ALLAH
g m
Menganjurk mmusia unfuk memperha
fikan
alam
r a k
langif, bumi, bbinfang-bintang, &af, lauf, tumbuhan,
bina-,
dsn
manusia ifu sendin'. Me1almUIperhafifian
tersebut manusia
ak;ul
menakpaf manfast h2ganak,
p ' f u
menyadazi kebesman
dan
keagungan
lkhan
dsn
P
PEANUT STRIPE VIRUS STRAiN INDONESIA:
VARlASl BIOLOGI, DETEKSI MOLEKULER, PENGKLONAN,
DAN DETERMINASI URUTAN NUKLEOTIDA 3'GENOM RNA
PStV, SERTA ANALISIS KERAGAMAN DAN FlLOGENETlKA
BERDASARKAN GEN
CP
DAN 3'UTR
Oleh
HASRI'ADI MAT
AKIN
.
PROGRAM PASCASARJANA
INSTITUT PERTANIAN BOGOR
RINGKASAN
HASRIADI MAT
AKIN.Peanut Strip
Virus
Strain Indonesia: Variasi Biologi,
Deteksi Molekuler, Pengklonan, dan Determinasi Urutan Nukleotida
3'Getlorn
RNA
PStV, serta Analisis Keragaman dan ' ~ i l o ~ e n e t i k a
Berdasarkan Cen
CP
dan
3'UTR
pi
bawah bimbingan Prof. Dr. Tr. Edi Guhardja, MSc sebrgai
ketua, Dr.
Ir.
Sudarsono,
MSe,
Prof. Dr.
Ir.
Rusmlah Suseao
MSc,
Dr?
Ir.
Hajrial Aswidineoor, MSc, Dr. 1r. Suharsono sebagai anggota]
Hasil kacang tanah di Indonesia tahun 1996 adalah 746.600 ton dengan luas
panen 696.600 Ha (1.07 tonha) (BPS, 1996). Di negara-negara penghasil kacang
tanah lain, seperti Korea Selatan, Jepang, dan RRC hasil rata-rata perhektar berturut-
turut adalah 1.77 ton, 1.72 ton, dan 1.92 ton (Xu, 1992). Rendahnya produktivitas
kacang tanah di Indonesia antara lain disebabkan serangan virus. Di Indonesia, PStV
p'aliig dominan menyerang kacang tanah dibandingkan dengan virus-virus yang lain.
Penelitian ini bertujuan: (1) mengisolasi PStV dari 12 propinsi di Indonesia, (2)
mengetahui variasi biologi isolat-isolat PStV berdasarkan simptomatologi
dan
patogenisitas beberapa kultivar kacang tanah, (5) mengembangkan metode. deteksi
molekuler, (4) melakukan pengklonan dan determinasi urutan nukleotida isolat-isolat
PStV dari berbagai daerah di Indonesia, dan (5) menganalisis keragaman dan
filogenetika berdasarkan urutan nukleotida gen CP, ?'UTR, serta urutan asam amino
CP PStV
Has2 isolasi virus menunjukkan dari contoh daun kacang tanah yang bergejala
belang yang dikumpulkan dari 12 propinsi di Indonesia semuanya terinfeksi oleh
virus. Semua isolat virus yang didapat menimbukan gejala lesio lokal pada inokulasi
mekanik pada
C.
amaranficoIor
dan
C.
quinoa.
Dan contoh daun tanaman kacang
tanah tersebut diperoleh 15 isolat virus yang merupakan satu isolat untuk rnasing-
masing propinsi kecuali untuk Propinsi Jawa Barat empat isolat.
HasiI penelitian ini menunjukkan isolat-isolat PStV mempunyai variasi biologi
tanah. Berdasarkan tipe gejala penyakit pada daun kadang tanah yang terinfeksi, 15
isolat PStV dikelompokkan nierijadi
5kelompok, yaitu:
mildmottle, hlotch, severe
bk>/ch slripe, chkcrolic ring mollle, dan
slrip.Berdasarkan patogenisitasnya, isolat-isolat PStV dikelompokkan menjadi isolat
kuat, sedang, dan kuat. Hasil studi
ini
menunjuklcan tiga faktor yang mempengdruhi
patogenisitas PStV, yaitu: faktor genetika PStV (asal isolat) dan genotipe kacang
tanah (kultivar), serta lingkungan (musim tanam).
Pengamh asal isolat & t ~
ditunjukkan oleh adanya keragaman respoti kacang tanah yang diinokulasi dengan
berbagai isolat PStV. Sedangkan pengamh kultivar terhadap patogenisitas isolat PStV
diamati dengan adanya perbedaan respon kedua kultivar Landak dan Gajah terhadap
infeksi isolat PStV. Patogenisitas isolat-isolat PStV juga dipengaruhi oleh kondisi
Iingkungan berdasarkan perbedaan patogenisitas isolat-isolat PStV pada musim hujan
dan musim kernarau.
PStV-TPSl yang termasuk kelompok mild mottle mempunyai tingkat
patogenisitas sedang pada kultivar
Landak
dan Gajah. Isolat-isolat yang termas.uk
kelompok blotch, blotch stripe, severe hlotch stripe, chlorotic ring-mode, dan stripe
umurnnya merupakan isolat kuat pada kultivar Landak dan Gajah, kecuali IPS12
merupakan isolat sedang pada kultivar Landak; sebaliknya TPS6, TPS7, dan TPS13
menjadi isolat sedang pada kultivar
Gajah.
Analisis keragaman pada tingkat urutan nukleotida gen CP, asam amino CP,
dan
3'UTR
menunjulckan isolat-isolat PStV yang berasal dari 12 propinsi di Indonesia
mempunyai keragaman yang rendah walaupun isolat-isolat tersebut mempunyai
karakter gejala
dan
patogenisitas yang berbeda. Hal
ini
kernungkman disebabkan gen
CP tidak b h n g s i sebagai pengendaii karakter gejala pada PStV.
Analisis flogenetika gen CP PStV yang berasal dari Indonesia, Thailand
,
dan
USA mengindikasikan PStV di wilayah tersebut berasal dari nenek moyang yang sama.
Pemisahan strain-strain menjadi tiga kelompok mc~rtc~phyIelic
menunjukkan PStV
sudah ada pada wilayah tersebut sejak wale yang sangat lama. Karalrter tetua PStV
hubungan antara PStV dengan subkelompok BCMV mhs;ldiiasikan salah satu dari
virus tersebut dapat menjadi caloti tetua.
Strain-strain PStV yang berasal dari Indonesia merupakan rekombinasi antara
PStV dengan BlCMV yang tejadi pada daerah CP13'UTR. Rekombinasi antara PStV
dan BlCMV dapat terjadi pada tanaman inang yang mengalami infeksi campuran. Hasil
pengamatan pada urutan nukleotida isolat-isolat PStV dari Indonesia menunjukkan
kejadian rekombiiasi
RNA
PStV dengan BlCMV pada posisi basa ke 9764
*(38nukleotida bagian hulu kodon stop).
Berdasarkan jarak genetika dari masing-masing isolat PStV Indonesia diduga
Bogor dan Malang merupakan daerah asal penyebaran PStV di Indonesia. Isolat
Kalimantan Tengah (PStV-PS14) m e ~ p d c a n
satu-satunya PStV yang berasal dari
Malang, sedangkan isolat-isolat yang lain berasal dari Bogor. Variasi morfologi gejala
penyakit yang besar dari isolat-isolat PStV yang berasal dari Bogor (IPS2,3,4,15) juga
genunjang dugaan tesebut yang menetapkan Bogor sebagai salah saki daerah asal
penyebaran PStV di Indonesia.
