KAJIAN DINAMIKA PERTUMBUHAN FITOPLANKTON
DAN KETERKAITANNYA DENGAN VARIABILITAS
INTENSITAS CAHAYA MATAHARI DAN
NUTRIEN INORGANIK TERLARUT
DI PERAIRAN TELUK BANTEN
ALIANTO
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Kajian Dinamika Pertumbuhan Fitoplankton dan Keterkaitannya Dengan Variabilitas Intensitas Cahaya Matahari dan Nutrien Inorganik Terlarut di Perairan Teluk Banten adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, September 2011
ABSTRACT
ALIANTO. Study of the Dynamic of Phytoplankton Growth and its Relation to Variability of Solar Radiation Intensity and Dissolved Inorganic Nutrient in Banten Bay. Under direction of ENAN M. ADIWILAGA, ARIO DAMAR, and ENANG HARRIS
The growth of phytoplankton in the waters mostly depends on sunlight intensity and dissolved inorganic nutrient present in euphotic zone. This research aimed to count phytoplankton growth including growth rate, doubling time, and generation time, and to analyze growth rate response of phytoplankton to solar radiation intensity and dissolved inorganic nutrient. Measurement of phytoplankton growth was conducted from April to September 2008 at five stations. Station 1 was situated at the mouth river, station 2 at the middle of the Bay toward the beach, station 3 at the middle of the bay, station 4 located at the middle toward outside the bay, and station 5 located outside the bay. Phytoplankton growth at each station was measured at various water depth. At station 1, measurement was done at three euphotic depth namely at 10%, 5%, and 1% from surface irradiance. At station 2, 3, and 4, growth measurement was conducted at 50%, 25%, 10%, and 5% from surface irradiance while at station 5, phytoplankton growth was measured at five euphotic depth namely 50%, 25%, 10%, 5%, and 1% from surface irradiance. Research result showed that phytoplankton growth occurred faster at intermediate light intensity and intermediate concentration of dissolved inorganic nutrient. However, the role of light intensity and dissolved inorganic nutrient in controlling phytoplankton growth was not synergic. Temporally, light intensity did not really control the growth of phytoplankton as compared to dissolved inorganic nutrient in term of DSi because sunlight intensity from April to September was relatively stable compared to DSi that tended to be various. Spatially, on the other hand, light intensity and dissolved inorganic nutrient in term of DIN, DIP, and DSi simultaneously controlled phytoplankton growth. This occurred due to light intensity was varied and tended to be homogenous from station 1 to station 4 and low at station 5. Different phenomena also occurred with DIN, DIP, and DSI which was varied and tended to be higher at station 1 and homogenous at station 1 to station 4. However, DSi tended to be more varied than DIN and DIP. Based on this finding, it was stated that DSi was more dominant in controlling phytoplankton growth in Banten Bay and categorized as fast growth rate ranging from 0.66-2.44 day-1, doubling time 0.34-2.04.day-1, and generation time ranging from 0.91-3.53 day-1.
ALIANTO. Kajian Dinamika Pertumbuhan Fitoplankton dan Keterkaitannya Dengan Variabilitas Intensitas Cahaya Matahari dan Nutrien Inorganik Terlarut di Perairan Teluk Banten. Dibimbing oleh ENAN M. ADIWILAGA, ARIO DAMAR, dan ENANG HARRIS
Pertumbuhan fitoplankton di perairan sangat tergantung pada ketersediaan intensitas cahaya matahari dan nutrien inorganik terlarut yang berada di zona eufotik. Permasalahannya adalah ketersediaan intensitas cahaya matahari dan nutrien inorganik terlarut ini tidak selalu berada pada kondisi yang serasi untuk menunjang pertumbuhan fitoplankton. Hal ini tentu akan menyebabkan pertumbuhan fitoplankton di perairan selalu berada pada kondisi yang tidak optimal. Berdasarkan hal ini maka penelitian ini dilakukan dengan berbagai pendekatan dengan melakukan pengukuran pertumbuhan fitoplankton pada beberapa kedalaman eufotik. Penelitian ini bertujuan untuk menghitung pertumbuhan fitoplankton yang meliputi laju pertumbuhan, waktu doubling, dan waktu generasi fitoplankton serta menganalisis respon laju pertumbuhan fitoplankton pada perubahan intensitas cahaya matahari dan nutrien inorganik terlarut.
Pengukuran pertumbuhan fitoplankton dilakukan dari bulan April sampai September pada lima stasiun. Stasiun satu berada pada muara sungai, stasiun dua berada pada perairan pantai, stasiun tiga berada pada bagian tengah teluk tapi lebih mengarah ke pantai, stasiun empat berada pada bagian tengah teluk tapi lebih mengarah ke luar teluk, dan stasiun lima berada pada luar teluk. Pengukuran pertumbuhan fitoplankton pada setiap stasiun dilakukan pada kedalaman yang berbeda-beda. Pada stasiun satu dilakukan pada tiga kedalaman eufotik yang secara berturut-turut terdiri dari kedalaman eufotik 10%, 5%, 1% dari cahaya permukaan. Pada stasiun dua, tiga, dan empat dilakukan pada empat kedalaman eufotik yang secara berturut-turut terdiri dari kedalaman eufotik 50%, 25%, 10%, dan 5% dari cahaya permukaan. Sedangkan pada stasiun lima dilakukan pada lima kedalaman eufotik yang secara berturut-turut terdiri dari kedalaman eufotik 50%, 25%, 10%, 5%, dan 1% dari cahaya permukaan.
Hasil uji anova menunjukkan bahwa laju pertumbuhan, waktu doubling, dan waktu generasi fitoplankton berbeda di zona eufotik, baik secara temporal maupun spasial. Laju pertumbuhan fitoplankton lebih tinggi pada zona eufotik 50% dan 25% dengan rata-rata sebesar 1.66 hari-1 dan 1.64 hari-1. Sedangkan waktu doubling fitoplankton di kedua zona eufotik ini dengan rata-rata sebesar 0.72 hari dan 0.92 hari serta waktu generasi fitoplankton dengan rata-rata sebesar 1.10 hari-1dan 1.30 hari-1. Hal ini berarti bahwa pada zona eufotik 50% dan 25% secara berturut-turut terjadi peningkatan biomassa fitoplankton sebesar 1.66 mg C hari-1dan 1.64 mg C hari-1dalam waktu rata-rata sebesar 0.72 hari dan 0.92 hari serta jumlah generasi fitoplankton baru yang dihasilkan dengan rata-rata sebesar 1.10 hari-1dan 1.30 hari-1.
rata-rata sebesar 3.53 hari-1terdapat pada bulan Mei. Hal ini berarti bahwa pada bulan Mei terjadi peningkatan biomassa fitoplankton sebesar 2.44 mg C hari-1 dengan waktu rata-rata sebesar 0.34 hari serta jumlah generasi fitoplankton baru yang dihasilkan dengan rata-rata sebesar 3.53 hari-1. Sedangkan secara spasial, laju pertumbuhan fitoplankton tertinggi dengan rata-rata berkisar dari 1.32-1.82 hari-1 dan waktu doubling dengan rata-rata berkisar dari 0.68-1.07 hari, serta waktu generasi fitoplankton dengan rata-rata berkisar dari 1.91-2.63 hari-1 secara berturut-turut terdapat pada stasiun satu, dua, tiga, dan empat. Hal ini berarti bahwa pada stasiun satu, dua, tiga, dan empat secara berturut-turut terjadi peningkatan biomassa fitoplankton dengan rata-rata berkisar dari 1.32-1.82 mg C hari-1 dalam waktu rata-rata berkisar 0.68-1.07 hari serta jumlah generasi fitoplankton baru yang dihasilkan dengan rata-rata berkisar dari 1.91-2.63 hari-1.
Hasil analisis korelasi pearson’s memperlihatkan adanya korelasi positif antara laju pertumbuhan fitoplankton dengan perubahan intensitas cahaya matahari. Hal ini menunjukkan bahwa komunitas fitoplankton dapat beradaptasi dengan berbagai level intensitas cahaya matahari. Hasil uji anova menunjukkan bahwa komunitas fitoplankton lebih respon dengan level intensitas cahaya matahari yang sedang dibandingkan dengan level intensitas cahaya matahari yang rendah maupun tinggi. Hal ini dapat dilihat dari laju pertumbuhan fitoplankton yang tinggi dengan rata-rata sebesar 2.44 hari-1terjadi pada level intensitas cahaya matahari dengan rata-rata berkisar dari 236.15-301.65 µmol foton m-2 s-1. Sedangkan pada intensitas cahaya matahari yang rendah dengan rata-rata sebesar 14.63 µmol foton m-2 s-1 dan tinggi dengan rata-rata sebesar 717.37 µmol foton m-2 s-1 diperoleh laju pertumbuhan fitoplankton secara berturut-turut hanya sebesar 0.75 hari-1dan 1.66 hari-1.
Hal yang sama terjadi pula pada nutrien inorganik terlarut terutama dalam bentuk DSi, dimana hasil analisis korelasi Pearson’s menunjukkan adanya korelasi positif antara laju pertumbuhan fitoplankton dengan perubahan konsentrasi DSi. Hal ini menunjukkan bahwa komunitas fitoplankton dapat merespon perubahan konsentrasi DSi dari konsentrasi yang rendah sampai tinggi atau dengan kata lain komunitas fitoplankton dapat beradaptasi pada semua level konsentrasi DSi. Walaupun demikian, komunitas fitoplankton lebih respon dengan konsentrasi DSi yang relatif sedang dibandingkan dengan konsentrasi DSi yang tinggi maupun rendah. Hal ini dapat dilihat dari uji anova menunjukkan bahwa komunitas fitoplankton dengan laju pertumbuhan tertinggi dengan rata-rata sebesar 2.44 hari -1
terjadi pada konsentrasi DSi yang sedang dengan rata-rata sebesar 82.64 μM. Sedangkan pada konsentrasi DSi yang tinggi dengan rata-rata sebesar 148.39μM, laju pertumbuhan fitoplankton yang diperoleh dengan rata-rata sebesar 1.82 hari-1. Pada konsentrasi DSi yang rendah dengan rata-rata sebesar 23.85 μM, laju pertumbuhan fitoplankton yang diperoleh hanya dengan rata-rata sebesar 0.93
μM.
berturut-turut pada stasiun satu, dua, tiga dan empat terjadi bersamaan dengan konsentrasi DIN dengan rata-rata berkisar dari 5.20-14.61 μM dan DIP dengan rata-rata berkisar dari 0.06-0.20μM.
