile
Struktur Braktea dan Bunga
Setiap spesies yang diamati memiliki karakteristik yang bervariasi, baik struktur braktea yang meliputi panjang, diameter, dan warna braktea, maupun struktur bunga. Perkembangan braktea Z. spectabile dapat dikelompokkan ke dalam lima tahap (fase). Fase pertama adalah saat braktea mulai membuka (10 %) dan berwarna kuning pucat yang mulai muncul akhir Januari sampai Februari. Panjang braktea rata-rata berkisar antara 3.97±2.07 cm dengan diameter antara 3.66±1.85 cm.
Fase dua ditandai dengan braktea yang mulai menguning dan berjumlah antara 20–30 helaian braktea. Kuncup bunga mulai tampak di dalam helaian braktea pada fase ini. Panjang dan diameter braktea bertambah dari fase pertama ke fase kedua masing-masing menjadi berkisar 7.07±2.47 cm dan 6.196±0.98 cm dalam waktu sekitar tiga minggu.
Fase tiga adalah fase individu bunga muncul dan mekar secara bergantian yang dimulai dari helaian braktea bagian bawah. Satu spika mengandung 30–40 helaian braktea. Penyerbukan sudah dapat dilakukan pada fase ini karena bunga yang diperlukan sudah tersedia. Waktu yang diperlukan dari fase kedua sampai fase ini adalah sekitar empat minggu, yaitu pada akhir bulan Maret.
Pada fase empat braktea mulai mengalami perubahan warna dari kuning ke merah. Tampak semburat-semburat merah yang muncul dari braktea bagian bawah dan semakin lama semakin ke atas. Bunga masih tetap muncul pada fase ini, biasanya berada pada helaian braktea bagian tengah. Panjang braktea mencapai 13.47±2.06 cm dengan diameter 7.49±0.41 cm. Fase ini terjadi 4–5 minggu setelah fase ketiga.
Tabel 1. Karakteristik braktea Z. spectabile pada lima fase perkembangan Fase Perkembangan Waktu Ukuran braktea(cm) Keterangan
Panjang Diameter 1 Braktea mulai
membuka
Januari -
Februari 3.97±2.07 3.66±1.85
Braktea berwarna
minggu 7.07±2.47 6.20±0.98
Jumlah helaian braktea
minggu 10.19±2.12 7.09±0.57
Dapat dilakukan
minggu 13.47±2.06 7.49±0.41 Bunga sudah mencapai braktea tengah
minggu 13.47±2.06 7.52±0.39 Bunga sudah jarang atau tidak muncul lagi
Gambar 7. Perkembangan braktea Z. spectabile: (A) Fase pertama (B) Fase dua (C) Fase tiga (D) Fase empat (E) Fase lima
A B C
Lama perkembangan braktea mulai dari fase pertama sampai fase terakhir memerlukan waktu sekitar 13–17 minggu. Pemanfaatan braktea Z. spectabile
sebagai tanaman hias, terutama bunga potong, dapat dilakukan pada semua fase perkembangannya, kecuali fase pertama karena brakteanya masih kuncup (Gambar 7). Fase tiga, empat dan lima baik digunakan sebagai bunga potong karena ukuran dan warna yang memiliki nilai komersil. Penentuan ukuran yang tepat tergantung tujuan penggunaan dalam dekorasi. Sebagai bunga potong, Z. spectabile memiliki masa pajang sekitar 14 hari.
Pada setiap helaian braktea terdapat satu bunga Z. spectabile. Bunga yang akan mekar ditandai dengan kuncup yang sudah berkembang hingga keluar braktea (Gambar 8A), yang dapat diidentifikasi sekitar pukul 09.00. Kelopak bunga mulai membuka (Gambar 8B) sekitar pukul 11.00 yang diikuti dengan munculnya mahkota bunga. Bunga yang mekar pada hari yang sama terjadi pada waktu yang hampir bersamaan, yaitu sekitar pukul 12.00 (Gambar 8C). Bunga mekar secara berurutan mulai dari braktea bagian bawah. Sejak kuncup bunga muncul dalam braktea sampai bunga mekar diperlukan waktu antara 7-9 minggu (fase dua sampai fase empat).