Sedangkan
penetapan Malang yang juga merupakan
daerah asal PStV didukung oleh hasil analisis tilogenetika yang menunjukkan bahwa
isolat
Milan6
(PStV-lb) dan isolat Kalimantan
Tengah
(PStV-IPS14) membentuk
garis evolusi yang berbeda dengan isolat-isolat PStV indonesia yang lain.
Deteksi menggunakan teknologi RT-PCR dapat
digunakan
untuk mendeteksi
PStV dalam daun kacang tanah dan vektor
A.
craccipra.
Hibridisasi dot blot dengaff
pelacak cDNA gen CP PStV yang dilabel dengan
digoxigenin(Ihg-cDNA) juga dapaf
PEANUT STRIPE VIRUS
STRAIN INDONESIA:
VARlASl BIOLOGI, DETEKSI MOLEKULER, PENGKLONAN,
DAN DETERMINASI URUTAN NUKLEOTIDA 3'GENOM RNA
PStV, SERTA ANALISIS KERAGAMAN DAN FILOGENETIKA
BERDASARKAN GEN
CP
DAN 3'UTR
*oleh
HASRIADI
MAT
AKIN
Disertasi sebagai salah satu syarat untuk mernperoleh gelar
Doktor
pada
Program Pascasa jana, lnstitut Pertanian Bogor
Program Studi Biologi
PROGRAM
PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
J U ~ U I
:
PEANUT
STRIPE
VIRUS
STRAJN
INDONESIA:
VARWI BIOLOGI,
DETEKSI
MOLEKULER,
PENGKLONAN,
DAN
DETERMINASI
URUTAN
NUKLEOTIDA
3' GENOM
RNA PStV,
SERTA
ANALISIS
KERAGAMAN
DAN
FILOGENETIKA BERDASARKAN
GEN CP
DAN
3'UTR
Nama Promovendus
:
Hasriadi Mat
Akin
IP
N R P
:
94526
Program Studi
:
Biologi
Menyetujui
1. Komisi Pembimbing
Prof.Dr.lr. Edi Guhardia,
MSCKetua
--
Dr.lr.Sudarsono. MSc
Prof. Dr. Ir. Rusmilah Suseno. MSc
Anggota
Anggota
Dr.lr. Hairiat Aswidinnoor. MSc
RIWAYAT HIDUP
Penulis adalah anak ke empat dari empat bersaudara yang dilahirkan pada tanggal 29
Juni 1957 di Kemantan Kebalai, Kabupaten Kerinci. Orang tuanya adalah H. Datuk Mat
Akin (almarhurn) dan Hj. Timatun Sawiyah. Pada tahun 1982 menikah dengan Urip Mulyati
dan dikaruniai empat orang putrali, Yulia Rahma Fitriana (15 th), Chandra Prasetyo Hadi (I I
th), Muhammad Yogie Fadly (5 th), dan HasriI Mulya Budiman (1 th).
Pendidian dasar dan menengah diselesaikan di Kabupaten Kerinci. Pada tahun 197 1
PenuBs menyelesaikan S D Negeri Kemantan Kebalai; pada tahun 1974 menyelesaikan SMP
Negeri Semurup; dan pada tahun 1977 menyelesaikan SMA Negeri Sungai Penuh Kerinci.
Pada tahun 1978 Penulis diterima sebagai mahasiswa IPB rnelalui Proyek Perintis 11 clan
tahun 1982 Penulis memeperoleh gelah Sarjana Pertanian dari Fakultas Pertanian IPB. Tahun
1994 Penulis mernperoleh gelar Magister Sains dalam bidang Fitopatologi dari P r o s a m
Pascasarjana Universitas Gadjah Mada, Yogyakarta dengan yudisium Cum Laude. Pada tahun 1994 panulis mernperoleh beasiswa dari Proyek Beasiswa Unggulan Pascasarjana
Dalam Negeri Proyek URGE (University Research for Garaduate Educarion) untuk mengikuti Program Doktor (53) di Program Pascasarjana IPB, Program Studi Biologi dengan
kekhususan Biologi Molekuler dan Bioteknologi. Tahun 1998 Penulis mengikuti program
San&vich untuk penelitian disertasi di Queensland Agi-iculrural Biotechnology Centre
(QABC) the University of Queensland, St. Lucia, Brisbane, Australia. _
Tchun 1982- 1983 Penulis beke j a sebagai Kepala Proyek Managemen Unit Proyek
Pembinaan dan Pengembangan Wilayah Transmiaasi (PMU-P3W Transterpadu) Teluk
Dalam Kalimantan Timur. Tahun 1983- 1984 bekerjz pada Direktorat Perlindungan Tanaman
Pangan Departemen Pertanian Jakarta. Tahun 1994-1986 Penulis pekerja sebagai Asisten
Kebun pada PT. Perkebunan X (sekarang PTPN VII). Bandar Lampung yang di tempatkan di
Perkebunan Kelapa Sawit Bekri. Sejak tahun 1956 Penulis rnenjadi staf edukatif pada
UCAPAN TERIMA KASIH
Puji syukur dipanjatkan kepada Allah SWT atas rahmat dan hidayah-Nya
Penulis dapat menyelesaikan penelitian clan penulisan disertasi ini.
Disertasi berjudul
Peanul
SZriipe
VuusStrain Indonesia: Variasi
Biologi, Deteksi molekuler, Pengklonan dan Determinasi Urutan
Nukleotida
3'Genom
RNA
PStV, serta Analisis Keragaman dan
Filogenetika Berdasarkaa Gen CP dan 3'UTR
disusun berdasarkanperrelitian yang dilaksrrnakaa di Laboratorium Biologi Motelder, Laboratorium Pusat Studi Pemuliaan Tanaman Jurusan Budidaya Pertanian, Rumah Kaca Jurusan Tanah
Fakultas Pertanian IPB, dan QueensZand AgricuIturaI Biotechnology Centre (QABC),
Department Primery Industry and Energy, Australia
Sebagian dari hasil penelitian
ini
telah d i p u bpada ~ ~Indonesian Journal o f Tropical Agriculture dan dipresentasikan pa& Indonesian Biotechnology Conference di Jakarta, 17-19 Juni 1997, dan Kongres Nasional Biologi di BandarLampung, 24-26 Juli 1997. Saat imi sedang dipmiapkan tiga tulisan dari has3
penelitii ini dan akan d i p u b b i k a n pada jumal yang telah bedaidiiasi
Selama penelitian dan penulisan disertasi ini Pen& mendapatkm pengarahan dan bhbhgau dari Tim Komisi P e m b i i . Oleh sebab ity Penulis mengucapkan terkna kasih kepada Prof. Dr. Ir. Edi Guhardja sebagai ketua komki,
Dr.Ir. Sudarsrzno, MSc, Prof. Dr. Ir. RusmiIah Suseno, Dr.Ir. HajriaI Aswidhumor,
MSc, d m Dr-Ir. Suharsono masing-masing sebagai anggota komisi atas bimbingan
dan
pengarahannya, dan kepada Dr. RG. Dietzgen sebagai pimpinan QABCdan
Dr.Collen M. Hi* sebagai Biotechnologist yang telah memberi kesempatan dan
b i i i a n selama melakukan penelitii di Australia.