©
Hak Cipta milik IPB, tahun 2011 Hak Cipta dilindungi Undang-undang1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah. b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
NUTRIEN INORGANIK TERLARUT
DI PERAIRAN TELUK BANTEN
ALIANTO
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Ilmu Perairan
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji Luar Komisi pada Ujian Tertutup :
1. Dr. Ir. Hefni Effendi, M.Phil
(Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor)
2. Dr. Ir. Niken T.M. Pratiwi, M.Si
(Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor)
Penguji Luar Komisi pada Ujian Terbuka :
1. Dr. Ir. Zahidah, M.S.
(Jurusan Perikanan, Fakultas Perikanan dan Ilmu Kelautan, Universitas Padjadjaran
Bandung)
2. Dr. Ir. Yusli Wardiatno, M.Sc
Teluk Banten
Nama : Alianto
NIM : C161060031
Disetujui Komisi Pembimbing
Dr. Ir. Enan M. Adiwilaga Ketua
Dr. Ir. Ario Damar, M.Si Prof. Dr. Ir. Enang Harris, M.S.
Anggota Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana
Ilmu Perairan
Prof. Dr. Ir. Enang Harris, M.S. Dr. Ir. Dahrul Syah, M.Sc.Ag.
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala limpahan rahmat dan karunia-Nya kepada penulis sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan April 2008 ini ialah produktivitas perairan, dengan judul Kajian Dinamika Pertumbuhan Fitoplankton dan Keterkaitannya Dengan Variabilitas Intensitas Cahaya Matahari dan Nutrien Inorganik Terlarut di Perairan Teluk Banten.
Pada kesempatan ini penulis menyampaikan ucapan terima kasih yang sebesar-besarnya kepada komisi pembimbing yang terdiri dari Bapak Dr. Ir. Enan M. Adiwilaga, selaku ketua komisi pembimbing, Bapak Dr. Ir. Ario Damar, M.Si., dan Bapak Prof. Dr. Ir. Enang Harris, M.S., selaku anggota komisi pembimbing yang telah banyak memberi arahan dan bimbingan kepada penulis. Disamping itu, penulis menyampaikan pula ucapan terima kasih yang sebesar-besarnya kepada penguji luar komisi pembimbing yang terdiri dari Bapak Dr. Ir. Hefni Effendi, M.Phil., Ibu Dr. Ir. Niken T.M. Pratiwi, M.Si., Ibu Dr. Ir. Zahidah, M.S., dan Bapak Dr. Ir. Yusli Wardiatno, M.Sc., yang telah berkenan menyumbangkan buah pikirannya untuk memperkaya tulisan ini.
Semoga karya ilmiah ini bermanfaat. Amin.
Bogor, September 2011
Pada kesempatan ini penulis menyampaikan ucapan terima kasih yang sebesar-besarnya secara berturut-turut kepada :
1. Kementerian Pendidikan Nasional, Direktorat Jenderal Pendidikan Tinggi, atas bantuan beasiswa BPPS tahun 2006 serta dukungan yang telah diberikan dengan secara berturut-turut memberikan dana penelitian melalui Program Hibah Bersaing tahun 2008 dan Hibah Doktor tahun 2011.
2. Kementerian Kelautan dan Perikanan, Direktorat Jenderal Kelautan, Pesisir, dan Pulau-Pulau Kecil, atas bantuan biaya penelitian dan penulisan disertasi melalui Program COREMAP tahun 2008.
3. Yayasan Toyota dan Astra, atas bantuan biaya penelitian yang telah diberikan pada tahun 2009.
4. Yayasan Supersemar, atas bantuan biaya penelitian yang telah diberikan pada tahun 2009.
5. Rektor universitas Negeri Papua, atas kesempatan yang telah diberikan kepada penulis untuk melanjutkan pendidikan S3.
6. Bapak Dr. Ir. Kardiyo Praptokardiyo, atas masukan, saran, dan nasehat yang diberikan kepada penulis dari semenjak menempuh studi pada program S2 sampai dengan S3.
7. Ayahanda, H. La Hamuna dan Ibunda Hj. Wa Nuru atas kasih sayang dan doa yang telah diberikan kepada penulis selama menuntut ilmu.
8. Mertua, Drs. Marlis dan Ibu Agustina Hermianti atas kasih sayang dan doa yang telah diberikan kepada penulis selama menuntut ilmu.
9. Istri, Yudistira Anggraini, Amd.Pi dan ananda Rhizki Solenia Alianto (7 tahun) dan Chalisa Alianto (2 tahun), atas kasih sayang, dukungan, dan pengorbanan yang telah diberikan kepada penulis selama menuntut ilmu. 10. Bapak Sawad dan tim Teluk Banten, atas segala pengorbanan saat
membantu penulis selama pengambilan data dan sampel di lapangan. 11. Ibu Ana Mariana dan seluruh staf laboratorium Produktivitas dan
Lingkungan Perairan, atas segala bantuannya kepada penulis saat melakukan analisis laboratorium.
12. Sahabat-sahabat terbaik selama studi dan seangkatan AIR 2006, Dr. Ir. Rahmadi Tambaru, M.Si., Ir. Safrudin La Abukena, M.Si., Ir. Sriati, M.Si., Bahtiar, S.Pi., M.Si., Yuliana, S.Pi., M.Si., Ir. Wellem Muskita, M.Si., Dr. Ir. Hengky Manopo, M.Sc., Dr. Ir. Muhammad Aris, S.Pi., M.Si., Ir. Frederika S. Pello, M.Si, Usman Madubun, S.Pi., M.Si., dan Kadri Laetje, S.Pi, atas bantuan dan kerjasama yang terjalin selama masa studi.
13. Berbagai pihak lainnya yang tidak sempat disebutkan namanya satu persatu yang memiliki andil dalam keberhasilan penulis sampai dengan menyelesaikan studi S3 di Program Studi Ilmu Perairan, Sekolah Pascasarjana, Institut Pertanian Bogor.
RIWAYAT HIDUP
Penulis dilahirkan di Wanci pada tanggal 5 Maret 1970 sebagai anak ketiga dari tujuh bersaudara dari pasangan H. La Hamuna dan Hj. Wa Nuru. Pendidikan sarjana di tempuh di Program Studi Manajemen Sumberdaya Perairan, Fakultas Perikanan, Universitas Pattimura, lulus pada tahun 1997. Pada tahun 2003, penulis diterima di Program Studi Ilmu Perairan pada Program Pascasarjana IPB dan menamatkannya pada tahun 2006. Kesempatan untuk melanjutkan ke program doktor pada program studi dan perguruan tinggi yang sama di peroleh pula pada tahun 2006. Beasiswa pendidikan pascasarjana diperoleh dari Kementerian Pendidikan Nasional.
Penulis bekerja sebagai staf dosen pada Program Studi Manajemen Sumberdaya Perairan, Jurusan Perikanan, Fakultas Peternakan, Perikanan dan Ilmu Kelautan, Universitas Negeri Papua.
Selama mengikuti program S3, penulis telah mempublikasikan beberapa karya ilmiah sebagai berikut :
1. Produktivitas primer fitoplankton dan keterkaitannya dengan unsur hara dan cahaya di perairan Teluk Banten. Disajikan dalam Konferensi Sains Kelautan dan Perikanan I Tahun 2007 di Bogor serta dipublikasi tahun 2008 pada Jurnal Ilmu-Ilmu Perairan dan Perikanan Indonesia15(1): 21-26.
2. Variasi spasial konsentrasi nutrien terlarut di zona eufotik perairan Teluk Banten. Disajikan dalam Seminar Nasional Kimia XVIII Tahun 2008. Yogyakarta.
3. Pengaruh air buangan pada produktivitas primer dan pertumbuhan terumbu karang. Disajikan dalam Simposium Terumbu Karang Nasional ke-2 Tahun 2008 di Jakarta serta dipublikasi dalam Prosiding Simposium Terumbu Karang Nasional ke-2.
4. Estimasi potensi ikan pelagis kecil berbasis produktivitas primer fitoplankton di perairan laut. Disajikan dalam Seminar Nasional Tahunan VI Hasil Penelitian Perikanan dan Kelautan Tahun 2009 di Yogyakarta serta dipublikasi dalam Prosiding Seminar Nasional Tahunan VI Hasil Penelitian Perikanan dan Kelautan..
5. Measurement of dissolved inorganic nutrient in euphotic zone the Banten Bay. Dipublikasi tahun 2009 padaIndonesian Journal of Chemistry9(2): 217-225. 6. Kajian tentang pengaruh perbedaan konsentrasi N, P dan Si pada akumulasi
biomassa fitoplankton di perairan Teluk Banten. Disajikan dalam Seminar Nasional Tahunan VIII Hasil Penelitian Perikanan dan Kelautan Tahun 2011. Yogyakarta.