Bunga Z. spectabile termasuk bunga lengkap dan bunga sempurna, terdiri dari kelopak, mahkota sebanyak tiga helai, satu helai labellum yang berwarna ungu dengan bintik kuning, pistil, dan stamen (Gambar 9). Warna mahkota bunga
Z. spectabile bervariasi dengan rentang dari yellow group 10C sampai yellow orange group 14D (Tabel 2). Pistil Z. spectabile lebih panjang daripada stamen, sehingga posisi kepala putik (stigma) lebih tinggi daripada antera. Menurut
Gambar 8. Bunga Z. spectabile: (A) Kuncup (B) Mulai mekar (C) Mekar
Darjanto dan Satifah (1990), tipe bunga dengan stilus yang panjang dan benang sari (stamen) yang pendek atau sebaliknya disebut heterostyly. Tipe bunga seperti ini termasuk ciri tanaman menyerbuk silang. Antera hanya dua buah yang menempel pada anther crest (pembungkus antera yang berbentuk seperti tanduk dan menopang stilus) serta filamen yang menyatu dengan bunga.
Gambar 9. Bunga Z. spectabile: (A) Bagian-bagian bunga (B) Antera (C) Stigma Jumlah ovul dalam setiap bunga berkisar 43.8±10.69 yang terletak dalam tiga lokus. Tingginya variasi jumlah ovul ini mempengaruhi jumlah biji yang
dapat terbentuk karena ovul merupakan bakal biji. Hasil perhitungan jumlah polen mencapai 71 680 per antera (Tabel 2). Jika dalam satu braktea Z. spectabile yang mekar penuh terdapat sekitar 85 bunga, maka potensi benih yang dapat terbentuk sekitar 3 723 benih/braktea dan jumlah polen dalam satu braktea mencapai 12 185 600 polen/braktea.
Tabel 2. Karakteristik bungaZ. spectabile
Variabel Ukuran Panjang bunga (cm) 6.8±0.35
Panjang labellum (cm) 3.17±0.24
Lebar labellum (cm) 1.36±0.10 Panjang mahkota (cm) 3.23±0.15 Lebar mahkota (cm) 0.72±0.05 Warna mahkota yellow group 10C – 14D Panjang pistil (cm) 5.84±0.25 Panjang antera (cm) 1.16±0.23 Jumlah ovul/ovarium 43.8±10.69 Jumlah polen/antera
Jumlah helaian braktea
71 680 97.4±17.4 Stigma
Anther crest
Antera
Labellum
diameter polen tersebut (Gambar 10). Polen yang tidak viabel tidak berkecambah (tetap bulat) atau pecah yang umumnya karena plasmolisis sehingga sitoplasma dalam polen keluar.
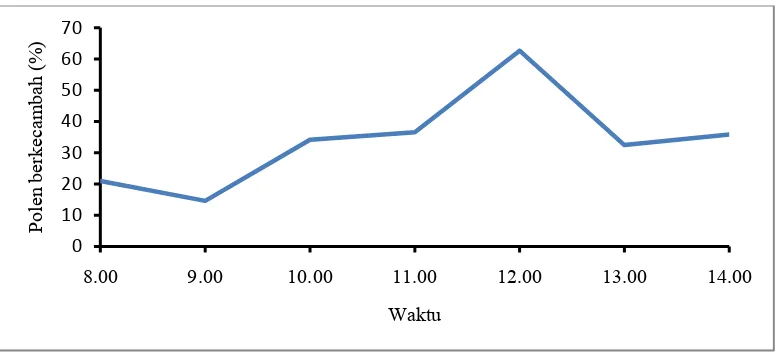
Gambar 11. Periode viabilitas polen Z. spectabile berdasarkan perkecambahan
Viabilitas polen antara pukul 08.00-09.00 masih rendah, berkisar antara 14.6-21 % dan terus meningkat hingga pukul 12.00 mencapai 62.7 % tetapi mulai menurun pada pukul 13.00 sekitar 32.5 % (Gambar 11). Data ini sejalan dengan hasil penelitian yang telah dilakukan sebelumnya pada Alpinia purpurata dan
Etlingera elatior, yang menunjukkan bahwa viabilitas polen meningkat pada saat bunga mekar penuh sekitar pukul 09.00-11.00 (Oktaviani, 2009). Data ini menunjukkan periode viabilitas polen yang dapat digunakan untuk penyerbukan terjadi antara pukul 10.00-14.00.