Ucapan terima kasih juga disampaikan kepada Rektor dan Direktur Program
Pascasarjana IPB atas kesempatan mengikuti program doktor di IPB; kepada Rektor
Universitas Lampung dan Dekan Fakdtas Pertanian yang telah memberi izin untuk menghtti program doktor di IPB. Ucapan terima kasih yang sama disampaikan
kepada Pemimpin Proyek URGE (University Reseach for Graduate Education)
melafui Beasiswa Unggulan Pascasarjana Dalam Negeri, Program Sanmvich untuk melakukan penelitii di Australia, dan Hibah
Ti
di bawah PI Dr.Ir. Sudarsono, MSc,Proyek AClAR (AustraZian Centre for Infernational Agricultural Research), dan P T .
Rasa terima h i h juga disampaikan kepada rekan-rekan mahsiswa dim teman- ternan di Laboratorium Biologi Molekuler Jurusan Budidaya Pertanian IPB yaitu: Ir.
Edy Irwansyah atas bantuan untuk menata slide proyektor dalam ujian terbuka, Ir.
Edy Batara Mulya Siregar, MSi Ir. Gustian, MS, Ir. Saloon Sinaga, MSi, Ir.
Ramadiayana, MSi, Ir. Elok Dwi Sulichantini, MSi,
Ir.
A k q Edi, MSi, Ir.Di
Dinarti, Ir. Yudha Hartanto, MSi, Ir. Kikin Hamzah Mutaqin, Ir. Rustikawati, MSi, Ir.
Catur Horizon, MSc, Ir. WaM Dinarto, MSi,
Ir.
Selvi Turnbelaka, MSi, Ir. Karskh, Ir. Sudirman Numba, MS,Ir.
Semuel Runtunuwu, MS, Yudiasyah, dan Didik atasbantuan clan kerja samanya s e b Penulis melakukan penelitian. Rasa terima kasih
juga disampaikan kepada rekan-rekan selama penelitian di QABC Austrdh yaitu:
Ir.
Dwi Hapsoro, MSc,
Ir.
Sholeh Avivi, MSi, Rhormda, Philp, Margaret, Sadeep, dan Ir.Emy Sulktiowati, MAgr atas bantuan
dan
kerja m y aRasa kagum
dan
hormat khusus dkampakm kepada kedua orang tua Penulis Hj. T i i t u n Sawiyah dan H. Mat Akin (almarhum) yang telah mencurahkan kasihsayang, mernbhbiing, mendo'akan
dan
mengantarkan anak-anaknya sampai mampuhidup mandiri. Penulis myadari tanpa dedikasi dan kerja keras beliau Penulis tidak mungkin dapat myelesaikan pedidikan sethggi
ini
Ucapan terima kasih juga disampaikan kepada ketiga kakak UrusLani, Djuf?i Mat Akin, SH, dan Zubaidah yang telah memberi dorongan moril dan materil selama Penulis memmtut ilmu.Kepada isteri
dan
aaak-anakk&
(Ana, Can, Ogi, clan Acil) yang se& mendampingi Penulis selama mengkuti tugas belajar di S2dan
53 juga diucapkan banyak terima kasih atas kesabaran dan k-a Semoga keberhashn ini menjadi kebanggaandan
pemicu untuk &pat berbuat yang lebih baik lagi di masa- masa yang akan datang.Akhirnya, diiringi do'a semoga seluruh keg* stud ini dapat bernilai ibadah dihadapan
AUah
SWT, baik bagi Penulis ~naupun semua phdc yang telah raembantu moril maupun materil selama penelitian dan penulisan dkxtasi Semoga hasidpenelitian ini dapat didayagudcan untuk kemajuan umat manusia
dan
h u pengetah-DAFTAR IS1
KINGKASAN ... ii
...
UCAPAN TERIMA KASM vii
DAFTARISI ... x . . .
DAFTAR TABEL xi
... ...
DAFTAR GAMB AR XIII
. . .
Latar Bdakang 2
. . .
Tujuan Penelitian 5
TINJAUAN PUSTAKA ... 7
. . .
Kacang Tanah
.
7A
Patogemsitas PStV ... 8 ...
Biologi Molekuler PStV 10
...
Variasi Molekuler Potyvirus 12
ISOLASI PStV DARI DAUN KACANG TANAH
YANG BERASAL DART 12 PROPINS1 DI INDONESIA ... 15
Pendahutuan ... 16
Bahan clan Metode ... 16
Hasil ... 21 Pembahasan ... 24
METODE DETEKSI DAN IDENTIFKASI ISOLAT-ISOLAT PStV
DENGAN TEKNIK RT-PCR DAN H I B m I S A S I DOT BLOT ... 26
... ...
Pendahuluan 27
...
Bahan dan Metode ... 28
Hasil . . . . . . 36
Pembahasan ... . . . 44
PATOGENISITAS BERBAGAT ISOLAT PStV PADA
...
B E B E W A KULTNAR KACANG TANAH 47
...
Pendahuiuan
...
Bahan dan Metode
...
Hasil
...
Pembahasan
...
..
KERAGAMAN ISOLAT-ISOLAT PStV BERDASARKAN
...
SIMPTOMATOLOGI PENYAKIT
...
Pendahuluan
...
Bahan dan Metode
...
Hasil
...
..
...
Pembabasan
PENGKLONAN DAN DETERMINASI URUTAN I*TUKLEOTIDA 3'GENOM RNA PStV SERTA ANALISIS K E R A G M DAN
FLOGENETIKA PStV DALAM SUBGROUP BCMV
...
Pendahuluan
...
Bahandan
Metode...
Hasil...
Pembahasan
...
..
...
.
.
.
...
KESTMPULAN DAN SARAN
...
DAFTARTABEL
1 . Protein-protein yang disintesis oleh genom PStV dan potyvirus lainnya
dalam sel tanaman inang..
.
. . ..
. . ..
. . . 1 32 Gejala infeksi beberapa
. .
virus yang menyerang kacang tanah padatanaman mdkator
.
.. . .
. . .. . .
. . . ..
. . . . .. .
. . .. .
. . .. . .
.. . .
..
. 173. Gejaia pada tanaman indikator yang digunakan untuk isolasi PStV.. . .
.
. . . . 224. primer oligonukleatida yang digunakan untuk deteksi PStV . . . 28
5. Kehilangan hasil kacang tanah kultivar Gajah akibat infeksi PStV
di berbagai lokasi di Indonesia.. . .
. . .
. . ..
..
. . ..
- . . . 496 . Patogenisitas isolat-isolat PStV pada dua kutivar Gajah dan Landak.. . . . 57
7. Patogenisitas isolat-isolat PStV pada kacang tanah kultivar Gajah
yang ditanam pada musim hujan dan kemarau.. . . 5 8
8. Pengaruh infeksi PStV terhadap pertumbuhan vegetatif kacang tanah
kutivar Gajah dan Landak..
.
. . . 599. Patogenisitas PStV pada m u s h hujan dan kemarau berdasarkan bobot
..
*
biji kermg.. . . 61
10. Pengelompokan isolat-isolat PStV berdasarkan gejala penyakit pada
kacang tanah kultivar Lamndak . . . 68
11. Gejala infeksi isolat-isolat PStV pada beberapa tanaman inang.. . . 7 0
12. Primer yang digunakan untuk determinasi -tan nukleotida cDNA 3'
Genom RNA PStV
. .