Halaman
DAFTAR TABEL... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN... xvii
PENDAHULUAN. ... 1
Latar Belakang. ... 1
Rumusan Masalah. ... 2
Tujuan dan Manfaat . ... 5
Kebaruan . ... 5
Hipotesis. ... 6
TINJAUAN PUSTAKA. ... 8
Karakteristik Teluk Banten. ... 8
Konsep Pertumbuhan Fitoplankton... 10
Pembatas Pertumbuhan Fitoplankton. ... 15
METODE PENELITIAN... 25
Lokasi dan Waktu Penelitian. ... 25
Menentukan Stasiun dan Titik Pengambilan Contoh... 25
Pengukuran Parameter Kualitas Air... 27
Analisis Data. ... 34
HASIL DAN PEMBAHASAN... 35
Hasil. ... 35
KedalamanSecchi discdan Koefisien Peredupan. ... 35
Zona Mixing dan Eufotik... 37
Salinitas... 42
Suhu. ... 44
pH... 47
Intensitas Cahaya Matahari... 49
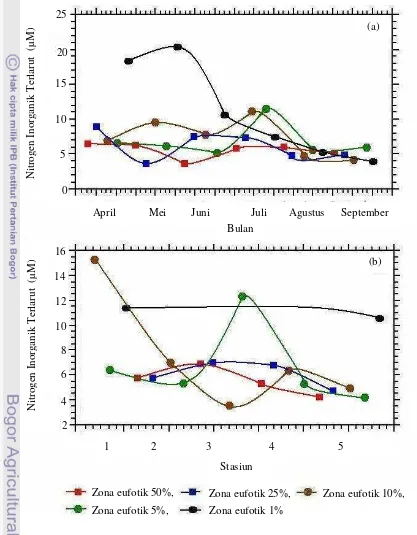
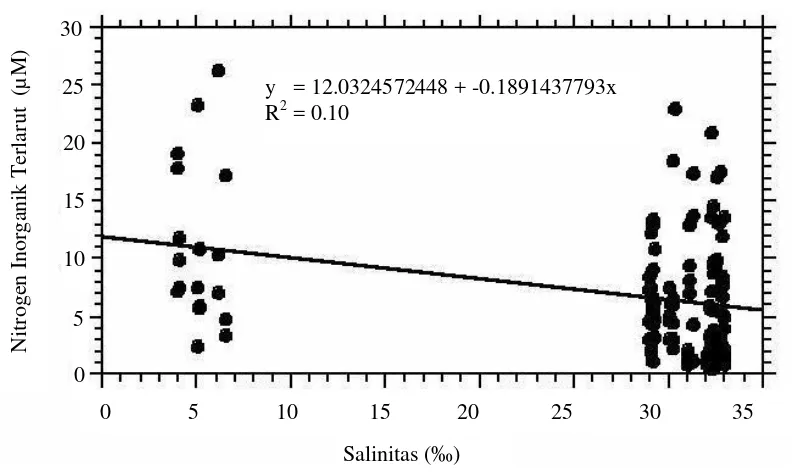
Nutrien Inorganik Terlarut. ... 53
Produktivitas Primer Fitoplankton. ... 74
Klorofil-a... 76
Kelimpahan dan Komposisi Fitoplankton. ... 78
Pertumbuhan Fitoplankton... 89
Pembahasan... 95
Status Pengukuran Pertumbuhan Fitoplankton . ... 95
Pertumbuhan Fitoplankton dan Perbandingannya Dengan Perairan Temperate dan Tropis Lainnya ... 101
Pembatas Pertumbuhan Fitoplankton ... 105
Respon Fitoplankton Pada Perubahan Intensitas Cahaya Matahari . ... 110
Respon Laju Fitoplankton Pada Perubahan Konsentrasi Nutrien Inorganik Terlarut. ... 121
Laju Pertumbuhan dan Biomassa Fitoplankton . ... 127
SIMPULAN DAN SARAN. ... 131
DAFTAR PUSTAKA. ... 132
Halaman
1 Posisi stasiun untuk pengukuran dan pengambilan sampel. ... 26
2 Parameter kualitas air laut yang akan diukur serta metode dan alat ukur yang digunakan... 29
3 Nilai salinitas (‰) dan suhu (°C) permukaan dan dasar Teluk Banten. ... 38
4 Komposisi fitoplankton di zona eufotik Teluk Banten. ... 81
5 Komposisi kelimpahan genera Bacillariophyceae. ... 85
6 Komposisi kelimpahan genera Dinophyceae... 87
7 Status pengukuran pertumbuhan fitoplankton di perairan Teluk Banten... 96
8 Variasi temporal pertumbuhan fitoplankton di Teluk Banten. ... 101
9 Variasi spasial pertumbuhan fitoplankton di Teluk Banten... 102
10 Kisaran rata-rata laju pertumbuhan fitoplankton pada beberapa lokasi di perairan temperate dan tropis... 104
11 Variasi temporal pertumbuhan fitoplankton sebagai fungsi dari intensitas cahaya matahari dan nutrien inorganik terlarut. ... 108
12 Variasi spasial pertumbuhan fitoplankton sebagai fungsi dari intensitas cahaya matahari dan nutrien inorganik terlarut. ... 108
13 Laju pertumbuhan fitoplankton pada enam level intensitas cahaya Matahari di zona eufotik Teluk Banten... 116
DAFTAR GAMBAR
Halaman
1 Diagram alir kerangka teoritis pemecahan masalah. ... 7
2 Arah arus pada musim barat (a) dan timur (b) di Teluk Banten ... 9
3 Berkurangnya ukuran sel setelah terjadinya pembelahan sel... 12
4 Pembelahan sel dan pembentukan frustula. ... 12
5 Kurva pertumbuhan fitoplankton. ... 14
6 Distribusi intensitas cahaya PAR di perairan laut... 17
7 Penetrasi intensitas cahaya matahari... 18
8 Rasio ketersediaan intensitas cahaya matahari dan nutrien inorganik terlarut di zona eufotik ... 19
9 Lokasi penelitian di perairan Teluk Banten. ... 25
10 Desain zonasi secara horisontal dan vertikal lokasi penelitian dilihat dari samping... 27
11 Profil kedalaman Secchi discdan koefisien peredupan Teluk Banten. ... 36
12 Profil kedalaman zona mixing dan eufotik Teluk Banten... 39
13 Profil rasio zona mixing dan eufotik Teluk Banten. ... 40
14 Profil salinitas di zona eufotik secara temporal (a) dan spasial (b) di Teluk Banten... 43
15 Korelasi salinitas dengan kedalaman Secchi disc. ... 44
16 Profil suhu di zona eufotik secara temporal (a) dan spasial (b) di Teluk Banten... 46
17 Profil pH di zona eufotik secara temporal (a) dan spasial (b) di Teluk Banten... 48
18 Korelasi pH dengan salinitas. ... 49
19 Profil intensitas cahaya matahari permukaan di Teluk Banten... 50
20 Profil intensitas cahaya matahari di zona eufotik secara temporal (a) dan spasial (b) di Teluk Banten... 52
21 Profil intensitas cahaya matahari sebagai fungsi salinitas ... 53
22 Profil nitrogen inorganik terlarut di zona eufotik secara temporal (a) dan spasial (b) di Teluk Banten... 55
24 Profil amonia nitrogen di zona eufotik secara temporal (a) dan
spasial (b) di Teluk Banten. ... 57 25 Profil amonia nitrogen sebagai fungsi salinitas. ... 58 26 Profil nitrat nitrogen di zona eufotik secara temporal (a) dan spasial (a)
di Teluk Banten... 60 27 Profil korelasi nitrat nitrogen sebagai fungsi zona mixing. ... 61 28 Profil distribusi nitrat nitrogen sebagai fungsi salinitas... 61 29 Profil nitrit nitrogen di zona eufotik secara temporal (a) dan spasial (a)
di Teluk Banten... 62 30 Profil distribusi nitrit nitrogen sebagai fungsi salinitas. ... 62 31 Profil fosfat inorganik terlarut di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 65 32 Profil fosfat inorganik terlarut sebagai fungsi salinitas. ... 66 33 Profil silika inorganik terlarut di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 67 34 Profil silika inorganik terlarut sebagai fungsi salinitas... 68 35 Profil rasio DIN : DIP di zona eufotik secara temporal (a) dan
spasial (b) di Teluk Banten. ... 70 36 Profil rasio DSi dan DIN di zona eufotik Teluk Banten. ... 72 37 Profil distribusi total kelimpahan, Bacillariophyceae, dan
Chaetoceros spp. sebagai fungsi rasio DSi:DIN... 73 38 Profil nilai produktivitas primer fitoplankton di zona eufotik
secara temporal (a) dan spasial di Teluk Banten... 75 39 Profil konsentrasi klorofil-a di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 77 40 Profil kelimpahan fitoplankton di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 79 41 Profil kelimpahan Bacillariophyceae di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 82 42 Profil kelimpahan Dinophyceae di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 83 43 Profil kelimpahanChaetocerosspp. di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 86 44 Profil kelimpahanNoctilucasp. di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 88 45 Profil laju pertumbuhan fitoplankton di zona eufotik secara temporal (a)
46 Profil waktu doubling fitoplankton di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 92 47 Profil waktu generasi fitoplankton di zona eufotik secara temporal (a)
dan spasial (b) di Teluk Banten... 94 48 Konsentrasi DIN (a) dan DIP (b) di zona eufotik
Teluk Banten. ... 107 49 Laju pertumbuhan fitoplankton pada enam level intensitas cahaya
matahari di zona eufotik 1% stasiun satu (a), zona eufotik 5%
stasiun satu (b), dua (c), tiga (d), dan empat (e)... 111 50 Laju pertumbuhan fitoplankton pada enam level intensitas cahaya
matahari di zona eufotik 10% stasiun satu (a), dua (b), tiga (c),
dan empat (d). ... 114 51 Laju pertumbuhan fitoplankton pada enam level intensitas cahaya
matahari di zona eufotik 25% stasiun dua (a), tiga (b), empat (c),
dan zona eufotik 50% stasiun dua (d), tiga (e), dan empat (f). ... 117 52 Laju pertumbuhan fitoplankton pada enam level silika inorganik
terlarut di zona eufotik 50% stasiun dua (a), tiga (b), empat (c),
Halaman
1 Hasil uji anova distribusi kedalamanSecchi discsecara temporal
di perairan Teluk Banten... 145
2 Hasil uji anova distribusi kedalamanSecchi discsecara spasial di perairan Teluk Banten... 145
3 Hasil analisis korelasi Pearson’s parameter fisika dan kimia serta biologi perairan Teluk Banten. ... 146
4 Hasil uji anova koefisien peredupan secara temporal di perairan Teluk Banten... 148
5 Hasil uji anova koefisien peredupan secara spasial di perairan Teluk Banten... 148
6 Hasil uji anova rasio zona mixing dan eufotik secara temporal di perairan Teluk Banten... 149
7 Hasil uji anova rasio zona mixing dan eufotik secara spasial di perairan Teluk Banten... 149
8 Hasil uji anova salinitas (‰) di zona eufotik Teluk Banten. ... 150
9 Hasil uji anova salinitas (‰) secara temporal Teluk Banten... 150
10 Hasil uji anova salinitas (‰) secara spasial Teluk Banten. ... 150
11 Hasil uji anova suhu (°C) di zona eufotik Teluk Banten. ... 151
12 Hasil uji anova suhu (°C) secara temporal Teluk Banten. ... 151
13 Hasil uji anova suhu (°C) secara spasial Teluk Banten. ... 151
14 Hasil uji anova pH di zona eufotik Teluk Banten... 152
15 Hasil uji anova pH secara temporal Teluk Banten... 152
16 Hasil uji anova pH secara spasial Teluk Banten. ... 153
17 Hasil uji anova intensitas cahaya matahari permukaan (umol foton m-2s-1) Permukaan di perairan Teluk Banten... 153
18 Intensitas cahaya matahari permukaan (umol foton m-2s-1) di perairan Teluk Banten... 155
19 Intensitas cahaya matahari (umol foton m-2s-1) di zona eufotik perairan Teluk Banten... 156
20 Hasil uji anova intensitas cahaya matahari (umol foton m-2s-1) di zona eufotik perairan Teluk Banten... 157
22 Hasil uji anova intensitas cahaya matahari secara spasial
di perairan Teluk Banten... 157 23 Konsentrasi nitrogen inorganik terlarut (μM) di perairan
Teluk Banten. ... 158 24 Hasil uji anova nitrogen inorganik terlarut (μM)
di zona eufotik perairan Teluk Banten ... 161 25 Hasil uji anova nitrogen inorganik terlarut (μM) secara
temporal di perairan Teluk Banten. ... 161 26 Hasil uji anova nitrogen inorganik terlarut (μM) secara
spasial di perairan Teluk Banten... 161 27 Hasil uji anova amonia nitrogen (μM) di zona eufotik
perairan Teluk Banten... 162 28 Hasil uji anova amonia nitrogen (μM) secara temporal
di perairan Teluk Banten... 162 29 Hasil uji anova amonia nitrogen (μM) secara
spasial di perairan Teluk Banten... 163 30 Hasil uji anova nitrat nitrogen (μM) di zona eufotik
perairan Teluk Banten... 163 31 Hasil uji Anova distribusi nitrat nitrogen secara temporal
di perairan Teluk Banten... 163 32 Hasil uji anova nitrat nitrogen (μM) secara spasial