Masa Reseptif Stigma
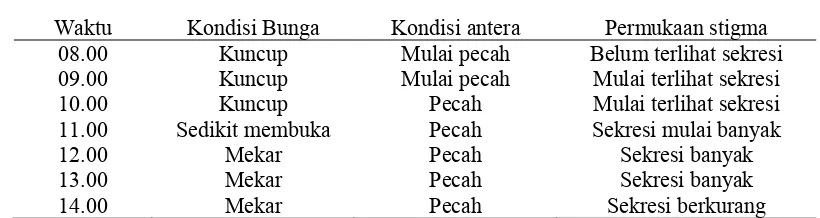
Pengamatan terhadap stigma dan antera menunjukkan bahwa antera sudah mulai pecah dan sekresi pada permukaan stigma sudah terlihat pada saat bunga masih kuncup pada pukul 09.00. Pada saat bunga mulai mekar pada pukul 11.00, antera telah pecah dan sekresi mulai bertambah (Tabel 3). Pada pukul 12.00, bunga sudah mekar penuh, antera sudah pecah dan sekresi pada permukaan stigma bertambah banyak. Banyaknya sekresi pada stigma yang diproduksi menunjukkan
0 10 20 30 40 50 60 70
8.00 9.00 10.00 11.00 12.00 13.00 14.00
Polen berkecam
bah
(%)
bahwa antara pukul 11.00-13.00 merupakan waktu yang baik untuk penyerbukan, karena pada saat itu stigma reseptif. Kondisi stilus melengkung sejak bunga kuncup hingga mekar. Hal ini berbeda dengan A. blepharocalyx yang mempunyai stilus fleksibel (flexistyly). Stigma pada pagi hari mengarah ke labellum
sedangkan pada sore hari di arah yang berlawanan. Pergerakan stilus ini berkaitan dengan masa reseptif stigma, yaitu pada saat stigma mengarah ke labellum (Sun et al., 2007).
Menurut Oktaviani (2009) A. purpurata dan E. elatior mulai mekar antara pukul 09.00-10.00 dan sekresi pada permukaan stigma sudah terlihat sesaat sebelum bunga mekar dan bertambah banyak beberapa saat setelah bunga mekar. Sekresi mulai berhenti antara pukul 12.00-15.00 yang mengindikasikan mulai
menurunnya reseptivitas stigma. Oleh karena itu, masa reseptif stigma
A. purpurata dan E. elatior terjadi antara pukul 09.00-12.00. Data ini sejalan dengan Z. spectabile sehingga kemungkinan untuk menyilangkan ketiga spesies ini dapat dilakukan karena masa reseptif terjadi pada waktu yang sama.
Tabel 3. Perubahan pada stigma dan antera Z. spectabile selama bunga mekar
Waktu Kondisi Bunga Kondisi antera Permukaan stigma 08.00 Kuncup Mulai pecah Belum terlihat sekresi 09.00 Kuncup Mulai pecah Mulai terlihat sekresi 10.00 Kuncup Pecah Mulai terlihat sekresi 11.00 Sedikit membuka Pecah Sekresi mulai banyak
12.00 Mekar Pecah Sekresi banyak
13.00 Mekar Pecah Sekresi banyak
14.00 Mekar Pecah Sekresi berkurang
Percobaan penyerbukan silang dilakukan untuk mendukung penentuan masa reseptif stigma. Bunga diasumsikan mulai dapat diserbuki pada pukul 08.00 karena dari antera sudah mulai pecah pada saat itu. Penyerbukan dihentikan pada pukul 14.00 karena sekresi sudah mulai berkurang.
Hasil pengamatan ini menunjukkan bahwa keberhasilan penyerbukan berkisar antara 0.23-0.63 (Tabel 4).