.. . .
.. . .
.. . .
.. .
..
..
. . ..
. . .. . .
. . . 7 813. Beberapa spesies subkelompok B C W yang digunakan dalam analisis
keragarnan dan filogenetika . .
.
. . ..
. . . S O14. Hasil seielisi klon rekombinan dengan teknik PCR.. . . . . . . . . 84
16. Variasi perbedaan antara PStV yang berasal dari Indonesia dengan
strain-strain dari BCMV.. ... 93
17. Persentase kesamaan urutan nukleotida gen CP dan asam amino CP
diantara isolat-isolat PStV yang berasai dari berbagai daerah di Indonesia.. . . 100
18. Persentase kesamaan urutan nukleotida 3'UTR diantra isolat-isolat PStV
yang berasal dari berbagai daerah di Indonesia.. . . 10 1
19. Persentase kesamaan urutan nukleotida gen CP dan asam amino CP antara
. . .
PStV strain Indonesia dan subkelornpok BCMV 102
19. 20. Persentase kesamaan urutan nukleotida 3'UTR antara
PStV strain Indonesia dan subkelompok BCMV ... 103
21. Korelasi antara gejda dan patogenisitas isolat-isolat PStV pada kuitivar
DAFTAR
GAMBAR
1 Skema tahapan penelitian yang dilalcukan . . . 6
2 Partikel PStV berbentuk tuk batang Ientur (f7rxirnrs rod) dan
(B) badan inklusi berbentuk cakra (pimuheel). . . .
3 Peta genom PStV dan protein-protein yang disintesis dari genom
RNA PStV ... 12
4 Peta asal isolat-isolat PStV dari 12 propinsi di Indonesia.. ... 19
. . .
5 Tahap-tahap isolasi PStV dari contoh d mkacang tanah dari lapang 23
6 Posisi primer pada genom RNA PStV dan cDNA berukuran 234 bp
dan 1,2 Kb hirsil amplifikasi RT-PCR . . . 29
7 Posisi hibridisasi pelacak PStV pada genom RNA PStV untuk deteksi PStV dengan hibridisasi dot blot.. ... 3 3 8 Spesifisitas deteksi RT-PCR mengynakan pasangan primer
PSTl& PST2, dan pasangan PSTI&PST4.. ...
9 Sensitifitas deteksi PStV dengan teknik RT-PCR menggunakan primer
PSTl d m PST2.. ... 3 7
1 0 Kondisi sampel sebelum dideteksi RT-PC dengan primer PSTl dan PST4 3 9
1 1 Deteksi PStV dari vektor (Aphis craccivora) dengan RT-PCR primer
PSTldanPST2 ... 39
12 Hasil amplifikasi cDNA 234 bp isolat-isolat PStV dari beberapa daerah di Indonesia dengm teknik RT-PCR menggunakan pasangan
primer PSTl dan PST2 dengan cetakan total RNA daun kacang tanah
yang terkSeksi PStV ... 30
13 Has3 deteksi isolat-isolat PStV pada daun kacang tanah dengan
teknik hibridisasi pelacak PStV.. . . . 42
15 A. Deteksi PStV pada benih yang didapatkan dari tanaman induk yang terinfeksi PStV dari lapang; B. Deteksi PStV pada benih yang didapatkan
dari tanaman induk yang terinfeksi PStV di laboratonurn.. . .
15 Persentase kandungan klorofil total dan bobot biji kering per tanaman kacang tanah kultivar Biawak dan Macan yang terinfeksi
isolat-isolat PStV dibandingkan dengan kontrol tanaman sehat ...
17 Persentase bobot biji, polong, dan berangkasan kering, serta jumlah polong pertanaman dibandingkan dengan kontrol kultivar Gajah yang terinfeksi isoiat-isolat PStV yang &tanam pada musim hujan dibandingkan dengan kontrol tanaman sehat ...
1 8 Persentase bobot biji, polong, dan berangkasan kering, serta jumlah polong pertanaman dibandingkan dengan kontrol kultivar Gajah yang terinfeksi isolat-isolat PStV yang ditanam pada musim kemarau dibandingkan dengan kontrot tanaman sehat.. ...
19 Persentase bobot biji polong, dan beritngkasan kering, serta jumlah polong pertanaman dibandingkan dengan kontrol kultivar Landak yang terinfeksi isolat-isolat PStV yang ditanam pada musim kemarau diband'igkan dengan kontrol tamanan sehat.. ...
20 Tipe gejala penyakit pada kacang tanah kulivar Landak akibat infeksi isolat PStV dari berbagai daerah di Indonesia ...
21 Dendogam keragaman isolat-isolat PStV dari berbagai daerah di Indonesia berdasarkan karakter simtomatologi penyakit. . .
22 Amplifikasi RT-PCR cDNA 1,2 Kb dari genom RNA PStV dengan primer PSTI dan PST4.. ...
23 Plasmid vektor @GEM-T Easy) yang di@an untuk rnengldon cDNA 1.2 Kb dari 3' genom RNA-PStV. . .
24 Hasil amplifikasi RT-PCR dengan primer PSTl dan PST2 dan hasil pemurnian cDNA 1,2Kb.. ...
2 5 Proses kIoning dan seleksi klon rekombinan pada media seleksi yang dapat menimbulkan wama putih untuk klon rekombinan dan biru untuk klon yang
. . .
26 Hasil amplifikasi cDNA 1.4 Kb dari klon E. cnli DH5a yang membawa plasmid rekombinan (pGEM-T::CP-PStV) dengan teknik PCR
menggunakan primer T7 dan SP6.. . . .. . . . . . .. . . ..
27 Hasil pernotongan plasmid rekombinan dengan E w R 1 . . . . . . . . . . . . .
28 Urutan nukleotida 3' genom RNA-PStV isolat PStV-TPS2 dan prediksi urutan asam amino protein NIb dan protein selubung PStV.. . .
.
. . .29 Pohon filogenetika PStV (Neigbmzr .Joining Tree) berdasarkan urutan nukleotida sen CP . . . . . . . . . . . . . . . . .
30 Pohon filogenetika PStV (Neighmrr Joining Tree) berdasarkan urutan
asam amino CP.. . .
.
. . .. .
.. . . .
. .. .
. . .. . ..
. . .. . .
. . . . ... .
. . . .. . ... .
. ..
. . .. ..3 1 Parsimnni Wee subkempok B C W berdasarkan urutan nukleotida gen CP.
32 Pohon filogenetika ( N e i g h r Joining Tree) subkelompok BCMV
berdasarkan urutan nukleotida gen CP.. . .
33 Pohon filogenetika (Neighmr Joining Tree) subkelompok BCMV
berdasarkan urutan asam amino C P . . . . . .. .
.
. . .34 Pohon filogenetika subkelompok (Neighmr Joining Tree) subkelompok
BCMV berdasarkan urutan nukleotida CP/3'UTR.. . . .
.
..35 Model pindah silang antara R N A BlCMV dengan R N A PStV rnembentuk PStV rekombinan.. . .
.