di perairan Teluk Banten... 164 33 Hasil uji anova nitrit nitrogen (μM) di zona eufotik
perairan Teluk Banten... 164 34 Hasil uji anova nitrit nitrogen (μM) secara temporal
di perairan Teluk Banten... 164 35 Hasil uji anova nitrit nitrogen (μM) secara spasial
di perairan Teluk Banten... 164 36 Hasil uji anova fosfat inorganik terlarut (μM) di zona eufotik
perairan Teluk Banten... 165 37 Hasil uji anova fosfat inorganik terlarut (μM) secara temporal
di perairan Teluk Banten... 165 38 Hasil uji anova fosfat inorganik terlarut (μM) secara spasial
di perairan Teluk Banten... 166 39 Hasil uji anova silika inorganik terlarut (μM) di zona eufotik
perairan Teluk Banten... 166 40 Hasil uji anova silika inorganik terlarut (μM) secara temporal
di perairan Teluk Banten... 166 41 Hasil uji anova silika inorganik terlarut (μM) secara spasial
di perairan Teluk Banten... 167 42 Rasio nutrien inorganik terlarut. ... 168 43 Hasil uji anova rasio DIN dan DIP di zona eufotik
perairan Teluk Banten... 171 44 Hasil uji anova Rasio DIN dan DIP secara temporal
di perairan Teluk Banten... 171 45 Hasil uji anova rasio DIN dan DIP secara spasial
di perairan Teluk Banten... 171 46 Hasil uji anova rasio DSi dan DIN di zona eufotik
perairan Teluk Banten... 172 47 Hasil uji anova Rasio DSi dan DIN secara temporal
di perairan Teluk Banten... 172 48 Hasil uji anova rasio DSi dan DIN secara spasial
di perairan Teluk Banten... 172 49 Nilai produktivitas primer fitoplankton di perairan
(mg C m-3jam-1) Teluk Banten. ... 173 50 Hasil uji anova produktivitas primer fitoplankton (mg C m-3jam-1)
di zona eufotik perairan Teluk Banten... 174 51 Hasil uji anova produktivitas primer fitoplankton (mg C m-3jam-1)
secara temporal di perairan Teluk Banten... 174 52 Hasil uji anova produktivitas primer fitoplankton (mg C m-3jam-1)
secara spasial di perairan Teluk Banten... 174 53 Nilai klorofil-a (mg chl-a m-3) dan karbon fitoplankton
(mg C m-3) di perairan Teluk Banten. ... 175 54 Hasil uji anova klorofil-a (mg chl-a m-3) di zona eufotik
perairan Teluk Banten... 176 55 Hasil uji anova klorofil-a (mg chl-a m-3) secara temporal
di perairan Teluk Banten... 176 56 Hasil uji anova klorofil-a (mg chl-a m-3) secara spasial
di perairan Teluk Banten... 176 57 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan April di zona
eufotik perairan Teluk Banten... 177 58 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan Mei di zona
eufotik perairan Teluk Banten... 178 59 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan Juni di zona
eufotik perairan Teluk Banten... 180 60 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan Juli di zona
61 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan Agustus
di zona eufotik perairan Teluk Banten... 183 62 Hasil analisis kelimpahan fitoplankton (sel l-1) pada bulan September
di zona eufotik perairan Teluk Banten... 184 63 Hasil uji anova kelimpahan fitoplankton (sel l-1) di zona eufotik
perairan Teluk Banten... 186 64 Hasil uji anova distribusi kelimpahan fitoplankton (sel l-1)
secara temporal di perairan Teluk Banten... 186 65 Hasil uji anova kelimpahan fitoplankton (sel l-1) secara spasial
di perairan Teluk Banten... 186 66 Hasil uji anova kelimpahan Bacillariophyceae (sel l-1) di zona eufotik
perairan Teluk Banten... 187 67 Hasil uji anova kelimpahan Bacillariophyceae (sel l-1) secara temporal
di perairan Teluk Banten... 187 68 Hasil uji anova kelimpahan Bacillariophyceae (sel l-1) secara spasial
di perairan Teluk Banten... 188 69 Hasil uji anova kelimpahan Dinophyceae (sel l-1) di zona eufotik
perairan Teluk Banten... 188 70 Hasil uji anova kelimpahan Dinophyceae (sel l-1) secara temporal
di perairan Teluk Banten... 188 71 Hasil uji anova kelimpahan Dinophyceae (sel l-1) secara spasial
di perairan Teluk Banten... 189 72 Hasil uji anova kelimpahanChaetocerosspp. (sel l-1) di zona eufotik
perairan Teluk Banten... 189 73 Hasil uji anova kelimpahanChaetocerosspp. (sel l-1) secara temporal
di perairan Teluk Banten... 189 74 Hasil uji anova distribusi kelimpahanChaetocerosspp. (sel l-1)
secara spasial di perairan Teluk Banten... 190 75 Hasil uji anova kelimpahanNoctilucaspp. (sel l-1) di zona eufotik
perairan Teluk Banten... 190 76 Hasil uji anova kelimpahanNoctilucaspp. (sel l-1) secara temporal
di perairan Teluk Banten... 190 77 Hasil uji anova kelimpahanNoctilucasp. (sel l-1) secara spasial
di perairan Teluk Banten... 191 78 Laju pertumbuhan, waktu doubling dan pembelahan sel fitoplankton
(hari-1) di perairan Teluk Banten... 192 79 Hasil uji anova laju pertumbuhan fitoplankton (hari l-1) di zona eufotik
perairan Teluk Banten... 194 80 Hasil uji anova laju pertumbuhan fitoplankton (hari l-1) secara temporal
di perairan Teluk Banten... 194 81 Hasil uji anova laju pertumbuhan fitoplankton (hari l-1) secara spasial
di perairan Teluk Banten... 195 82 Hasil uji anova waktu doubling fitoplankton (hari l-1) di zona eufotik
di perairan Teluk Banten... 195 83 Hasil uji anova waktu doubling fitoplankton (hari l-1) secara temporal
di perairan Teluk Banten... 196 84 Hasil uji anova waktu doubling fitoplankton (hari l-1) secara spasial
di perairan Teluk Banten... 196 85 Hasil uji anova pembelahan sel fitoplankton (hari l-1) di zona eufotik
di perairan Teluk Banten... 197 86 Hasil uji anova pembelahan sel fitoplankton (hari l-1) secara temporal
di perairan Teluk Banten... 197 87 Hasil uji anova pembelahan sel fitoplankton (hari l-1) secara spasial
PENDAHULUAN
Latar Belakang
Pada dasarnya, dinamika pertumbuhan fitoplankton terjadi sebagai pengaruh dari variabilitas intensitas cahaya matahari dan nutrien di perairan. Pertumbuhan fitoplankton akan berlangsung cepat bila ditunjang oleh ketersediaan intensitas cahaya matahari dan nutrien inorganik terlarut terutama dalam bentuk nitrogen (N), fosfat (P) dan silika (Si) yang cukup untuk pertumbuhannya. Namun pada kenyataannya interaksi antara kedua komponen penting ini selalu bervariasi di zona eufotik. Hal ini tentu akan menyebabkan fitoplankton memberikan respon yang bervariasi karena tidak semua fitoplankton dapat beradaptasi dengan level intensitas cahaya yang tinggi maupun rendah (Jager et al. 2008). Selain itu, diperkirakan dari 20 000 jenis fitoplankton hanya sebagian kecil yang berperan penting dalam siklus nutrien (Falkowski dan Raven 1997), seperti N (Doreet al. 2002), P (Benitez-Nelson dan Karl 2002) dan Si (Brown et al. 2003). Sebagai konsekuensi dari variasi level intensitas cahaya dan konsentrasi N, P dan Si menyebabkan pertumbuhan fitoplankton di perairan alami menjadi lambat (Ornolfsdottiret al. 2004).
untuk tumbuh pada dua kondisi yang berbeda ini sangat ditentukan oleh kemampuannya untuk beradaptasi dengan kondisi cahaya yang bervariasi serta memanfaatkan N, P dan Si yang berada di zona eufotik. Fenomena seperti ini dapat terjadi pada semua perairan, termasuk perairan Teluk Banten.
Perairan Teluk Banten merupakan salah perairan yang memiliki tipe perairan yang tercampur sempurna atau well-mixed (Nurhayati 2002). Kondisi perairan seperti ini tentu akan sangat berpengaruh pada distribusi vertikal zona eufotik dan konsentrasi N, P dan Si di kolom air. Hal ini terlihat dari hasil penelitian sebelumnya yang memperlihatkan bervariasinya zona eufotik serta distribusi konsentrasi N, P, dan Si yang homogen di perairan teluk ini (Alianto et al. 2008a; Alianto et al, 2008b; Alianto dan Damar 2009; Aliantoet al. 2009b). Selain berpengaruh pada distribusi vertikal zona eufotik serta N, P, dan Si, perairan teluk dengan tipe tercampur sempurna ini akan menyebabkan pula fitoplankton tersuspensi di kolom air bersama dengan N, P, dan Si dengan level intensitas cahaya matahari yang bervariasi (Ghosalet al. 2000). Berkaitan dengan uraian ini serta didukung oleh beberapa uraian sebelumnya di atas, maka dapat diidentifikasi yang menjadi permasalahan adalah “apakah potensi tumbuh fitoplankton serasi atau tidak dengan level intensitas cahaya matahari dan distribusi konsentrasi N, P dan Si di perairan Teluk Banten”.
Rumusan Masalah
dengan massa air laut yang dalam pergerakannya akan mengalami percampuran. Konsekuensi dari percampuran dua massa air ini tentu akan mempengaruhi level intensitas cahaya matahari dan ketersediaan DIN, DIP, dan DSi di kolom air. Kondisi ini pada akhirnya akan berpengaruh pada bervariasinya zona eufotik dan mixing yang terbentuk di perairan pantai, tengah maupun perairan luar teluk. Pembentukan kedua zona ini sangat ditentukan oleh pengaruh proses percampuran dari dua massa air ini pada setiap bagian perairan.
Pada perairan pantai yang dangkal umumnya yang dominan pengaruh dari darat yang dicirikan oleh percampuran yang sempurna (Huismann et al. 2006) dengan tingkat kekeruhan yang tinggi. Konsekuensinya menyebabkan konsentrasi DIN, DIP, dan DSi di zona ini relatif tinggi karena mendapat suplai dari aliran sungai dan run-off serta dari proses pengadukan dari dasar perairan. Sebaliknya penetrasi intensitas cahaya matahari hanya mencapai lapisan permukaan, sehingga zona eufotik yang terbentuk relatif dangkal. Konsekuensinya sudah jelas level intensitas cahaya matahari rendah karena terserap oleh partikel tersuspensi di zona ini. Oleh karena itu dapat dikatakan bahwa di zona pantai yang tercampur sempurna ini penetrasi cahaya matahari umumnya rendah dibandingkan dengan zona mixing yang meliputi seluruh kolom perairan dan umumnya yang menjadi pembatas pertumbuhan fitoplankton adalah cahaya (Hassett et al. 1997). Dengan demikian fitoplankton akan tersebar diseluruh kolom air dan untuk pertumbuhannya sangat tergantung pada kemampuan fitoplankton itu sendiri untuk memanfaatkan DIN, DIP, dan DSi dengan level intensitas cahaya matahari yang rendah (Jageret al. 2008).