Tabel 4. Jumlah biji Z. spectabile yang dihasilkan pada masing-masing perlakuan waktu penyerbukan silang
Penyerbukan (pkl) Σ bunga
diserbuk Σ buah (%) Σ biji/buah (%) KP
08.00 15 14 (93.3) 29.5 (67.4) 0.62
09.00 15 14 (93.3) 26.9 (61.5) 0.58
10.00 15 13 (86.7) 31.7 (72.3) 0.63
11.00 15 11 (66.7) 23.5 (53.6) 0.36
12.00 15 14 (93.3) 27.5 (62.9) 0.59
13.00 15 9 (48.9) 20.0 (45.7) 0.23
14.00 15 14 (93.3) 24.6 (56.1) 0.52
Keterangan: angka dalam kurung menunjukkan persentase; KP = keberhasilan penyerbukan
Tabel di atas menunjukkan jumlah biji hasil penyerbukan meningkat dari pukul 08.00-10.00 yang berkisar antara 61.5-72.3 % . Pada pukul 11.00 jumlah biji/buah mulai menurun dan pada pukul 13.00 hanya 45.7 % biji yang terbentuk. Berdasarkan persentase pembentukan buah dan biji waktu yang paling tepat untuk penyerbukan berkisar antara pukul 08.00-12.00. Apabila dikombinasikan dengan banyaknya sekresi pada stigma yang dihasilkan, maka dapat ditentukan bahwa waktu yang paling tepat untuk penyerbukan adalah pukul 10.00-12.00. Penyerbukan sebelum pukul 10.00 dapat dilakukan dengan berhasil, akan tetapi untuk alasan praktis tidak disarankan karena pada waktu tersebut bunga masih kuncup. Pembukaan kelopak bunga secara paksa dapat memperpendek kesegarannya sehingga kelopak gugur lebih awal. Walaupun setelah pukul 12.00 penyerbukan juga masih dapat dilakukan dengan berhasil, akan tetapi pada cuaca terik dikhawatirkan polen sudah kurang viabel, sehingga akan menurunkan jumlah biji/buah. Masa reseptif stigma Z. spectabile lebih pendek dibandingkan dengan Alpinia purpurata yang terjadi antara pukul 08.00-12.00 (Oktaviani, 2009).
Persilangan intergenus Alpinia, Etlingera dan Zingiber secara fenologi dapat dilakukan karena masa reseptif stigma dan periode polen viabel yang hampir bersamaan, sehingga kemungkinan mendapat varian baru lebih besar. Kendala utama yang dihadapi dalam persilangan intergenus diduga kekerabatan genetik yang terlalu jauh sehingga biji tidak terbentuk (Yunira, 2009). Oleh karena itu kompatibilitas antar genus perlu diamati lebih jauh.
Keberhasilan Reproduksi
Tipe penyerbukan yang tepat menentukan keberhasilan reproduksi pada suatu tanaman. Penyerbukan yang dilakukan terhadap spesies Z. spectabile ini diantaranya penyerbukan terbuka (alami), penyerbukan sendiri (selfing), dan penyerbukan silang buatan (intraspesies) (Gambar 12). Penyerbukan terbuka tidak menghasilkan biji sehingga ovarium yang tidak berkembang. Perkembangan diameter ovarium pada penyerbukan sendiri dan silang (intraspesies) meningkat tajam pada 3 minggu pertama setelah penyerbukan (Gambar 12), diduga pada periode itu embrio berkembang. Pada minggu ke empat sampai ke sembilan setelah penyerbukan diameter ovarium tidak banyak mengalami pembesaran yang memberikan indikasi periode akumulasi cadangan makanan. Pada minggu ke sepuluh diameter ovarium mengalami pembesaran karena pada saat itu ovarium sudah mulai merekah yang menandakan bahwa biji sudah masak (Gambar 12), diduga pada saat itu masak fisiologis tercapai.
Gambar 12. Perkembangan diameter ovarium Z. spectabile setelah penyerbukan pada tiga tipe penyerbukan
0 0.5 1 1.5 2
2 3 4 5 6 7 8 9 10
Diam
eter Ovary
(cm
)
Minggu Setelah Penyerbukan
Salah satu kendala yang menyebabkan penyerbukan terbuka tidak berhasil (Tabel 5) diduga adalah terbatasnya serangga penyerbuk yang dapat membantu menempelkan polen pada permukaan stigma, karena posisi stigma lebih tinggi daripada antera. Namun pada penelitian sebelumnya (Yunira, 2009), penyerbukan terbuka yang dilakukan pada Zingiber ottensi berhasil membentuk buah dan biji masing-masing 0.1 dan 0.4 dengan tingkat keberhasilan reproduksi (KR) sebesar 0.04. Salah satu faktor yang membedakan keberhasilan penyerbukan terbuka antara Z. spectabile dan Z. ottensi diduga adalah vektor penyerbuk karena morfologi bunga kedua bunga ini sama, yaitu stigma berada di atas antera. Pada Z. spectabile tidak ditemukan adanya serangga yang berperan sebagai vektor sehingga tidak terjadi penyerbukan.