. . . ... .36 Analisis multipasangan urutan nukleotida gen CP subkelompok B C W
37 AnaIisis multipasangan urutan asam amino CP CP subkelompok B C W .
3 8 Analisis multipasangan urutan nukleotida CP/3 'WTR subkeIompok BCMY
3 9 Urutan nukleotida RNA ribosorn kacang tanah hasil amplifikasi RT-PCR yang tidak spesifik dengan primer PSTl dan PST4
PENDAHULUAN
Latar Belakang
Kacang tanah (Amachis w g a e a L.) merupakan salah satu dari enarn
komoditas terpenting di dunia. Sebagai tanaman kacang-kacangan sumber protein dan
lernak nabati, kacang tanah menempati posisi kedua setelah kedelai. Biji kacang tanah
dietahui mengandung protein (25-35%), lemak (43-55%), nikotimin, thiamin, dan
vitamin E (Ashley, 1984; Cooper-Bland et al., 1992). SeIain dapat dionsumsi
Iangsung, kacang tanah juga digunakan sebagai bahan baku industri makanan.
I n t e d k a s i dan ekstensifikasi pertaoian merupakan usaha-usaha yang dapat
ditempuh untuk peningkatan produksi kacang tanah. Di Indonesia us&-usaha
intensijjlcasi penanaman kacang tanah &pat menyebabkan timbulnya cekarnan biologi
dan meningkatnya laju epiderni penyakit sehingga menurunkan hasil panen. Sebaliknya
ekstens- pada lahan-lahn marjinal menghadapi kendala cekaman lingkungan.
Untuk itu diperlukan dukungan yang berupa penemuan varietas u n g y l berdaya hasil
tinggi tahan terhadap hama dan penyakit, atau toleran terhadap cekaman -an.
Perrnintaan kacang
tanah
di Indonesia dalam dasawarsa terakhir ini terusmeningkat dim diperkirakan pada tahun 2000 akan mencapai 1,9 juta ton (Kasno,
1993). Hasil k a a g tanah di Indonesia pada tahun 1996 adalah 746.600 ton, luas
p m 696.600 ha, dan harif rata-rata 1,07 ton/ha (BPS, 1996). Hasil rata-rata ini
mas* tergolong rendah b i dibandigkan dengan hasil rata-rata di negara-negara
penghasil kacang tanah lainnya, seperti Korea Selatan (1,77 tonha), Jepang (1,78
Rendahnya produktivitas kacang tanah di Indonesia antara lain disebabkan
oleh pengynaan benih yang kurang bennutu, pemupukan dan teknik budidaya yang
belum tepat, serta serangan hama dan patogen yang belum dapat diatasi dengan efektif.
Epidemi penyakit virus dapat merupakan kendala biologi utama budidaya h a g tanah
&I Indonesia. Tujuh jenis virus yang secara alamiah dapat menginfeksi kacang tanah
adalah p e a t mottle virus (PeMoV), pemnrf stripe virus (PStV), p e a t mosaic virus
(PMV),peanut crinkle 171rus (PCV), rugrose leaf curl virus, cowpea mild mottle virus
(CMMV), dan tomato sported wilt virus (TSWV) (Roechan et al., 1978; L k k a ,
1986).
Di Indonesia, PStV paling dorninan menyerang kacang tanah dibandingkan
dengan virus-virus yang lain. PStV diietahui terdapat hampir di seluruh pertanaman
kacang tanah dan merupakan salah satu penyebab rendahnya daya hasi kacang tanah
(Saleh ei al., 1989). Salah satu faktor penyebab PStV menjadi e n d e d di lokasi-
lokasi budidaya kacang tanah di Indonesia adalah kemampuan virus ini untuk menular
melalui benih (seedborne). Penularan melalui benih dapat tejadi sejalan dengan
penyebaran benih dari satu daerah ke daerah lain. Selain i t y PStV dapat bertahan dan
mas& infektii selama penyimpanan benih. Sampai saat ini belum dikembangkan
metode yang efektif untuk deteksi PStV dalarn benih kacang tanah terinfeksi karena
secara visual antara benih yang sehat d m terinfeksi PStV tidak dapat dibedakan.
Deteksi molekuler yang cepat, tepat, dan spesifik terhadap PStV sangat diperlukan
metode deteksi molekuler PStV juga diperlukan untuk melacak keberadaan transgen
dalam tanaman transgenik yang dihasilkan.
Berbeda dengan patogen dari golongan bakteri dan jamur yang dapat diatasi
dengan aplikasi pestisida, sampai saat ini belum ditemukan pestisida yang dapat
menekan laju replikasi virus. Beberapa tindakan yang direkomendasikan untuk
pengendalian virus tumbuhan adalah (1) penanaman benih bebas virus, (2) pergiliran
tanaman dan eradikasi tanaman sumber penularan, (3) sanitasi y l m a inang virus, dan
(4) aplikasi insektisida untuk mengendalikan vektor (Direktorat Perlindungan Tanaman
Pangan, 1989). Namun demikian hasil penelitian Saleh et al., (1 99 1 ) menunjukkan
aplikasi insektisida, penyiangan g u h a dan eradikasi tanaman sakit kurang efektif
untuk mengatasi PStV. Sebaliknya, pen- benih kacang tanah bebas PStV
dapat menurunkan laju epidemi penyakit belang di lapang.
Untuk mengatasi epidemi penyakit belang secara komprehensif perlu
dikembangkan metode pengendalian hama terpadu (PHT) pa& budidaya kacang
tanah. Beberapa komponen pengendalian dapat dikembangkan untuk mencegah
terjadinya epidemi penyakit tersebut adalah ( 1 ) pengynaan be& bebas PStV, (2)
monitoring insiden penyakit secara dini (emly warning system), ( 3 ) pengynaan varietas resisten, dan (4) pengendalian vektor.
Tanaman yang resisten terhadap suatu strain patogen dapat menjadi rentan
terhadap strain yang lain. Oleh sebab itu pengetahuan tentang variasi biologi PStV
yang ada di bdagai daerah diIndonesia sangat diperlukan untuk mendukung program
Pengynaan teknologi DNA rekombinan atau rekayasa genetika untuk
perbaikan ketahanan kacang tanah terhadap PStV perlu dikaji karena sampai saat mi
belum ditemukan kultivar kacang tanah komersial yang tahan Rekayasa genetika
memerlukan pengetahuan dasar tentang gen CP-PStV (gen pen~andi coat protezn
PStV) Gen CP-PStV dapat dikonstruksi sebagai gen anti PStV dan diintroduksikan ke dalam genom kacang tanah Ketahanan tanaman transgenik dipergamhi oleh tingkat
kesamam urutan nukleotida gen tersebut dengan strain virus yang menyerang Oleh
sebab itu, pengklonan dan analisis umtan nukleotida gen CP isolat-isolat PStV dari
berbagai daerah di Indonesia perlu dilakukan untuk menentukan klon yang terbaik
untuk gen anti PStV di Indonesia.
Dari sudut pengembangan ilmu, studi biologi moIekuler PStV diperlukan
sebagai model dalam pengembangan bioteknologi untuk tujuan yang lebih luas
Beberapa aspek lain dari studi biologi ~ 0 l e k ~ l e r virus tumbuhan adalah penggunaan
virus tumbuhan mtuk memproduksi vaksii untuk marmsia dan hewan (Featherstone,
1996). Yusibov el al.. (1 9973 fnkmhzat vaksin melalui fisi protein dengan menyisipkan
gen yang mengbihpresikan antigen rabies dan m - 1 pada gen p&yandi C P - A H V
(alfalfa mode wrus].