Pada perairan tengah teluk yang relatif dangkal yang merupakan zona transisi antara pantai dan laut biasanya dicirikan oleh pengaruh darat yang mulai berkurang. Kondisi ini tentu akan menyebabkan penetrasi cahaya matahari cenderung lebih dalam dibandingkan dengan perairan pantai. Konsekuensinya zona eufotik yang terbentuk di bagian perairan ini dapat mencapai dasar perairan karena pengaruh kekeruhan mulai berkurang. Kondisi ini akan menyebabkan perairan memiliki distribusi konsentrasi DIN, DIP, dan DSi lebih rendah dari perairan pantai karena sumbernya hanya berasal dari dasar perairan (Huismann et al. 2006). Sedangkan sumber utama DIN, DIP, dan DSi di perairan berasal dari
daratan yang mengalir melalui aliran sungai-sungai disekitarnya dan juga run-off dari daratan. Berdasarkan karakteristik perairan seperti yang telah diuraikan ini, maka yang menjadi pembatas pertumbuhan fitoplankton adalah nutrien. Dengan demikian fitoplankton yang dapat tumbuh dengan baik adalah kelompok fitoplankton yang dapat memanfaatkan dan beradaptasi dengan konsentrasi DIN, DIP, dan DSi yang rendah dan dapat beradaptasi pula dengan level cahaya yang mulai cenderung tinggi (Goldman 2003).
Pada perairan luar teluk lebih didominasi oleh pengaruh perairan laut yang umumnya dicirikan oleh kondisi perairan yang jernih, sehingga penetrasi intensitas cahaya matahari dapat mencapai perairan yang lebih dalam. Konsekuensinya zona eufotik yang terbentuk lebih dalam atau dapat mencapai separuh dari kedalaman perairan. Pada perairan laut yang dalam umumnya terjadi stratifikasi antara lapisan permukaan yang dinamakan sebagai zona eufotik dan lapisan bawah yang dinamakan sebagai zona mixing (Diehl 2002). Konsekuensi dari stratifikasi kolom air ini adalah pada zona eufotik konsentrasi DIN, DIP, dan DSi rendah dan pada zona mixing tinggi. Suplai DIN, DIP, DSi di zona eufotik hanya bersumber dari proses remineralisasi bahan organik (Hirose dan Kamiya 2003). Oleh karena itu yang menjadi pembatas pertumbuhan fitoplankton di zona eufotik ini adalah nutrien (Hassett et al. 1997). Dengan demikian fitoplankton yang dapat tumbuh di perairan ini adalah kelompok fitoplankton yang mempunyai kemampuan untuk beradaptasi dibawah kondisi nutrien yang rendah serta mempunyai kemampuan untuk mengatur posisinya di zona eufotik (Phlips et al. 1999) dengan level intensitas cahaya matahari yang tinggi (Jageret al. 2008).
cahaya matahari serta DIN, DIP, dan DSi di zona eufotik maka sangat diperlukan penelitian tentang pertumbuhan fitoplankton di zona eufotik pada ketiga bagian perairan ini. Bila pertumbuhan fitoplankton cepat, maka dapat dikatakan bahwa intensitas cahaya matahari serta DIN, DIP, dan DSi sesuai untuk pertumbuhan fitoplankton. Sebaliknya, bila laju pertumbuhan fitoplankton lambat, maka dapat dikatakan bahwa intensitas cahaya dan nutrien kurang sesuai untuk pertumbuhan fitoplankton. Pendekatan dan kerangka teoritis yang digunakan untuk menguraikan fenomena tersebut disajikan pada Gambar 1.
Tujuan dan Manfaat
Berdasarkan uraian secara detail pada latar belakang penelitian, maka penelitian ini mempunyai tujuan sebagai berikut :
1) menghitung pertumbuhan fitoplankton yang meliputi laju pertumbuhan, waktu doubling dan waktu generasi fitoplankton di zona eufotik,
2) menganalisis respon laju pertumbuhan fitoplankton pada perubahan ketersediaan intensitas cahaya matahari, dan
3) menganalisis respon laju pertumbuhan fitoplankton pada perubahan distribusi konsentrasi nutrien inorganik terlarut di zona eufotik baik secara temporal maupun spasial.
Sedangkan manfaat yang diharapkan dari penelitian ini adalah dapat mengetahui dan memahami ekologi fitoplankton Teluk Banten terutama yang meliputi informasi tentang dinamika pertumbuhan fitoplankton pada berbagai level intensitas cahaya matahari dan nutrien inorganik terlarut terutama dalam bentuk-bentuk DIN, DIP dan DSi.
Kebaruan
dan setiap berapa lama terjadi peningkatan biomassa fitoplankton direpresentasikan oleh waktu doubling fitoplankton serta jumlah generasi fitoplankton baru yang dihasilkan direpresentasikan oleh waktu generasi fitoplankton. Informasi ini diperoleh secarain situpada berbagai kombinasi level intensitas cahaya matahari dan nutrien inorganik terlarut.
Pertumbuhan fitoplankton dengan kombinasi intensitas cahaya matahari yang tinggi dan nutrien inorganik terlarut yang rendah merupakan representasi dari bagian permukaan perairan dan luar teluk. Pertumbuhan fitoplankton dengan kombinasi intensitas cahaya matahari dan nutrien inorganik terlarut yang sedang merupakan representasi dari bagian tengah kolom perairan dan tengah teluk. Sedangkan pertumbuhan fitoplankton dengan kombinasi intensitas cahaya matahari yang rendah dan nutrien inorganik terlarut yang tinggi merupakan representasi dari bagian yang mendekati dasar perairan dan pantai atau muara sungai. Selain itu, diperoleh juga informasi tentang parameter mana di antara intensitas cahaya matahari dan dan nutrien inorganik terlarut yang paling berperan pada terjadinya dinamika pertumbuhan fitoplankton.
Hipotesis
Hipotesis yang digunakan pada penelitian ini adalah sebagai berikut : 1. Pertumbuhan fitoplankton akan berlangsung lambat pada perairan dengan
intensitas cahaya matahari yang rendah dan tinggi, sebaliknya akan berlangsung cepat pada perairan dengan intensitas cahaya matahari yang sedang.
Karakteristik Teluk Banten
Teluk Banten terletak di pesisir bagian utara Kabupaten Serang yang dibatasi oleh Tanjung Piatu disebelah barat dan Tanjung Pontang di sebelah timur. Teluk yang merupakan sebuah teluk terbuka ini memiliki panjang rentangan mulut lebih kurang 16 km dengan luas sekitar 120 km2 (Douvenet al. 2000). Perairan Teluk Banten merupakan perairan yang dangkal, umumnya kedalamannya kurang dari 20 meter (Booijet al. 2001). Pada bagian dalam teluk terdapat pulau-pulau kecil, hanya satu pulau kecil yang berpenghuni dan sebagian besar pulau-pulau lainnya tidak berpenghuni. Teluk ini menerima aliran air tawar yang mengalir melalui sungai-sungai disekitarnya. Aliran air tawar ini sebagian besar mengandung air buangan yang berasal dari air buangan rumah tangga, kegiatan pertanian, peternakan maupun industri yang berada disekitarnya. Air buangan rumah tangga maupun industri dan kegiatan lainnya sebagian besar berasal dari 16 kecamatan yang berada disekitar teluk ini dan sebagian besar atau bahkan mencapai 100% air buangan ini mengalir ke kedalam Teluk Banten (Heun 1996).
terdiri atas pasir, lumpur pasiran, pasir lumpuran, pasir lanauan, lanau lumpuran, lumpur lanauan, dan pasir kerikilan (Helfinalis 2002).
Perairan Teluk Banten yang memiliki tipe pasang surut campuran dengan unsur ganda (mixed semidiurnal) yang lebih menonjol dengan tunggang pasang surut tak lebih dari 1.1 m (Ongkosongo dan Wijonarko 2004). Tipe pasang surut dan tunggang pasang surut seperti ini sangat berperan penting dalam proses pembentukan zona mixing sebagai konsekuensi dari percampuran massa air tawar dan massa air laut yang banyak mengandung nutrien. Selanjutnya massa air tawar akan terdistribusi pada seluruh bagian Teluk Banten melalui pergerakan arus. Kecepatan arus pada bagian pantai berkisar dari 0-5 cm s-1, bagian tengah berkisar dari 5-10 cm s-1 dan mengarah ke bagian luar teluk berkisar dari 10-35 cm s-1 (Ongkosongo dan Wijonarko 2004) menyebabkan massa air tawar terdistribusi secara tidak merata pada setiap bagian perairan. Selanjutnya dinyatakan bahwa massa air ini akan terbawa oleh arus menuju ke timur pada musim barat dan berubah menuju ke barat pada musim timur (Gambar 2). Hal ini tentu akan berpengaruh pula pada distribusi penetrasi intensitas cahaya matahari dan nutrien terutama dalam bentuk-bentuk terlarut, seperti DIN, DIP, dan DSi berbeda pada setiap bagian perairan teluk. Kondisi seperti ini akan berpengaruh pada respon fitoplankton yang berbeda pula pada setiap bagian perairan.
Gambar 2 Arah arus pada musim barat (a) dan timur (b) di Teluk Banten (Ongkosongo dan Wijonarko 2004).
Konsep Pertumbuhan Fitoplankton
Pengertian Pertumbuhan
Pada prinsipnya pertumbuhan secara umum didefinisikan sebagai proses berkembangnya organisme. Bila ditinjau secara mendalam pengertian dari definisi pertumbuhan ini, maka dapat dikatakan bahwa pertumbuhan identik dengan gerak, sehingga secara ekologi pertumbuhan lebih dikenal sebagai kinetik pertumbuhan (Riley dan Chester 1971). Selanjutnya Riley dan Chester (1971) mendefinisikan kinetik pertumbuhan sebagai perubahan dalam jumlah sel hidup per unit waktu atau volume. Berkaitan dengan ini, Causton dan Venus (1981) menyatakan bahwa perubahan jumlah sel ini terjadi melalui dua mekanisme, yaitu (1) mekanisme yang berkaitan dengan perubahan secara kualitatif seperti terjadinya pembentukan sel, jaringan dan organ-organ tubuh, dan (2) mekanisme yang berkaitan dengan perubahan secara kuantitatif seperti perkembangan organisme. Selanjutnya dinyatakan bahwa sebagai hasil akhir dari kedua mekanisme ini khususnya pada organisme uniselullar seperti fitoplankton adalah terjadinya pembelahan sel dan peningkatan jumlah sel organisme atau fitoplankton.
penting terutama untuk dapat membedakan antara laju pertumbuhan, waktu doubling, dan waktu generasi fitoplankton.