Penyerbukan sendiri pada Z. spectabile (Tabel 5) berhasil membentuk buah dan biji., sedangkan pada A. purpurata, penyerbukan sendiri tidak berhasil membentuk buah, yang memberikan indikasi adanya fenomena self incompatible
(Yunira, 2009). Berdasarkan kondisi tersebut, bunga A. purpurata tidak perlu diemaskulasi terlebih dahulu apabila akan diserbuk silang dengan polen dari bunga lain.
Tabel 5. Jumlah buah dan biji/buah pada masing-masing tipe penyerbukan pada Z. spectabile
Penyerbukan Σ Bunga Σ Buah (%)
Rata-rata Σ Biji/Lokus Σ
Biji/Buah ISI
Diserbuk Pinggir Tengah Pinggir
Alami:
Terbuka 15 0 0 0 0 0
0.92 Buatan:
Sendiri 15 12 (80) 9.3±1.5 6.0±2.5 8.7±0.8 24.0±4.5 Silang 15 13(86.7) 5.0±2.5 3.8±2.1 3.7±2.5 11.7±6.3
benih dan hanya sedikit yang tidak berhasil. Penyerbukan sendiri menghasilkan 24.0±4.5 biji/buah, sedangkan penyerbukan silang menghasilkan 11.7±6.3 biji/buah. Jumlah biji yang dihasilkan dari penyerbukan sendiri lebih banyak daripada penyerbukan silang. Salah satu faktor yang mempengaruhi jumlah biji yang dihasilkan diduga adalah polen yang digunakan. Penyerbukan sendiri menggunakan polen yang berasal dari bunga itu sendiri, sehingga waktu yang diperlukan untuk memindahkan polen ke stigma lebih pendek jika dibandingkan pada penyerbukan silang yang menggunakan polen dari bunga yang berbeda. Menurut Hoekstra (1982) polen tidak dapat bertahan lama, harus segera dikecambahkan setelah terpisah dari antera kecuali diberi perlakuan suhu dan kelembaban yang optimum.
Gambar 13.Perkembangan ovarium Z. spectabile setelah penyerbukan silang: (A) Ovarium membesar (8-10 minggu) (B) Ovarium mulai pecah (10-11 minggu) (C) Biji sudah masak pada ketiga lokus
Persentase keberhasilan penyerbukan silang yang ditunjukkan oleh persentase buah yang terbentuk pada Z. spectabile lebih tinggi dibandingkan dengan A. purpurata yang hanya berkisar antara 43.2-66 % (Yunira, 2009). Hal ini memberikan indikasi bahwa tingkat keberhasilan Z. spectabile yang diserbuk silang lebih besar sehingga peluang untuk meningkatkan keragaman dari spesies ini cukup besar.
Lokus pinggir
Lokus pinggir
Lokus tengah
Tapeinochilos ananassae
Struktur Braktea dan Bunga
Seperti pada Z. spectabile, perkembangan braktea T. ananassae juga dapat dikelompokkan ke dalam lima fase. Fase pertama banyak ditemukan pada bulan Januari, yaitu pada saat braktea mulai membuka dan berwarna merah pucat. Kuncup bunga belum muncul pada fase ini. Panjang braktea rata-rata berkisar 4.74±0.85 cm dengan diameter 5.86±2.15 cm.
Fase dua dimulai pada pertengahan Februari dengan braktea yang semakin membuka sekitar 2-3 baris helaian dan kuncup bunga sudah muncul dari braktea. Fase tiga dari perkembangan braktea T. ananassae ditandai dengan bunga mekar, sehingga penyerbukan dapat dilakukan. Waktu yang diperlukan sekitar tujuh minggu setelah fase pertama, yang jatuh pada pertengahan bulan Maret.
