Keragaman Dan Pemetaan Penyebab Bulai Jagung Di 13 Provinsi Indonesia
Teks penuh
Gambar
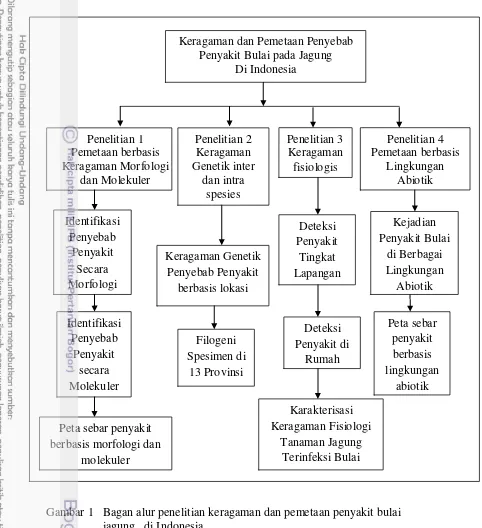


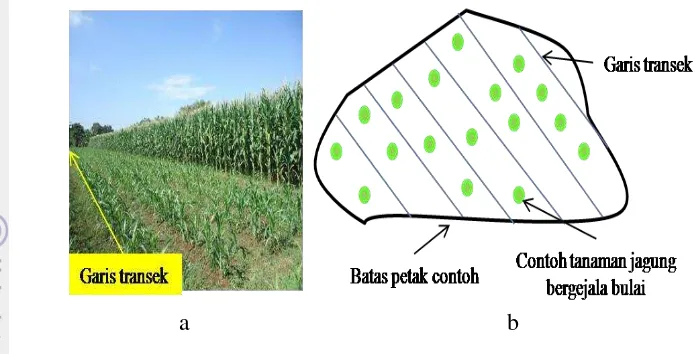
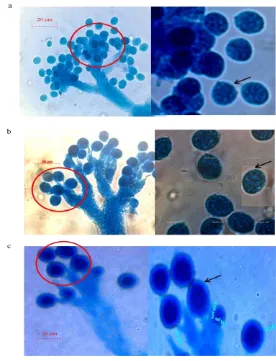
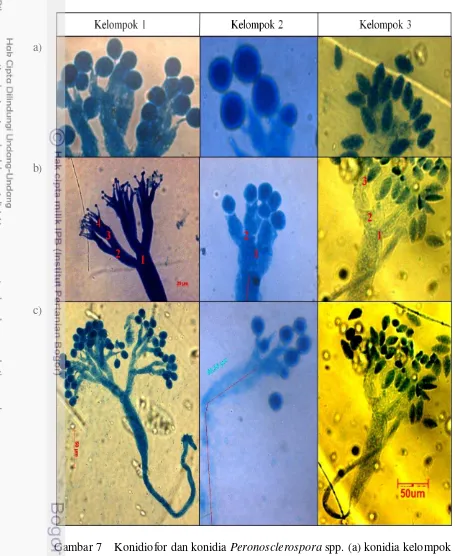

Dokumen terkait
Berdasarkan hasil penelitian ini diketa- hui bahwa penyakit bulai telah tersebar luas pada tanaman jagung di semua kabupaten yang di Pulau Madura yaitu
Pengaruh Penggunaan Beberapa Varietas dan Aplikasi Pseudomonas fluorescens untuk Mengendalikan Penyakit Bulai (Peronosclerospora maydis) pada Tanaman Jagung (Zea mays
Hasil kajian menunjukkan: (1) di lokasi Kediri penyakit bulai menurunkan produksi 12-80%, sedangkan di lokasi Pasuruan penyakit bulai menurunkan produksi 20-38%; (2) meskipun
Salah satu kendala dalam budidaya jagung adalah penyakit bulai yang disebabkan Peronosclerospora maydis Penelitian ini bertujuan untuk mengetahui pengaruh berupa
Pengaruh Paenibacillus polymyxa dan Pseudomonas fluorescens dalam molase terhadap keterjadian penyakit bulai (Perenosclerospora maydis L.) pada tanaman jagung manis.. The
Hasil penelitian hubungan faktor lingkungan terhadap laju infeksi dan pola sebaran penyakit bulai pada tanaman jagung di kabupaten jombang adalah sebagai
Skripsi dengan judul : Pengelompokan Genotipe Jagung Berdasarkan Tingkat Ketahanan terhadap Penyakit Bulai (Peronosclerospora maydis) dan Daya Hasilnya, telah diuji
Penelitian ini bertujuan untuk menentukan karakteristik morfometri dan tingkat patogenisitas strain Peronosclerospora yang diambil dari beberapa lokasi sentra produksi jagung