Pada beberapa uraian di atas dapat dikatakan secara umum bahwa pertumbuhan merupakan proses-proses fisiologi yang terjadi di dalam tubuh fitoplankton itu sendiri (lihat uraian siklus reproduksi dan fase pertumbuhan fitoplankton). Hal ini berkaitan dengan pernyataan Ornolfsdottir et al. (2004) bahwa pertumbuhan fitoplankton merupakan proses-proses fisiologi, dimana akumulasi biomassa merupakan fungsi dari laju pertumbuhan. Sedangkan laju pertumbuhan itu sendiri didefinisikan sebagai jumlah karbon yang diserap melalui fotosintesis dalam unit waktu atau per unit volume (Riley dan Chester 1971), atau terjadinya peningkatan biomassa per unit waktu (Miller 2004; Woodet al. 2005). Sedangkan banyaknya atau jumlah fitoplankton yang terbentuk atau dihasilkan selama proses pertumbuhan disebut dengan waktu generasi (Riley dan Chester 1971). Waktu yang diperlukan oleh populasi untuk meningkat sampai 100% (Parsons et al. 1984a; Lalli dan Parsons 1995) atau waktu yang diperlukan oleh populasi untuk mencapai ukuran meningkatkan jumlahnya (Lalli dan Parsons 1995) disebut dengan waktu doubling.
Siklus Hidup Fitoplankton
Mekanisme reproduksi fitoplankton sangat bervariasi, ada yang vegetatif, aseksual atau seksual dan tahap resting (Riley dan Chester 1971; Garisson 1984; Tomas 1997; dan Miller 2004). Beberapa fitoplankton dapat menggunakan satu atau lebih mekanisme ini tergantung pada kondisi sekitarnya. Rilley dan Chester (1971) menyatakan bahwa proses vegetatif tidak langsung meliputi pembelahan protoplasma, melainkan terjadi pembelahan sel. Garisson (1984) menyatakan selama pembelahan sel akan terbentuk setengah theca baru dari theca yang sama yang terbentuk pada bagian dalam theca sel induk, dan ukuran sel akan berkurang setelah terjadinya pembelahan sel berulangkali (Gambar 3). Untuk mengembalikan ukuran sel seperti semula dilakukan dengan pembentukan frustula (Tomas 1997). Hal ini diawali oleh perbesaran nukleus dengan duplikasi
Gambar 3 Berkurangnya ukuran sel setelah terjadinya pembelahan sel pada diatom (Hendey 1964inGarrison 1984).
DNA yang direpresentasikan oleh badan golgi (Gambar 4a) dan nukleus ini terbagi oleh adanya plasmalemma (Gambar 4b). Valve berkembang karena terjadinya pembentukan silikalemma yang lengkap (Gambar 4c) dan deposit baru (Gambar 4d) disekitar silika deposit visikles dan sel-sel dipisahkan (Gambar 4e).
Perbaikan ukuran sel seperti yang telah diuraikan di atas dilakukan melalui reproduksi seksual dengan pembentukan zygot yang diikuti oleh pembentukan sel berukuran besar yang dinamakan dengan auxospora (Garisson 1984). Selanjutnya dinyatakan bahwa auxospora ini akan berkembang ke dalam sel vegetatif sampai
Gambar 4 Pembelahan sel dan pembentukan frustula (Dawson 1973).
French 1954inGarisson 1984). Resting spore umumnya terbentuk sebagai respon pada kondisi lingkungan yang tidak menguntungkan (Tomas 1997). Selanjutnya dinyatakan bahwa terdapat tiga bentuk resting spora, yaitu exogeneous resting spora (resting spora dewasa yang tidak berkaitan dengan theca sel induk), semiendogenous resting spora (spora hypovalve yang tertutup dengan satu theca sel induk), endogenous resting spore (semua spora tertutup dengan frustula sel induk).
Fase Pertumbuhan Fitoplankton
Pertumbuhan fitoplankton pada prinsipnya melalui lima fase, yaitu fase diam (lag phase), fase eksponensial (eksponential phase), fase lambat (retardation phase), fase stabil (stationary phase), dan fase kematian (death phase) (Fogg 1965; Riley dan Chester 1971; Millero 2006) (Gambar 5). Pertumbuhan fitoplankton pada tahap fase diam, belum terjadi peningkatan jumlah sel (Fogg 1965). Hal yang sama dinyatakan pula oleh Wood et al. (2005) bahwa belum terdapat pertumbuhan fitoplankton pada fase diam. Pada fase ini baru terjadi akumulasi energi di dalam sel (Pollingher 1988) atau reaksi enzimatis di dalam sel sampai terbentuknya suatu medium baru (lihat uraian peran intensitas cahaya dan nutrien inorganik terlarut pada pertumbuhan fitoplankton), yaitu berupa penggabungan karbon dioksida kedalam produk karbon organik selama reaksi gelap berlangsung (Behrenfeld et al. 2002; Miller 2004). Selanjutnya dinyatakan pula bahwa proses-proses ini merupakan kelanjutan dari produk reaksi terang yang menghasilkan energi cahaya untuk fotosistem I dan II (PS I, PS II), transport elektron, serta generasi ATP (adenosine triphosphate) dan NADPH (nicotinamide adenine dinucloitide phosphate).
Pertumbuhan fitoplankton pada tahap fase eksponensial, dicirikan oleh cepatnya peningkatan jumlah sel beberapa kali secara eksponensial (Fogg 1965; Riley dan Chester 1971). Memasuki fase lambat, pertumbuhan fitoplankton mulai berkurang. Riley dan Chester (1971) menyatakan bahwa berkurangnya pertumbuhan fitoplankton disebabkan oleh (1) berkurangnya konsentrasi nutrien
Gambar 5 Kurva pertumbuhan fitoplankton (Fogg 1965; Riley dan Chester 1971; Millero 2006).
merupakan penyebab berakhirnya fase lambat dan menjadi faktor pembatas, sehingga populasi sel maksimum pada akhir fase eksponensial yang selanjutnya akan berkurang dan proposinya akan sebanding dengan pada saat konsentrasi nutrien awal. (2) inhibition yang disebabkan oleh toksik atau terhambatnya pertumbuhan karena toksik yang diproduksi oleh sel mereka sendiri atau organisme lain, dan (3) berkurangnya fotosintesis yang disebabkan oleh shading. Fase lambat mulai masuk ke fase stabil dicirikan oleh ketersediaan nutrien mulai berkurang karena telah digunakan untuk pertumbuhan selama fase eksponensial. Sehingga pada tahap fase eksponensial tidak terjadi lagi peningkatan jumlah sel, walaupun dengan jangka waktu yang relatif singkat (Pollingher 1988).
dari yang hidup dalam bentuk cyst atau resting (Garrison 1984). Selanjutnya dinyatakan bahwa fitoplankton dalam bentuk resting dapat hidup pada periode dengan kondisi perairan yang kurang baik, seperti pada temperatur yang ekstrim dan berkurangnya nutrien. Ketika kondisi perairan mulai mendukung pertumbuhan, sel-sel resting akan terbawa ke dalam atau tertahan oleh mixing yang selanjutnya akan berkembang dan sel-sel akan mulai aktif tumbuh. Oleh karena itu bentuk-bentuk resting menjadi sesuatu yang penting bagi keberlangsungan hidup fitoplankton di laut.
Pembatas Pertumbuhan Fitoplankton
Intensitas Cahaya Matahari
Peran Intensitas Cahaya Matahari Pada Pertumbuhan Fitoplankton
Intensitas cahaya berperan sebagai sumber energi untuk mentransformasi nutrien menjadi biomassa alga baru (lihat uraian peran nutrien inorganik terlarut pada pertumbuhan fitoplankton) melalui proses fotosintesis (Cloern 1999). Pembentukan biomassa alga baru melalui proses fotosintesis terjadi melalui proses yang multitahap dan tergabung pada dua reaksi (Valiela 1984; Behrenfeld et al. 2002; Geider dan MacIntyre 2002), yaitu reaksi cahaya (tergantung pada cahaya) dan reaksi gelap (tidak tergantung pada cahaya). Selanjutnya dinyatakan oleh Geider dan MacIntyre (2002) bahwa pada reaksi cahaya terjadi absorpsi foton yang dilakukan oleh antena sebagai penangkap cahaya, perpindahan energi ke pusat reaksi fotosistem I dan II (PSI dan PSII), transpor elektron dari H2O (air) ke NADP (nicotinamide adenine dinucloitide phosphate), generasi ATP (adenosine triphosphate) dan NADPH (nicotinamide adenine dinucloitide phosphate). Pada kondisi normal, terjadinya proses-proses tersebut pada reaksi cahaya berberlangsung menurut reaksi berikut (Geider dan MacIntyre 2002)
8 foton + 2 H2O + 2 NADP++ ADP + 3 Pi O
2+ 2 H-+ 2 NADPH + 3 ATP
dan NADPH akan digunakan pada reaksi gelap untuk mereduksi CO2 (karbon dioksida) menjadi bahan organik melalui reaksi berikut (Valiela 1984)
12 NADH2 + 18 ATP + 6 CO2 C6H12O6+ 6 H2O + 18 ADP + 18 P + 12 NAD
Selanjutnya dinyatakan bahwa bahan organik yang dihasilkan pada reaksi ini merupakan produksi primer yang sebenarnya atau biomassa alga baru. Pembentukan biomassa alga baru melalui dua reaksi ini (reaksi terang dan gelap) sangat tergantung pada intensitas cahaya. Hal ini berkaitan dengan pernyataan Behrenfeldet al. 2002 bahwa pada intensitas cahaya rendah, fotosintesis dibatasi oleh kemampuan cahaya yang dihasilkan pada PSII. Selanjutnya dinyatakan bahwa pada intensitas cahaya saturasi fotosintesis dibatasi oleh kemampuan pengambilan karbon pada reaksi gelap. Pada titik ini fitoplankton tidak dapat menggunakan cahaya dan laju fotosintesis menjadi asimtot dan penggunaan cahaya sangat tergantung pada kelompok fitoplankton (Valiela 1984).
Pengaruh Intensitas Cahaya Matahari Pada Pertumbuhan Fitoplankton
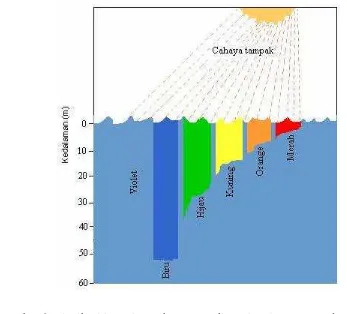
mencapai 10 meter dan berkurang di perairan keruh sekitar 1 atau 2 meter (Raymont 1963).
Gambar 6 Distribusi intensitas cahaya PAR di perairan laut (Castro dan Huber 2005).