Perubahan braktea pada fase empat tidak terlalu signifikan, yaitu hanya pada ukuran braktea. Ukuran panjang dan diameter braktea berturut-turut mencapai 10.97±1.09 cm dan 9.82±0.57 cm. Pada fase terakhir, yaitu fase lima, braktea sudah mekar penuh. Panjang braktea mencapai 13.74±2.53 cm sedangkan diameternya tidak mengalami perubahan (Tabel 6).
Tabel 6. Karakteristik braktea T. ananassae pada lima fase perkembangan
Fase Perkembangan Waktu Ukuran braktea(cm) Keterangan Panjang Diameter
minggu 5.35±1.02 7.09±1.93
Helaian braktea
minggu 8.63±1.26 9.57±0.62
Dapat dilakukan
minggu 10.97±1.09 9.82±0.57
Bunga sudah
Lama perkembangan braktea mulai fase pertama sampai fase lima sekitar 14-18 minggu. Beberapa braktea mengalami perkembangan yang abnormal, braktea memendek dan membentuk roset. Braktea abnormal ini ditemukan pada tanaman yang mendapat cahaya matahari dengan intensitas yang lebih tinggi dibandingkan dengan yang normal karena paranet mulai rusak pada beberapa bagian. Pertumbuhan memanjang braktea yang terlalu banyak menerima sinar matahari terhambat, dan menjadi abnormal (Gambar 14F).
Gambar 14. Perkembangan braktea T. ananassae: (A) Fase pertama (B) Fase dua
(C) Fase tiga (D) Fase empat (E) Fase lima (F) Braktea abnormal Bunga T. ananassae yang mekar ditandai dengan kuncup yang sudah berkembang hingga keluar braktea (Gambar 15A), yang dapat diidentifikasi sekitar pukul 08.00. Kelopak bunga mulai membuka (Gambar 15B) antara pukul 12.00-13.00 yang diikuti dengan munculnya stigma. Bunga mekar penuh pada keesokan harinya yaitu sekitar pukul 06.00-07.00 (Gambar 15C). Bunga T. ananassae yang mekar akan layu dalam waktu yang cukup lama pada hari yang sama ataupun keesokan harinya.
A B C
Gambar 15. Bunga T. ananassae: (A) Kuncup (B) Mulai mekar (C) Mekar penuh Bunga T. ananassae termasuk bunga lengkap dan bunga sempurna, mempunyai kelopak, struktur mahkota sebanyak tiga helai yang berwarna kuning (yellow orange 17A), satu helai labellum dengan ukuran relatif kecil, pistil, dan stamen. Dua buah antera terletak tepat di bawah stigma (Gambar 16A) dan menempel satu sama lain seolah-olah satu kesatuan. Stilus memanjang dan bersatu dengan ovarium yang terbagi menjadi dua lokus. Jumlah total ovul dalam ke dua lokus bervariasi berkisar 46.4±8.46. Hasil perhitungan jumlah polen dalam
satu antera mencapai ±62 760 (Tabel 7).
Tabel 7. Karakteristik bunga T. ananassae
Variabel Ukuran
Panjang bunga 3.7±0.23 cm
Panjang labellum 1.17±0.08 cm
Lebar labellum 0.44±0.05 cm
Panjang mahkota 1.53±0.08 cm
Lebar mahkota 0.66±0.02 cm
Warna mahkota Yellow orange 17A
Panjang pistil 2.39±0.14 cm
Panjang antera 0.84±0.07 cm
Jumlah ovul/ovarium 46.4±8.46 Jumlah polen/antera
Jumlah helaian braktea
±62 760 132.2±6.7
pengecambah yang terlalu tinggi untuk spesies T. ananassae. Polen T. ananassae
yang paling banyak berkecambah hanya mencapai 0.07 %, yaitu pada pukul 09.00, selebihnya hanya mencapai 0.05 % atau tidak berkecambah.