Teknik biologi molekuier telah menyediakan cara baru untuk klasifikasi,
identifikasi, clan studi kekerabatan antar virus
&uhan
Karakterisasi molekuler genCP dan 3'- (untrmlaied region) banyak digunakan untuk mengetahui hubungan
antara speues-spesies potyvirus (Atreya, 1992) Taksonomi Potywrus, sebagm
besamya variasi diantara anggota kelornpok potyvhs (Ward & Shukla, 1990). Shukla
dan Ward (1988) m ~ b a n d i n & a n susunan asam amino CP 17 strain dari 8 spesies
potyvirus. Hasid studi tersebut menunjukkan kesamaan asarn amino 38-71% untuk
spesies yang berbeda; 90-99% untuk strain-strain dalam spesies virus yang sama; dan
73-83% untuk virus-virus yang berbeda tetapi masih dalarn satu subkelompok. Siudi
variasi molekuler isolat-isolat PStV dalarn penelitian ini bertujuan untuk rnengetahui
keragaman dan evolusi isolat-isolat PStV yang berasal dari berbagai daerah di
Indonesia dengan penekanan subgmp
B C M V
(bean common mosaic v i m ) .Tujuan
Penelitian
Secara urnurn tujuan penelitian ini adalah (1) mengisolasi PStV dari 12 propinsi
di Indonesia, (2) m e n g w v m biologi PStV berdasarkan simptomatoIogi d m
patogenisitas isolat-isoht PStV pack beberapa Mtivar kacang tanah, (3) rnengem-
banp;kan metode deteksi moIekuler PStV dengan teknik RT-PCR dan DNA hibridisasi
dot blot, (4) mengklon
d m
rnendeterrninasi urutan nukleotida isolat-isolat PStV dariberbagai daerah di Indonesia, (5) menganalisis keragaman dan mogenetika PStV
berdasarkan urutan nukleotida gen CP, 3'UTR, dan urutan asam amino CP-PStV.
Tahap-tahap perwbaan yang dilakukan untuk mencapai sasaran penelitian ini
Koleksi Contoh daun kacang tanah terinfeksi v i m dari
PLPq : Lsolasi PStV dari
virus-virus lain
I
PtSk Metode PLBS: d o n j n g ,
molekuler d
-an-
1. Variasi bidogi berdasarkan simptomatologi penyakit
2. Variasi biologi berdasarkan patogenisitas isobt-
PStV pada bebrapa kultivar kacang tanah 3. Metode d-i mdekuler PStV dengan RT-PCR
dan hibridiii dot Mot
4. Variasi biologi Masarkan unrtan nukleoiida gen CP, dan asam amino CP, serta analisis keragaman dan
filogenetika PStV dalam subkdompok BCMV
Garnbar 1. Skema tahapan penelitian yang dilakukan.
TINJAUAN PUSTAKA
Kacang Tanah
Kacang tanah (Arachis hypogaea L.) adalah tanarnan polong-polongan yang
termasuk subfamif Legurninoceae, famili Papilionaceae. Morfologi kacang tanah yang
dibudidayakan bervariasi untuk sifat-sifat ~ertumbuhan dan sifat-sifat khas lainnya,
tetapi semuanya mempunyai karakter pemasakan buah yang tejadi di bawah
permukaan tanah (geokarpi). Pola percabangan di atas tanah pada dasarnya terdiri atas
satu batang pokok (monopodial) (Ashley, 1984)
Arachjs spp. berasal dari daerah tropika dan subtropik Amerika Selatan dari
Argentina sampai Amazon. A. hypogaea yang dibudidayakan dan A. monicola yang
turnbuh liar keduanya adalah dotetraploid (2n=4x-40) sehingga persilangan kedua
spesies hi menghasillcan turunan yang fertil. Diperkirakan perubahan A. menticola
(nenek moyang kacang tanah bar) menjadi A. W g m a terjadi melalui proses seleksi
yang sengaja d i h h k a n menjelang tahun 1500 M sudah tersebar di Arnerika Selatan,
Meksiio, dan Karibia Kacang tanah dibawa oleh bangsa Portugis ke Aiiika dan India
pa& abad ke-16 dan pada waktu yang sama bangsa Spanyol mengintroduksii
kacang tanah ke negara-negara Pasifik Selatan seperti: RRC, Indonesia, dan
Mdagaskar (Ashley, 1984; Van der Maesen & Sornaatmadja, 1992).
Penyebaran kacang tanah sekarang meliputi daerah-daerah tropika dan
subtropika yang terletak diantara 40" LU sampai 40" LS (Ashley, 1984). Budidaya
kacang tanah dilakukan pada tanah ringan, netral atau basa, dengan curah hujan atau
curah hujan di bawah 600 mrn lebih cocok untuk kultivar-kultivar kacang tanah yang
berumur pendek. Sedangkan di daerah-daerah yang agak kering sampai agak basah
dengan curah hujan 600-900 mrn lebih sesuai untuk kultivar-kultivar yang berumur
panjang. Kacang tanah biasa ditanam pada ketinggian 1500 m di atas permukaan laut
dengan kisaran suhu 25-35" C (Ashley, 1984).
Patogenisitas PStV
PStV addah salah satu spesies genus Potyvirus, dan fam* Potyviridae (Francki
et al.. 1991). Potyvirus paling banyak menimbulkan kerugian hasil pertanian
dibandiigkan dengan virus-virus dari genus yang lainnya. Hal ini disebabkan oleh
jumlah spesies Potyvirus yang banyak, penyebaran yang mudah melalui kutu daun
secara nonpersisten yang sulit dikendalikan, infeksinya pada tanaman h a n g
menimbulkan gejala nekrosis, klorosis, dan kerdil (Lmdbo er al., 1992; Tomaru,
1994).
..,
V i s addah parasit pada tingkat molekuler karena secara alarniah virus tidak
mempunyai perangkat sistem metabolisme untuk memperoleh energi. Oleh sebab itu,
virus memerlukan tanaman inang sebagai sumber energi untuk perkembangannya.
Dengan bantuan sel tanaman inang, vims mensintesis protein stmkturd d m fhngsional
yang diperlukan untuk r e p W s i dan transmisi virus dalam jaringan tanaman
(Matthews, 1992). Infeks'~ PStV menginduksi terbentuknya badan inklusi berbentuk
cakra @imvheel) pada jaringan daun tanaman kacang tanah (Gambar 2B). Hajimorad
et al. , (1 996) melaporkan protein Nla (~nrclear incltrsion b e ) dan Nlb terdapat dalam
Sampai saat ini behm ditemukan varietas kacang tanah yang resisten terhadap
PStV. Hasil seleksi 1 1.000 galur kacang tanah koleksi ICRISAT (International
Crops Reseach Institute for Semi-Arid Tropics) yang dilakukan di Indonesia menunjukkan semua galur tersebut rentan terhadap PStV (Saleh & Baliadi, 1988).
Pengujian yang dilakukan diiuar negeri, seperti di USA 244 galur, RRC 663 galur, dan
India 59 galur, juga tidak diternukan satupun galur kacang tanah yang resisten
terhadap PStV @emski & Reddy, 1988; Ebo et al., 1989; Xu, 1988).