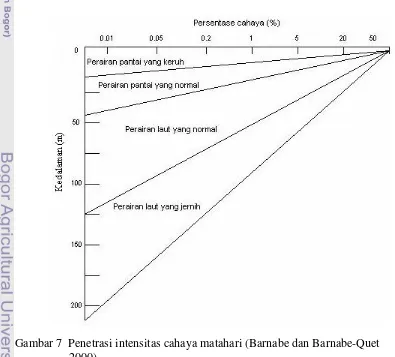
Distribusi intensitas cahaya akan semakin berkurang dalam perjalanannya menembus kolom air karena terserap oleh bahan-bahan tersuspensi (Gambar 7) seperti fitoplankton dan bahan-bahan terlarut serta molekul air itu sendiri (Wyatt dan Jackson 1989). Penyerapan cahaya oleh bahan-bahan tersuspensi ini di kolom air bervariasi tergantung dari zona mixing yang terbentuk (Kemp dan Boynton 1984). Pada perairan muara sungai, zona mixing umumnya relatif dalam sampai ke dasar perairan dengan tingkat kekeruhan yang tinggi (Cervetto et al. 2002). Pada perairan ini, berkurangnya intensitas cahaya di kolom air karena diserap oleh molekul air sebesar 10%, fitoplankton sebesar 40%, bahan-bahan terlarut sebesar 30%, dan bahan-bahan selain fitoplankton tergantung pada karakter massa air berkisar dari 2.9-43.8% (Kishino 1994). Sedangkan pada perairan laut, zona mixing yang terbentuk relatif rendah karena adanya stratifikasi kolom air dengan perairan yang jernih. Pada perairan ini, berkurangya intensitas
cahaya karena diserap oleh molekul air lebih dari 90%, fitoplankton sebesar 1.4%, dan bahan-bahan terlarut sebesar 3.1% (Kishino 1994).
Perbedaan penyerapan cahaya seperti yang telah diuraikan di atas, tentu akan menyebabkan pula perbedaan koefisien atenuasi di setiap bagian perairan. Koefisien atenuasi di perairan dengan tingkat kekeruhan yang tinggi biasanya berkisar dari 1.3 meter-1 (Kishino 1994) sampai 5.56 meter-1 (Gocke dan Lenz 2004). Sedangkan koefisien atenuasi di perairan yang jernih biasanya berkisar dari 0.04 sampai 0.05 meter-1 (Collier et al. 1973). Koefisien atenuasi yang besar mununjukkan intensitas cahaya yang menembus kolom air kecil (Huisman et al. 1999). Selanjutnya dinyatakan nilai koefisien atenuasi yang kecil menunjukkan semakin besar intensitas cahaya yang menembus kolom air. Bervariasinya koefisien atenuasi ini akan berpengaruh pula pada bervariasinya zona mixing dan eufotik. Zona mixing merupakan suatu lapisan homogen yang dicirikan oleh perbedaan suhu sampai 1°C dari suhu permukaan (Laevastu dan
Hayes 1981) dan salinitas lebih kecil dari 2‰ (Livingston 2001). Sedangkan zona eufotik merupakan suatu zona yang masih memperoleh cahaya 1% dari intensitas cahaya permukaan (Huisman 1999).
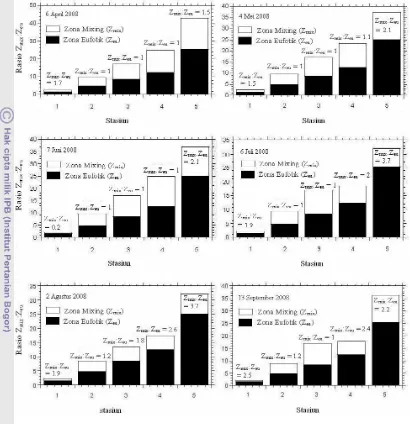
Perbedaan nilai koefisien atenuasi seperti yang telah diuraikan di atas akan berpengaruh pada bervariasinya zona mixing (Zmix) dan eufotik (Zeu) yang pada akhirnya akan berpengaruh pada laju pertumbuhan fitoplankton (Alpine dan Cloern 1988; Huisman et al. 1999; Damar 2003). Selanjutnya dinyatakan bahwa bila rasio zona mixing dan eufotik lebih kecil dari satu (Gambar 8A), hal ini menunjukkan sebagai indikator bahwa laju pertumbuhan fitoplankton akan tinggi. Laju pertumbuhan fitoplankton yang tinggi ini disebabkan karena fitoplankton berada pada kolom air dengan intensitas cahaya yang tinggi (Damar 2003) dan biasanya diperkirakan sebesar 115.92 µmol foton m-2s-1 (Alpine dan Cloern 1988). Nilai ini tidak berbeda jauh dengan nilai yang dinyatakan oleh Koch et al. (2004) bahwa laju pertumbuhan fitoplankton akan meningkat pada intensitas cahaya yang berkisar dari 81.08-104.25 µmol foton m-2 s-1. Laju pertumbuhan fitoplankton pada kondisi cahaya ini biasanya berkisar dari 0.3-1.5 hari-1 dengan waktu doubling 0.5-2.5 hari (Kochet al. 2004).
Gambar 8 Rasio ketersediaan cahaya di zona eufotik (Damar 2003).
Laju pertumbuhan fitoplankton yang tinggi seperti yang telah disebutkan di atas biasanya terjadi di perairan dangkal (Gabric dan Parslow 1989) seperti muara sungai (Koch et al. 2004) dan perairan yang zona eufotiknya mixing (Gocke dan Lenz 2004). Bila rasio zona mixing dan eufotik sama dengan satu (Gambar 8B), hal ini menunjukkan sebagai indikator pertumbuhan fitoplankton berlangsung sedang (Damar 2003). Hal ini disebabkan karena fitoplankton tertahan pada lapisan yang mendapat cahaya (Damar 2003) dengan intensitas cahaya yang hanya cukup untuk pertumbuhannya (Mallin dan Paerl 1992). Tertahannya fitoplankton dilapisan ini disebabkan karena zona mixing hanya terdapat pada sekitar bagian bawah zona eufotik dengan intensitas cahaya yang hanya cukup untuk tetap berlangsungnya pertumbuhan fitoplankton (Mallin dan Paerl 1992). Intensitas cahaya yang cukup untuk tetap tumbuhnya fitoplankton dengan laju pertumbuhan yang sedang adalah sebesar 46.33 µmol foton m-1 s-1 (Koch et al. 2004). Laju pertumbuhan fitoplankton pada kondisi cahaya yang cukup biasanya berkisar dari 0.5-1 hari-1(Banse 1982).
Nutrien Inorganik Terlarut
Peran Nutrien Inorganik Terlarut Pada Pertumbuhan Fitoplankton
Nutrien inorganik terlarut yang dibutuhkan untuk pertumbuhan fitoplankton terutama terdapat dalam bentuk DIN seperti nitrat-nitrogen (NO3--N), nitrit-nitrogen (NO2--N), dan ammonia-nitrogen (NH3-N); DIP dalam bentuk ortofosfat (PO43--P), dan DSi dalam bentuk asam ortosilikat (Si(OH)2-Si). Furnas (1992) mendeskripsikan peran dari masing-masing unsur ini adalah sebagai berikut, (1) nitrogen berperan dalam pembentukan asam amino, asam nukleit, dan kofaktor metabolik; (2) fosfat berperan dalam pembentukan asam nukleit, fosfolipit, serta energi untuk metabolisme; dan (3) silikat berperan dalam pembentukan dinding sel diatom. Selain itu, pada diatom silikat berperan juga sebagai penterjemah dalam sintesis jaringan DNA (Darley 1982; Miller 2004). Pengambilan unsur-unsur ini terutama dilakukan melalui dua mekanisme, yaitu (1) transpor aktif yang meliputi transpor aktif primer yang mengambil atau menerima masukan langsung nutrien dan transpor aktif sekunder yang mengarahkan nutrien ke sistem transpor aktif primer, dan (2) difusi (Wheller 1983).
Parsonset al. (1984a) menyatakan bahwa pengambilan DIN, DIP, dan DSi seperti yang telah diuraikan di atas dilakukan melalui dua tahap, yaitu pengambilan dari luar sel dan penggunaannya oleh sel fitoplankton. Bila ketersediaan ketiga nutrien ini cukup untuk pertumbuhan fitoplankton (lihat uraian distribusi dan rasio inorganik terlarut), maka sebagian besar akan digunakan untuk sintesis protoplasma baru yang dijelaskan oleh formula
160CO2 + 122H2O + 16NO3- + PO
43- cahaya (CH2O)106(NH3)16H3PO4 + 138O2 karbon dioksida air nitrat ortofosfat protoplasma baru oksigen
dengan produk akhir yang utama adalah protein (Fogg 1975). Komponen protein yang penting terdiri atas ribonucleic acid (RNA) dan deoxyribonucleic acid (DNA) (Wheller 1983). Selanjutnya dinyatakan bahwa setelah mengalami proses ini (lihat uraian fase pertumbuhan fitoplankton), akan terjadi atau disertai pembelahan sel fitoplankton. Namun pada kenyataannya di perairan proses-proses
ini selalu berlangsung pada kondisi yang tidak optimal karena ketersediaan cahaya (lihat uraian intensitas cahaya) dan DIN, DIP dan DSi selalu bervariasi, baik spasial maupun temporal.
Pengaruh Nutrien Inorganik Terlarut Pada Pertumbuhan Fitoplankton
Ketersediaan DIN dan DIP yang cukup untuk pertumbuhan fitoplankton seperti yang telah disebutkan pada sub bagian peran nutrien inorganik terlarut dicirikan oleh pengambilan nutrien ini yang mencapai nilai minimal untuk pertumbuhan fitoplankton. Fitoplankton untuk dapat tumbuh, minimal memerlukan nutrien terutama N dan P dengan rasio 16:1 (Riley dan Chester 1971). Rasio ini merupakan rasio N dan P dalam sel fitoplankton yang umumnya dikenal dengan rasio Redfield dan ini merupakan rata-rata rasio optimal bagi pertumbuhan seluruh komunitas fitoplankton (Lagus et al. 2004). Selanjutnya dinyatakan bahwa Rasio Redfield digunakan sebagai indikator pembatas pertumbuhan fitoplankton yang ditentukan dari selisih konsentrasi N dan P di perairan. Setiap jenis fitoplankton mempunyai tingkat pemanfaatan N dan P yang berbeda dan dari 20 000 jenis fitoplankton hanya sebagian kecil yang berperan dalam pemanfaatan N dan P (Falkowski dan Raven 1997). Hal ini tentu akan berpengaruh pada bervariasinya rasio N dan P dalam sel fitoplankton (Tomaskyet al. 1999) yang mana rasionya bisa lebih besar dari rasio Redfield atau sebaliknya.