Tabel 8.Persentase polen T. ananassae yang berkecambah pada tiap waktu pengambilan sampel
No Waktu pengambilan polen (pk) Perkecambahan (%)
1 08.00 0.00
2 09.00 0.07
3 10.00 0.00
4 11.00 0.05
5 12.00 0.03
6 13.00 0.00
7 14.00 0.00
Rendahnya persentase perkecambahan polen T. ananassae pada saat bunga mekar diduga karena polen sudah tidak viabel. Antera sudah pecah pada saat bunga mekar. Pengamatan viabilitas polen sebelum bunga mekar penuh perlu dilakukan untuk menjelaskan hal ini. Ketidak-sesuaian media pengecambah polen juga dapat menjadi penyebab rendahnya perkecambahan polen, karena masing-masing spesies memerlukan media yang berbeda (Warid, 2009). Kondisi lingkungan pada saat bunga mekar juga menjadi faktor yang dapat mempengaruhi viabilitas polen. Viabilitas polen yang dipanen pada saat hari cerah cenderung lebih tinggi dibandingkan dengan polen yang dipanen pada saat hari hujan (Wahyudin, 1999).
Pengamatan viabilitas polen pada Z. spectabile ataupun pada A. purpurata
dan E. elatior (Oktaviani, 2009) menunjukkan bahwa polen viabel antara pukul 09.00-12.00. Dalam pengamatan ini antara waktu tersebut juga menunjukkan viabilitas yang lebih tinggi daripada waktu lainnya, akan tetapi perkecambahan yang sangat rendah tidak dapat digunakan untuk penentuan periode viabilitas polen.
Masa reseptif stigma
mekar, antera sudah mulai pecah pada pukul 08.00, akan tetapi sekresi pada permukaan stigma belum terlihat (Tabel 9). Pada pukul 12.00, bunga sudah mekar, antera sudah pecah dan sekresi pada permukaan stigma mulai banyak. Pengamatan sekresi pada stigma menunjukkan bahwa produksi sekresi meningkat sekitar pukul 12.00-14.00, sehingga waktu tersebut diduga merupakan waktu yang baik untuk penyerbukan, karena pada saat itu stigma reseptif. Data ini perlu dikaji ulang terkait iklim sewaktu pengamatan dilakukan karena bunga ini berbunga sepanjang tahun.
Tabel 9. Perubahan stigma dan antera T. ananassae selama bunga mekar
Waktu Kondisi Bunga Kondisi antera Permukaan stigma 08.00 Mekar Pecah Belum terlihat sekresi 09.00 Mekar Pecah Belum terlihat sekresi 10.00 Mekar Pecah Belum terlihat sekresi 11.00 Mekar Pecah Belum terlihat banyak
12.00 Mekar Pecah Sekresi mulai banyak 13.00 Mekar Pecah Sekresi mulai banyak 14.00 Mekar Pecah Sekresi mulai banyak Penentuan masa reseptif juga didukung melalui percobaan penyerbukan silang. Bunga diasumsikan mulai dapat diserbuki pada pukul 08.00 karena antera sudah mulai pecah pada saat itu. Penyerbukan dihentikan pada pukul 14.00.
Tabel 10. Jumlah biji yang dihasilkan pada masing-masing perlakuan waktu penyerbukan pada T. ananassae
Perlakuan (pk) Σ bunga
diserbuk Σ buah (%) Σ biji/buah (%) KP
08.00 15 2 (13.3) 12.7 (27.4) 0.04
09.00 15 1 (6.7) 2.4 (5.2) 0
10.00 15 1 (6.7) 4.4 (9.5) 0.01
11.00 15 2 (13.3) 7.8 (16.8) 0.02
12.00 15 0 0.0 (0) 0
13.00 15 1 (6.7) 4.3 (9.3) 0.01
14.00 15 0 0.0 (0) 0
Keterangan: Angka dalam kurung menunjukkan persentase, KP = keberhasilan penyerbukan
biji/buah masing-masing adalah 6.7 % dan 9.7 % dengan persentase keberhasilan penyerbukan (KP) berkisar antara 0.01-0.04.
Tabel di atas menunjukkan jumlah biji hasil penyerbukan paling banyak dihasilkan dari penyerbukan pukul 08.00 dengan rata-rata 12.70 biji/buah, sedangkan penyerbukan pada pukul 12.00 dan 14.00 tidak menghasilkan biji. Percobaan penyerbukan menunjukkan hasil yang berbeda dengan pengamatan sekresi pada stigma. Berdasarkan hasil percobaan penyerbukan silang, waktu penyerbukan yang baik adalah pukul 08.00-11.00.