Epidemi penyakit belang dimulai dari benih yang terinfeksi PStV sebagai
sumber inokulum pada awaI pertumbuhan kacang tanah, kemudian disebarkan melalui
serangga vektor secara nonpersisten ke tanaman-tanaman sekitarnya. Beberapa spesies
kutu daun yang &pat menularkan PStV adalah Aphis craccivora, A. glycines, A. ~im~cola, Myzus percisae, dan Schzzaphzs rohrndwentris (Natural et al., 1989). Di
Indonesia PStV ditularkan oleh A. craccivora, A. glycznes, A. citricola, RhopaIosiphwn nqidis.
R
pedi, Hysteroneura senfaria. A. gosspii, A. pomi, S.r o t u n d i ~ e ~ s , dan M. persicae (Saleh & Horn, 1989; Suprapto, 1991).
S e h kacang tanah, PStV juga mensinfeksi secara sistematik Glycine mm,
Nicoriana benthami-, Tnyoiium incantaturn, T visiculosum. T subterraneum. Vigna unguzculata, V. r d a t a , Desmonium tr$!orium, Inakgoofera amoena, Peuralza phaseoides. StyIosenthes capitata, S. acraba (Demski et al.. 1984; Natural et al.,
1989; Wonhkaew & DoUet, 1990). Di Indonesia PStV secara alami menyerang
Wongkaew dan Dollet ( 1 990) mengelompokkan 24 isolat PStV, dari berbagai
negara, menjadi delapan kelompok, yaitu: mild moftle, blotch, stripe, blotch-stripe.
blotch-CP-N, chlorotic ring-mottle, chIorotic rzng-paffem, dan neffoticQ"e.
Pengelompokkan tersebut berdasarkan gejala yang timbul pa& &un kacang tanah dan
tanaman indikator lainnya.
Biologi Molekuler PStV
PStV berbentuk batang lentur (.~?'exiars rod} dan bemkuran 12x752 n m
(Gambar 2A). Virion terdiri atas satu utas RNA (ssRNA) dengan bobot molekul (BM)
3100 kDa dan protein selubung yang terdiri atas subunit-subunit protein dengan BM
3 1 kDa (Demski e f aL, 1984). PStV secara in dtro menjadi inaktif pada suhu 65"
selama 10 menit, tit& batas pengenceran dalarn cairan perasan daun kacang tanah
selama 3 hari @PI, 1990).
Genom PStV berukuran 10059 nt tidak t e m u k ekor polidenilat @oli-A).
Sebagai mana genom Potyvirus lainoya, pada ujung b e a n 5' genom RNA terdapat
protein (VPg) yang terikat secara kovalen pada RNA dan bagian ujung 3' terdapat
poli-A (Gunashingheet et al.. 1994; Shaw et al.. 1990). Genom RNA tersebut terdiri
atas satu kerangka baca (open reacfing3gframe) yang meliputi 95% total RNA. Kodon
awal terletak 134-136 d m kodon stop 9768-9770 nukleotida dari bagian ujung bagan
Gambar 2. (A) Partikel PStV berbetuk batang lentur (fi'exious rad) dan (B) badan
Ekspresi genom RNA-PStV akan membentuk satu molekul poliprotein yang
selanjutnya diproses oleh ensim protease menjadi molekui-molekul protein yang
diperlukan untuk replikasi dan patogenesis tumbuhan (Gambar 3). Beberapa jenis
protein yang diekspresikan oleh genom PStV dan fingsi biologinya dalam patogenesis
tanarnan dapat dilihat pada Tabel I .
5' Genom ssRNA PStV (70059
nO
3'P I H C A o P3 6K CI
e
I
pdipoteinI
Eel
1 7 1
*,
p,
,
T
i
[image:192.537.41.462.21.567.2]I-.[
E l
lnrl
1
7
Gambar 3 . Peta genom PStV dan protein-protein yang disintesis dari genom RNA PStV (Gunasinghe et al., 1994).
Secara umum proses replikasi virus yang genomnya RNA berutas tunggal
(+ssFtNA) terjadi melalui beberapa tahap, yaitu: (1) virion mas& ke dalam sitoplasma
sel tanaman inang, (2) pelepasan CP dari RNA virus, (3) RNA berasosiasi dengan
ribosom tanarnan inang dan sintesis polimerase untuk replikasi RNA, (4) sintesis utas
negatif RNq (5) sintesis utas positif RNA d m mRNA CP menggunakan utas negatif
virion yang te jadi melalui penggabungan antara utas positif RNA dengan CP, dan (8)
penyebaran virion ke sel-sel sekelilingnya melalui plasmodesmata (%=hews, 1992).
Tabel 1. Protein-protein yang disintesis oleh genorn PStV dan potyvirus lainnya dalam sel tanaman inang (Gunashinghe et al., 1994; Shaw et al., 1990).
Coat protein {CP) mtein pembuqkus, trannnisi melalui
Variasi Moleknler Potyvirus
Gen CP dan 3'UTR banyak digunakan sebagai dasar untuk ident.5ka-4, studi
kerag- dan analisis filogenetika potyvirus. Bagian tengah urutan asam amino CP
potyvirus hampir sama untuk virus-virus dalam kelompok potyvirus; sebaliknya pada
ujung amino (N-) dan karboksil (C-) bervariasi dari satu kelompok virus ke kelompok
yang lain (Liidbo ef al., 1992).
Shukla dan Ward (1988) men- urutan asam amino CP unruk
mengetahui hubungan kekerabatan 17 strain dari delapan spesies potyvirus. Hasil
kajian tersebut menunjukkan spesies-spesies yang berbeda mempunyai kesamaan 38-
71% sebaliknya untuk strain-strain dalam spesies virus yang sama homologinya 90-
9%. Frenkel et al.. (1 989) melaporkan hasil studi homologi 3'UTR antar strain dari
[image:193.534.45.461.20.576.2]tersebut menunjukkan homologi urutan nukleotida 39-53% untuk spesies virus yang
berbeda, dan 83-99% untuk strain dalam spesies virus yang sama.
Hasil penelitian Teycheney dan Dietzgen (1994) menunjukkan antara isolat
PStV-Ib (berasal dari Indonesia) dengan isolat PStV-USa dan PStV-USb @eras& dari USA) mempunyai kesamaan 98% pada urutan nukleotida 3'UTR. Kesamaan umtan
asam amino CP dan nukleotida 3'UTR antara PStV dengan PeMoV @ e m t mottle
virus) adalah 66,7% dan 34,6%. Berdasarkan kriteria penggolongan yang diajukan
oleh Shukla dan Ward (1988) dan Frankel et al.. (1989) kedua virus diklasifikasikan
ke dalam spesies virus yang berbeda. Kesamaan antara PStV dengan B C W - N U
pada urutan nukleotida 3'UTR adalah 96,6% sehingga kedua virus tersebut
diklas-kan sebagai strain dalam spesies virus yang sarna (Collmer e f al., 1996).
Berdasarkan reaksi serologi BCMV dibagi menjadi dua subkelompok yaitu
serotipe A clan B (Vetten et d., 1992). Mink e f ad., ( 1 994) menysulkan pemisahkan
subkelompok B C W serotipe A menjadi B C W dm serotipe B menjadi BCMNV.