Prins 2002). Sebaliknya, bila rasio N:Si berkurang sampai 0.09 menunjukkan terjadinya penurunan pemanfaatan Si oleh fitoplankton (Harrisonet al. 2004). Bila kondisi ini berlanjut, fitoplankton dapat memanfaatkan N dan P untuk pertumbuhannya (Hutchins dan Bruland 1998).
Laju pertumbuhan fitoplankton sangat tergantung pada kandungan N, P dan Si dalam sel yang sebagian besar sangat ditentukan oleh konsentrasi unsur ini di perairan (Cuvin-Aralar 2004) dan meningkat secara hiperbolik dengan ketersediaan nutrien (Brown et al. 2003). Konsentrasi tertinggi N, P dan Si terutama dalam bentuk inorganik terlarut biasanya lebih besar dari 30 µM untuk DIN, 4 µM untuk DIP dan 100 µM untuk DSi (Hager 1994 in Cloern 1999). Konsentrasi unsur-unsur ini di perairan teluk homogen di kolom air karena pengaruh mixing pasang surut (Cheriton et al. 2009). Hal inilah yang menjadi salah satu faktor penyebab laju pertumbuhan fitoplankton lebih tinggi (Gabric dan Parslow 1989) dibandingkan dengan bagian perairan lainnya (Gambar 8C). Laju pertumbuhan fitoplankton yang tinggi umumnya berkisar dari 0.26-2.75 hari-1 (MacIntyreet al. 2002). Pada kondisi konsentrasi nutrien tinggi laju pertumbuhan fitoplankton akan meningkat hiperbolik dan menurun bila telah berada pada kondisi saturasi (Valiela 1984).
Sedangkan mengarah ke bagian tengah teluk laju pertumbuhan fitoplankton mulai berkurang atau sedang. Hal ini dicirikan oleh proporsi N dan P yang terdapat di perairan sama dengan rasio Redfield (Riley dan Chester 1971) atau ketersediaan N dan P konsentrasinya hanya maksimum pada level rasio optimal untuk pertumbuhan fitoplankton (Laguset al. 2004). Pada kondisi seperti ini, laju pertumbuhan fitoplankton berkisar dari 0.5-1 hari-1 (Banse 1982). Ketersediaan DIN, DIP dan DSi di bagian teluk ini homogen pada kolom air, namun konsentrasinya mulai berkurang (Gambar 8B). Hal ini disebabkan karena letaknya yang mulai jauh dari daratan karena sumber utama nutrien ini berasal dari daratan yang masuk melalui sungai dan run-off dan terbawa ke bagian tengah teluk oleh gerakan mixing surut. Nutrien ini, sebelum mencapai bagian tengah teluk telah berkurang konsentrasinya karena telah dimanfaatkan oleh fitoplankton di perairan pantai untuk pertumbuhannya. Penyebab lainnya adalah nutrien ini
ditransformasi dengan cepat menjadi partikulat melalui sorption, flokulasi di bagian perairan yang salinitasnya rendah (Kemp dan Boynton 1984).
METODE PENELITIAN
Lokasi dan Waktu Penelitian
Penelitian dilaksanakan di perairan Teluk Banten, Propinsi Banten. Secara geografis lokasi penelitian berada pada posisi koordinat 10606'-10607’ Bujur Timur dan 5506’-5508’ Lintang Selatan (Gambar 9). Penelitian ini dilaksanakan dalam kurung waktu 6 bulan (April 2008 sampai September 2008) yang mewakili dua musim, yaitu musim peralihan I (hujan ke kemarau) dan kemarau. Penelitian berlangsung dalam enam kali sampling dengan interval waktu setiap sampling selama 30 hari yang meliputi tiga kali sampling pada musim peralihan I dan berikutnya tiga kali sampling pada musim kemarau.
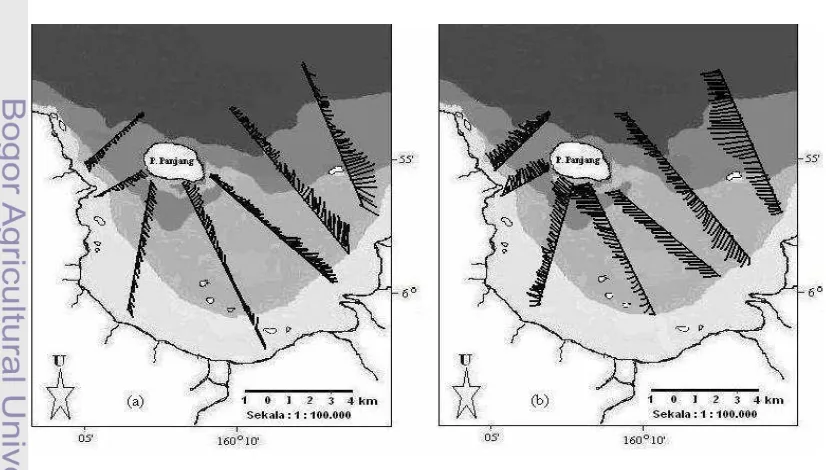
Gambar 9 Lokasi penelitian di perairan Teluk Banten.
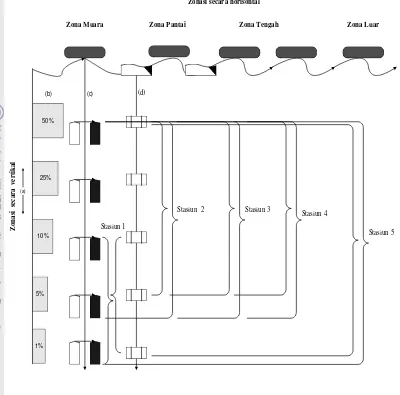
Menentukan Stasiun dan Titik Pengambilan Contoh
mengarah luar teluk, dan stasiun lima berlokasi di perairan luar teluk (Tabel 1 dan Gambar 10). Dasar pertimbangan penentuan lokasi penelitian secara horisontal dengan lima stasiun adalah distribusi vertikal intensitas cahaya matahari akan semakin dalam ke arah laut. Sebaliknya distribusi DIN, DIP, DSi, dan fitoplankton itu sendiri akan semakin berkurang ke arah laut. Dengan demikian, atas pertimbangan ini diperkirakan pertumbuhan fitoplankton akan bervariasi mengikuti level intensitas cahaya matahari dan keberadaan ketiga bentuk nutrien inorganik terlarut tersebut.
Tabel 1 Posisi stasiun untuk pengukuran dan pengambilan sampel
Stasiun Lokasi Posisi
Bujur Timur Lintang Selatan 1 Muara sungai 106°09'57.1" 06°01'24.1"
2 Pantai 106°10'01.4" 06°00'47.0"
3 Tengah mengarah ke pantai 106°09'48.3" 05°59'00.9" 4 Tengah mengarah ke luar 106°10'56.6" 05°56'36.1" 5 Luar teluk 106°10'30.0" 05°53'26.2"
50%
25%
10%
5%
Zona Pantai Zona Tengah Zona Luar
(a)
Stasiun 2 Stasiun 3 Stasiun 4
Stasiun 5
Gambar 10 Desain zonasi secara horisontal dan vertikal lokasi penelitian dilihat dari samping, (a) kedalaman yang akan ditentukan berdasarkan hasil pengukuran intensitas cahaya matahari, (b) kedalaman inkubasi, (c) pengukuran parameter kualitas air laut, dan (d) pengambilan contoh air.
Pengukuran Parameter Kualitas Perairan
Fisika Perairan
Mengukur Zona Mixing
Zona mixing ditentukan dari hasil bacaan Conductivity Thermometer Depth (CTD). Zona ini ditentukan berdasarkan ketebalan lapisan turbulen atau terbentuknya suatu kedalaman yang mempunyai ciri yang hampir sama, dimana
kedalaman ini merupakan suatu kedalaman yang dicirikan oleh perbedaan suhu yang mempunyai kisaran sampai 1 °C dari suhu permukaan (Laevastu dan Hayes 1981) (lihat uraian pengukuran suhu) dan kisaran salinitas yang tidak lebih dari 2 ‰ (Livingston 2003) (Lihat uraian pengukuran salinitas).
Mengukur Zona Eufotik
Zona eufotik ditentukan berdasarkan persentase penetrasi intensitas cahaya matahari sampai 1% dari cahaya permukaan di kolom air. Intensitas cahaya matahari permukaan diukur dengan alat Automatic Weather Station (AWS) tipe JY 106 dari Badan Meteorologi dan Geofisika Serang (Tabel 2). Besarnya intensitas cahaya matahari yang dapat menembus kolom air dihitung dengan menggunakan persamaan eksponensial atau hukum Beer-Lambert (Walsby 2001) sebagai berikut :
kz o z I e
I (3.1)
keterangan :
Iz = intesitas cahaya pada kedalamanz Io = intensitas cahaya permukaan
k = adalah koefisien peredupan (diketahui dari persamaan 3.2), dan Z = adalah kedalaman
Koefisien peredupan diketahui dari hasil pengukuran kecerahan perairan. Kecerahan perairan (dalam satuan meter) diukur dengan menggunakan keping Secchi disc(Sd) yang berdiameter 30 cm. Setelah itu koefisien peredupan dihitung dengan menggunakan persamaan sebagai berikut (Tillmanet al. 2000) :
Mengukur Suhu Perairan
Pengukuran suhu perairan dilakukan bersamaan dengan parameter kimia perairan, seperti salinitas dan pH yang dilakukan dengan menggunakan Conductivity Thermometer Depth (CTD) (Tabel 2). Pengukuran suhu, salinitas, dan pH dilakukan dengan cara menurunkan CTD dari permukaan perairan hingga ke dasar perairan. Cara seperti ini dilakukan pada masing-masing zona (zona muara sungai, pantai, tengah, dan luar teluk).
Tabel 2 Parameter kualitas air laut yang akan diukur serta metode dan alat ukur yang digunakan
Parameter Satuan Metode dan Alat Analisis
Tempat Analisis
Oksigen terlarut mg l-1 Oksigen, titrasi Winkler In situ NH3-N μM Brusin, spektrofotometer Eks situ*
NO3-N μM Sulfanilik, spektrofotometer Eks situ*
NO2-N μM Penat, spektrofotometer Eks situ*
PO4-P μM Asam molibdat, spektrofotometer Eks situ*
Si(OH)4-Si μM Molibdosilika, spektrofotometer Eks situ*
Fitoplankton Sel l-1 Pencacahan, mikroskop Eks situ*
Klorofil-a mg m-3 Spektrofotometer Eks situ* Intensitas cahaya
matahari μmol foton m-2s-1 Automatic weather station In situ
Kecerahan Meter Visual,Secchi disc In situ
Suhu °C CTD In situ
Salinitas ‰ CTD In situ
pH pH Orion In situ
* Laboratorium Produktivitas dan Lingkungan Perairan, Fakultas Perikanan dan Kelautan, Institut Pertanian Bogor
Kimia Perairan
Mengukur Konsentrasi Nutrien Inorganik Terlarut