Waktu untuk penyerbukan adalah kombinasi antara masa reseptif stigma dan keberhasilan penyerbukan silang karena viabilitas polen sangat rendah sehingga tidak dapat digunakan untuk menentukan waktu penyerbukan yang baik. Hasil pengamatan stigma menunjukkan bahwa periode stigma reseptif terjadi antara pukul 12.00-14.00, sedangkan berdasarkan percobaan penyerbukan silang, antara pukul 08.00-11.00 berhasil membentuk biji paling banyak. Oleh karena itu penyerbukan akan berhasil apabila dilakukan antara pukul 09.00-11.00.
Keberhasilan Reproduksi
Penyerbukan yang dilakukan terhadap spesies T. ananassae diantaranya penyerbukan terbuka (alami), penyerbukan sendiri (selfing), dan penyerbukan silang buatan (intraspesies). Ovarium mengalami perkembangan (Gambar 18) baik pada penyerbukan alami, penyerbukan sendiri ataupun penyerbukan silang, namun pada penyerbukan alami tidak dihasilkan biji. Ovarium diduga berkembang seiring perkembangan braktea, meskipun tidak terjadi pembuahan.
Gambar 18. Perkembangan diameter ovarium T. ananassae setelah peyerbukan 0.6
0.7 0.8 0.9 1
silang sendiri terbuka
Diam
eter Ovarium
(cm
)
Tipe Penyerbukan
Ovarium pada bunga dari penyerbukan terbuka tetap segar dan membengkak sampai akhir pengamatan, akan tetapi biji tidak terbentuk (Tabel 11). Demikian halnya dengan bunga yang diserbuk sendiri dan silang yang tidak menghasilkan biji. Hasil pengamatan ini memberi indikasi terbentuknya buah partenokarpi, yaitu buah yang terbentuk tanpa melalui proses fertilisasi sehingga tidak terdapat biji karena biji yang sedang tumbuh menjadi rusak pada tahap awal perkembangannya (Widajati et al., 2008).
Tabel 11. Jumlah buah dan biji yang terbentuk dari tiap tipe penyerbukan pada T. ananassae
Penyerbukan Σ bunga diserbuk
Σ buah (%)
Σ biji/lokus Σ
Biji/buah ISI Kiri Kanan
Alami:
Terbuka 15 0 0 0 0
1 Buatan:
Sendiri 15 2(13.3) 2.7 2.8 5.5
Silang 15 2(13.3) 3.2 4.1 7.3
Persentase pembentukan buah pada penyerbukan sendiri sama dengan penyerbukan silang, yaitu 13.3 % sehingga intensitas inkompatibilitas berdasarkan
index of self of incompatibility (ISI) sebesar 1. Data ini mengelompokkan
T. ananassae ke dalam kategori completely self compatible. Penyerbukan silang menghasilkan sekitar 7.3 biji/buah dan penyerbukan sendiri menghasilkan biji sekitar 5.5 biji/buah (Gambar 19).
Gambar 19. (A) Ovarium yang mulai pecah ((B) Biji sudah masak (C) Butiran biji T. ananassae berwarna hitam
Keberhasilan penyerbukan T. ananassae baik penyerbukan sendiri ataupun silang hampir sama, sehingga upaya perbanyakan melalui biji dari penyerbukan
sendiri ataupun upaya peningkatan keragaman dari penyerbukan silang memiliki peluang yang sama. Oleh karena itu tanaman ini baik untuk dikembangkan dan diteliti lebih lanjut.
Rendahnya biji yang terbentuk diduga karena viabilitas polen yang rendah karena antera sudah pecah sejak bunga mekar pada pagi hari. Upaya untuk meningkatkan viabilitas polen perlu dilakukan agar produksi biji meningkat. Dari struktur morfologi T. ananassae memungkinkan untuk menyerbuk sendiri. Data penyerbukan menunjukkan bahwa spesies ini completely self compatible, sehingga seharusnya produksi biji secara alami tinggi. Tidak terbentuknya biji dari penyerbukan terbuka perlu dikaji lebih lanjut, yang diduga juga karena rendahnya viabilitas polen dan terbatasnya vektor polen.