Studi molekuler virus-virus yang temasuk ke dalam subkelompok BCMV
menggunakan urutan nukleutida 3 ' m genom RNA membuktikan BCMV
mempakan virus yang berbeda dengan BCMNV ( b e m common mosaic necrosis
virus) (Berger et aL, 1997). Analisis filogenetika menempatkan PStV ke dalam
subkelompok BCMV (Murphy et al.. 1995; Berger et al.. 1997). Analisis urutan asarn
amino CP menunjukkan PStV, B I C W (hlackeye cowpea mosaic virus), dan A M
ISOLASI PStV DARI DAUN
KACANG TANAH YANG
PENDAHULUAN
PStV diketahui telah tersebar di Asia Tenggara, Cina, India, dan USA (Demski
et al., 1984; Reddy, 1985; Natural el al., 1989). Di Indonesia PStV merupakan virus yang paling dominan yang menyerang kacang tanah (Saleh el al., 1989). Salah satu
penyebab PStV menjadi endernik di lokasi-lokasi penanaman kacang tanah di
Indonesia adalah sifat penularannya yang dapat tejadi melalui benih sehingga
memungkinkan PStV tersebar dari satu daerah ke daerah yang lain sejalan dengan
penyebaran b e d .
Penelitian ini bertujuan untuk mengisolasi PStV dari kacang tanah yang berasal
dari 12 propinsi di Indonesia. Isolasi dilakukan dengan tanaman indikator spesifik
untuk PStV. Isolat-isolat yang didapat akan diynakan untuk percobaan selanjutnya.
BAHAN
DAN
METODE
Tempat
dan
Waktu Penelitian
Penelitian ini ddakukan di Laboratorium Biologi MoleMer, Jumsan Budidaya
Pertanian, Fakultas Pertanian, IPB. Percobaan dilakukan dari bulan Oktober 1995
sampai Pebruari 1996.
Contoh Tanaman Sakit dari Lapang
Contoh daun kacang tanah sakit yang diduga terinfeksi PStV dikurnpulkan
kuning (stripe). Contoh daun dipotong-potong dan dikering anginkan selama 3 hari
dan disimpan dalamfreezer.
Contoh daun tersebut digerus dalam bufer inokulasi (0,OlM kalium fosfat, pH
7,O; 0,OlM Na-DIECA), kemudian disaring dengan dua lapis kain kasa. Cairan
perasan yang didapat digunakan sebagai inokulum untuk uji inokulasi mekanik pada
tanaman indikator (Chenopodium mnmanficolor Coste & Reyn).
Isolasi PStV dari Virus Lain
Isolasi virus bertujuan untuk memisahkan PStV dari virus-virus lain yang
menyerang kacang tanah di lapang. Isolsi virus tersebut ddakukan menggunakan
tanaman indikator yang menunjukkan g e j a spesifik untuk infeksi PStV. Seberapa
virus yang dapat menimbufkan gejala belang pada kacang tanah di lapang antara lain
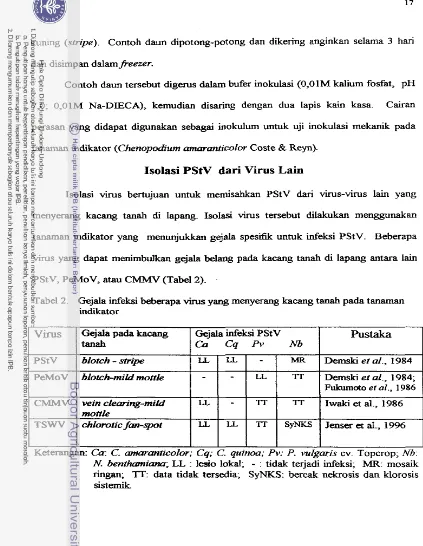
PStV, PeMoV, atau CMh4V (Tabel 2).
Tabel 2. Gejala infeksi beberapa virus yang menyerang kacang tanah pada tanaman indikator
Virus
PStV
I I I I I I I
Keterangan: Ca: C. amaranticolor; Cq;
C.
guinoa;PV:
P.
~wlgaris cv. Topcrop;Nb:
N.
benfhmniaraa; LL : lesio lokal;-
: tidak terjadi infeksi; MR: mosaikringan, TT: data tidak tersedia; SyNKS: bercak nekrosis dan klorosis sisternik
PeMoV
CMMY
TSWV
Pustaka
Dernski e f al., 1984
Gejala pada kacang
tanah
biotch-mild mottle
w i n clearing-mild motile
chlorotic fan-qmt
Gejala infeksi PStV
Ca
Cq h.Nb
blotch-
stripe1
tLI
LL-
1
M R-
LL-
-
LL LL 1TS Y ~
Demski et al., 1984; Fukumoto ef al., 1986
Iwaki et al., 1986
[image:197.537.42.465.17.563.2]1 . Isolasi
PStV
d a r i P e M o VPemisahan antara PStV dengan PeMoV dilakukan dengan menginokulasikan
cairan perasan daun kacang tanah yang menunjukkan gejala belang pada C.
amarmticolor. Inokulasi PStV pada daun C. maranticolor akan menimbuIkan gejala lesio lokal. Untuk memperoleh isolat virus yang murni dilakukan isolasi bercak tunggal
pada daun C. amarmticolor. h s i o lokal yang timbul pada daun C. mnaranticoIor
diisolasi dengan dua kali isolasi lesio lokal tunggal. Satu lesio lokal yang timbul pada
daun C. maranticolor diisolasi dan d i p a k a n sebagai inomurn untuk inokulasi
d a m C. amaranticolor yang kedua. Daun C. amarmticolor yang diinokulasi dengan inokulum yang berasal dari lesio lokal kedua digunakan untuk identifikasi PStV
menggunakan tanaman indikator.
2. Isofasi PStV d a r i CMMV
Pemisahan antara PStV dengan C M M V diiakukan pada tanaman indikator C.
quinoa yang hanya dapat diinfeksi oleh PStV. Inokulasi PStV pada C. guinoa akan
menimbulkan gejala lesio l o w , sebabknya C. quirwa tidak dapat dinfiehi oleh
Gambar 4 Peta asal isolat-isolat PStV dari 12 propinsi di Indonesia
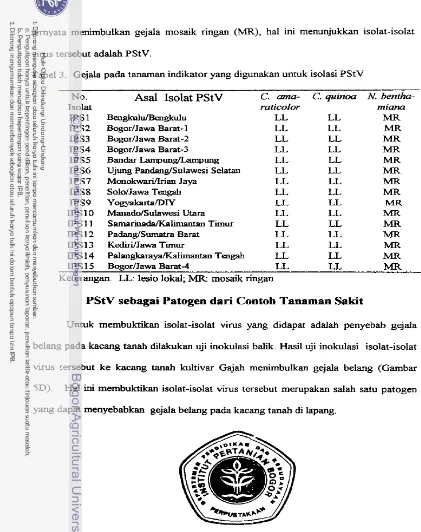
Asal Isolat PStV: P S I - Bengkulu, IPS2 Jawa Barat-1, P S 3 . Jawa
Barat-2, IPS4 Jawa Barat-3, IPSS. Lampung, IPS6 Sdawesi Selatan,
IPS7. Irian Jaya; IPS8. Jawa tengah; P S 9 . Yogyakarta, IPS10 Sulawesi
Utara, IPS1 1 Kalimantan Timur, IPS12 Sumatra Barat, IPS 13 Jawa
[image:199.534.42.461.24.553.2]3. Isolasi PStV dari TSWV
T S W d m PStV tidak dapat di